Products Derived from Buchenavia tetraphylla Leaves Have In Vitro Antioxidant Activity and Protect Tenebrio molitor Larvae against Escherichia coli-Induced Injury

 , , ,
, , , 
Abstract
:1. Introduction
2. Results
2.1. Comparison of the Antioxidant Activity of Extracts Obtained from B. tetraphylla Leaves
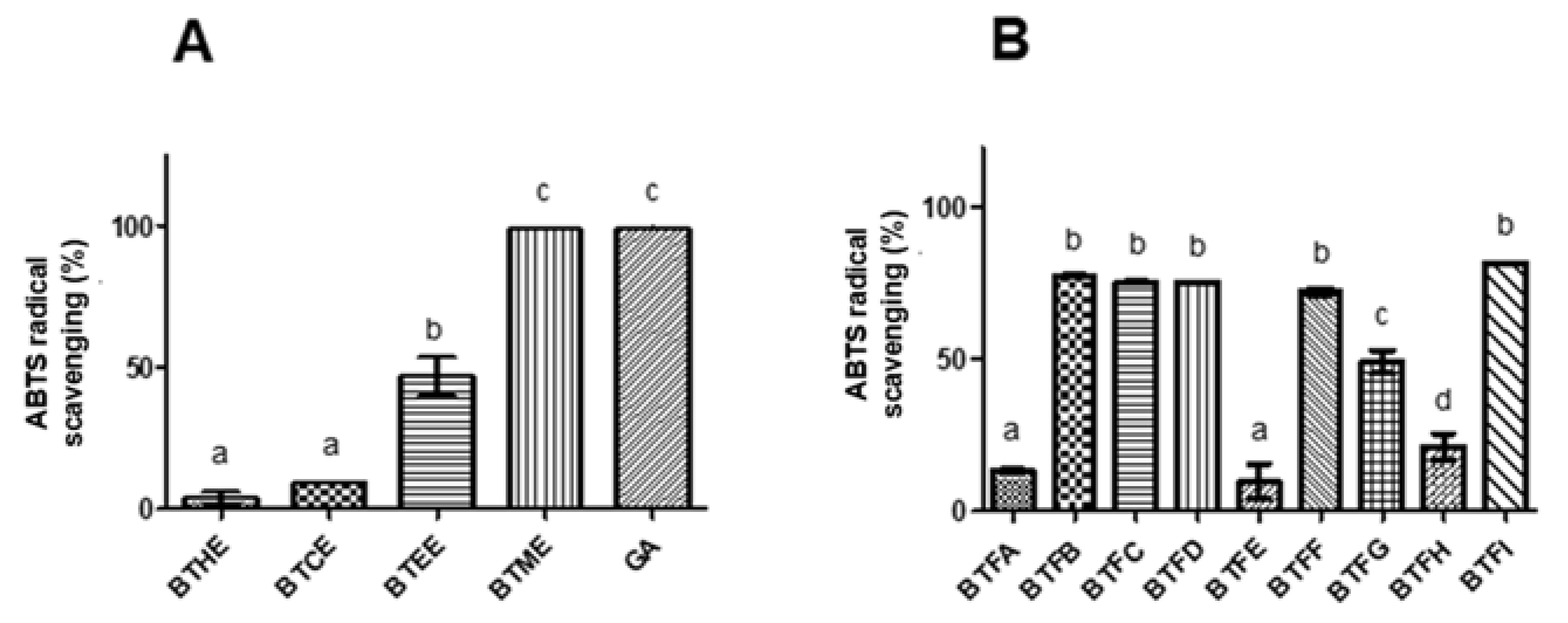
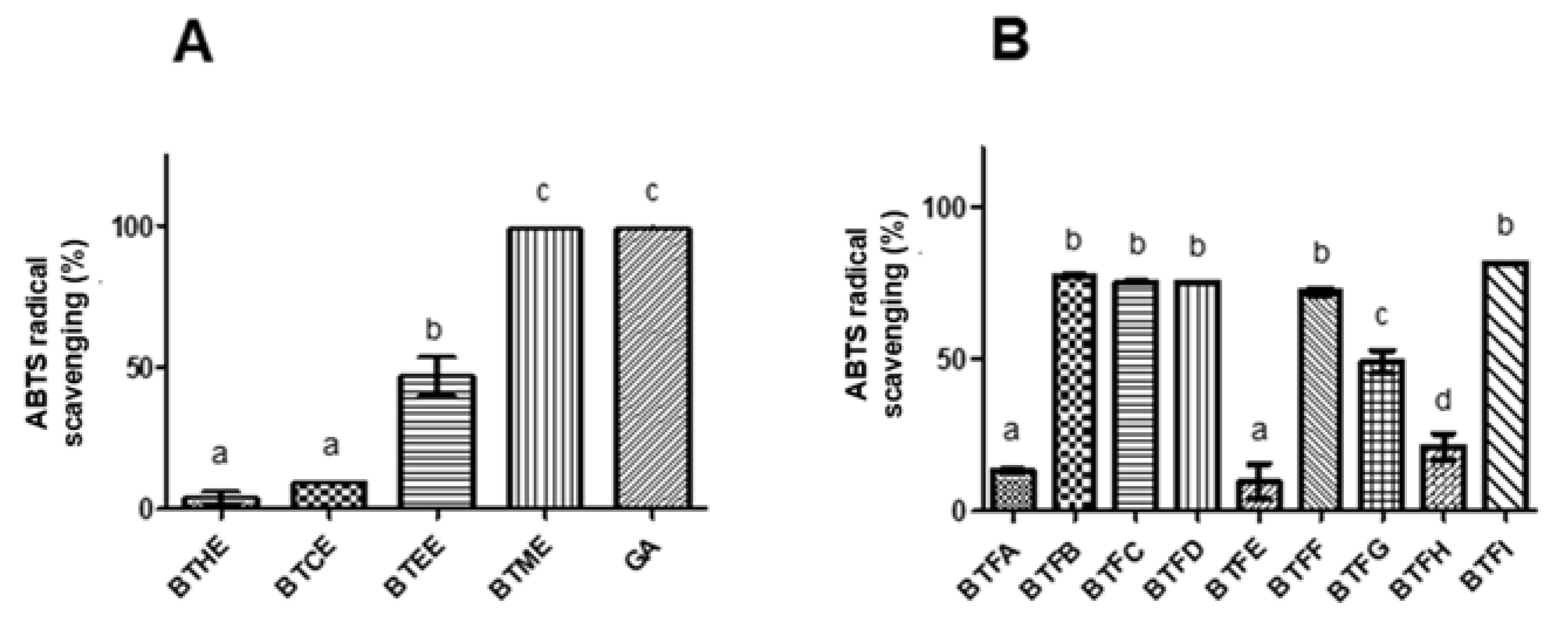
2.2. Comparison of Phenolic and Flavonoid Content and Antioxidant Activity of Fractions Obtained from Methanolic Extract of B. tetraphylla Leaves
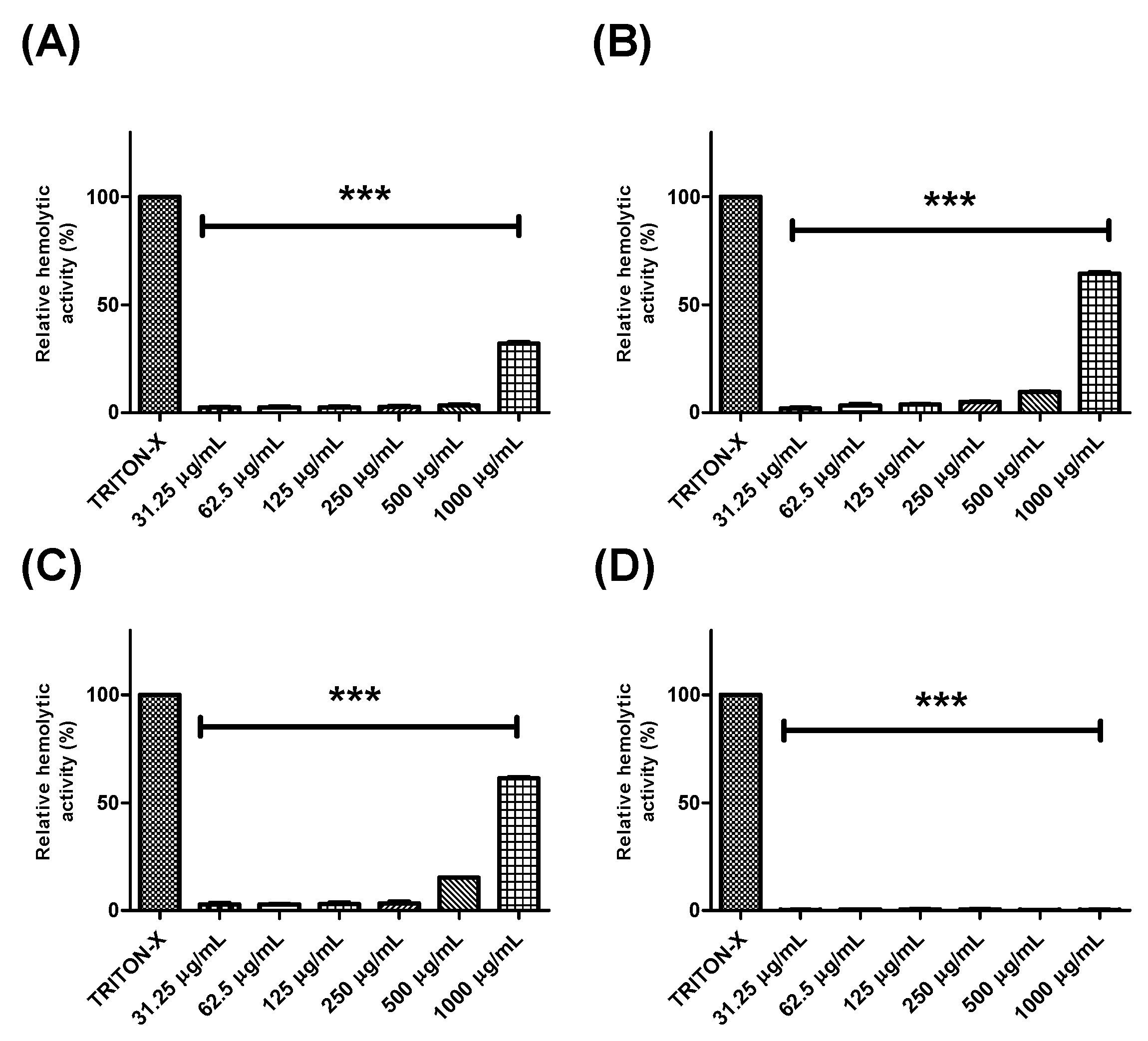
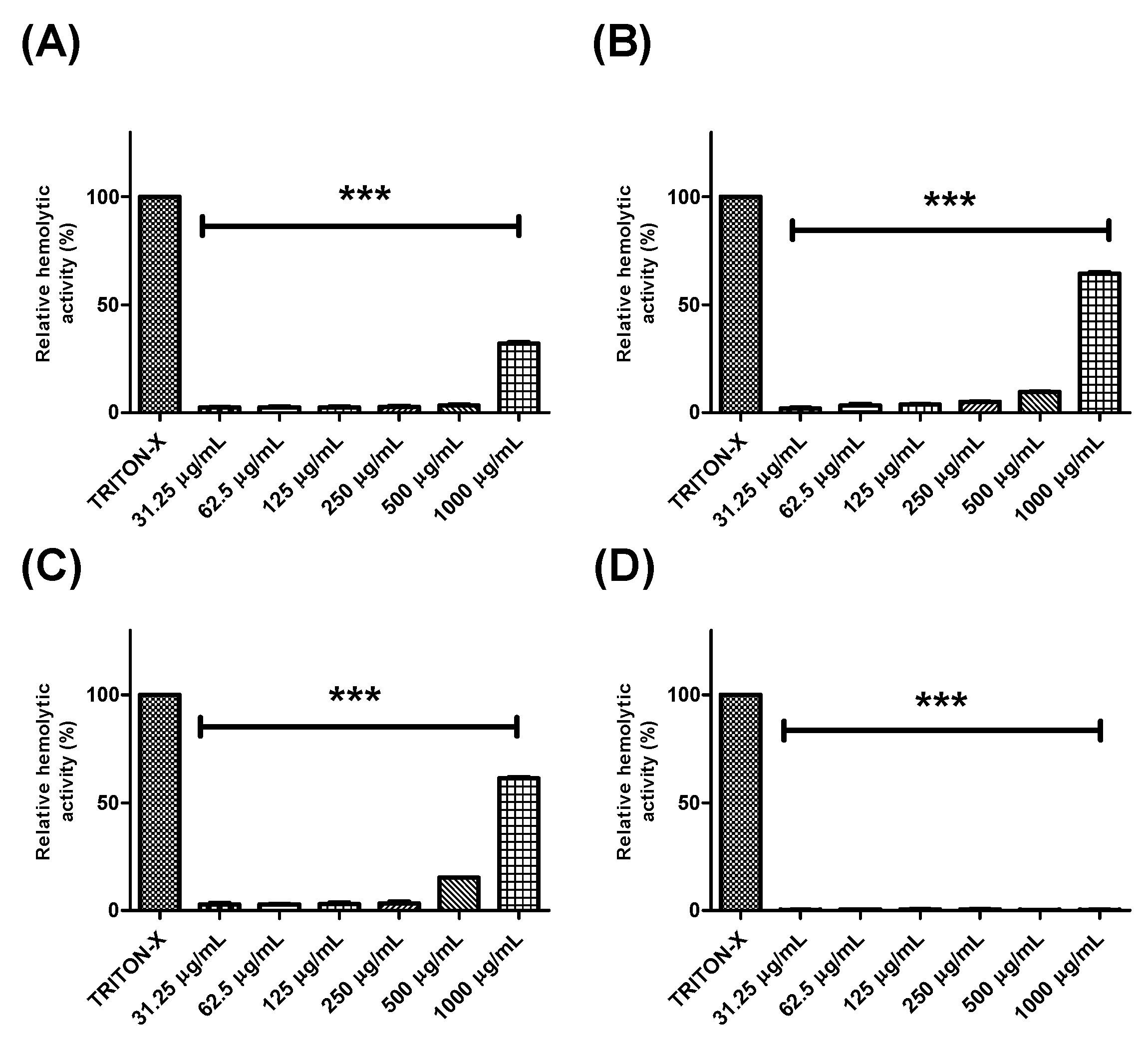
2.3. Evaluation of the Hemolytic Effects of Extracts and Fractions from Buchenavia tetraphylla Leaves
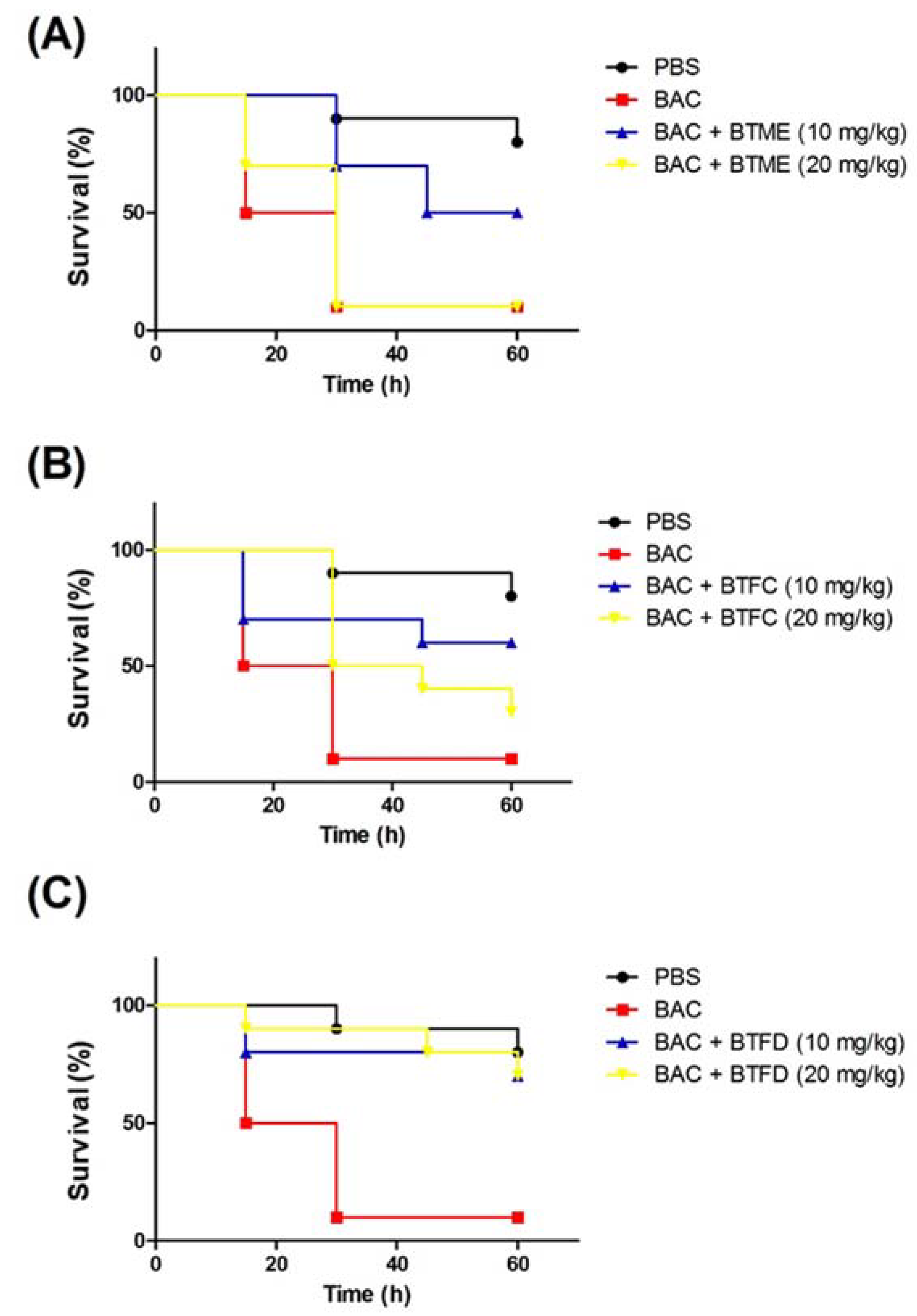
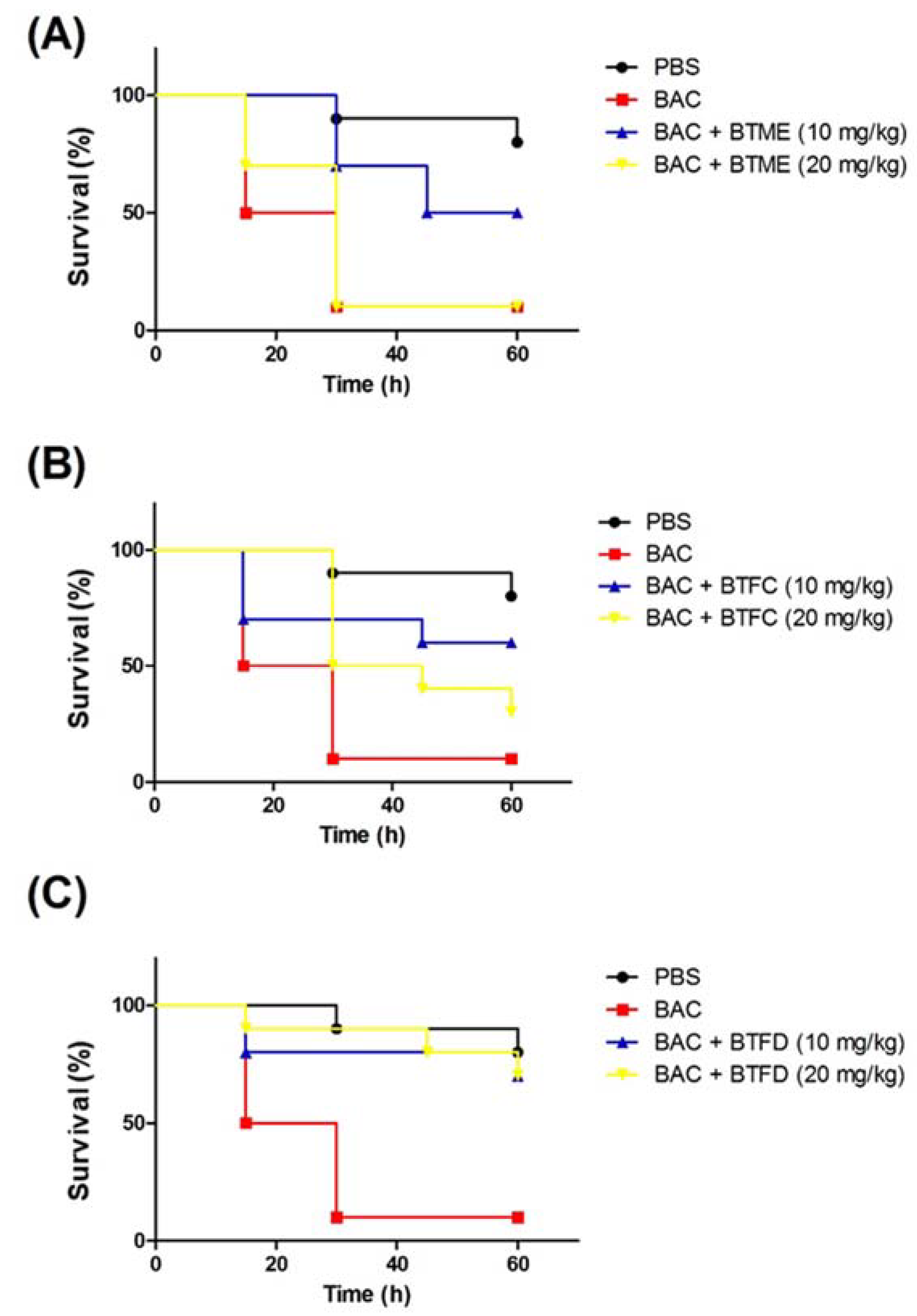
2.4. Effects of Extracts and Fractions from Buchenavia tetraphylla Leaves on the Survival of Tenebrio molitor larvae Submitted to Stress Induced by Heat-Killed Escherichia coli
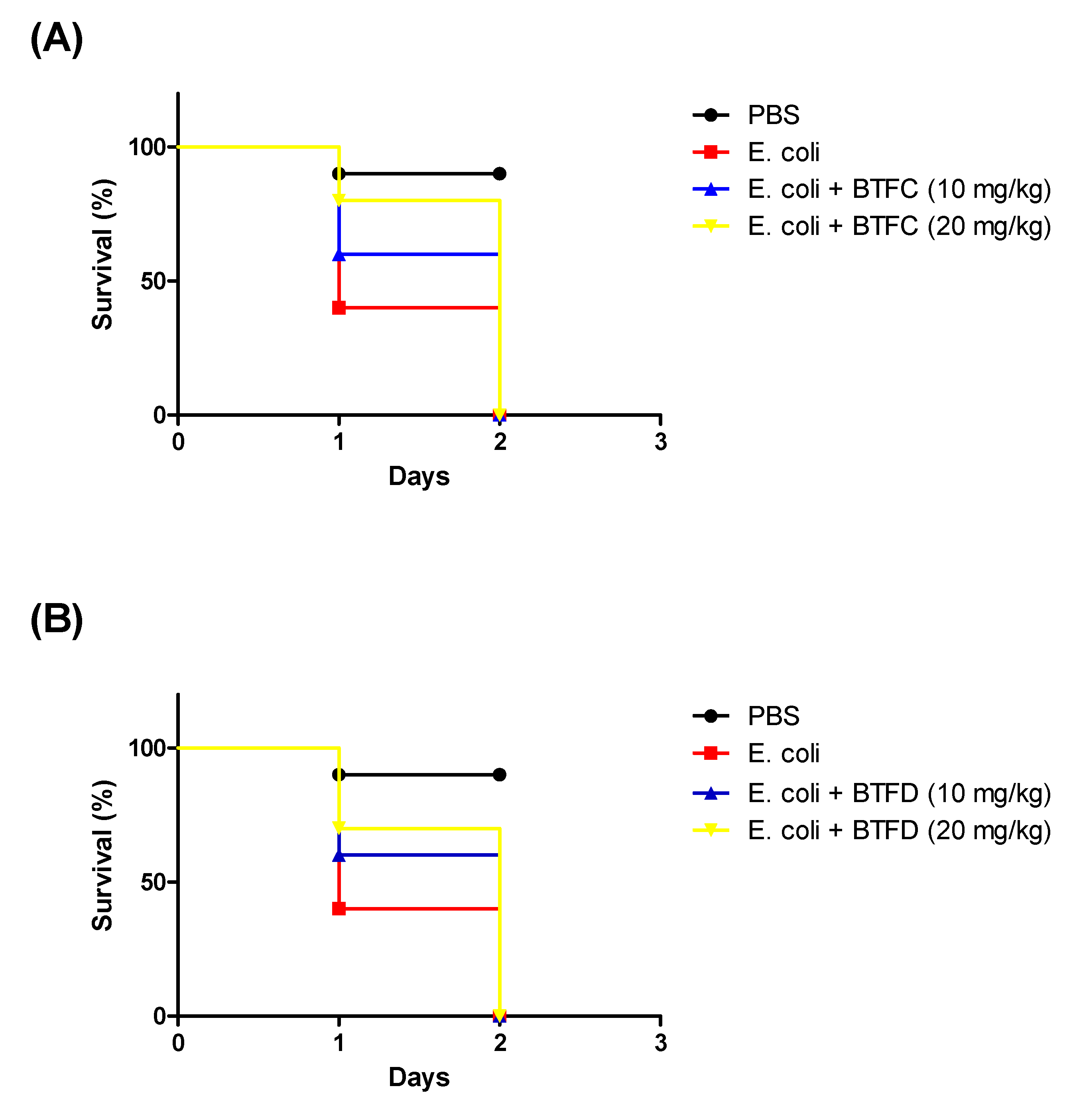
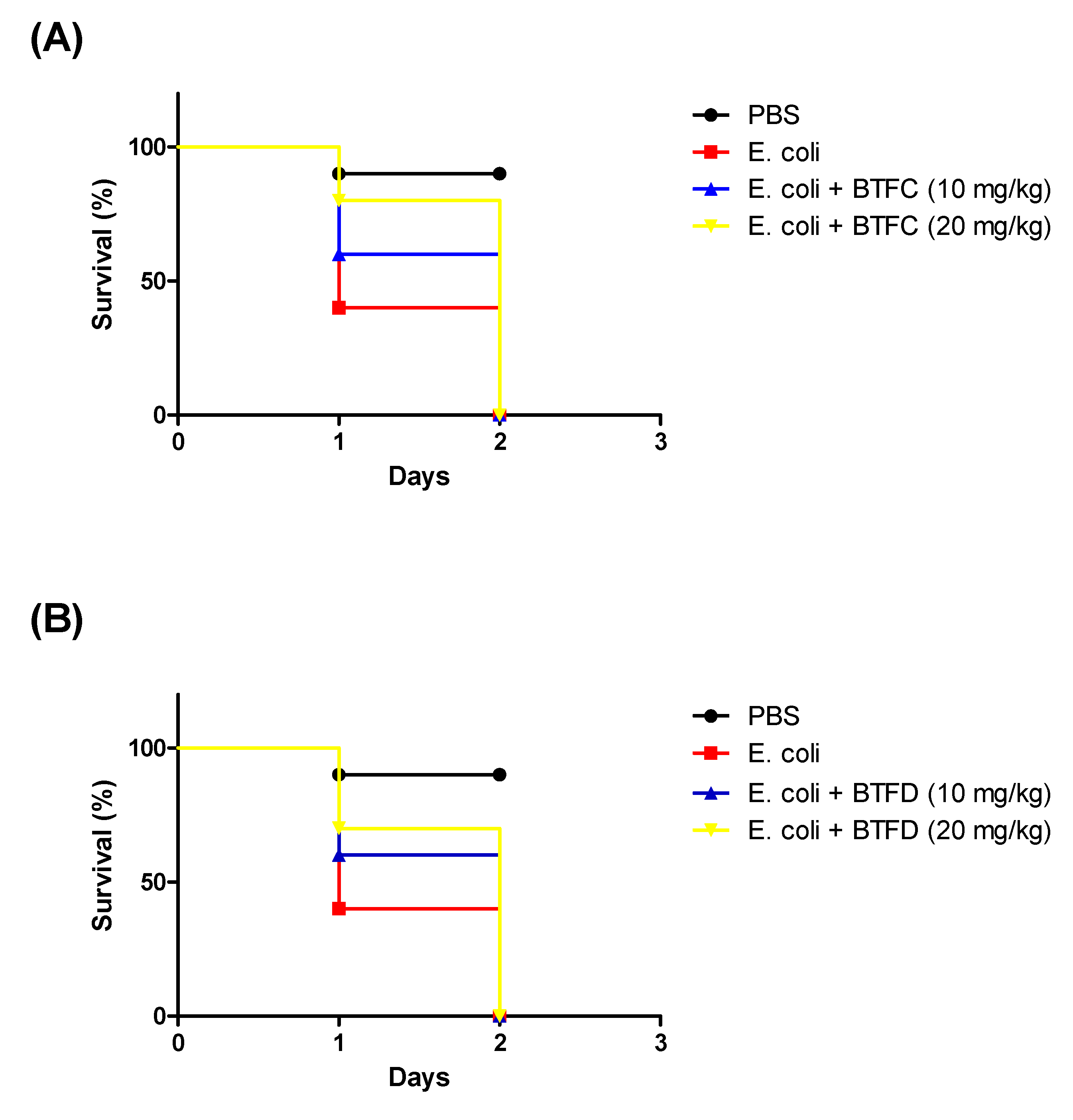
2.5. Effects of Fractions from Buchenavia tetraphylla Leaves in the Lifespan of Tenebrio molitor larvae Infected by Enteroaggregative Escherichia coli
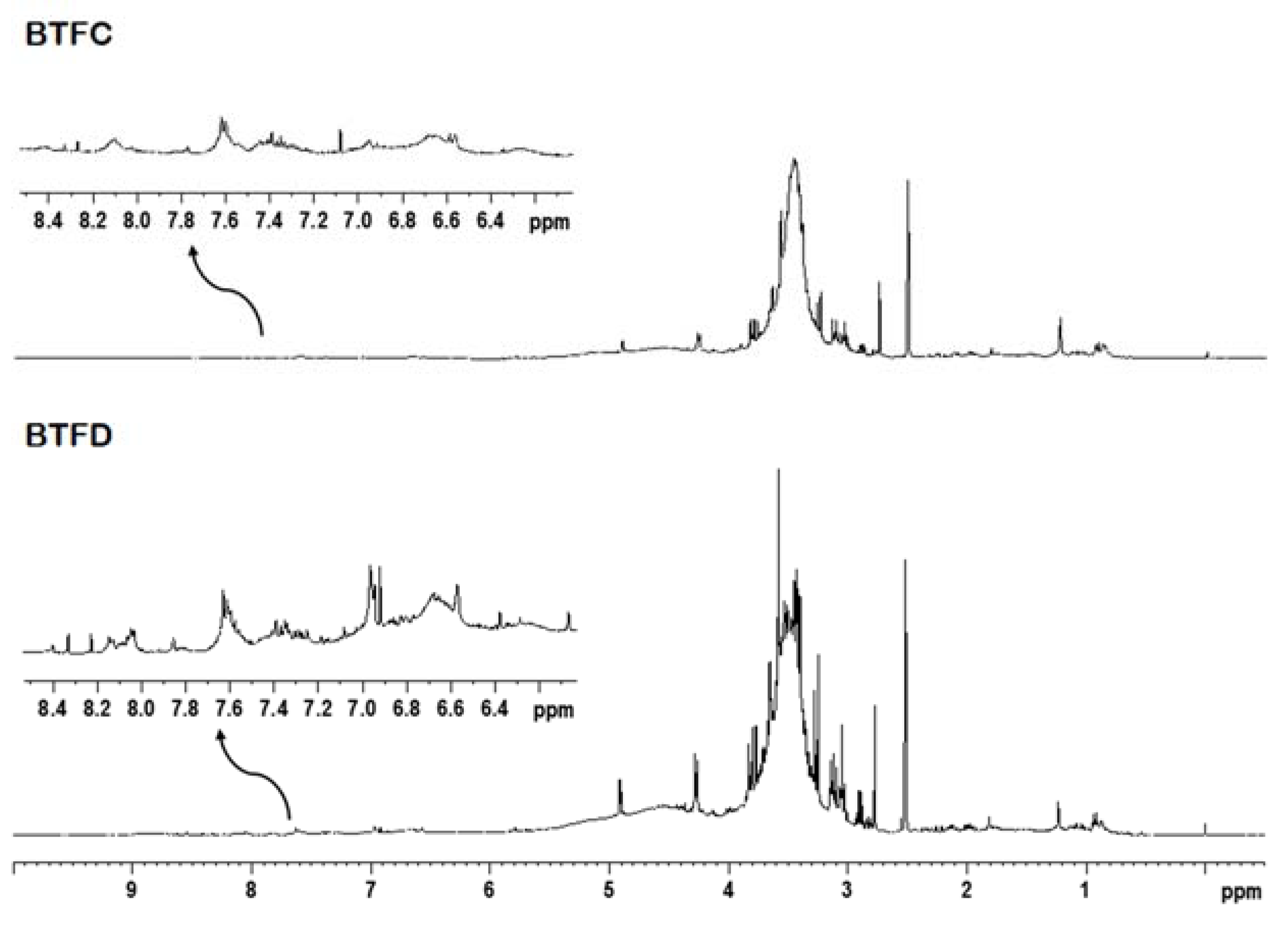
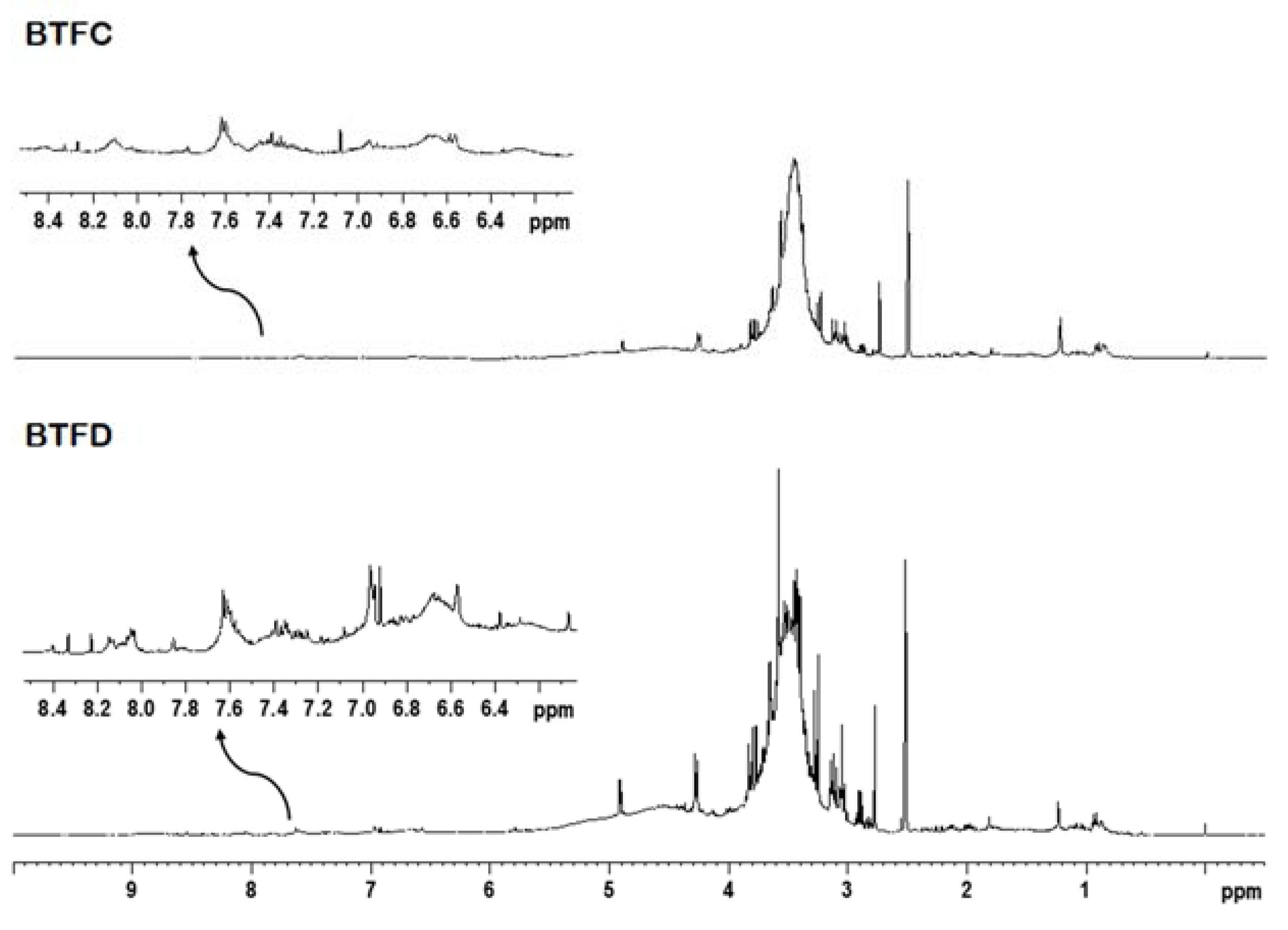
2.6. Nuclear Magnetic Resonance Analysis of Buchenavia tetraphylla Leaves
3. Discussion
4. Material and Methods
4.1. Collection and Extract Preparation
4.2. Fractionation of the Methanolic Extract
4.3. Total Phenolic Content
4.4. Flavonoid Content
4.5. DPPH Assay
4.6. ABTS Assay
4.7. Hemolytic Activity
4.8. Toxicity Model Using Heat-Killed E. coli
4.9. Infection Model Using Enteroaggregative E. coli
4.10. Nuclear Magnetic Resonance Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Poprac, P.; Jomova, K.; Simunkova, M.; Kollar, V.; Rhodes, C.J.; Valko, M. Targeting Free Radicals in Oxidative Stress-Related Human Diseases. Trends Pharmacol. Sci. 2017, 38, 592–607. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, M.J.; Salazar, L.; Grijalva, M. Oxidative Stress Modulation and ROS-Mediated Toxicity in Cancer: A review on in vitro models for plant-derived compounds. Oxid. Med. Cell. Longev. 2017, 2017, 4586068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikara, S.; Nagaprashantha, L.D.; Singhal, J.; Horne, D.; Awasthi, S.; Singhal, S.S. Oxidative stress and dietary phytochemicals: Role in cancer chemoprevention and treatment. Cancer Lett. 2018, 413, 122–134. [Google Scholar] [CrossRef] [PubMed]
- De Andrade Teles, R.B.; Diniz, T.C.; Costa Pinto, T.C.; de Oliveira Junior, R.G.; Gama, E.S.M.; de Lavor, E.M.; Fernandes, A.W.C.; de Oliveira, A.P.; de Almeida Ribeiro, F.P.R.; da Silva, A.A.M.; et al. Flavonoids as therapeutic agents in alzheimer’s and parkinson’s diseases: A systematic review of preclinical evidences. Oxid. Med. Cell. Longev. 2018, 2018, 7043213. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H. Mitochondrial dysfunction and oxidative stress in asthma: Implications for mitochondria-targeted antioxidant therapeutics. Pharmaceuticals 2011, 4, 429–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sillar, J.R.; Germon, Z.P.; DeIuliis, G.N.; Dun, M.D. The role of reactive oxygen species in acute myeloid leukaemia. Int. J. Mol. Sci. 2019, 20, 6003. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, J.P.; de Castro, A.A.; Soares, F.V.; da Cunha, E.F.F.; Ramalho, T.C. Future therapeutic perspectives into the alzheimer’s disease targeting the oxidative stress hypothesis. Molecules 2019, 24, 4410. [Google Scholar] [CrossRef] [Green Version]
- Valko, M.; Leibfritz, D.; Moncol, J.; Cronin, M.T.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Nascimento da Silva, L.C.; Bezerra Filho, C.M.; Paula, R.A.; Silva, E.S.C.S.; Oliveira de Souza, L.I.; Silva, M.V.; Correia, M.T.; Figueiredo, R.C. In vitro cell-based assays for evaluation of antioxidant potential of plant-derived products. Free Radic. Res. 2016, 50, 801–812. [Google Scholar] [CrossRef]
- Halliwell, B. Free radicals and antioxidants: Updating a personal view. Nutr. Rev. 2012, 70, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, L.C.; Alves, N.M.; de Castro, M.C.; Higino, T.M.; da Cunha, C.R.; Pereira, V.R.; da Paz, N.V.; Coelho, L.C.; Correia, M.T.; de Figueiredo, R.C. pCramoll and rCramoll as new preventive agents against the oxidative dysfunction induced by hydrogen peroxide. Oxid. Med. Cell. Longev. 2015, 2015, 520872. [Google Scholar] [PubMed] [Green Version]
- Cacciatore, I.; Fornasari, E.; Baldassarre, L.; Cornacchia, C.; Fulle, S.; Di Filippo, E.S.; Pietrangelo, T.; Pinnen, F. A potent (R)-alpha-bis-lipoyl derivative containing 8-hydroxyquinoline scaffold: Synthesis and biological evaluation of its neuroprotective capabilities in sh-sy5y human neuroblastoma cells. Pharmaceuticals 2013, 6, 54–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marranzano, M.; Rosa, R.L.; Malaguarnera, M.; Palmeri, R.; Tessitori, M.; Barbera, A.C. Polyphenols: Plant sources and food industry applications. Curr. Pharm. Des. 2018, 24, 4125–4130. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, J.S.; Santos, M.; Silva, L.K.R.; Pereira, L.C.L.; Santos, I.A.; da Silva Lannes, S.C.; da Silva, M.V. Natural antioxidants used in meat products: A brief review. Meat Sci. 2019, 148, 181–188. [Google Scholar] [CrossRef] [PubMed]
- Lourenco, S.C.; Moldao-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132. [Google Scholar] [CrossRef] [Green Version]
- da Silva, L.C.; da Silva, C.A., Jr.; de Souza, R.M.; Jose Macedo, A.; da Silva, M.V.; dos Santos Correia, M.T. Comparative analysis of the antioxidant and DNA protection capacities of Anadenanthera colubrina, Libidibia ferrea and Pityrocarpa moniliformis fruits. Food Chem. Toxicol. 2011, 49, 2222–2228. [Google Scholar] [CrossRef]
- Teles Fujishima, M.A.; Silva, N.; Ramos, R.D.S.; Batista Ferreira, E.F.; Santos, K.; Silva, C.; Silva, J.O.D.; Campos Rosa, J.M.; Santos, C. An Antioxidant Potential, Quantum-Chemical and Molecular Docking Study of the Major Chemical Constituents Present in the Leaves of Curatella americana Linn. Pharmaceuticals 2018, 11, 72. [Google Scholar] [CrossRef] [Green Version]
- Agra, M.F.; Baracho, G.S.; Nurit, K.; Basilio, I.J.; Coelho, V.P. Medicinal and poisonous diversity of the flora of “Cariri Paraibano”, Brazil. J. Ethnopharmacol. 2007, 111, 383–395. [Google Scholar] [CrossRef]
- Beutler, J.A.; Cardellina, J.H., 2nd; McMahon, J.B.; Boyd, M.R.; Cragg, G.M. Anti-HIV and cytotoxic alkaloids from Buchenavia capitata. J. Nat. Prod. 1992, 55, 207–213. [Google Scholar] [CrossRef]
- De Oliveira, Y.L.; Nascimento da Silva, L.C.; da Silva, A.G.; Macedo, A.J.; de Araujo, J.M.; Correia, M.T.; da Silva, M.V. Antimicrobial activity and phytochemical screening of Buchenavia tetraphylla (Aubl.) R. A. Howard (Combretaceae: Combretoideae). Sci. World J. 2012, 2012, 849302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavalcanti Filho, J.R.; Silva, T.F.; Nobre, W.Q.; Oliveira de Souza, L.I.; Silva, E.S.F.C.S.; Figueiredo, R.C.; de Gusmao, N.B.; Silva, M.V.; Nascimento da Silva, L.C.; Correia, M.T. Antimicrobial activity of Buchenavia tetraphylla against Candida albicans strains isolated from vaginal secretions. Pharm. Biol. 2017, 55, 1521–1527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, L.M.; Calvert, M.B.; Sperry, J. Flavoalkaloids-Isolation, Biological Activity, and Total Synthesis. Alkaloids Chem. Biol. 2017, 77, 85–115. [Google Scholar] [PubMed]
- Becker, K.; Schroecksnadel, S.; Gostner, J.; Zaknun, C.; Schennach, H.; Uberall, F.; Fuchs, D. Comparison of in vitro tests for antioxidant and immunomodulatory capacities of compounds. Phytomedicine 2014, 21, 164–171. [Google Scholar] [CrossRef]
- Pellegrini, N.; Vitaglione, P.; Granato, D.; Fogliano, V. Twenty-five years of total antioxidant capacity measurement of foods and biological fluids: Merits and limitations. J. Sci. Food Agric. 2018. [Google Scholar] [CrossRef]
- Yun, E.J.; Lee, S.H.; Kim, S.; Kim, S.H.; Kim, K.H. Global profiling of metabolic response of Caenorhabditis elegans against Escherichia coli O157: H7. Process. Biochem. 2017, 53, 36–43. [Google Scholar] [CrossRef]
- Canteri de Souza, P.; Custodio Caloni, C.; Wilson, D.; Sergio Almeida, R. An Invertebrate Host to Study Fungal Infections, Mycotoxins and Antifungal Drugs: Tenebrio molitor. J. Fungi 2018, 4, 125. [Google Scholar] [CrossRef] [Green Version]
- Guo, Z.; Pfohl, K.; Karlovsky, P.; Dehne, H.W.; Altincicek, B. Dissemination of Fusarium proliferatum by mealworm beetle Tenebrio molitor. PLoS ONE 2018, 13, e0204602. [Google Scholar] [CrossRef]
- Czarniewska, E.; Urbanski, A.; Chowanski, S.; Kuczer, M. The long-term immunological effects of alloferon and its analogues in the mealworm Tenebrio molitor. Insect Sci. 2018, 25, 429–438. [Google Scholar] [CrossRef]
- McGonigle, J.E.; Purves, J.; Rolff, J. Intracellular survival of Staphylococcus aureus during persistent infection in the insect Tenebrio molitor. Dev. Comp. Immunol. 2016, 59, 34–38. [Google Scholar] [CrossRef]
- Seong, J.H.; Jo, Y.H.; Seo, G.W.; Park, S.; Park, K.B.; Cho, J.H.; Ko, H.J.; Kim, C.E.; Patnaik, B.B.; Jun, S.A.; et al. Molecular Cloning and Effects of Tm14-3-3zeta-Silencing on Larval Survivability Against, E. coli and C. albicans in Tenebrio molitor. Genes 2018, 9, 330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, P.C.; Morey, A.T.; Castanheira, G.M.; Bocate, K.P.; Panagio, L.A.; Ito, F.A.; Furlaneto, M.C.; Yamada-Ogatta, S.F.; Costa, I.N.; Mora-Montes, H.M.; et al. Tenebrio molitor (Coleoptera: Tenebrionidae) as an alternative host to study fungal infections. J. Microbiol. Methods 2015, 118, 182–186. [Google Scholar] [CrossRef] [Green Version]
- Johnston, P.R.; Makarova, O.; Rolff, J. Inducible defenses stay up late: Temporal patterns of immune gene expression in Tenebrio molitor. G3 2013, 4, 947–955. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Kim, S.K.; So, Y.I.; Park, H.Y.; Li, X.H.; Yeom, D.H.; Lee, M.N.; Lee, B.L.; Lee, J.H. Protease IV, a quorum sensing-dependent protease of Pseudomonas aeruginosa modulates insect innate immunity. Mol. Microbiol. 2014, 94, 1298–1314. [Google Scholar] [CrossRef] [PubMed]
- Tindwa, H.; Jo, Y.H.; Patnaik, B.B.; Lee, Y.S.; Kang, S.S.; Han, Y.S. Molecular cloning and characterization of autophagy-related gene TmATG8 in Listeria-invaded hemocytes of Tenebrio molitor. Dev. Comp. Immunol. 2015, 51, 88–98. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.T.; Lee, M.R.; Lee, S.J.; Kim, S.; Nai, Y.S.; Kim, J.S. Tenebrio molitor Gram-negative-binding protein 3 (TmGNBP3) is essential for inducing downstream antifungal Tenecin 1 gene expression against infection with Beauveria bassiana JEF-007. Insect Sci. 2018, 25, 969–977. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Ze, S.Z.; Stanley, D.W.; Yang, B. Parasitization by Scleroderma guani influences expression of superoxide dismutase genes in Tenebrio molitor. Arch. Insect Biochem. Physiol. 2014, 87, 40–52. [Google Scholar] [CrossRef]
- Li, X.; Liu, Q.; Lewis, E.E.; Tarasco, E. Activity changes of antioxidant and detoxifying enzymes in Tenebrio molitor (Coleoptera: Tenebrionidae) larvae infected by the entomopathogenic nematode Heterorhabditis beicherriana (Rhabditida: Heterorhabditidae). Parasitol. Res. 2016, 115, 4485–4494. [Google Scholar] [CrossRef] [Green Version]
- Gulevsky, A.K.; Relina, L.I.; Grishchenkova, Y.A. Variations of the antioxidant system during development of the cold-tolerant beetle, Tenebrio molitor. Cryo Lett. 2006, 27, 283–290. [Google Scholar]
- Medina-Gomez, H.; Farriols, M.; Santos, F.; Gonzalez-Hernandez, A.; Torres-Guzman, J.C.; Lanz, H.; Contreras-Garduno, J. Pathogen-produced catalase affects immune priming: A potential pathogen strategy. Microb. Pathog. 2018, 125, 93–95. [Google Scholar] [CrossRef]
- Peixoto, H.; Roxo, M.; Silva, E.; Valente, K.; Braun, M.; Wang, X.; Wink, M. Bark Extract of the Amazonian Tree Endopleura uchi (Humiriaceae) Extends Lifespan and Enhances Stress Resistance in Caenorhabditis elegans. Molecules 2019, 24, 915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohn, I.; Raschke, S.; Aschner, M.; Tuck, S.; Kuehnelt, D.; Kipp, A.; Schwerdtle, T.; Bornhorst, J. Treatment of Caenorhabditis elegans with small selenium species enhances antioxidant defense systems. Mol. Nutr. Food Res. 2019, 63, e1801304. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, G.R.; Brighenti, F.L.; Delbem, A.C.; Delbem, A.C.; Khouri, S.; Gontijo, A.V.; Pascoal, A.C.; Salvador, M.J.; Koga-Ito, C.Y. Antifungal activity of extracts and isolated compounds from Buchenavia tomentosa on Candida albicans and non-albicans. Future Microbiol. 2015, 10, 917–927. [Google Scholar] [CrossRef] [PubMed]
- Teodoro, G.R.; Gontijo, A.V.L.; Salvador, M.J.; Tanaka, M.H.; Brighenti, F.L.; Delbem, A.C.B.; Delbem, A.C.B.; Koga-Ito, C.Y. Effects of acetone fraction from Buchenavia tomentosa aqueous extract and gallic acid on Candida albicans biofilms and virulence factors. Front. Microbiol. 2018, 9, 647. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez-Burgos, E.; Gomez-Serranillos, M.P. Terpene compounds in nature: A review of their potential antioxidant activity. Curr. Med. Chem. 2012, 19, 5319–5341. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef]
- Dai, J.; Mumper, R.J. Plant phenolics: Extraction, analysis and their antioxidant and anticancer properties. Molecules 2010, 15, 7313–7352. [Google Scholar] [CrossRef]
- Castaneda-Arriaga, R.; Perez-Gonzalez, A.; Reina, M.; Alvarez-Idaboy, J.R.; Galano, A. Comprehensive investigation of the antioxidant and pro-oxidant effects of phenolic compounds: A double-edged sword in the context of oxidative stress? J. Phys. Chem. B 2018, 122, 6198–6214. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Viticult. 1965, 16, 144–158. [Google Scholar]
- Woisky, R.G.; Salatino, A. Analysis of propolis: Some parameters and procedures for chemical quality control. J. Apicult. Res. 1998, 37, 99–105. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BTHE | BTCE | BTEE | BTME | Trolox | |
|---|---|---|---|---|---|
| Yield (%) | 5.30 | 14.48 | 10.45 | 14.44 | - |
| Phenolic compounds content (mg GAE/mg) | 9.45 ± 1.29 a | 26.53 ± 0.50 b | 116.65 ± 10.26 c | 123.03 ± 1.23 c | - |
| Flavonoid content (mg QE/mg) | 10.03 ± 0.14 a | 14.28 ± 0.48 b | 24.92 ± 0.45 c | 108.89 ± 0.06 d | - |
| DPPH (EC50 μg/mL) | 6826.45 | 3779.98 | 562.75 | 79.04 | 44.10 |
| BTFA | BTFB | BTFC | BTFD | BTFE | BTFF | BTFG | BTFH | BTFI | |
|---|---|---|---|---|---|---|---|---|---|
| Yields (%) | 0.10 | 0.29 | 1.22 | 2.53 | 2.04 | 0.96 | 0.46 | 0.56 | 0.98 |
| Phenolic contents (μg GAE/μg) | 49.44 ± 1.86 a | 107.20 ± 7.23 b | 155.67 ± 3.40 c | 168.98 ± 1.81 c | 49.80 ± 6.52 a | 72.83 ± 1.13 d | 127.62 ± 15.60 e | 55.61 ± 3.01 a | 110.10 ± 0.62 b |
| Flavonoid contents (μg QE/μg) | 16.80 ± 1.76 a | 12.65 ± 0.76 a | 68.27 ± 2.35 b | 56.01 ± 4.52 c | 14.01 ± 1.72 a | 4.28 ± 0.11 d | 45.27 ± 3.37 e | 0.86 ± 0.21 d | 39.29 ± 2.36 e |
| DPPH (EC50 μg/mL) | 2480.22 | 562.75 | 50.41 | 237.76 | 4132.98 | 2355.09 | 294.38 | 2578.18 | 376.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, T.F.; Cavalcanti Filho, J.R.N.; Barreto Fonsêca, M.M.L.; Santos, N.M.d.; Barbosa da Silva, A.C.; Zagmignan, A.; Abreu, A.G.; Sant’Anna da Silva, A.P.; Lima, V.L.d.M.; Silva, N.H.d.; et al. Products Derived from Buchenavia tetraphylla Leaves Have In Vitro Antioxidant Activity and Protect Tenebrio molitor Larvae against Escherichia coli-Induced Injury. Pharmaceuticals 2020, 13, 46. https://doi.org/10.3390/ph13030046
Silva TF, Cavalcanti Filho JRN, Barreto Fonsêca MML, Santos NMd, Barbosa da Silva AC, Zagmignan A, Abreu AG, Sant’Anna da Silva AP, Lima VLdM, Silva NHd, et al. Products Derived from Buchenavia tetraphylla Leaves Have In Vitro Antioxidant Activity and Protect Tenebrio molitor Larvae against Escherichia coli-Induced Injury. Pharmaceuticals. 2020; 13(3):46. https://doi.org/10.3390/ph13030046
Chicago/Turabian StyleSilva, Tiago Fonseca, José Robson Neves Cavalcanti Filho, Mariana Mirelle Lima Barreto Fonsêca, Natalia Medeiros dos Santos, Ana Carolina Barbosa da Silva, Adrielle Zagmignan, Afonso Gomes Abreu, Ana Paula Sant’Anna da Silva, Vera Lúcia de Menezes Lima, Nicácio Henrique da Silva, and et al. 2020. "Products Derived from Buchenavia tetraphylla Leaves Have In Vitro Antioxidant Activity and Protect Tenebrio molitor Larvae against Escherichia coli-Induced Injury" Pharmaceuticals 13, no. 3: 46. https://doi.org/10.3390/ph13030046
APA StyleSilva, T. F., Cavalcanti Filho, J. R. N., Barreto Fonsêca, M. M. L., Santos, N. M. d., Barbosa da Silva, A. C., Zagmignan, A., Abreu, A. G., Sant’Anna da Silva, A. P., Lima, V. L. d. M., Silva, N. H. d., Dutra, L. M., Almeida, J. R. G. d. S., Silva, M. V. d., Correia, M. T. d. S., & Nascimento da Silva, L. C. (2020). Products Derived from Buchenavia tetraphylla Leaves Have In Vitro Antioxidant Activity and Protect Tenebrio molitor Larvae against Escherichia coli-Induced Injury. Pharmaceuticals, 13(3), 46. https://doi.org/10.3390/ph13030046