Methylxanthines: Potential Therapeutic Agents for Glioblastoma

 , , , and
, , , and
Abstract
1. Introduction
2. Phosphodiesterases in Glioblastoma (GBM)
3. Methylxanthines
3.1. Caffeine
3.2. Theophylline
3.3. Theobromine
4. Methylxanthines and Cellular Differentiation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Louis, D.N.; Perry, A.; Reifenberger, G.; von Deimling, A.; Figarella-Branger, D.; Cavenee, W.K.; Ohgaki, H.; Wiestler, O.D.; Kleihues, P.; Ellison, D.W. The 2016 World Health Organization Classification of Tumors of the Central Nervous System: A summary. Acta Neuropathol. 2016, 131, 803–820. [Google Scholar] [CrossRef] [PubMed]
- Ostrom, Q.T.; Gittleman, H.; Liao, P.; Vecchione-Koval, T.; Wolinsky, Y.; Kruchko, C.; Barnholtz-Sloan, J.S. CBTRUS Statistical Report: Primary brain and other central nervous system tumors diagnosed in the United States in 2010–2014. Neuro Oncol. 2017, 19, v1–v88. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, G.S.; Dzhenkov, D.; Ghenev, P.; Iliev, B.; Enchev, Y.; Tonchev, A.B. Cell biology of glioblastoma multiforme: From basic science to diagnosis and treatment. Med. Oncol. 2018, 35, 27. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Taillibert, S.; Kanner, A.; Read, W.; Steinberg, D.; Lhermitte, B.; Toms, S.; Idbaih, A.; Ahluwalia, M.S.; Fink, K.; et al. Effect of Tumor-Treating Fields Plus Maintenance Temozolomide vs. Maintenance Temozolomide Alone on Survival in Patients with Glioblastoma: A Randomized Clinical Trial. JAMA 2017, 318, 2306–2316. [Google Scholar] [CrossRef] [PubMed]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Otani, Y.; Kurozumi, K.; Date, I. Phenotypic Transition as a Survival Strategy of Glioma. Neurol. Med. Chir. 2016, 56, 387–395. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Cesarini, V.; Martini, M.; Vitiani, L.R.; Gravina, G.L.; Di Agostino, S.; Graziani, G.; D’Alessandris, Q.G.; Pallini, R.; Larocca, L.M.; Rossi, P.; et al. Type 5 phosphodiesterase regulates glioblastoma multiforme aggressiveness and clinical outcome. Oncotarget 2017, 8, 13223–13239. [Google Scholar] [CrossRef]
- Vengoji, R.; Macha, M.A.; Batra, S.K.; Shonka, N.A. Natural products: A hope for glioblastoma patients. Oncotarget 2018, 9, 22194–22219. [Google Scholar] [CrossRef]
- Shahcheraghi, S.H.; Zangui, M.; Lotfi, M.; Ghayour-Mobarhan, M.; Ghorbani, A.; Jaliani, H.Z.; Sadeghnia, H.R.; Sahebkar, A. Therapeutic Potential of Curcumin in the Treatment of Glioblastoma Multiforme. Curr. Pharm. Des. 2019, 25, 333–342. [Google Scholar] [CrossRef]
- Le, C.T.; Leenders, W.P.J.; Molenaar, R.J.; van Noorden, C.J.F. Effects of the Green Tea Polyphenol Epigallocatechin-3-Gallate on Glioma: A Critical Evaluation of the Literature. Nutr. Cancer 2018, 70, 317–333. [Google Scholar] [CrossRef]
- Arnaud, M.J. Pharmacokinetics and metabolism of natural methylxanthines in animal and man. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2011; pp. 33–91. [Google Scholar]
- Azevedo, M.F.; Faucz, F.R.; Bimpaki, E.; Horvath, A.; Levy, I.; de Alexandre, R.B.; Ahmad, F.; Manganiello, V.; Stratakis, C.A. Clinical and molecular genetics of the phosphodiesterases (PDEs). Endocr. Rev. 2014, 35, 195–233. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, S.; Hadjighassem, M.; Vousooghi, N.; Parvaresh, M.; Arbabi, F.; Amini, N.; Joghataei, M.T. The Role of Protein Kinase B Signaling Pathway in Anti-Cancer Effect of Rolipram on Glioblastoma Multiforme: An In Vitro Study. Basic Clin. Neurosci. 2017, 8, 325–336. [Google Scholar] [CrossRef] [PubMed]
- Moon, E.-Y.; Lee, G.-H.; Lee, M.-S.; Kim, H.-M.; Lee, J.-W. Phosphodiesterase inhibitors control A172 human glioblastoma cell death through cAMP-mediated activation of protein kinase A and Epac1/Rap1 pathways. Life Sci. 2012, 90, 373–380. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Teraoka, A.; Matsutani, M.; Machiyama, N.; Tsuchida, T.; Hatanaka, H.; Sano, K. B-6. Effect of Cyclic AMP on the Malignant Brain Tumor Cells. Neurol. Med. Chir. 1971, 11, 231. [Google Scholar] [CrossRef]
- Sato, S.; Sugimura, T.; Yoda, K.; Fujimura, S. Morphological Differentiation of Cultured Mouse Glioblastoma Cells Induced by Dibutyryl Cyclic Adenosine Monophosphate. Cancer Res. 1975, 35, 2494–2499. [Google Scholar]
- Takanaga, H.; Yoshitake, T.; Hara, S.; Yamasaki, C.; Kunimoto, M. cAMP-induced astrocytic differentiation of C6 glioma cells is mediated by autocrine interleukin-6. J. Biol. Chem. 2004, 279, 15441–15447. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, N.; Miwa, S.; Hitomi, Y.; Nakamura, H.; Tsuchiya, H.; Yachie, A. Theobromine, the primary methylxanthine found in Theobroma cacao, prevents malignant glioblastoma proliferation by negatively regulating phosphodiesterase-4, extracellular signal-regulated kinase, Akt/mammalian target of rapamycin kinase, and nuclear fact. Nutr. Cancer 2014, 66, 419–423. [Google Scholar] [CrossRef]
- Stewart, D.J.; Hugenholtz, H.; DaSilva, V.; Benoit, B.; Richard, M.; Russell, N.; Maroun, J.; Verma, S. Cytosine arabinoside plus cisplatin and other drugs as chemotherapy for gliomas. Semin. Oncol. 1987, 14, 110–115. [Google Scholar]
- Janss, A.J.; Levow, C.; Bernhard, E.J.; Muschel, R.J.; McKenna, W.G.; Sutton, L.; Phillips, P.C. Caffeine and staurosporine enhance the cytotoxicity of cisplatin and camptothecin in human brain tumor cell lines. Exp. Cell Res. 1998, 243, 29–38. [Google Scholar] [CrossRef]
- Chen, Y.; Chou, W.C.; Ding, Y.M.; Wu, Y.C. Caffeine inhibits migration in glioma cells through the ROCK-FAK pathway. Cell. Physiol. Biochem. 2014, 33, 1888–1898. [Google Scholar] [CrossRef]
- Sinn, B.; Tallen, G.; Schroeder, G.; Grassl, B.; Schulze, J.; Budach, V.; Tinhofer, I. Caffeine Confers Radiosensitization of PTEN-Deficient Malignant Glioma Cells by Enhancing Ionizing Radiation-Induced G1 Arrest and Negatively Regulating Akt Phosphorylation. Mol. Cancer Ther. 2010, 9, 480–488. [Google Scholar] [CrossRef] [PubMed]
- Ku, B.M.; Lee, Y.K.; Jeong, J.Y.; Ryu, J.; Choi, J.; Kim, J.S.; Cho, Y.W.; Roh, G.S.; Kim, H.J.; Cho, G.J.; et al. Caffeine inhibits cell proliferation and regulates PKA/GSK3β pathways in U87MG human glioma cells. Mol. Cells 2011, 31, 275–279. [Google Scholar] [CrossRef] [PubMed]
- Kleppisch, T. Phosphodiesterases in the central nervous system. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 71–92. [Google Scholar]
- Yarla, N.S.; Gali, H.; Pathuri, G.; Smriti, S.; Farooqui, M.; Panneerselvam, J.; Kumar, G.; Madka, V.; Rao, C.V. Targeting the paracrine hormone-dependent guanylate cyclase/cGMP/phosphodiesterases signaling pathway for colorectal cancer prevention. Semin. Cancer Biol. 2019, 56, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Murray, F.; Zahno, A.; Kanter, J.R.; Chou, D.; Suda, R.; Fenlon, M.; Rassenti, L.; Cottam, H.; Kipps, T.J.; et al. Cyclic nucleotide phosphodiesterase profiling reveals increased expression of phosphodiesterase 7B in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2008, 105, 19532–19537. [Google Scholar] [CrossRef] [PubMed]
- Vatter, S.; Pahlke, G.; Deitmer, J.W.; Eisenbrand, G. Differential phosphodiesterase expression and cytosolic Ca2+ in human CNS tumour cells and in non-malignant and malignant cells of rat origin. J. Neurochem. 2005, 93, 321–329. [Google Scholar] [CrossRef]
- Goldhoff, P.; Warrington, N.M.; Limbrick, D.D., Jr.; Hope, A.; Woerner, B.M.; Jackson, E.; Perry, A.; Piwnica-Worms, D.; Rubin, J.B. Targeted inhibition of cyclic AMP phosphodiesterase-4 promotes brain tumor regression. Clin. Cancer Res. 2008, 14, 7717–7725. [Google Scholar] [CrossRef] [PubMed]
- Rowther, F.B.; Wei, W.; Dawson, T.P.; Ashton, K.; Singh, A.; Madiesse-Timchou, M.P.; Thomas, D.G.T.; Darling, J.L.; Warr, T. Cyclic nucleotide phosphodiesterase-1C (PDE1C) drives cell proliferation, migration and invasion in glioblastoma multiforme cells in vitro. Mol. Carcinog. 2016, 55, 268–279. [Google Scholar] [CrossRef]
- Michibata, H.; Yanaka, N.; Kanoh, Y.; Okumura, K.; Omori, K. Human Ca2+/calmodulin-dependent phosphodiesterase PDE1A: Novel splice variants, their specific expression, genomic organization, and chromosomal localization. Biochim. Biophys. Acta 2001, 1517, 278–287. [Google Scholar] [CrossRef]
- Reed, T.M.; Browning, J.E.; Blough, R.I.; Vorhees, C.V.; Repaske, D.R. Genomic structure and chromosome location of the murine PDE1B phosphodiesterase gene. Mamm. Genome 1998, 9, 571–576. [Google Scholar] [CrossRef]
- Loughney, K.; Martins, T.J.; Harris, E.A.; Sadhu, K.; Hicks, J.B.; Sonnenburg, W.K.; Beavo, J.A.; Ferguson, K. Isolation and characterization of cDNAs corresponding to two human calcium, calmodulin-regulated, 3′,5′-cyclic nucleotide phosphodiesterases. J. Biol. Chem. 1996, 271, 796–806. [Google Scholar] [CrossRef]
- Rosman, G.J.; Martins, T.J.; Sonnenburg, W.K.; Beavo, J.A.; Ferguson, K.; Loughney, K. Isolation and characterization of human cDNAs encoding a cGMP-stimulated 3′,5′-cyclic nucleotide phosphodiesterase. Gene 1997, 191, 89–95. [Google Scholar] [CrossRef]
- Doecke, J.D.; Wang, Y.; Baggerly, K. Co-localized genomic regulation of miRNA and mRNA via DNA methylation affects survival in multiple tumor types. Cancer Genet. 2016, 209, 463–473. [Google Scholar] [CrossRef] [PubMed]
- Meacci, E.; Taira, M.; Moos, M.; Smith, C.J.; Movsesian, M.A.; Degerman, E.; Belfrage, P.; Manganiello, V. Molecular cloning and expression of human myocardial cGMP-inhibited cAMP phosphodiesterase. Proc. Natl. Acad. Sci. USA 1992, 89, 3721–3725. [Google Scholar] [CrossRef] [PubMed]
- Yasmeen, S.; Akram, B.H.; Hainsworth, A.H.; Kruuse, C. Cyclic nucleotide phosphodiesterases (PDEs) and endothelial function in ischaemic stroke. A review. Cell. Signal. 2019, 61, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Miki, T.; Taira, M.; Hockman, S.; Shimada, F.; Lieman, J.; Napolitano, M.; Ward, D.; Taira, M.; Makino, H.; Manganiello, V.C. Characterization of the cDNA and gene encoding human PDE3B, the cGIP1 isoform of the human cyclic GMP-inhibited cyclic nucleotide phosphodiesterase family. Genomics 1996, 36, 476–485. [Google Scholar] [CrossRef] [PubMed]
- Houslay, M.D.; Baillie, G.S.; Maurice, D.H. cAMP-Specific phosphodiesterase-4 enzymes in the cardiovascular system: A molecular toolbox for generating compartmentalized cAMP signaling. Circ. Res. 2007, 100, 950–966. [Google Scholar] [CrossRef] [PubMed]
- Conti, M.; Richter, W.; Mehats, C.; Livera, G.; Park, J.-Y.; Jin, C. Cyclic AMP-specific PDE4 phosphodiesterases as critical components of cyclic AMP signaling. J. Biol. Chem. 2003, 278, 5493–5496. [Google Scholar] [CrossRef] [PubMed]
- Ramezani, S.; Vousooghi, N.; Kapourchali, F.R.; Hadjighasem, M.; Hayat, P.; Amini, N.; Joghataei, M.T. Rolipram potentiates bevacizumab-induced cell death in human glioblastoma stem-like cells. Life Sci. 2017, 173, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Bao, Z.; Feng, Y.; Wang, H.; Zhang, C.; Sun, L.; Yan, Z.; Liu, Q.; Guo, T.; Li, M.; Yang, X.; et al. Integrated analysis using methylation and gene expression microarrays reveals PDE4C as a prognostic biomarker in human glioma. Oncol. Rep. 2014, 32, 250–260. [Google Scholar] [CrossRef]
- Lin, C.-S.; Chow, S.; Lau, A.; Tu, R.; Lue, T.F. Human PDE5A gene encodes three PDE5 isoforms from two alternate promoters. Int. J. Impot. Res. 2002, 14, 15–24. [Google Scholar] [CrossRef]
- Corton, M.; Blanco, M.J.; Torres, M.; Sanchez-Salorio, M.; Carracedo, A.; Brion, M. Identification of a novel mutation in the human PDE6A gene in autosomal recessive retinitis pigmentosa: Homology with the nmf28/nmf28 mice model. Clin. Genet. 2010, 78, 495–498. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, V.K.L.; Takiuti, J.T.; Jauregui, R.; Lima, L.H.; Tsang, S.H. Structural disease progression in PDE6-associated autosomal recessive retinitis pigmentosa. Ophthalmic Genet. 2018, 39, 610–614. [Google Scholar] [CrossRef]
- Piriev, N.I.; Viczian, A.S.; Ye, J.; Kerner, B.; Korenberg, J.R.; Farber, D.B. Gene structure and amino acid sequence of the human cone photoreceptor cGMP-phosphodiesterase alpha’ subunit (PDEA2) and its chromosomal localization to 10q24. Genomics 1995, 28, 429–435. [Google Scholar] [CrossRef] [PubMed]
- Brooks, M.D.; Jackson, E.; Warrington, N.M.; Luo, J.; Forys, J.T.; Taylor, S.; Mao, D.D.; Leonard, J.R.; Kim, A.H.; Piwnica-Worms, D.; et al. PDE7B is a novel, prognostically significant mediator of glioblastoma growth whose expression is regulated by endothelial cells. PLoS ONE 2014, 9, e107397. [Google Scholar] [CrossRef] [PubMed]
- Han, P.; Fletcher, C.F.; Copeland, N.G.; Jenkins, N.A.; Yaremko, L.M.; Michaeli, T. Assignment of the mouse Pde7A gene to the proximal region of chromosome 3 and of the human PDE7A gene to chromosome 8q13. Genomics 1998, 48, 275–276. [Google Scholar] [CrossRef] [PubMed]
- Hetman, J.M.; Soderling, S.H.; Glavas, N.A.; Beavo, J.A. Cloning and characterization of PDE7B, a cAMP-specific phosphodiesterase. Proc. Natl. Acad. Sci. USA 2000, 97, 472–476. [Google Scholar] [CrossRef] [PubMed]
- Goto, M.; Murakawa, M.; Kadoshima-Yamaoka, K.; Tanaka, Y.; Inoue, H.; Murafuji, H.; Hayashi, Y.; Miura, K.; Nakatsuka, T.; Nagahira, K.; et al. Phosphodiesterase 7A inhibitor ASB16165 suppresses proliferation and cytokine production of NKT cells. Cell. Immunol. 2009, 258, 147–151. [Google Scholar] [CrossRef]
- Wang, P.; Wu, P.; Egan, R.W.; Billah, M.M. Human phosphodiesterase 8A splice variants: Cloning, gene organization, and tissue distribution. Gene 2001, 280, 183–194. [Google Scholar] [CrossRef]
- Hayashi, M.; Shimada, Y.; Nishimura, Y.; Hama, T.; Tanaka, T. Genomic organization, chromosomal localization, and alternative splicing of the human phosphodiesterase 8B gene. Biochem. Biophys. Res. Commun. 2002, 297, 1253–1258. [Google Scholar] [CrossRef]
- Basole, C.P.; Nguyen, R.K.; Lamothe, K.; Vang, A.; Clark, R.; Baillie, G.S.; Epstein, P.M.; Brocke, S. PDE8 controls CD4+ T cell motility through the PDE8A-Raf-1 kinase signaling complex. Cell. Signal. 2017, 40, 62–72. [Google Scholar] [CrossRef]
- Wang, H.; Sun, T.; Hu, J.; Zhang, R.; Rao, Y.; Wang, S.; Chen, R.; McLendon, R.E.; Friedman, A.H.; Keir, S.T.; et al. miR-33a promotes glioma-initiating cell self-renewal via PKA and NOTCH pathways. J. Clin. Investig. 2014, 124, 4489–4502. [Google Scholar] [CrossRef] [PubMed]
- Guipponi, M.; Scott, H.S.; Kudoh, J.; Kawasaki, K.; Shibuya, K.; Shintani, A.; Asakawa, S.; Chen, H.; Lalioti, M.D.; Rossier, C.; et al. Identification and characterization of a novel cyclic nucleotide phosphodiesterase gene (PDE9A) that maps to 21q22.3: Alternative splicing of mRNA transcripts, genomic structure and sequence. Hum. Genet. 1998, 103, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Dorner-Ciossek, C.; Kroker, K.S.; Rosenbrock, H. Role of PDE9 in Cognition. Adv. Neurobiol. 2017, 17, 231–254. [Google Scholar] [PubMed]
- Fujishige, K.; Kotera, J.; Michibata, H.; Yuasa, K.; Takebayashi, S.; Okumura, K.; Omori, K. Cloning and characterization of a novel human phosphodiesterase that hydrolyzes both cAMP and cGMP (PDE10A). J. Biol. Chem. 1999, 274, 18438–18445. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, V.; Zhong, S.; Lin, H.; Xu, W.; Bradaia, A.; Steidl, E.; Gleyzes, M.; Wadel, K.; Buisson, B.; Padovan-Neto, F.E.; et al. Phosphodiesterase 10A Inhibition Improves Cortico-Basal Ganglia Function in Huntington’s Disease Models. Neuron 2016, 92, 1220–1237. [Google Scholar] [CrossRef] [PubMed]
- Yin, D.; Ogawa, S.; Kawamata, N.; Tunici, P.; Finocchiaro, G.; Eoli, M.; Ruckert, C.; Huynh, T.; Liu, G.; Kato, M.; et al. High-resolution genomic copy number profiling of glioblastoma multiforme by single nucleotide polymorphism DNA microarray. Mol. Cancer Res. 2009, 7, 665–677. [Google Scholar] [CrossRef]
- Peng, T.; Gong, J.; Jin, Y.; Zhou, Y.; Tong, R.; Wei, X.; Bai, L.; Shi, J. Inhibitors of phosphodiesterase as cancer therapeutics. Eur. J. Med. Chem. 2018, 150, 742–756. [Google Scholar] [CrossRef]
- Monteiro, J.P.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. Structure-Bioactivity Relationships of Methylxanthines: Trying to Make Sense of All the Promises and the Drawbacks. Molecules 2016, 21, 974. [Google Scholar] [CrossRef]
- Ashihara, H.; Mizuno, K.; Yokota, T.; Crozier, A. Xanthine Alkaloids: Occurrence, Biosynthesis, and Function in Plants. Prog. Chem. Org. Nat. Prod. 2017, 105, 1–88. [Google Scholar]
- Basnet, R.M.; Zizioli, D.; Guarienti, M.; Finazzi, D.; Memo, M. Methylxanthines induce structural and functional alterations of the cardiac system in zebrafish embryos. BMC Pharmacol. Toxicol. 2017, 18, 72. [Google Scholar] [CrossRef]
- Morfin Maciel, B.M.; Castillo Morfin, B.M. Theophylline, a new look to an old drug. Rev. Alerg. Mex. 2010, 57, 112–122. [Google Scholar] [PubMed]
- Chen, J.F.; Eltzschig, H.K.; Fredholm, B.B. Adenosine receptors as drug targets—What are the challenges? Nat. Rev. Drug Discov. 2013, 12, 265–286. [Google Scholar] [CrossRef]
- Fishman, P.; Bar-Yehuda, S.; Synowitz, M.; Powell, J.D.; Klotz, K.N.; Gessi, S.; Borea, P.A. Adenosine receptors and cancer. In Handbook of Experimental Pharmacology; Springer: Berlin/Heidelberg, Germany, 2009; pp. 399–441. [Google Scholar]
- Torres, A.; Vargas, Y.; Uribe, D.; Jaramillo, C.; Gleisner, A.; Salazar-Onfray, F.; Lopez, M.N.; Melo, R.; Oyarzun, C.; San Martin, R.; et al. Adenosine A3 receptor elicits chemoresistance mediated by multiple resistance-associated protein-1 in human glioblastoma stem-like cells. Oncotarget 2016, 7, 67373–67386. [Google Scholar] [CrossRef]
- Durrant, K.L. Known and hidden sources of caffeine in drug, food, and natural products. J. Am. Pharm. Assoc. 2002, 42, 625–637. [Google Scholar] [CrossRef]
- Jones, A.W. Review of Caffeine-Related Fatalities along with Postmortem Blood Concentrations in 51 Poisoning Deaths. J. Anal. Toxicol. 2017, 41, 167–172. [Google Scholar] [CrossRef] [PubMed]
- Poole, R.; Kennedy, O.J.; Roderick, P.; Fallowfield, J.A.; Hayes, P.C.; Parkes, J. Coffee consumption and health: Umbrella review of meta-analyses of multiple health outcomes. BMJ 2017, 359, j5024. [Google Scholar] [CrossRef]
- Durrant, K.L.; Grosso, G.; Godos, J.; Galvano, F.; Giovannucci, E.L.; Astorino, T.A.; Liu, S.; Alkadhi, K.A.; Addicott, M.; Farah, A.; et al. Coffee, Caffeine, and Health Outcomes: An Umbrella Review. Annu. Rev. Nutr. 2017, 37, 131–156. [Google Scholar]
- Cappelletti, S.; Daria, P.; Sani, G.; Aromatario, M. Caffeine: Cognitive and Physical Performance Enhancer or Psychoactive Drug? Curr. Neuropharmacol. 2015, 13, 71–88. [Google Scholar] [CrossRef]
- Lipton, R.B.; Diener, H.C.; Robbins, M.S.; Garas, S.Y.; Patel, K. Caffeine in the management of patients with headache. J. Headache Pain 2017, 18, 107. [Google Scholar] [CrossRef]
- Temple, J.L.; Hostler, D.; Martin-Gill, C.; Moore, C.G.; Weiss, P.M.; Sequeira, D.J.; Condle, J.P.; Lang, E.S.; Higgins, J.S.; Patterson, P.D. Systematic Review and Meta-analysis of the Effects of Caffeine in Fatigued Shift Workers: Implications for Emergency Medical Services Personnel. Prehosp. Emerg. Care 2018, 22, 37–46. [Google Scholar] [CrossRef]
- Baratloo, A.; Rouhipour, A.; Forouzanfar, M.M.; Safari, S.; Amiri, M.; Negida, A. The Role of Caffeine in Pain Management: A Brief Literature Review. Anesthesiol. Pain Med. 2016, 6, e33193. [Google Scholar] [CrossRef] [PubMed]
- Arnold, A.C.; Ramirez, C.E.; Choi, L.; Okamoto, L.E.; Gamboa, A.; Diedrich, A.; Raj, S.R.; Robertson, D.; Biaggioni, I.; Shibao, C. Combination ergotamine and caffeine improves seated blood pressure and presyncopal symptoms in autonomic failure. Front. Physiol. 2014, 5, 270. [Google Scholar] [CrossRef] [PubMed]
- Szopa, A.; Poleszak, E.; Wyska, E.; Serefko, A.; Wośko, S.; Wlaź, A.; Pieróg, M.; Wróbel, A.; Wlaź, P. Caffeine enhances the antidepressant-like activity of common antidepressant drugs in the forced swim test in mice. Naunyn-Schmiedebergs Arch. Pharmacol. 2016, 389, 211–221. [Google Scholar] [CrossRef]
- Ning, Y.L.; Yang, N.; Chen, X.-Y.; Zhao, Z.A.; Zhang, X.Z.; Chen, X.Y.; Li, P.; Zhao, Y.; Zhou, Y.G. Chronic caffeine exposure attenuates blast-induced memory deficit in mice. Chin. J. Traumatol. 2015, 18, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.T.; Jackson, E.K.; Wisniewski, S.R.; Gillespie, D.G.; Puccio, A.M.; Clark, R.S.; Dixon, C.E.; Kochanek, P.M. Increases in Cerebrospinal Fluid Caffeine Concentration are Associated with Favorable Outcome after Severe Traumatic Brain injury in Humans. J. Cereb. Blood Flow Metab. 2008, 28, 395–401. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Geiger, N.H.; Soliman, M.L.; Hui, L.; Geiger, J.D.; Chen, X. Caffeine, Through Adenosine A3Receptor-Mediated Actions, Suppresses Amyloid-β Protein Precursor Internalization and Amyloid-β Generation. J. Alzheimers Dis. 2015, 47, 73–83. [Google Scholar] [CrossRef]
- Khadrawy, Y.A.; Salem, A.M.; El-Shamy, K.A.; Ahmed, E.K.; Fadl, N.N.; Hosny, E.N. Neuroprotective and Therapeutic Effect of Caffeine on the Rat Model of Parkinson’s Disease Induced by Rotenone. J. Diet. Suppl. 2017, 14, 553–572. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wang, Z.; Jin, Y.; Guo, J. Association between tea and coffee consumption and brain cancer risk: An updated meta-analysis. World J. Surg. Oncol. 2019, 17, 51. [Google Scholar] [CrossRef]
- Jiang, J.; Lan, Y.Q.; Zhang, T.; Yu, M.; Liu, X.Y.; Li, L.H.; Chen, X.P. The in vitro effects of caffeine on viability, cycle cycle profiles, proliferation, and apoptosis of glioblastomas. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 3201–3207. [Google Scholar]
- Sun, F.; Han, D.F.; Cao, B.Q.; Wang, B.; Dong, N.; Jiang, D.H. Caffeine-induced nuclear translocation of FoxO1 triggers Bim-mediated apoptosis in human glioblastoma cells. Tumor Biol. 2016, 37, 3417–3423. [Google Scholar] [CrossRef]
- Chen, J.C.; Hwang, J.H.; Chiu, W.H.; Chan, Y.C. Tetrandrine and Caffeine Modulated Cell Cycle and Increased Glioma Cell Death via Caspase-Dependent and Caspase-Independent Apoptosis Pathways. Nutr. Cancer 2014, 66, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.C.; Hwang, J.H. Effects of caffeine on cell viability and activity of histone deacetylase 1 and histone acetyltransferase in glioma cells. Tzu Chi Med. J. 2016, 28, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.S.; Han, K.S.; Ku, B.M.; Lee, Y.K.; Hong, J.; Shin, H.Y.; Almonte, A.G.; Woo, D.H.; Brat, D.J.; Hwang, E.M.; et al. Caffeine-mediated inhibition of calcium release channel inositol 1,4,5-trisphosphate receptor subtype 3 blocks glioblastoma invasion and extends survival. Cancer Res. 2010, 70, 1173–1183. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.C.; Ding, Y.M.; Hueng, D.Y.; Chen, J.Y.; Chen, Y. Caffeine suppresses the progression of human glioblastoma via cathepsin B and MAPK signaling pathway. J. Nutr. Biochem. 2016, 33, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, G.; D’Amico, A.G.; Rasà, D.M.; Saccone, S.; Federico, C.; Magro, G.; Cavallaro, S.; D’Agata, V. Caffeine Effect on HIFs/VEGF Pathway in Human Glioblastoma Cells Exposed to Hypoxia. Anti-Cancer Agents Med. Chem. 2018, 18, 1432–1439. [Google Scholar] [CrossRef]
- Conde, V.R.; Alves, M.G.; Oliveira, P.F.; Silva, B.M. Tea (Camellia sinensis (L.)): A putative anticancer agent in bladder carcinoma? Anti-Cancer Agents Med. Chem. 2015, 15, 26–36. [Google Scholar] [CrossRef]
- Onatibia-Astibia, A.; Franco, R.; Martinez-Pinilla, E. Health benefits of methylxanthines in neurodegenerative diseases. Mol. Nutr. Food Res. 2017, 61, 1600670. [Google Scholar] [CrossRef]
- Barnes, P.J. Theophylline in chronic obstructive pulmonary disease: New horizons. Proc. Am. Thorac. Soc. 2005, 2, 334–339. [Google Scholar] [CrossRef]
- Barnes, P.J. Theophylline. Pharmaceuticals 2010, 3, 725–747. [Google Scholar] [CrossRef]
- Koul, D.; Wang, S.; Wu, S.; Saito, N.; Zheng, S.; Gao, F.; Kaul, I.; Setoguchi, M.; Nakayama, K.; Koyama, K.; et al. Preclinical therapeutic efficacy of a novel blood-brain barrier-penetrant dual PI3K/mTOR inhibitor with preferential response in PI3K/PTEN mutant glioma. Oncotarget 2017, 8, 21741–21753. [Google Scholar] [CrossRef]
- Gallelli, L.; Falcone, D.; Cannataro, R.; Perri, M.; Serra, R.; Pelaia, G.; Maselli, R.; Savino, R.; Spaziano, G.; D’Agostino, B. Theophylline action on primary human bronchial epithelial cells under proinflammatory stimuli and steroidal drugs: A therapeutic rationale approach. Drug Des. Dev. Ther. 2017, 11, 265–272. [Google Scholar] [CrossRef] [PubMed]
- Domvri, K.; Zarogoulidis, K.; Zogas, N.; Zarogoulidis, P.; Petanidis, S.; Porpodis, K.; Kioseoglou, E.; Hohenforst-Schmidt, W. Potential synergistic effect of phosphodiesterase inhibitors with chemotherapy in lung cancer. J. Cancer 2017, 8, 3648–3656. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.L.; Hsu, Y.J.; Chen, Y.; Wang, Y.W.; Huang, S.M. Theophylline exhibits anti-cancer activity via suppressing SRSF3 in cervical and breast cancer cell lines. Oncotarget 2017, 8, 101461–101474. [Google Scholar] [CrossRef] [PubMed]
- Rusconi, M.; Conti, A. Theobroma cacao L., the Food of the Gods: A scientific approach beyond myths and claims. Pharmacol. Res. 2010, 61, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Kreider, J.W.; Rosenthal, M.; Lengle, N. Cyclic adenosine 3′,5′-monophosphate in the control of melanoma cell replication and differentiation. J. Natl. Cancer Inst. 1973, 50, 555–558. [Google Scholar] [CrossRef] [PubMed]
- Pozner, J.; Papatestas, A.E.; Fagerstrom, R.; Schwartz, I.; Saevitz, J.; Feinberg, M.; Aufses, A.H., Jr. Association of tumor differentiation with caffeine and coffee intake in women with breast cancer. Surgery 1986, 100, 482–488. [Google Scholar] [CrossRef] [PubMed]
- Tomita, K.; Tsuchiya, H. Enhancement of cytocidal and antitumor effect of cisplatin by caffeine in human osteosarcoma. Clin. Ther. 1989, 11, 43–52. [Google Scholar] [PubMed]


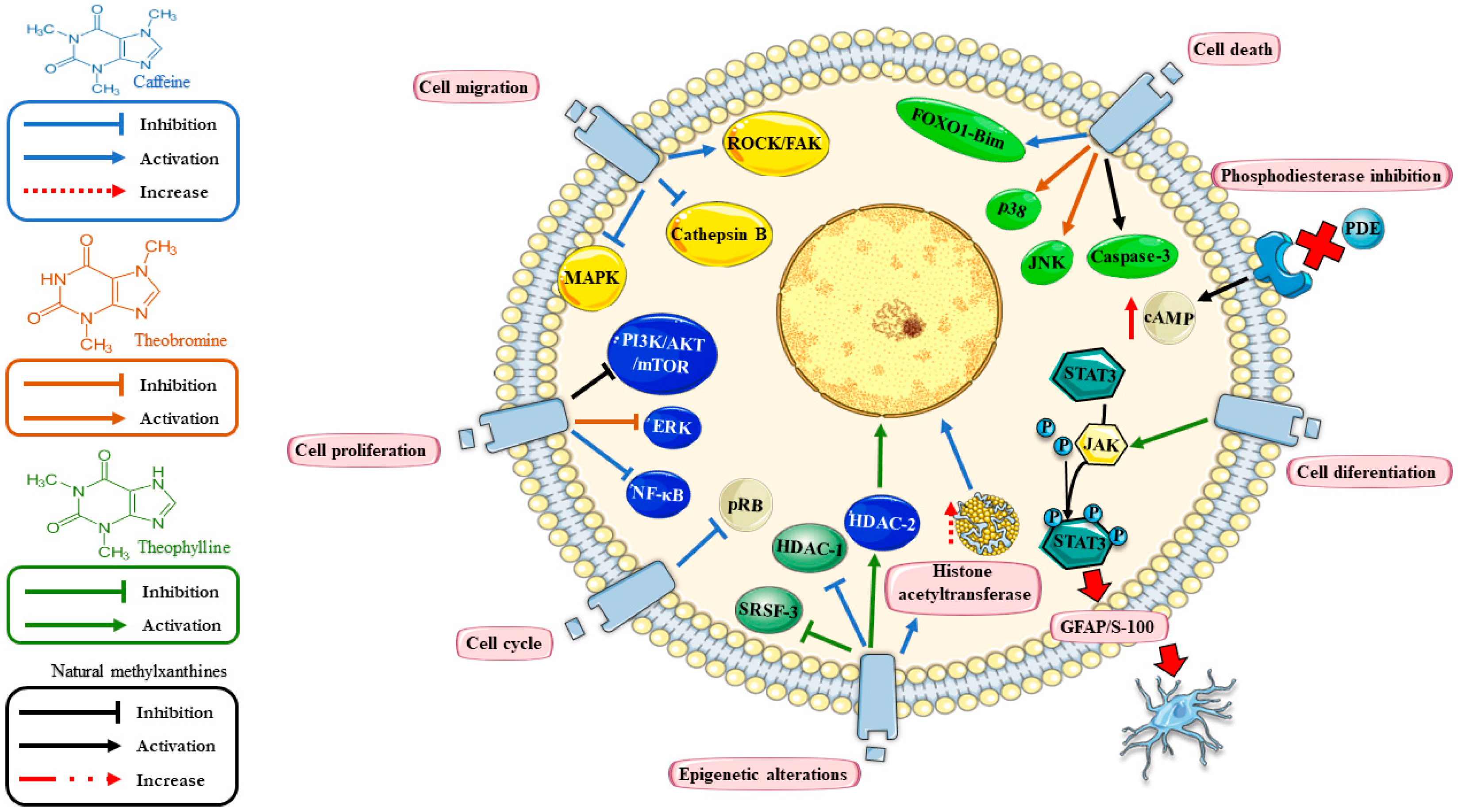{kind=link}
| Main Author (Reference) | Methylxanthine | Type of Study | Relevant Methodology | Relevant Results |
|---|---|---|---|---|
| Moon et al., 2012 [14] | Theophylline | In vitro | A-172 and U87MG cell lines | Reduces the survival and proliferation |
| Nagai et al., 1971 [15] | Theophylline | In vitro | Human glioblastoma cells and glioma cells induced by MC in C57 black mouse | Induces morphological changes |
| Sato et al., 1975 [16] | Theophylline | In vitro | Mouse glioma cell line | Induces glial-like morphological changes and expression of S-100 protein. |
| Takanaga et al., 2004 [17] | Theophylline | In vitro | C6 cell line | N6,2′-O-dibutyryl cAMP (Bt2AMP) and theophylline caused delayed phosphorylation of STAT3 and expression of GFAP. |
| Sugimoto et al., 2014 [18] | Theobromine | In vitro | U87MG cell line | Anti-tumoral and anti-inflammatory effects. Inhibits proliferation and induces apoptosis. |
| Stewart et al., 1987 [19] | Caffeine | Clinical | 25 patients with gliomas Caffeine added to cytosine arabinoside plus cisplatin | Presence of caffeine-induced seizures 48% of the patients responded. |
| Janss et al., 1998 [20] | Caffeine | In vitro | U251 glioma cells | Caffeine reduced the ID50 and ID90 of cisplatin promoting apoptosis. |
| Chen et al., 2014 [21] | Caffeine | In vitro | C6 and U87MG cell lines | Caffeine decreases migration by inhibition of ROCK-focal adhesion complex pathway. |
| Sinn et al., 2010 [22] | Caffeine | In vitro | U87MG, T98G and U373MG cells lines | Inhibits PI3K, downregulating the PI3K/Akt pathway and induces apoptosis. |
| Ku et al., 2011 [23] | Caffeine | In vivo | Mouse xenograft model of GBM | Inhibits of the IP3R3. |
| Phosphodiesterase | Gene Chromosome | Substrate | Main Function | Participation in GBM | Ref. |
|---|---|---|---|---|---|
| PDE1 | PDE1A, B, C 2q32.1, 12q13.2, 7q14.3 | cAMP and cGMP | Promotes cell proliferation and migration | PDE1C is overexpressed on GBM | [29,30,31,32] |
| PDE2 | PDE2A 11q13.4 | cAMP and cGMP | Regulates endothelial permeability and proliferation and nNOS expression. | PDE2A is overexpressed in low grade glioma | [33,34] |
| PDE3 | PDE3A, B 12q12.2, 11p15.2 | cAMP and cGMP | Smooth muscle contraction, insulin signaling, blood vessel formation, and antiapoptotic and anti-inflammatory pathways | N. D. | [35,36,37] |
| PDE4 | PDE4A, B, C, D 19p13.2, 1p31.3, 19p13.11, 5p11.2-q12.1 | cAMP | Promotes blood vessel formation, monocyte and macrophage activation, and antiapoptotic and anti-inflammatory pathways | PDE4 promotes the tumor growth Hypermethylation of the PDE4C promoter is associated with high malignant grade and reduced overall survival | [36,38,39,40,41] |
| PDE5 | PDE5A 4q26 | cGMP | Regulates cell signaling | PDE5 is overexpression correlates with longer overall survival, and its inhibition induces an invasive phenotype of GBM | [7,42] |
| PDE6 | PDE6A, B, C 5q32, 4p16.3, 10q24 | cGMP | Participates in rod and cone photoreceptor function | N. D. | [43,44,45] |
| PDE7 | PDE7A, B 8q13, 6q23-24 | cAMP | Modulation of T-cell proliferation | PDE7B overexpression induces tumor growth | [46,47,48,49] |
| PDE8 | PDE8A, B 15q25.3, 15q13.3 | cAMP | Controls T cells and breast cancer cells motility | PDE8A expression correlates with an increased overall survival | [50,51,52,53] |
| PDE9 | PDE9A 21q22.3 | cGMP | Participates in synaptic plasticity and cognitive function | N. D. | [54,55] |
| PDE10 | PDE10A 6q26 | cAMP and cGMP | Regulates intracellular signaling and controls striatal gene expression | PDE10A is deleted on GBM tissue | [56,57,58] |
| PDE11 | PDE11A 2q31.2 | cAMP and cGMP | Contributes to sperm development | N. D. | [59] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Pérez, D.; Reyes-Vidal, I.; Chávez-Cortez, E.G.; Sotelo, J.; Magaña-Maldonado, R. Methylxanthines: Potential Therapeutic Agents for Glioblastoma. Pharmaceuticals 2019, 12, 130. https://doi.org/10.3390/ph12030130
Pérez-Pérez D, Reyes-Vidal I, Chávez-Cortez EG, Sotelo J, Magaña-Maldonado R. Methylxanthines: Potential Therapeutic Agents for Glioblastoma. Pharmaceuticals. 2019; 12(3):130. https://doi.org/10.3390/ph12030130
Chicago/Turabian StylePérez-Pérez, Daniel, Iannel Reyes-Vidal, Elda Georgina Chávez-Cortez, Julio Sotelo, and Roxana Magaña-Maldonado. 2019. "Methylxanthines: Potential Therapeutic Agents for Glioblastoma" Pharmaceuticals 12, no. 3: 130. https://doi.org/10.3390/ph12030130
APA StylePérez-Pérez, D., Reyes-Vidal, I., Chávez-Cortez, E. G., Sotelo, J., & Magaña-Maldonado, R. (2019). Methylxanthines: Potential Therapeutic Agents for Glioblastoma. Pharmaceuticals, 12(3), 130. https://doi.org/10.3390/ph12030130