Thermal Imaging Reliability for Estimating Grain Yield and Carbon Isotope Discrimination in Wheat Genotypes: Importance of the Environmental Conditions

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Experimental Conditions
2.2. Evaluations
2.2.1. Grain Yield and Carbon Isotope Discrimination
2.2.2. Thermography
2.3. Statistical Design and Data Analysis
3. Results
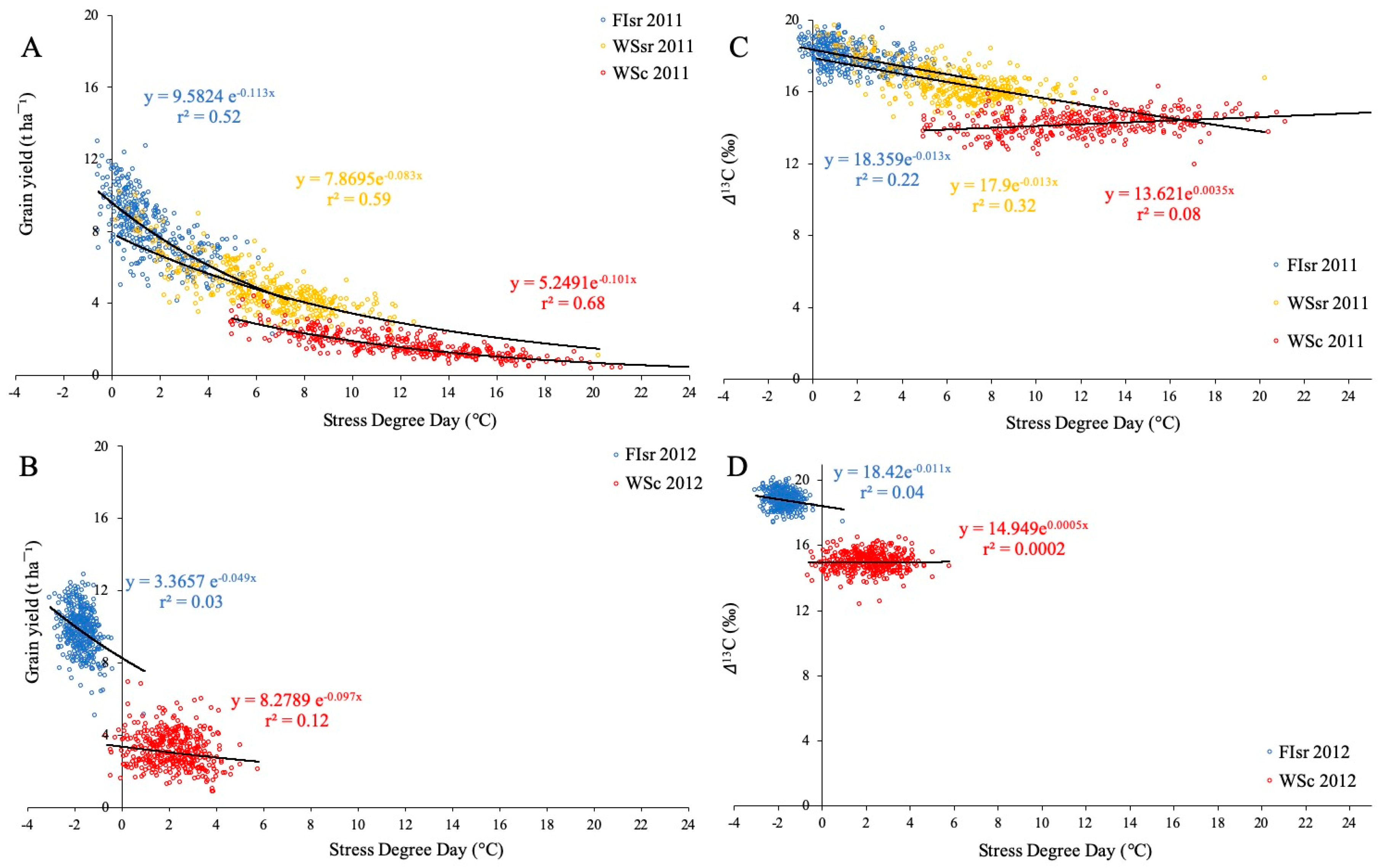
3.1. Environmental Conditions, Grain Yield, Carbon Isotope Discrimination, and Stress Degree Days
3.2. Relationships between Grain Yield and Canopy and Ambient Temperatures in 384 Wheat Genotypes
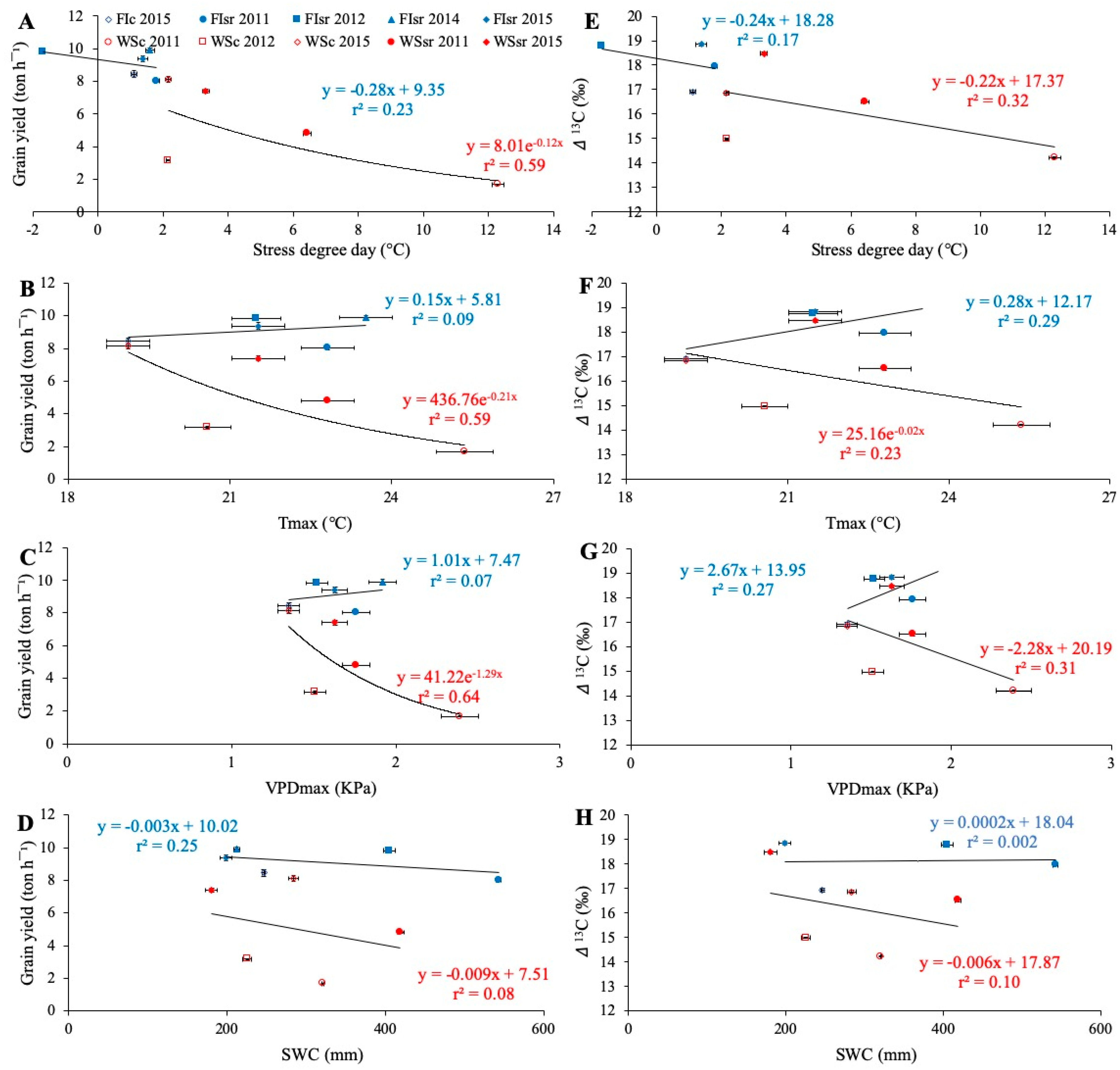
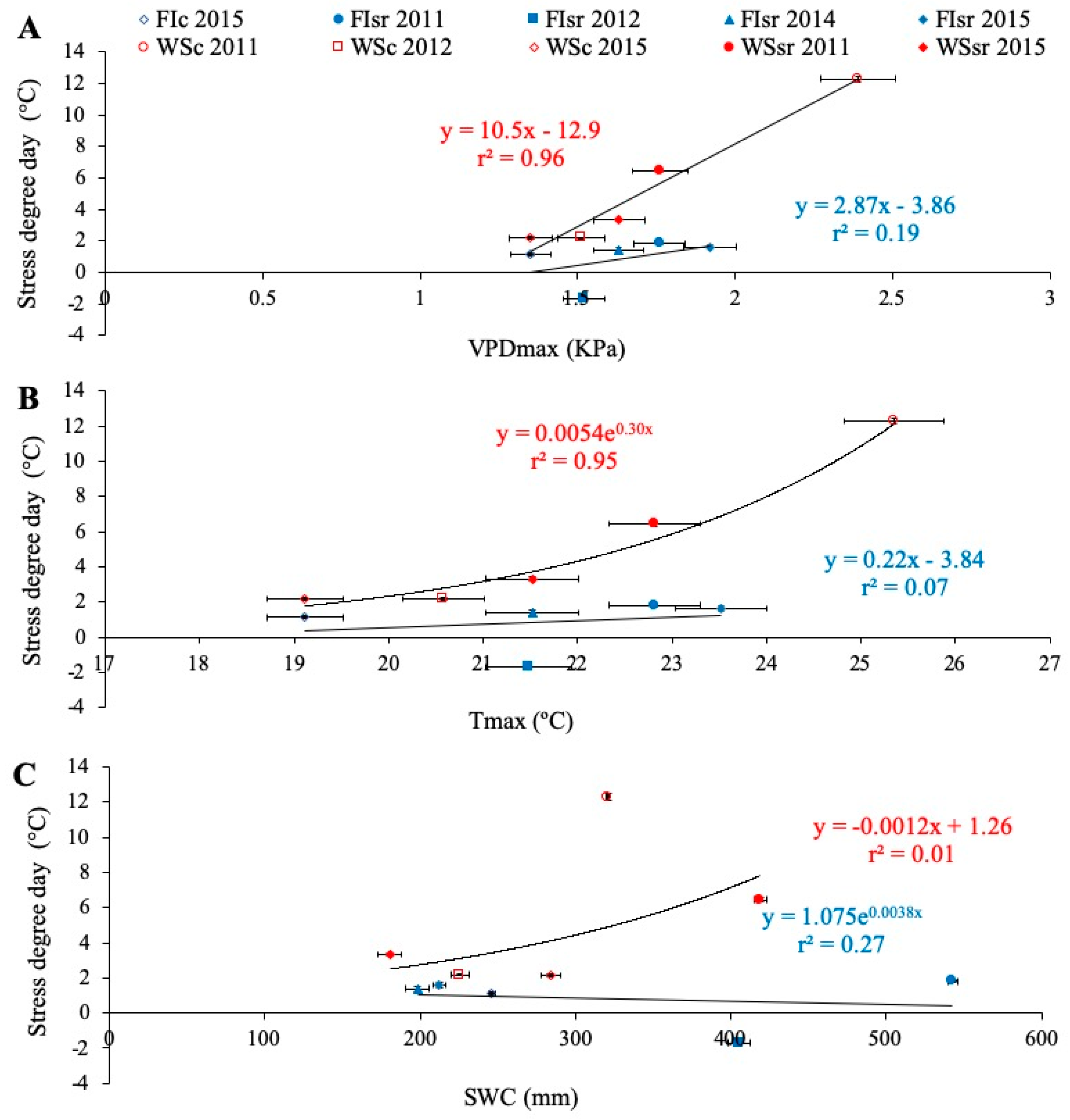
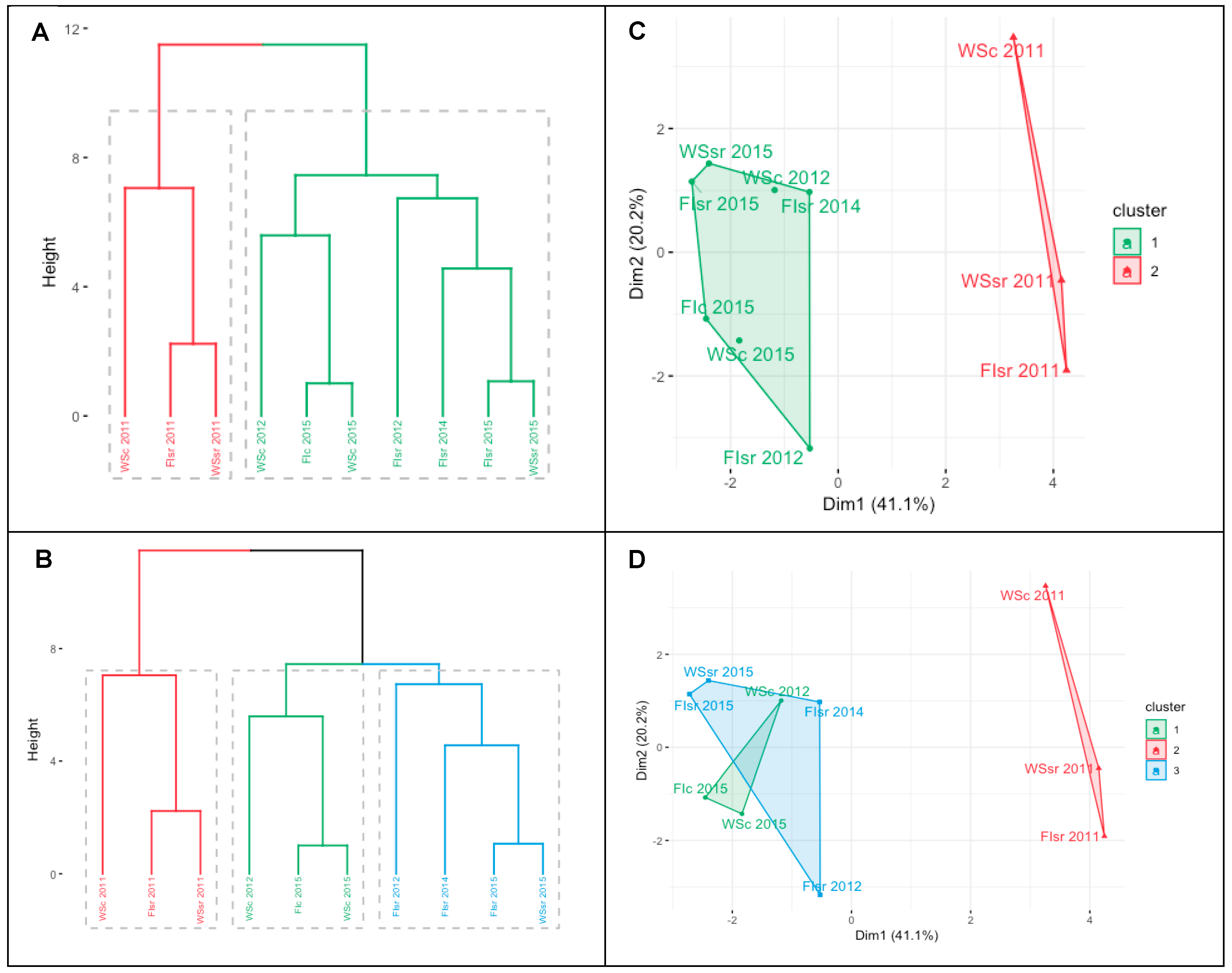
3.3. Environmental Effects on Grain Yield, Carbon Isotope Discrimination, and Stress Degree Day
4. Discussion
4.1. Environmental Effects on Grain Yield and Carbon Isotope Discrimination
4.2. The Potential of Stress Degree Day to Estimate Grain Yield
5. Conclusions and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Tanner, C.B. Plant Temperatures 1. Agron. J. 1963, 55, 210–211. [Google Scholar] [CrossRef]
- Zia, S.; Spohrer, K.; Merkt, N.; Wenyong, D.; He, X.; Müller, J. Non-invasive water status detection in grapevine (Vitis vinifera L.) by thermography. Int. J. Agric. Biol. Eng. 2009, 2, 46–54. [Google Scholar] [CrossRef]
- Jones, H.G.; Stoll, M.; Santos, T.; Sousa, C.D.; Chaves, M.M.; Grant, O.M. Use of infrared thermography for monitoring stomatal closure in the field: Application to grapevine. J. Exp. Bot. 2002, 53, 2249–2260. [Google Scholar] [CrossRef] [PubMed]
- Leinonen, I.; Jones, H.G. Combining thermal and visible imagery for estimating canopy temperature and identifying plant stress. J. Exp. Bot. 2004, 55, 1423–1431. [Google Scholar] [CrossRef] [PubMed]
- Möller, M.; Alchanatis, V.; Cohen, Y.; Meron, M.; Tsipris, J.; Naor, A.; Cohen, S. Use of thermal and visible imagery for estimating crop water status of irrigated grapevine. J. Exp. Bot. 2007, 58, 827–838. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.S.; Pan, Y.J.; Zhao, X.Q.; Dwivedi, D.; Zhu, L.H.; Ali, J.; Li, Z.K. Drought-induced site-specific DNA methylation and its association with drought tolerance in rice (Oryza sativa L.). J. Exp. Bot. 2010, 62, 1951–1960. [Google Scholar] [CrossRef] [PubMed]
- Zia, S.; Spohrer, K.; Wenyong, D.; Spreer, W.; Romano, G.; Xiongkui, H.; Joachim, M. Monitoring physiological responses to water stress in two maize varieties by infrared thermography. Int. J. Agric. Biol. Eng. 2011, 4, 7–15. [Google Scholar] [CrossRef]
- Jackson, R.D.; Idso, S.B.; Reginato, R.J.; Pinter, P.J., Jr. Canopy temperature as a crop water stress indicator. Water Resour. Res. 1981, 17, 1133–1138. [Google Scholar] [CrossRef]
- Luquet, D.; Bégué, A.; Vidal, A.; Clouvel, P.; Dauzat, J.; Olioso, A.; Tao, Y. Using multidirectional thermography to characterize water status of cotton. Remote Sens. Environ. 2003, 84, 411–421. [Google Scholar] [CrossRef]
- Jones, H.G. Application of thermal imaging and infrared sensing in plant physiology and ecophysiology. Adv. Bot. Res. 2004, 41, 107–163. [Google Scholar] [CrossRef]
- Amani, I.; Fischer, R.A.; Reynolds, M.P. Canopy temperature depression association with yield of irrigated spring wheat cultivars in a hot climate. J. Agron. Crop Sci. 1996, 176, 119–129. [Google Scholar] [CrossRef]
- Cossani, C.M.; Pietragalla, J.; Reynolds, M. Temperatura del dosel vegetal y características de la relación planta-agua. In Fitomejoramiento fisiológico I: Enfoques interdisciplinarios para mejorar la adaptación del cultivo; Reynolds, M.P., Pask, A.J.D., Mullan, D.M., Chávez-Dulanto, P.N., Eds.; CIMMYT: Ciudad de Mexico, Mexico, 2013; pp. 60–68. ISBN 978-607-8263-18-9. [Google Scholar]
- Fischer, R.A.; Rees, D.; Sayre, K.D.; Lu, Z.M.; Condon, A.G.; Saavedra, A.L. Wheat yield progress associated with higher stomatal conductance and photosynthetic rate, and cooler canopies. Crop Sci. 1998, 38, 1467–1475. [Google Scholar] [CrossRef]
- Leinonen, I.; Grant, O.M.; Tagliavia, C.P.P.; Chaves, M.M.; Jones, H.G. Estimating stomatal conductance with thermal imagery. Plant Cell Environ. 2006, 29, 1508–1518. [Google Scholar] [CrossRef] [PubMed]
- Grant, O.M.; Tronina, Ł.; Jones, H.G.; Chaves, M. Exploring thermal imaging variables for the detection of stress responses in grapevine under different irrigation regimes. J. Exp. Bot. 2007, 58, 815–825. [Google Scholar] [CrossRef] [PubMed]
- Prashar, A.; Jones, H.G. Infra-red thermography as a high-throughput tool for field phenotyping. Agronomy 2014, 4, 397–417. [Google Scholar] [CrossRef]
- Prashar, A.; Jones, H.G. Assessing drought responses using thermal infrared imaging. In Environmental Responses in Plants. Methods and Protocols, Methods in Molecular Biology; Duque, P., Ed.; Humana Press: New York, NY, USA, 2016; Volume 1398, pp. 209–219. [Google Scholar] [CrossRef]
- Blum, A.; Shpiler, L.; Golan, G.; Mayer, J. Yield stability and canopy temperature of wheat genotypes under drought-stress. Field Crops Res. 1989, 22, 289–296. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Farquhar, G.D. Carbon isotope discrimination is positively correlated with grain yield and dry matter production in field-grown wheat. Crop Sci. 1987, 27, 996–1001. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Hubick, K.T.; Condon, A.G.; Richards, R.A. Carbon isotope fractionation and plant water-use efficiency. In Stable Isotopes in Ecological Research; Rundel, P.W., Ehleringer, J.R., Nagy, K.A., Eds.; Springer: New York, NY, USA, 1989; pp. 21–40. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Farquhar, G.D. The effect of variation in soil water availability, vapour pressure deficit and nitrogen nutrition on carbon isotope discrimination in wheat. Aust. J. Agric. Res. 1992, 43, 935–947. [Google Scholar] [CrossRef]
- Araus, J.L.; Amaro, T.; Casadesus, J.; Asbati, A.; Nachit, M.M. Relationships between ash content, carbon isotope discrimination and yield in durum wheat. Funct. Plant Biol. 1998, 25, 835–842. [Google Scholar] [CrossRef]
- Araus, J.L.; Villegas, D.; Aparicio, N.; Del Moral, L.F.; El Hani, S.; Rharrabti, Y.; Royo, C. Environmental factors determining carbon isotope discrimination and yield in durum wheat under Mediterranean conditions. Crop Sci. 2003, 43, 170–180. [Google Scholar] [CrossRef]
- Lobos, G.A.; Matus, I.; Rodriguez, A.; Romero-Bravo, S.; Araus, J.L.; del Pozo, A. Wheat genotypic variability in grain yield and carbon isotope discrimination under mediterranean conditions assessed by spectral reflectance. J. Integr. Plant Biol. 2014, 56, 470–479. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Bosquet, L.; Albrizio, R.; Nogués, S.; Araus, J.L. Dual Δ13C/δ18O response to water and nitrogen availability and its relationship with yield in field-grown durum wheat. Plant Cell Environ. 2010, 34, 418–433. [Google Scholar] [CrossRef] [PubMed]
- Garriga, M.; Romero-Bravo, S.; Estrada, F.; Escobar, A.; Matus, I.; del Pozo, A.; Astudillo, C.; Lobos, G. Assessing wheat traits by spectral reflectance: Do we really need to focus on predicted trait-values or directly identify the elite genotypes group? Front. Plant Sci. 2017, 8, 280. [Google Scholar] [CrossRef] [PubMed]
- Moretti, C.L.; Mattos, L.M.; Calbo, A.G.; Sargent, S.A. Climate changes and potential impacts on postharvest quality of fruit and vegetable crops: A review. Food Res. Int. 2010, 43, 1824–1832. [Google Scholar] [CrossRef]
- Lobos, G.A.; Hancock, J.F. Breeding blueberries for a changing global environment: A review. Front. Plant Sci. 2015, 6, 782. [Google Scholar] [CrossRef] [PubMed]
- Camargo, A.V.; Lobos, G.A. Latin America: A development pole for phenomics. Front. Plant Sci. 2016, 7, 1729. [Google Scholar] [CrossRef] [PubMed]
- Lobos, G.A.; Camargo, A.V.; del Pozo, A.; Araus, J.L.; Ortiz, R.; Doonan, J.H. Plant Phenotype and phenomics for plant breeding. Front. Plant Sci. 2017, 8, 2181. [Google Scholar] [CrossRef]
- Sadras, V.O.; Reynolds, M.P.; De la Vega, A.J.; Petrie, P.R.; Robinson, R. Phenotypic plasticity of yield and phenology in wheat, sunflower and grapevine. Field Crops Res. 2009, 110, 242–250. [Google Scholar] [CrossRef]
- Mora, F.; Castillo, D.; Lado, B.; Matus, I.; Poland, P.; Belzile, F.; von Zitzewitz, J.; del Pozo, A. Genome-wide association mapping of agronomic traits and carbon discrimination in a worldwide germplasm collection of spring wheat using SNP markers. Mol. Breed. 2015, 35. [Google Scholar] [CrossRef]
- del Pozo, A.; Yáñez, A.; Matus, I.A.; Tapia, G.; Castillo, D.; Sanchez-Jardón, L.; Araus, J.L. Physiological traits associated with wheat yield potential and performance under water-stress in a mediterranean environment. Front. Plant Sci. 2016, 7, 987. [Google Scholar] [CrossRef]
- Zhang, D.; Du, Q.; Zhang, Z.; Jiao, X.; Song, X.; Li, J. Vapour pressure deficit control in relation to water transport and water productivity in greenhouse tomato production during summer. Sci. Rep. 2017, 7, 43461. [Google Scholar] [CrossRef] [PubMed]
- Hernández, J.; Lobos, G.A.; Matus, I.; del Pozo, A.; Silva, P.; Galleguillos, M. Using ridge regression models to estimate grain yield from field spectral data in bread wheat (Triticum aestivum L.) grown under three water regimes. Remote Sens. 2015, 7, 2109–2126. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Allen, R.G.; Pereira, L.S.; Raes, D.; Smith, M. Crop Evapotranspiration: Guidelines for Computing Crop Water Requirements. In FAO Irrigation and Drainage Paper N° 56; FAO: Rome, Italy, 1998; 300p. [Google Scholar]
- Fuentes, S.; De Bei, R.; Pech, J.; Tyerman, S. Computational water stress indices obtained from thermal image analysis of grapevine canopies. Irrig. Sci. 2012, 30, 523–536. [Google Scholar] [CrossRef]
- Idso, S.B.; Jackson, R.D.; Reginato, R.J. Remote sensing of crop yields. Science 1977, 196, 19–25. [Google Scholar] [CrossRef]
- Jackson, R.D.; Reginato, R.J.; Idso, S.B. Wheat canopy temperature: A practical tool for evaluating water requirements. Water Resour. Res. 1977, 13, 651–656. [Google Scholar] [CrossRef]
- Nerurkar, P.; Shirke, A.; Chandane, M.; Bhirud, S. Empirical analysis of data clustering algorithms. Procedia Comput. Sci. 2018, 125, 770–779. [Google Scholar] [CrossRef]
- Murtagh, F.; Legendre, P. Ward’s hierarchical agglomerative clustering method: Which algorithms implement ward’s criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]
- Ryan, A.C.; Dodd, I.C.; Rothwell, S.A.; Jones, R.; Tardieu, F.; Draye, X.; Davies, W.J. Gravimetric phenotyping of whole plant transpiration responses to atmospheric vapour pressure deficit identifies genotypic variation in water use efficiency. Plant Sci. 2016, 251, 101–109. [Google Scholar] [CrossRef]
- Farquhar, G.D.; O’Leary, M.H.; Berry, J.A. On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves. Funct. Plant Biol. 1982, 9, 121–137. [Google Scholar] [CrossRef]
- Acevedo, E.H.; Baginsky, C.G.; Solar, B.R.; Ceccarelli, S. Discriminación isotópica de C13 y su relación con el rendimiento y la eficiencia de transpiracion de genotipos locales y mejorados de cebada bajo diferentes condiciones hídricas. Inv. Agric. 1997, 17, 41–54. [Google Scholar]
- Rebetzke, G.J.; Condon, A.G.; Farquhar, G.D.; Appels, R.; Richards, R.A. Quantitative trait loci for carbon isotope discrimination are repeatable across environments and wheat mapping populations. Theor. Appl. Genet. 2008, 118, 123–137. [Google Scholar] [CrossRef] [PubMed]
- del Pozo, A.; Castillo, D.; Inostroza, L.; Matus, I.; Méndez, A.M.; Morcuende, R. Physiological and yield responses of recombinant chromosome substitution lines of barley to terminal drought in a mediterranean type environment. Ann. Appl. Biol. 2012, 160, 157–167. [Google Scholar] [CrossRef]
- Araus, J.L.; Cabrera-Bosquet, L.; Serret, M.D.; Bort, J.; Nieto-Taladriz, M.T. Comparative performance of δ13C, δ18O and δ15N for phenotyping durum wheat adaptation to a dryland environment. Funct. Plant Biol. 2013, 40, 595–608. [Google Scholar] [CrossRef]
- Passioura, J.B. Grain yield, harvest index, and water use of wheat. J. Aust. Inst. Agric. Sci. 1977, 43, 117–120. [Google Scholar]
- French, R.J.; Schultz, J.E. Water use efficiency of wheat in a Mediterranean-type environment. I. The relation between yield, water use and climate. Aust. J. Agric. Res. 1984, 35, 743–764. [Google Scholar] [CrossRef]
- Sun, H.Y.; Liu, C.M.; Zhang, X.Y.; Shen, Y.J.; Zhang, Y.Q. Effects of irrigation on water balance, yield and WUE of winter wheat in the north China plain. Agric. Water Manag. 2006, 85, 211–218. [Google Scholar] [CrossRef]
- Li, F.M.; Liu, X.L.; Li, S.Q. Effects of early soil water distribution on the dry matter partition between roots and shoots of winter wheat. Agric. Water Manag. 2001, 49, 163–171. [Google Scholar] [CrossRef]
- Anderson, D.B. Relative humidity or vapor pressure deficit. Ecology 1936, 17, 277–282. [Google Scholar] [CrossRef]
- Dreccer, M.F.; Fainges, J.; Whish, J.; Ogbonnaya, F.C.; Sadras, V.O. Comparison of sensitive stages of wheat, barley, canola, chickpea and field pea to temperature and water stress across Australia. Agric. For. Meteorol. 2018, 248, 275–294. [Google Scholar] [CrossRef]
- Rashid, M.A.; Andersen, M.N.; Wollenweber, B.; Zhang, X.; Olesen, J.E. Acclimation to higher VPD and temperature minimized negative effects on assimilation and grain yield of wheat. Agric. For. Meteorol. 2018, 248, 119–129. [Google Scholar] [CrossRef]
- Sharifi, M.R.; Rundel, P.W. The effect of vapour pressure deficit on carbon isotope discrimination in the desert shrub Larrea tridentata (creosote bush). J. Exp. Bot. 1993, 44, 481–487. [Google Scholar] [CrossRef]
- Franks, P.J. Stomatal control and hydraulic conductance, with special reference to tall trees. Tree Physiol. 2004, 24, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Blum, A.; Mayer, J.; Gozlan, G. Infrared thermal sensing of plant canopies as a screening technique for dehydration avoidance in wheat. Field Crops Res. 1982, 5, 137–146. [Google Scholar] [CrossRef]
- Reynolds, M.P.; Balota, M.; Delgado, M.I.B.; Amani, I.; Fischer, R.A. Physiological and morphological traits associated with spring wheat yield under hot, irrigated conditions. Funct. Plant Biol. 1994, 21, 717–730. [Google Scholar] [CrossRef]
- Olivares-Villegas, J.J.; Reynolds, M.P.; McDonald, G.K. Drought-adaptive attributes in the Seri/Babax hexaploid wheat population. Funct. Plant Biol. 2007, 34, 189–203. [Google Scholar] [CrossRef]
- Idso, S.B.; Reginato, R.J.; Clawson, K.L.; Anderson, M.G. On the stability of non-water-stressed baselines. Agric. For. Meteorol. 1984, 32, 177–182. [Google Scholar] [CrossRef]
- Hatfield, J.L. Measuring plant stress with an infrared thermometer. HortScience 1990, 25, 1535–1538. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Trial code | n | Dates | Means | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Sowing | Anthesis | Grain filling | Harvesting | GY (t ha−1) | Δ13C (‰) | SDD (°C) | Tmax (°C) | VPDmax (kPa) | SWC (mm) | |||
| FIsr 2011 | 384 | 31 Aug. 2011 | 24 Nov. 2011 | 22 Dec. 2011 | 11 Jan. 2012 | 8.03 | 18.0 | 1.81 | 23.4 | 1.76 | 542.7 | |
| FIsr 2012 | 384 | 07 Aug. 2012 | 05 Nov. 2012 | 19 Nov. 2012 | 28 Jan. 2013 | 9.83 | 18. 8 | −1.70 | 21.5 | 1.52 | 404.8 | |
| FIsr 2014 | 64 | 27 Aug. 2014 | 24 Nov. 2014 | 17 Dec. 2014 | 22 Jan. 2015 | 9.90 | - | 1.60 | 23.5 | 1.92 | 212.3 | |
| FIsr 2015 | 64 | 29 Jul. 2015 | 20 Nov. 2015 | 08 Dec. 2015 | 25 Jan. 2016 | 9.38 | 18.8 | 1.38 | 21.5 | 1.63 | 198.3 | |
| FIc 2015 | 64 | 18 May 2015 | 23 Oct. 2015 | 12 Nov. 2015 | 23 Dec. 2015 | 8.46 | 16.9 | 1.12 | 19.1 | 1.35 | 246.2 | |
| FI | Average | 9.12 | 18.1 | 0.84 | 21.8 | 1.64 | 320.9 | |||||
| FI | SD | 0.84 | 0.90 | 1.44 | 1.79 | 0.22 | 148.9 | |||||
| FI | Min. | 8.03 | 16.9 | −1.70 | 19.1 | 1.35 | 198.3 | |||||
| FI | Max. | 9.90 | 18.8 | 1.81 | 23.5 | 1.92 | 542.7 | |||||
| WSsr 2011 | 384 | 31 Aug. 2011 | 24 Nov. 2011 | 22 Dec. 2011 | 11 Jan. 2012 | 4.81 | 16.5 | 6.44 | 23.4 | 1.76 | 418.8 | |
| WSc 2011 | 384 | 07 Sep. 2011 | 29 Nov. 2011 | 13 Dec. 2011 | 05 Jan. 2012 | 1.68 | 14.2 | 12.29 | 25.4 | 2.39 | 320.9 | |
| WSc 2012 | 384 | 23 May 2012 | 11 Sep. 2012 | 25 Oct. 2012 | 23 Dec. 2012 | 3.18 | 15.0 | 2.17 | 20.6 | 1.51 | 225.5 | |
| WSsr 2015 | 64 | 29 Jul. 2015 | 20 Nov. 2015 | 08 Dec. 2015 | 25 Jan. 2016 | 7.40 | 18.5 | 3.31 | 21.5 | 1.63 | 180.4 | |
| WSc 2015 | 64 | 18 May 2015 | 23 Oct. 2015 | 12 Nov. 2015 | 23 Dec. 2015 | 8.13 | 16.9 | 2.16 | 19.1 | 1.35 | 283.8 | |
| WS | Average | 5.04 | 16.2 | 5.27 | 22.0 | 1.74 | 285.9 | |||||
| WS | SD | 2.45 | 1.49 | 4.29 | 2.18 | 0.36 | 91.8 | |||||
| WS | Min. | 1.68 | 14.2 | 2.16 | 19.1 | 1.35 | 180.4 | |||||
| WS | Max. | 8.13 | 18.5 | 12.3 | 25.4 | 2.39 | 418.8 | |||||
| GY | −0.81 ** | - | - | - | - |
| Δ13C | −0.71 * | 0.92 ** | - | - | - |
| Tmax | 0.64 * | −0.38 | −0.16 | - | - |
| VPDmax | 0.80 ** | −0.52 | −0.35 | 0.95 ** | - |
| SWC | 0.06 | −0.06 | −0.05 | 0.29 | 0.16 |
| SDD | GY | Δ13C | Tmax | VPDmax |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romero-Bravo, S.; Méndez-Espinoza, A.M.; Garriga, M.; Estrada, F.; Escobar, A.; González-Martinez, L.; Poblete-Echeverría, C.; Sepulveda, D.; Matus, I.; Castillo, D.; et al. Thermal Imaging Reliability for Estimating Grain Yield and Carbon Isotope Discrimination in Wheat Genotypes: Importance of the Environmental Conditions. Sensors 2019, 19, 2676. https://doi.org/10.3390/s19122676
Romero-Bravo S, Méndez-Espinoza AM, Garriga M, Estrada F, Escobar A, González-Martinez L, Poblete-Echeverría C, Sepulveda D, Matus I, Castillo D, et al. Thermal Imaging Reliability for Estimating Grain Yield and Carbon Isotope Discrimination in Wheat Genotypes: Importance of the Environmental Conditions. Sensors. 2019; 19(12):2676. https://doi.org/10.3390/s19122676
Chicago/Turabian StyleRomero-Bravo, Sebastián, Ana María Méndez-Espinoza, Miguel Garriga, Félix Estrada, Alejandro Escobar, Luis González-Martinez, Carlos Poblete-Echeverría, Daniel Sepulveda, Ivan Matus, Dalma Castillo, and et al. 2019. "Thermal Imaging Reliability for Estimating Grain Yield and Carbon Isotope Discrimination in Wheat Genotypes: Importance of the Environmental Conditions" Sensors 19, no. 12: 2676. https://doi.org/10.3390/s19122676
APA StyleRomero-Bravo, S., Méndez-Espinoza, A. M., Garriga, M., Estrada, F., Escobar, A., González-Martinez, L., Poblete-Echeverría, C., Sepulveda, D., Matus, I., Castillo, D., del Pozo, A., & Lobos, G. A. (2019). Thermal Imaging Reliability for Estimating Grain Yield and Carbon Isotope Discrimination in Wheat Genotypes: Importance of the Environmental Conditions. Sensors, 19(12), 2676. https://doi.org/10.3390/s19122676