Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth


Abstract
1. Introduction
2. Phage Isolation Methods
2.1. Overview of Isolation Protocols
2.2. Environmental Samples
2.3. Sample Processing
2.4. Choice of Isolation Host(s) and Enrichment Culture Variations
2.5. A Further Note about Hosts
2.6. Post-Infection Processing
3. Detection and Characterization of Phages for Phage Therapy
3.1. A Note about Cocktails
3.2. Desirable Characteristics of Phages for Phage Therapy
- Ability to clear a culture of target bacteria or some other measure of phage virulence
- Obligately lytic growth or lack of lysogenic potential
- Transduction potential
- Screening for toxin genes
- Host range
3.3. Phage Virulence (Culture Clearing)
3.4. Obligately Lytic Growth and Gene Screening
- Many temperate phages are capable of genetic transduction which can carry genes from one bacterium to another with the potential of increasing virulence of the recipient bacterium. As well, some species of bacteria use transducing phages to move pathogenicity islands into strains lacking them at even higher efficiency than random transduction by including packaging signals on the pathogenicity islands [128,129,130].
3.5. Host Range
3.6. Other Properties
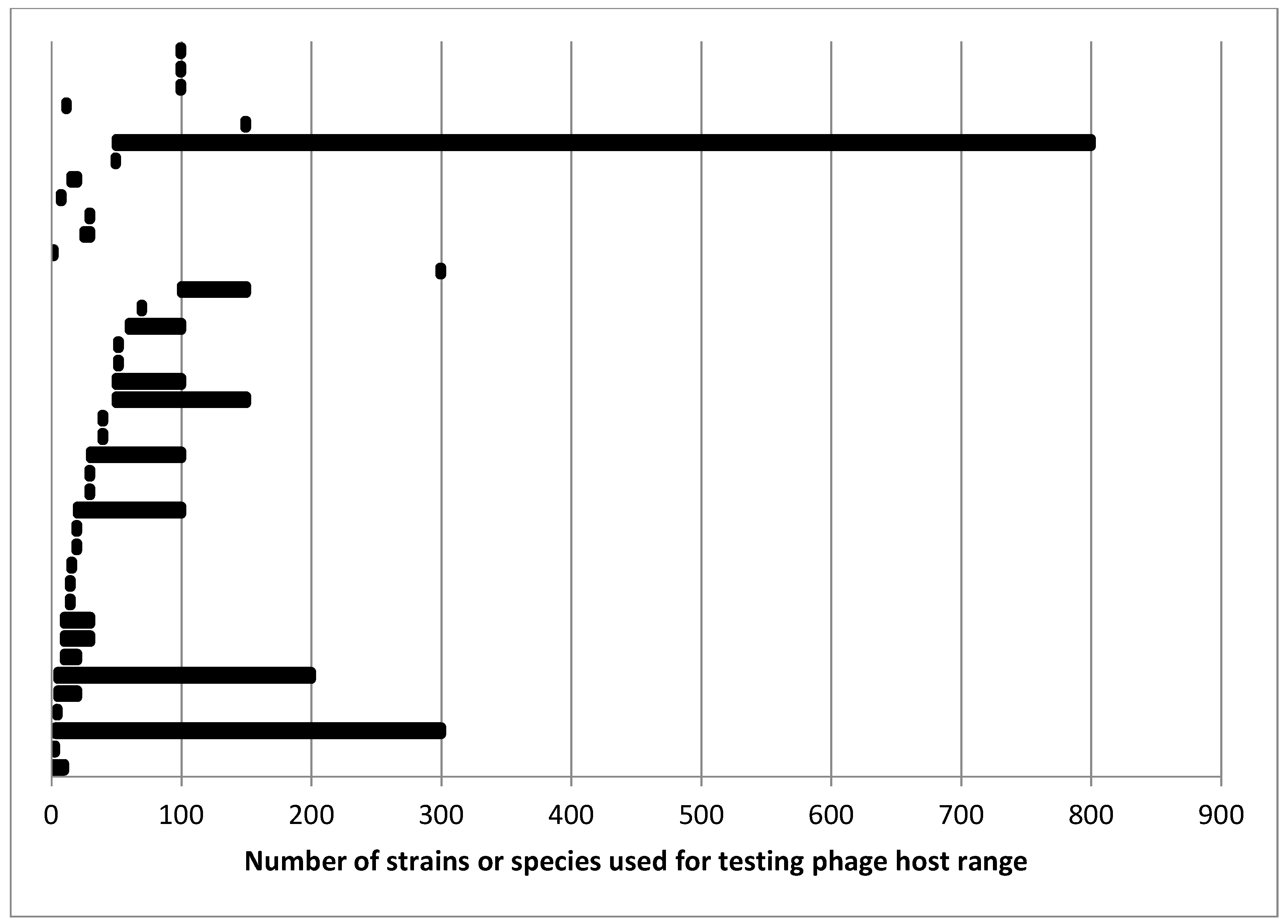
4. Current Practices in Host Range Testing
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Sillankorva, S. Isolation of Bacteriophages for Clinically Relevant Bacteria. In Bacteriophage Therapy: From Lab to Clinical Practice; Azeredo, J., Sillankorva, S., Eds.; Humana Press: New York, NY, USA, 2017; pp. 23–30. [Google Scholar]
- Carvalho, C.; Susano, M.; Fernandes, E.; Santos, S.; Gannon, B.; Nicolau, A.; Gibbs, P.; Teixeira, P.; Azeredo, J. Method for bacteriophage isolation against target Campylobacter strains. Lett. Appl. Microbiol. 2010, 50, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Van Twest, R.; Kropinski, A.M. Bacteriophage enrichment from water and soil. Methods Mol. Biol. 2009, 501, 15–21. [Google Scholar] [PubMed]
- Sulakvelidze, A.; Alavidze, Z.; Morris, J.G., Jr. Bacteriophage therapy. Antimicrob. Agents Chemother. 2001, 45, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Leibovici, L.; Shraga, I.; Drucker, M.; Konigsberger, H.; Samra, Z.; Pitlik, S.D. The benefit of appropriate empirical antibiotic treatment in patients with bloodstream infection. J. Intern. Med. 1998, 244, 379–386. [Google Scholar] [CrossRef] [PubMed]
- Ross, A.; Ward, S.; Hyman, P. More Is Better: Selecting for Broad Host Range Bacteriophages. Front. Microbiol. 2016, 7, 1352. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.J.; Hyman, P. Phage choice, isolation, and preparation for phage therapy. Curr. Pharm. Biotechnol. 2010, 11, 2–14. [Google Scholar] [CrossRef] [PubMed]
- Lobocka, M.; Hejnowicz, M.S.; Gagala, U.; Weber-Dabrowska, B.; Wegrzyn, G.; Dadlez, M. The First Step to Bacteriophage Therapy: How to Choose the Correct Phage. In Phage Therapy: Current Research and Applications; Borysowski, J., Miedzybrodzki, R., Gorski, A., Eds.; Caister Academic Press: Norfalk, UK, 2018; pp. 23–67. [Google Scholar]
- Melo, L.D.; Sillankorva, S.; Ackermann, H.W.; Kropinski, A.M.; Azeredo, J.; Cerca, N. Isolation and characterization of a new Staphylococcus epidermidis broad-spectrum bacteriophage. J. Gen. Virol. 2014, 95, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, A.; Sillankorva, S.; Quinta, R.; Henriques, A.; Sereno, R.; Azeredo, J. Isolation and characterization of bacteriophages for avian pathogenic, E. coli strains. J. Appl. Microbiol. 2009, 106, 1919–1927. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T.; Garcia, P.; Mullany, P.; Aminov, R. Editorial: Phage Therapy: Past, Present and Future. Front. Microbiol. 2017, 8, 981. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.W.; Audurier, A.; Berthiaume, L.; Jones, L.A.; Mayo, J.A.; Vidaver, A.K. Guidelines for bacteriophage characterization. Adv. Virus Res. 1978, 23, 1–24. [Google Scholar] [PubMed]
- Gencay, Y.E.; Birk, T.; Sorensen, M.C.; Brondsted, L. Methods for Isolation, Purification, and Propagation of Bacteriophages of Campylobacter jejuni. Methods Mol. Biol. 2017, 1512, 19–28. [Google Scholar] [PubMed]
- Bhunchoth, A.; Phironrit, N.; Leksomboon, C.; Chatchawankanphanich, O.; Kotera, S.; Narulita, E.; Kawasaki, T.; Fujie, M.; Yamada, T. Isolation of Ralstonia solanacearum-infecting bacteriophages from tomato fields in Chiang Mai, Thailand, and their experimental use as biocontrol agents. J. Appl. Microbiol. 2015, 118, 1023–1033. [Google Scholar] [CrossRef] [PubMed]
- Chibani-Chennoufi, S.; Sidoti, J.; Bruttin, A.; Kutter, E.; Sarker, S.; Brussow, H. In vitro and in vivo bacteriolytic activities of Escherichia coli phages: Implications for phage therapy. Antimicrob. Agents Chemother. 2004, 48, 2558–2569. [Google Scholar] [CrossRef] [PubMed]
- Debartolomeis, J.; Cabelli, V.J. Evaluation of an Escherichi coli host strain for enumeration of F male-specific bacteriophages. Appl. Environ. Microbiol. 1991, 57, 1301–1305. [Google Scholar]
- Bachrach, G.; Leizerovici-Zigmond, M.; Zlotkin, A.; Naor, R.; Steinberg, D. Bacteriophage isolation from human saliva. Lett. Appl. Microbiol. 2003, 36, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Tylenda, C.A.; Calvert, C.; Kolenbrander, P.E.; Tylenda, A. Isolation of Actinomyces bacteriophage from human dental plaque. Infect. Immunity 1985, 49, 1–6. [Google Scholar]
- Serwer, P.; Hayes, S.J.; Zaman, S.; Lieman, K.; Rolando, M.; Hardies, S.C. Improved isolation of undersampled bacteriophages: Finding of distant terminase genes. Virology 2004, 329, 412–424. [Google Scholar] [CrossRef] [PubMed]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Demeler, B.; Hardies, S.C. Isolation of novel large and aggregating bacteriophages. Methods Mol. Biol. 2009, 501, 55–66. [Google Scholar] [PubMed]
- Serwer, P. Restoring logic and data to phage-cures for infectious disease. AIMS Microbiol. 2017, 3, 706–712. [Google Scholar] [CrossRef]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Hardies, S.C. Propagating the missing bacteriophages: A large bacteriophage in a new class. Virol. J. 2007, 4, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Serwer, P.; Wright, E.T.; Chang, J.T.; Liu, X. Enhancing and initiating phage-based therapies. Bacteriophage 2014, 4, e961869. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, M.; Liu, Q.; Song, X.; Wang, D.; Ma, Y.; Shao, H.; Jiang, Y. Complete Genomic Sequence of Bacteriophage H188: A Novel Vibrio kanaloae Phage Isolated from Yellow Sea. Curr. Microbiol. 2016, 72, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, M.B.; Waterbury, J.B.; Chisholm, S.W. Cyanophages infecting the oceanic cyanobacterium Prochlorococcus. Nature 2003, 424, 1047–1051. [Google Scholar] [CrossRef] [PubMed]
- Stenholm, A.R.; Dalsgaard, I.; Middelboe, M. Isolation and characterization of bacteriophages infecting the fish pathogen Flavobacterium psychrophilum. Appl. Environ. Microbiol. 2008, 74, 4070–4078. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.P.; Gong, T.; Jost, G.; Liu, W.H.; Ye, D.Z.; Luo, Z.H. Isolation and characterization of five lytic bacteriophages infecting a Vibrio strain closely related to Vibrio owensii. FEMS Microbiol. Lett. 2013, 348, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Jakhetia, R.; Talukder, K.A.; Verma, N.K. Isolation, characterization and comparative genomics of bacteriophage SfIV: A novel serotype converting phage from Shigella flexneri. BMC. Genom. 2013, 14, 677. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Wei, X.; Ma, Y.; Yin, Z.; Li, H.; Lin, W.; Wang, X.; Li, C.; Shen, Z.; Zhao, R.; et al. Isolation and characterization of a bacteriophage phiEap-2 infecting multidrug resistant Enterobacter aerogenes. Sci. Rep. 2016, 6, 28338. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.M.; Nilsson, A.S. Isolation of phages for phage therapy: A comparison of spot tests and efficiency of plating analyses for determination of host range and efficacy. PLoS ONE 2015, 10, e0118557. [Google Scholar]
- Karumidze, N.; Kusradze, I.; Rigvava, S.; Goderdzishvili, M.; Rajakumar, K.; Alavidze, Z. Isolation and characterisation of lytic bacteriophages of Klebsiella pneumoniae and Klebsiella oxytoca. Curr. Microbiol. 2013, 66, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Jensen, E.C.; Schrader, H.S.; Rieland, B.; Thompson, T.L.; Lee, K.W.; Nickerson, K.W.; Kokjohn, T.A. Prevalence of broad-host-range lytic bacteriophages of Sphaerotilus natans, Escherichia coli, and Pseudomonas aeruginosa. Appl. Environ. Microbiol. 1998, 64, 575–580. [Google Scholar] [PubMed]
- Liu, J.; Yan, R.; Zhong, Q.; Ngo, S.; Bangayan, N.J.; Nguyen, L.; Lui, T.; Liu, M.; Erfe, M.C.; Craft, N.; et al. The diversity and host interactions of Propionibacterium acnes bacteriophages on human skin. ISME. J. 2015, 9, 2078–2093. [Google Scholar] [CrossRef] [PubMed]
- Farrar, M.D.; Howson, K.M.; Bojar, R.A.; West, D.; Towler, J.C.; Parry, J.; Pelton, K.; Holland, K.T. Genome sequence and analysis of a Propionibacterium acnes bacteriophage. J. Bacteriol. 2007, 189, 4161–4167. [Google Scholar] [CrossRef] [PubMed]
- Ronda, C.; Lopez, R.; Garcia, E. Isolation and characterization of a new bacteriophage, Cp-1, infecting Streptococcus pneumoniae. J. Virol. 1981, 40, 551–559. [Google Scholar] [PubMed]
- Rasool, M.H.; Yousaf, R.; Siddique, A.B.; Saqalein, M.; Khurshid, M. Isolation, Characterization, and Antibacterial Activity of Bacteriophages Against Methicillin-Resistant Staphylococcus aureus in Pakistan. Jundishapur. J. Microbiol. 2016, 9, e36135. [Google Scholar] [CrossRef] [PubMed]
- Nikkhahi, F.; Soltan Dallal, M.M.; Alimohammadi, M.; Rahimi, F.A.; Rajabi, Z.; Fardsanei, F.; Imeni, S.M.; Torabi, B.P. Phage therapy: Assessment of the efficacy of a bacteriophage isolated in the treatment of salmonellosis induced by Salmonella enteritidis in mice. Gastroenterol. Hepatol. Bed. Bench. 2017, 10, 131–136. [Google Scholar] [PubMed]
- Ackermann, H.W.; Caprioli, T.; Kasatiya, S.S. A large new Streptococcus bacteriophage. Can. J. Microbiol. 1975, 21, 571–574. [Google Scholar] [CrossRef] [PubMed]
- Mattila, S.; Ruotsalainen, P.; Jalasvuori, M. On-Demand Isolation of Bacteriophages Against Drug-Resistant Bacteria for Personalized Phage Therapy. Front. Microbiol. 2015, 6, 1271. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, K.R.; Otieno, J.R.; Thanki, A.; Blades, M.J.; Millard, A.D.; Browne, H.P.; Lawley, T.D.; Clokie, M.R. As Clear as Mud? Determining the Diversity and Prevalence of Prophages in the Draft Genomes of Estuarine Isolates of Clostridium difficile. Genome Biol. Evol. 2015, 7, 1842–1855. [Google Scholar] [CrossRef] [PubMed]
- Suttle, C.A.; Chan, A.M.; Cottrell, M.T. Use of ultrafiltration to isolate viruses from seawater which are pathogens of marine phytoplankton. Appl. Environ. Microbiol. 1991, 57, 721–726. [Google Scholar] [PubMed]
- Millard, A.D. Isolation of cyanophages from aquatic environments. Methods Mol. Biol. 2009, 501, 33–42. [Google Scholar] [PubMed]
- Czajkowski, R.; Ozymko, Z.; Lojkowska, E. Application of zinc chloride precipitation method for rapid isolation and concentration of infectious Pectobacterium spp. and Dickeya spp. lytic bacteriophages from surface water and plant and soil extracts. Folia Microbiol. 2016, 61, 29–33. [Google Scholar] [CrossRef] [PubMed]
- Poulos, B.T.; John, S.G.; Sullivan, M.B. Iron Chloride Flocculation of Bacteriophages from Seawater. Methods Mol. Biol. 2018, 1681, 49–57. [Google Scholar] [PubMed]
- Betz, J.V.; Anderson, K.E. Isolation and characterization of bacteriophages active on clostridium sporogenes. J. Bacteriol. 1964, 87, 408–415. [Google Scholar] [PubMed]
- Dafale, N.; Lakhe, S.; Yadav, K.; Purohit, H.; Chakrabarti, T. Concentration and recovery of coliphages from water with bituminous coal. Water Environ. Res. 2008, 80, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Trubl, G.; Solonenko, N.; Chittick, L.; Solonenko, S.A.; Rich, V.I.; Sullivan, M.B. Optimization of viral resuspension methods for carbon-rich soils along a permafrost thaw gradient. PeerJ. 2016, 4, e1999. [Google Scholar] [CrossRef] [PubMed]
- Williamson, K.; Corzo, K.; Drissi, C.; Buckingham, J.; Thompson, C.; Helton, R. Estimates of viral abundance in soils are strongly influenced by extraction and enumeration methods. Biol. Fertil. Soils 2013, 49, 857–869. [Google Scholar] [CrossRef]
- Piracha, Z.; Saeed, U.; Khurshid, A.; Chaudhary, W. Isolation and Partial Characterization of Virulent Phage Specific against Pseudomonas Aeruginosa. Glob. J. Med. Res. C Microbi. Pathol. 2014, 14, 1–9. [Google Scholar]
- Rombouts, S.; Volckaert, A.; Venneman, S.; Declercq, B.; Vandenheuvel, D.; Allonsius, C.N.; Jang, H.B.; Briers, Y.; Noben, J.P.; Klumpp, J.; et al. Characterization of Novel Bacteriophages for Biocontrol of Bacterial Blight in Leek Caused by Pseudomonas syringae pv. porri. Front. Microbiol. 2016, 7, 279. [Google Scholar] [CrossRef] [PubMed]
- Lone, A.; Anany, H.; Hakeem, M.; Aguis, L.; Avdjian, A.-C.; Bouget, M.; Atashi, A.; Brovko, L.; Rochefort, D.; Griffiths, M.W. Development of prototypes of bioactive packaging materials based on immobilized bacteriophages for control of growth of bacterial pathogens in foods. Int. J. Food Microbiol. 2016, 217, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Anany, H.; Chen, W.; Pelton, R.; Griffiths, M.W. Biocontrol of Listeria monocytogenes and Escherichia coli O157:H7 in meat by using phages immobilized on modified cellulose membranes. Appl. Environ. Microbiol. 2011, 77, 6379–6387. [Google Scholar] [CrossRef] [PubMed]
- Tolba, M.; Minikh, O.; Brovko, L.Y.; Evoy, S.; Griffiths, M.W. Oriented immobilization of bacteriophages for biosensor applications. Appl. Environ. Microbiol. 2010, 76, 528–535. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.; Abedon, S. Bacteriophage: Overview. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Filippov, A.A.; Sergueev, K.V.; He, Y.; Huang, X.Z.; Gnade, B.T.; Mueller, A.J.; Fernandez-Prada, C.M.; Nikolich, M.P. Bacteriophage-resistant mutants in Yersinia pestis: Identification of phage receptors and attenuation for mice. PLoS ONE 2011, 6, e25486. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.L.; Hsieh, P.F.; Huang, Y.T.; Lee, W.C.; Tsai, Y.T.; Su, P.A.; Pan, Y.J.; Hsu, C.R.; Wu, M.C.; Wang, J.T. Isolation of a bacteriophage and its depolymerase specific for K1 capsule of Klebsiella pneumoniae: Implication in typing and treatment. J. Infect. Dis. 2014, 210, 1734–1744. [Google Scholar] [CrossRef] [PubMed]
- Chan, B.K.; Sistrom, M.; Wertz, J.E.; Kortright, K.E.; Narayan, D.; Turner, P.E. Phage selection restores antibiotic sensitivity in MDR Pseudomonas aeruginosa. Sci. Rep. 2016, 6, 26717. [Google Scholar] [CrossRef] [PubMed]
- Hatfull, G.F. The secret lives of mycobacteriophages. Adv. Virus Res. 2012, 82, 179–288. [Google Scholar] [PubMed]
- Kumar, G.P.N.; Sundarrajan, S.; Paul, V.D.; Nandini, S.; Saravanan, R.S.; Hariharan, S.; Sriram, B.; Padmanabhan, S. Use of prophage free host for achieving homogenous population of bacteriophages: New findings. Virus Res. 2012, 169, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Adams, M.H. Bacteriophages; InterScience: New York, NY, USA, 1959. [Google Scholar]
- Raya, R.R.; Hebert, E.M. Isolation of phage via induction of lysogens. Methods Mol. Biol. 2009, 501, 23–32. [Google Scholar] [PubMed]
- Hurwitz, B.L.; Ponsero, A.; Thornton, J., Jr.; U’Ren, J.M. Phage hunters: Computational strategies for finding phages in large-scale omics datasets. Virus Res. 2018, 244, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed]
- Akhter, S.; Aziz, R.K.; Edwards, R.A. PhiSpy: A novel algorithm for finding prophages in bacterial genomes that combines similarity- and composition-based strategies. Nucleic Acids Res. 2012, 40, e126. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M. Phage Therapy—Everything Old is New Again. Can. J. Infect. Dis. Med. Microbiol. 2006, 17, 297–306. [Google Scholar] [CrossRef] [PubMed]
- Ghugare, G.S.; Nair, A.; Nimkande, V.; Sarode, P.; Rangari, P.; Khairnar, K. Membrane filtration immobilization technique—A simple and novel method for primary isolation and enrichment of bacteriophages. J. Appl. Microbiol. 2017, 122, 531–539. [Google Scholar] [CrossRef] [PubMed]
- Sillankorva, S.; Pleteneva, E.; Shaburova, O.; Santos, S.; Carvalho, C.; Azeredo, J.; Krylov, V. Salmonella Enteritidis bacteriophage candidates for phage therapy of poultry. J. Appl. Microbiol. 2010, 108, 1175–1186. [Google Scholar] [CrossRef] [PubMed]
- Yu, P.; Mathieu, J.; Li, M.; Dai, Z.; Alvarez, P.J. Isolation of Polyvalent Bacteriophages by Sequential Multiple-Host Approaches. Appl. Environ. Microbiol. 2016, 82, 808–815. [Google Scholar] [CrossRef] [PubMed]
- Mapes, A.C.; Trautner, B.W.; Liao, K.S.; Ramig, R.F. Development of expanded host range phage active on biofilms of multi-drug resistant Pseudomonas aeruginosa. Bacteriophage 2016, 6, e1096995. [Google Scholar] [CrossRef] [PubMed]
- Merabishvili, M.; Pirnay, J.P.; De, V.D. Guidelines to Compose an Ideal Bacteriophage Cocktail. Methods Mol. Biol. 2018, 1693, 99–110. [Google Scholar] [PubMed]
- Agboluaje, M.; Sauvageau, D. Bacteriophage Production in Bioreactors. Methods Mol. Biol. 2018, 1693, 173–193. [Google Scholar] [PubMed]
- Grieco, S.H.; Wong, A.Y.; Dunbar, W.S.; MacGillivray, R.T.; Curtis, S.B. Optimization of fermentation parameters in phage production using response surface methodology. J. Ind. Microbiol. Biotechnol. 2012, 39, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Sechaud, J.; Kellenberger, E. Early lysis produced by chloroform in bacteria infected by bacteriophage. Ann. Inst. Pasteur 1956, 90, 102–106. [Google Scholar]
- Ackermann, H.W. Tailed bacteriophages: The order caudovirales. Adv. Virus Res. 1998, 51, 135–201. [Google Scholar] [PubMed]
- d’Hérelle, F.; Smith, G.H. The Bacteriophage and Its Behavior; The Williams &Wilkins, Co.: Baltimore, MD, USA, 1926. [Google Scholar]
- Oliveira, H.; Pinto, G.; Oliveira, A.; Noben, J.P.; Hendrix, H.; Lavigne, R.; Lobocka, M.; Kropinski, A.M.; Azeredo, J. Characterization and genomic analyses of two newly isolated Morganella phages define distant members among Tevenvirinae and Autographivirinae subfamilies. Sci. Rep. 2017, 7, 46157. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Zhang, J.; Niu, Y.D.; Cui, N.; Ma, Y.; Cao, F.; Jin, L.; Li, Z.; Xu, Y. Isolation and characterization of a “phiKMV-like” bacteriophage and its therapeutic effect on mink hemorrhagic pneumonia. PLoS ONE 2015, 10, e0116571. [Google Scholar] [CrossRef] [PubMed]
- Pallavali, R.R.; Degati, V.L.; Lomada, D.; Reddy, M.C.; Durbaka, V.R.P. Isolation and in vitro evaluation of bacteriophages against MDR-bacterial isolates from septic wound infections. PLoS ONE 2017, 12, e0179245. [Google Scholar] [CrossRef] [PubMed]
- Kesik-Szeloch, A.; Drulis-Kawa, Z.; Weber-Dabrowska, B.; Kassner, J.; Majkowska-Skrobek, G.; Augustyniak, D.; Lusiak-Szelachowska, M.; Zaczek, M.; Gorski, A.; Kropinski, A.M. Characterising the biology of novel lytic bacteriophages infecting multidrug resistant Klebsiella pneumoniae. Virol. J. 2013, 10, 100. [Google Scholar] [CrossRef] [PubMed]
- Anne, J.; Wohlleben, W.; Burkardt, H.J.; Springer, R.; Puhler, A. Morphological and molecular characterization of several actinophages isolated from soil which lyse Streptomyces cattleya or, S. venezuelae. J. Gen. Microbiol. 1984, 130, 2639–2649. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.R. Simple colorimetric microplate test of phage lysis in Salmonella enterica. J. Microbiol. Methods 2007, 69, 394–398. [Google Scholar] [CrossRef] [PubMed]
- Waterbury, J.B.; Valois, F.W. Resistance to co-occurring phages enables marine synechococcus communities to coexist with cyanophages abundant in seawater. Appl. Environ. Microbiol. 1993, 59, 3393–3399. [Google Scholar] [PubMed]
- Thomas, E.L.; Corbel, M.J. Isolation of a phage lytic for several Brucella species following propagation of Tbilisi phage in the presence of mitomycin, C. Arch. Virol. 1977, 54, 259–261. [Google Scholar] [CrossRef] [PubMed]
- Corbel, M.J.; Thomas, E.L. Description of a new phage lytic for several Brucella species. J. Biol. Stand. 1976, 4, 195–201. [Google Scholar] [CrossRef]
- Douglas, J.T.; Elberg, S.S. Isolation of Brucella melitensis phage of broad biotype and species specificity. Infect. Immunity 1976, 14, 306–308. [Google Scholar]
- Skurnik, M.; Strauch, E. Phage therapy: Facts and fiction. Int. J. Med. Microbiol. 2006, 296, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Hamdi, S.; Rousseau, G.M.; Labrie, S.J.; Tremblay, D.M.; Kourda, R.S.; Ben, S.K.; Moineau, S. Characterization of two polyvalent phages infecting Enterobacteriaceae. Sci. Rep. 2017, 7, 40349. [Google Scholar] [CrossRef] [PubMed]
- Kutter, E. Phage host range and efficiency of plating. Methods Mol. Biol. 2009, 501, 141–149. [Google Scholar] [PubMed]
- Cooper, C.J.; Denyer, S.P.; Maillard, J.-Y. Rapid and quantitative automated measurement of bacteriophage activity against cystic fibrosis isolates of Pseudomonas aeruginosa. J. Appl. Microbiol. 2011, 110, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Biswas, B.; Vincent, L.; Mokashi, V.; Schuch, R.; Bishop-Lilly, K.A.; Sozhamannan, S. Development of a high throughput assay for indirectly measuring phage growth using the OmniLog™ system. Bacteriophage 2012, 2, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Estrella, L.A.; Quinones, J.; Henry, M.; Hannah, R.M.; Pope, R.K.; Hamilton, T.; Teneza-mora, N.; Hall, E.; Biswajit, B. Characterization of novel Staphylococcus aureus lytic phage and defining their combinatorial virulence using the OmniLog® system. Bacteriophage 2016, 6, e1219440. [Google Scholar] [CrossRef] [PubMed]
- LaVergne, S.; Hamilton, T.; Biswas, B.; Kumaraswamy, M.; Schooley, R.T.; Wooten, D. Phage therapy for a multidrug-resistant Acinetobacter baumannii craniectomy site infection. Open Forum Inf. Dis. 2018, 5, ofy064. [Google Scholar] [CrossRef] [PubMed]
- Schooley, R.T.; Biswas, B.; Gill, J.J.; Hernandez-Morales, A.; Lancaster, J.; Lessor, L.; Barr, J.J.; Reed, S.L.; Rohwer, F.; Benler, S.; et al. Development and use of personalized bacteriophage-based therapeutic cocktails to treat a patient with a disseminated resistant Acinetobacter baumannii infection. Antimicrob. Agents Chemother. 2017, 61, e00954-17. [Google Scholar] [CrossRef] [PubMed]
- Serwer, P.; Hayes, S.J.; Thomas, J.A.; Griess, G.A.; Hardies, S.C. Rapid determination of genomic DNA length for new bacteriophages. Electrophoresis 2007, 28, 1896–1902. [Google Scholar] [CrossRef] [PubMed]
- Quiroz, E.; Recalde, J.; Arias, M.; Seqqat, R.; Vinueza, C.; Ayala, L. Isolation of Lytic Bacteriophages for Nanobiocontrol of Pathogenic and Antibiotic Resistant Salmonella Present in Poultry in Ecuador. Biol. Med. 2016, 8, 287–292. [Google Scholar] [CrossRef]
- Chan, B.K.; Abedon, S.T. Phage therapy pharmacology phage cocktails. Adv. Appl. Microbiol. 2012, 78, 1–23. [Google Scholar] [PubMed]
- Kutter, E.; Borysowski, J.; Miedzybrodzki, R.; Gorski, A.; Weber-Dabrowska, B.; Kutateladze, M.; Alavidze, Z.; Goderdzishvili, M.; Adamia, R. Clinical Phage Therapy. In Phage Therapy: Current Research and Applications; Borysowski, J., Miedzybrodzki, R., Gorski, A., Eds.; Caister Academic Press: Norfalk, UK, 2018; pp. 257–288. [Google Scholar]
- Zschach, H.; Joensen, K.G.; Lindhard, B.; Lund, O.; Goderdzishvili, M.; Chkonia, I.; Jgenti, G.; Kvatadze, N.; Alavidze, Z.; Kutter, E.M.; et al. What Can We Learn from a Metagenomic Analysis of a Georgian Bacteriophage Cocktail? Viruses 2015, 7, 6570–6589. [Google Scholar] [CrossRef] [PubMed]
- McCallin, S.; Alam, S.S.; Barretto, C.; Sultana, S.; Berger, B.; Huq, S.; Krause, L.; Bibiloni, R.; Schmitt, B.; Reuteler, G.; et al. Safety analysis of a Russian phage cocktail: From metagenomic analysis to oral application in healthy human subjects. Virology 2013, 443, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Villarroel, J.; Larsen, M.V.; Kilstrup, M.; Nielsen, M. Metagenomic Analysis of Therapeutic PYO Phage Cocktails from 1997 to 2014. Viruses 2017, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- McCallin, S.; Sarker, S.A.; Sultana, S.; Oechslin, F.; Brussow, H. Metagenome analysis of Russian and Georgian Pyophage cocktails and a placebo-controlled safety trial of single phage versus phage cocktail in healthy Staphylococcus aureus carriers. Environ. Microbiol. 2018, 20, 3278–3293. [Google Scholar] [CrossRef] [PubMed]
- Hagens, S.; Loessner, M.J. Bacteriophage for biocontrol of foodborne pathogens: Calculations and considerations. Curr. Pharm. Biotechnol. 2010, 11, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Ackermann, H.-W.; Tremblay, D.; Moineau, S. Long-term bacteriophage preservation. World Fed. Cult. Coll. Newslett. 2004, 38, 35–40. [Google Scholar]
- Abedon, S. Phage therapy pharmacology: Calculating phage dosing. Adv. Appl. Microbiol. 2011, 77, 1–40. [Google Scholar] [PubMed]
- Abedon, S.T. Kinetics of phage-mediated biocontrol of bacteria. Foodborne Pathog. Dis. 2009, 6, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Levin, B.R.; Bull, J.J. Population and evolutionary dynamics of phage therapy. Nat. Rev. Microbiol. 2004, 2, 166–173. [Google Scholar] [CrossRef] [PubMed]
- Lenski, R.E. Experimental studies of pleiotropy and epistasis in Escherichia coli. I. Variation in competitive fitness among mutants resistant to virus T4. Evolution 1988, 42, 425–432. [Google Scholar] [PubMed]
- Atterbury, R.J.; Van Bergen, M.A.P.; Ortiz, F.; Lovell, M.A.; Harris, J.A.; De Boer, A.; Wagenaar, J.A.; Allen, V.M.; Barrow, P.A. Bacteriophage therapy to reduce Salmonella colonization of broiler chickens. Appl. Environ. Microbiol. 2007, 73, 4543–4549. [Google Scholar] [CrossRef] [PubMed]
- Zahid, M.S.; Udden, S.M.; Faruque, A.S.; Calderwood, S.B.; Mekalanos, J.J.; Faruque, S.M. Effect of phage on the infectivity of Vibrio cholerae and emergence of genetic variants. Infect. Immunity 2008, 76, 5266–5273. [Google Scholar] [CrossRef] [PubMed]
- Hall, A.R.; De Vos, D.; Friman, V.P.; Pirnay, J.P.; Buckling, A. Effects of sequential and simultaneous applications of bacteriophages on populations of Pseudomonas aeruginosa in vitro and in wax moth larvae. Appl. Environ. Microbiol. 2012, 78, 5646–5652. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Vandenheuvel, D.; Martínez, B.; Rodríguez, A.; Lavigne, R.; Garcia, P. Two phages, phiIPLA-RODI and phiIPLA-C1C, lyse mono- and dual-species Staphylococcal biofilms. Appl. Environ. Microbiol. 2015, 81, 3336–3348. [Google Scholar] [CrossRef] [PubMed]
- Ormala, A.M.; Jalasvuori, M. Phage therapy: Should bacterial resistance to phages be a concern, even in the long run? Bacteriophage 2013, 3, e24219. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.Y.K.; Wallin, M.; Lin, Y.; Leung, S.S.Y.; Wang, H.; Morales, S.; Chan, H.K. Phage therapy for respiratory infections. Adv. Drug Deliv. Rev. 2018, 133, 76–86. [Google Scholar] [CrossRef] [PubMed]
- Oechslin, F.; Piccardi, P.; Mancini, S.; Gabard, J.; Moreillon, P.; Entenza, J.M.; Resch, G.; Que, Y.A. Synergistic Interaction Between Phage Therapy and Antibiotics Clears Pseudomonas Aeruginosa Infection in Endocarditis and Reduces Virulence. J. Infect. Dis. 2017, 215, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Bacteriophages as Drugs: The Pharmacology of Phage Therapy. In Phage Therapy: Current Research and Applications; Borysowski, J., Miedzybrodzki, R., Gorski, A., Eds.; Caister Academic Press: Norfalk, UK, 2018; pp. 69–100. [Google Scholar]
- Leung, C.Y.J.; Weitz, J.S. Modeling the synergistic elimination of bacteria by phage and the innate immune system. J. Theor. Biol. 2017, 429, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Roach, D.R.; Leung, C.Y.; Henry, M.; Morello, E.; Singh, D.; Di Santo, J.P.; Weitz, J.S.; Debarbieux, L. Synergy between the host immune system and bacteriophage is essential for successful phage therapy against an acute respiratory pathogen. Cell Host Microbe 2017, 22, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Balogh, B.; Nga, N.T.T.; Jones, J.B. Relative level of bacteriophage multiplication in vitro or in phyllosphere may not predict in planta efficacy for controlling bacterial leaf spot on tomato caused by Xanthomonas perforans. Front. Microbiol. 2018, 9, 2176. [Google Scholar] [CrossRef] [PubMed]
- Shan, J.; Ramachandran, A.; Thanki, A.M.; Vukusic, F.B.I.; Barylski, J.; Clokie, M.R.J. Bacteriophages are more virulent to bacteria with human cells than they are in bacterial culture; insights from HT-29 cells. Sci. Rep. 2018, 8, 5091. [Google Scholar] [CrossRef] [PubMed]
- Olszak, T.; Zarnowiec, P.; Kaca, W.; Danis-Wlodarczyk, K.; Augustyniak, D.; Drevinek, P.; De, S.A.; McClean, S.; Drulis-Kawa, Z. In vitro and in vivo antibacterial activity of environmental bacteriophages against Pseudomonas aeruginosa strains from cystic fibrosis patients. Appl. Microbiol. Biotechnol. 2015, 99, 6021–6033. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Otto, G.; Molineux, I.J. In vivo growth rates are poorly correlated with phage therapy success in a mouse infection model. Antimicrob. Agents Chemother. 2012, 56, 949–954. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Ecology of anti-biofilm agents II: Bacteriophage exploitation and biocontrol of biofilm bacteria. Pharmaceuticals 2015, 8, 559–589. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, D.; Rodríguez-Rubio, L.; Martínez, B.; Rodríguez, A.; García, P. Bacteriophages as weapons against bacterial biofilms in the food industry. Front. Microbiol. 2016, 7, 825. [Google Scholar] [CrossRef] [PubMed]
- Pires, D.P.; Melo, L.D.R.; Vilas Boas, D.; Sillankorva, S.; Azeredo, J. Phage therapy as an alternative or complementary stratgy to prevent and control biofilm-related infections. Curr. Opin. Microbiol. 2017, 39, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Semler, D.D.; Lynch, K.H.; Dennis, J.J. The promise of bacteriophage therapy for Burkholderia cepacia complex respiratory infections. Front. Cell. Infect. Microbiol. 2011, 1, 27. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.; Abedon, S.T. Phage ecology of bacterial pathogenesis. In Bacteriophage Ecology; Abedon, S.T., Ed.; Cambridge University Press: Cambridge, UK, 2008; pp. 353–385. [Google Scholar]
- Schneider, C.L. Bacteriophage-Mediated Horizontal Gene Transfer: Transduction. In Bacteriophages; Harper, D., Abedon, S., Burrowes, B., McConville, M., Eds.; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Novick, R.P.; Ram, G. Staphylococcal pathogenicity islands-movers and shakers in the genomic firmament. Curr. Opin. Microbiol. 2017, 38, 197–204. [Google Scholar] [CrossRef] [PubMed]
- Penades, J.R.; Chen, J.; Quiles-Puchalt, N.; Carpena, N.; Novick, R.P. Bacteriophage-mediated spread of bacterial virulence genes. Curr. Opin. Microbiol. 2015, 23, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Fogg, P.C.; Allison, H.E.; Saunders, J.R.; McCarthy, A.J. Bacteriophage lambda: A paradigm revisited. J. Virol. 2010, 84, 6876–6879. [Google Scholar] [CrossRef] [PubMed]
- Fattah, K.R.; Mizutani, S.; Fattah, F.J.; Matsushiro, A.; Sugino, Y. A comparative study of the immunity region of lambdoid phages including Shiga-toxin-converting phages: Molecular basis for cross immunity. Genes Genet. Syst. 2000, 75, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Lwoff, A. Lysogeny. Bacteriol. Rev. 1953, 17, 269–337. [Google Scholar] [PubMed]
- Jackson, L.E.; Spendlove, R.S. Methods for Isolating Phage and for Controlling Microorganism Populations with the Phage. U.S. Patent 8,821,855, 2 September 2014. [Google Scholar]
- Russell, D.A. Sequencing, Assembling, and Finishing Complete Bacteriophage Genomes. Methods Mol. Biol. 2018, 1681, 109–125. [Google Scholar] [PubMed]
- Hatfull, G.F.; Hendrix, R.W. Bacteriophages and their genomes. Curr. Opin. Virol. 2011, 1, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Fauconnier, A. Guidelines for bacteriophage product certification. Methods Mol. Biol. 2018, 1693, 253–268. [Google Scholar] [PubMed]
- Philipson, C.W.; Voegtly, L.J.; Lueder, M.R.; Long, K.A.; Rice, G.K.; Frey, K.G.; Biswas, B.; Cer, R.Z.; Hamilton, T.; Bishop-Lilly, K.A. Characterizing phage genomes for therapeutic applications. Viruses 2018, 10, 188. [Google Scholar] [CrossRef] [PubMed]
- Lehman, S.M.; Mearns, G.; Rankin, D.; Cole, R.A.; Smrekar, F.; Branston, S.D.; Morales, S. Design and preclinical development of a phage product for the treatment of antibiotic-resistant Staphylococcus aureus infections. Viruses 2019, 11, 88. [Google Scholar] [CrossRef] [PubMed]
- Manoharadas, S.; Blasi, U. Genetically engineered phage as antimicrobials and biodetectors. In Phage Therapy: Current Research and Applications; Borysowski, J., Miedzybrodzki, R., Gorski, A., Eds.; Caister Academic Press: Norfalk, UK, 2018; pp. 343–355. [Google Scholar]
- Costa, A.R.; Milho, C.; Azeredo, J.; Pires, D.P. Synthetic biology to engineer bacteriophage genomes. In Bacteriophage Therapy: From Lab to Clinical Practice; Azeredo, J., Sillankorva, S., Eds.; Humana Press: New York, NY, USA, 2017; pp. 285–300. [Google Scholar]
- Pires, D.P.; Cleto, S.; Sillankorva, S.; Azeredo, J.; Lu, T.K. Genetically Engineered Phages: A Review of Advances over the Last Decade. Microbiol. Mol. Biol. Rev. 2016, 80, 523–543. [Google Scholar] [CrossRef] [PubMed]
- Hyman, P.; Abedon, S.T. Bacteriophage host range and bacterial resistance. Adv. Appl. Microbiol. 2010, 70, 217–248. [Google Scholar] [PubMed]
- Labrie, S.J.; Samson, J.E.; Moineau, S. Bacteriophage resistance mechanisms. Nat. Rev. Microbiol. 2010, 8, 317–327. [Google Scholar] [CrossRef] [PubMed]
- Laanto, E.; Hoikkala, V.; Ravantti, J.; Sundberg, L.R. Long-term genomic coevolution of host-parasite interaction in the natural environment. Nat. Commun. 2017, 8, 111. [Google Scholar] [CrossRef] [PubMed]
- Stern, A.; Sorek, R. The phage-host arms race: Shaping the evolution of microbes. Bioessays 2011, 33, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Rivas, L.; Coffey, B.; McAuliffe, O.; McDonnell, M.J.; Burgess, C.M.; Coffey, A.; Ross, R.P.; Duffy, G. In vivo and ex vivo evaluations of bacteriophages e11/2 and e4/1c for use in the control of Escherichia coli O157:H7. Appl. Environ. Microbiol. 2010, 76, 7210–7216. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.A.; McNair, K.; Faust, K.; Raes, J.; Dulith, B.E. Computational approaches to predict bacteriophage-host relationships. FEMS Microbiol. Rev. 2016, 40, 258–272. [Google Scholar] [CrossRef] [PubMed]
- Villarroel, J.; Kleinheinz, K.A.; Jurtz, V.I.; Zschach, H.; Lund, O.; Nielsen, M.; Larsen, M.V. HostPhinder: A phage host prediction tool. Viruses 2016, 8, 116. [Google Scholar] [CrossRef] [PubMed]
- Galiez, C.; Siebert, M.; Enault, F.; Vincent, J.; Soding, J. WisH: Who is the host? Predicting prokaryotic hosts from metagenomic phage contigs. Bioinformatics 2017, 33, 3113–3114. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chen, X.; Hu, Y.; Zhu, G.; White, A.P.; Koster, W. Evolution and Sequence Diversity of FhuA in Salmonella and Escherichia. Infect. Immunity 2018, 86. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, J.P.; Blanc-Potard, A.B. Molecular evolution of Salmonella enterica serovar Typhimurium and pathogenic Escherichia coli: From pathogenesis to therapeutics. Infect. Genet. Evol. 2008, 8, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, C.; Kaniuk, N.; Frirdich, E. Molecular insights into the assembly and diversity of the outer core oligosaccharide in lipopolysaccharides from Escherichia coli and Salmonella. J. Endotoxin. Res. 2003, 9, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Achtman, M. Insights from genomic comparisons of genetically monomorphic bacterial pathogens. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2012, 367, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Keim, P.; Gruendike, J.M.; Klevytska, A.M.; Schupp, J.M.; Challacombe, J.; Okinaka, R. The genome and variation of Bacillus anthracis. Mol. Aspects Med. 2009, 30, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.J.; Gill, J.J. The habits of highly effective phages: Population dynamics as a framework for identifying therapeutic phages. Front. Microbiol. 2014, 5, 618. [Google Scholar] [CrossRef] [PubMed]
- Dhar, B.; Singh, B.D.; Singh, R.B.; Srivastava, J.S.; Singh, V.P.; Singh, R.M. Occurrence and distribution of rhizobiophages in Indian soils. Acta Microbiol. Pol. 1979, 28, 319–324. [Google Scholar] [PubMed]
- Ackermann, H.W. Basic phage electron microscopy. Methods Mol. Biol. 2009, 501, 113–126. [Google Scholar] [PubMed]
- Endersen, L.; Buttimer, C.; Nevin, E.; Coffey, A.; Neve, H.; Oliveira, H.; Lavigne, R.; O’Mahony, J. Investigating the biocontrol and anti-biofilm potential of a three phage cocktail against Cronobacter sakazakii in different brands of infant formula. Int. J. Food Microbiol. 2017, 253, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Revazishvili, T.; Dean, T.; Butani, A.; Verratti, K.; Bishop-Lilly, K.A.; Sozhamannan, S.; Sulakvelidze, A.; Rajanna, C. A Yersinia pestis-specific, lytic phage preparation significantly reduces viable, Y. pestis on various hard surfaces experimentally contaminated with the bacterium. Bacteriophage 2012, 2, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Aziz, R.K.; Ackermann, H.-W.; Petty, N.K.; Kropinski, A.M. Essential steps in characterizing bacteriophages; Biology, taxonomy, and genome analysis. Methods Mol. Biol. 2018, 1681, 197–216. [Google Scholar] [PubMed]
- Pope, W.H.; Jacobs-Sera, D. Annotation of bacteriophage genome sequences using DNA Master: An overview. Methods Mol. Biol. 2018, 1681, 217–230. [Google Scholar] [PubMed]
- McNair, K.; Aziz, R.K.; Pusch, G.D.; Overbeck, R.; Dutilh, B.E.; Edwards, R. Phage genome annotation using the RAST pipeline. Methods Mol. Biol. 2018, 1681, 231–238. [Google Scholar] [PubMed]
- Turner, D.; Sutton, J.M.; Reynolds, D.M.; Sim, E.M.; Petty, N.K. Visualization of phage genomic data; Comparative genomics and publication quality diagrams. Methods Mol. Biol. 2018, 1681, 239–260. [Google Scholar] [PubMed]
- Simmonds, P.; Aiewsakun, P. Virus classification—Where do you draw the line? Arch. Virol. 2018, 163, 2037–2046. [Google Scholar] [CrossRef] [PubMed]
- Tolstoy, I.; Kropinski, A.M.; Brister, J.R. Bacteriophage taxonomy: An evolving discipline. Methods Mol. Biol. 2018, 1693, 57–71. [Google Scholar] [PubMed]
- Parmar, K.M.; Dafale, N.A.; Tikariha, H.; Purohit, H.J. Genomic characterization of key bacteriophages to formulate the potential biocontrol agent to combat enteric pathogenic bacteria. Arch. Microbiol. 2018, 200, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Feng, Y.; Zong, Z. Two new lytic bacteriophages of the Myoviridae family against carbapenem-resistant Acinetobacter baumannii. Front. Microbiol. 2018, 9, 850. [Google Scholar] [CrossRef] [PubMed]
- Kropinski, A.M. Practical advice on the one-step growth curve. Methods Mol. Biol. 2018, 1681, 41–47. [Google Scholar] [PubMed]
- Hyman, P.; Abedon, S.T. Practical methods for determining phage growth parameters. Methods Mol. Biol. 2009, 501, 175–202. [Google Scholar] [PubMed]
- Fan, N.; Qi, R.; Yang, M. Isolation and characterization of a virulent bacteriophage infecting Acinetobacter johnsonii from activated sludge. Res. Microbiol. 2017, 168, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Xing, S.; Sun, Q.; Pei, G.; Cheng, S.; Liu, Y.; An, X.; Zhang, X.; Qu, Y.; Tong, Y. Characterization and complete genome sequence analysis of a novel virulent Siphoviridae phage against Staphylococcus aureus isolated from bovine mastitis in Xinjiang, China. Virus Genes 2017, 53, 464–476. [Google Scholar] [CrossRef] [PubMed]
- Lingohr, E.; Frost, S.; Johnson, R.P. Determination of bacteriophage genome size by pulsed-field gel electrophoresis. Methods Mol. Biol. 2009, 502, 19–25. [Google Scholar] [PubMed]
- Krasowska, A.; Biegalska, A.; Augustyniak, D.; Los, M.; Richert, M.; Lukaszewicz, M. Isolation and characterization of phages infecting Bacillus subtilis. BioMed Res. Internat. 2105, 2015, 179597. [Google Scholar]
- Czajkowski, R.; Ozymko, Z.; de Jager, V.; Siwinska, J.; Smolarska, A.; Ossowicki, A.; Narajczyk, M.; Lojkowska, E. Genomic, proteomic and morphological characterization of two novel broad host lytic bacteriophages φPD10.3 and φPD23.1 infecting pectinolytic Pectobacterium spp. And Dickeya spp. PLoS ONE 2015, 10, e0119812. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Liao, G.; Liu, C.; Jiang, X.; Lin, M.; Zhao, C.; Tao, J.; Huang, Z. Characterization of bacteriophage HN48 and its protective effects in Nile tilapia Oreochromis niloticus against Streptococcus agalactiae infections. J. Fish. Dis. 2018, 41, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Abedon, S.T. Phage therapy dosing: The problem(s) with multiplicity of infection (MOI). Bacteriophage 2016, 6, e1220348. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration. Microbiology Data for Systemic Antibacterial Drugs—Development, Analysis, and Presentation Guidance for Industry, Revision 2; Food and Drug Administration: Silver Spring, MD, USA, 2018; Available online: https://www.fda.gov/downloads/drugs/guidancecomplianceregulatoryinformation/guidances/ucm182288.pdf (accessed on 21 December 2018).
- Bolmstrom, A.; Karlsson, A.; Engelhardt, A.; Ho, P.; Petersen, P.J.; Bradford, P.A.; Jones, C.H. Validation and reproducibility assessment of tigecycline MIC determinations by Etest. J. Clin. Microbiol. 2007, 45, 2474–2479. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Method | Description | Advantages | Limitations * | Example References |
|---|---|---|---|---|
| Spot testing | A plate is inoculated with host bacteria to form a lawn, then small drops of phage filtrate are placed on the surface. After incubation a zone of lysis indicates presence of phage. | Simple. Allows testing of multiple phage filtrates on the same plate. | Host must grow to confluence on solid media. Prone to false positives due to bacterial killing by media components or phage binding that does not lead to a productive infection. | [76,77] |
| Plaque testing | Increasing dilutions of phage filtrate are mixed with bacteria and placed on plate surface by spreading or soft agar overlay. After incubation plate is checked for the appearance of plaques. | Demonstrates productive phage growth. Plaque appearance can suggest lytic vs. temperate life cycle. Plaque size may suggest phage size due to diffusion effects (larger phages diffuse more slowly, etc.) | Host must grow to confluence on solid media. Not all phages are capable of forming plaques even on productive hosts due to limited diffusion in agar or low productivity. | [78,79,80] |
| Culture lysis | Phage filtrate added to broth culture of bacteria and incubated. Monitored for cell lysis as indicated by loss of culture turbidity. Metabolic dyes can be used instead of turbidity to assess bacterial metabolic activity [81]. | Useful for bacteria that will not grow to confluence on solid media as well as any bacteria that grows well in broth. Can be adapted to automation using spectrophotometry to measure turbidity either in single tube or multiple well plates. | As with spot testing, can have false positives due to non-productive lysis. Cell debris from cells lysed during early infections may bind to and inactivate free phages, interfering with later infections. Hosts that rapidly evolve phage resistant mutants will cause false negatives as mutants maintain turbidity. | [25,82] |
| Routine test dilution (RTD) | Phage lysates are diluted to the point of producing just less than confluent lysis on a plate. | Useful with phage that do not form distinct plaques or very small indistinct plaques. | Prone to false positives when media components are not highly diluted. | [83,84,85] † |
| Property | Description (Reference for Methodology) | References (Examples) |
|---|---|---|
| Efficiency of plating (EOP) | Number of plaques or lysis measurements are compared to a reference phage/host combination for relative EOP. Number of plaques compared to the number of phage particles (as determined by a non-culture method such as epifluoresence microscopy counting) used for infection is the absolute EOP. EOP between any phage pair may vary on different hosts. Often performed when multiple phages have been isolated to establish which are more or less virulent, especially when planning to combine phages to make a cocktail. EOP of each cocktail component prevents use of phages with very low killing efficiency [60]. | [157] |
| Phage morphology by electron microscopy | Until the advent of next-generation sequencing of phage genomes, morphology was considered essential for classifying novel phages and phages of different morphologies were often used to make a broader, more diverse phage cocktail. Classification is now more commonly done by genome comparisons but knowing the morphology is still useful for quickly and easily showing diversity in a phage mixture [158]. | [159,160] |
| Whole genome sequence | If not determined earlier, this can confirm lack of toxin genes and ability to form a lysogen. Also somewhat useful for identifying related phages for inferences of some gene functions, for example [135]. For overviews of sequencing methods see [62,135,161,162,163,164]. Phage taxonomic classification is still under development so using sequence data for this is potentially limited [165,166]. | [167,168] |
| One step growth curve | Phages with long latent periods may be less useful as therapeutic phages. Burst size can also be determined if the treatment will be with fewer phage than can immediately infect all the bacteria infecting the patient. As discussed elsewhere in this paper, it is also important to remember that growth rates in the laboratory may not reflect growth rates in patients, in biofilms, and in other potential phage application locations [60,169,170]. | [171,172] |
| Pulse-field gel electrophoresis (PFGE) | PFGE is a modification of conventional agarose gel electrophoresis which uses a variable electric voltage to drive DNA migration instead of the standard continuous voltage. This allows for the separation of much larger pieces of DNA. It can be used to directly measure the size of a phage genome. It can also be used for restriction fragment length polymorphism (RFLP) analysis when multiple phages have been isolated to show that each phage is different by comparing restriction enzyme digested genomic DNA although depending on the size of the phage genome, this differentiation can also be done using conventional gel electrophoresis. [173] | [174,175] |
| Multiple strains of one species | 14 (29%) |
| Closely related species | 13 (27%) |
| Distantly related species | 4(8%) |
| As diverse as possible | 11(22%) |
| Multiple answers | 7 (14%) |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyman, P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals 2019, 12, 35. https://doi.org/10.3390/ph12010035
Hyman P. Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals. 2019; 12(1):35. https://doi.org/10.3390/ph12010035
Chicago/Turabian StyleHyman, Paul. 2019. "Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth" Pharmaceuticals 12, no. 1: 35. https://doi.org/10.3390/ph12010035
APA StyleHyman, P. (2019). Phages for Phage Therapy: Isolation, Characterization, and Host Range Breadth. Pharmaceuticals, 12(1), 35. https://doi.org/10.3390/ph12010035