
Acute Sleep Deprivation Induces a Local Brain Transfer Information Increase in the Frontal Cortex in a Widespread Decrease Context

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design of the Study
2.2. EEG Acquisition and Preprocessing
- Maximum amplitude lower than ±150 mV to avoid electrode-related artifacts or remaining ocular contamination.
- Absolute power in the 35 to 45 Hz band lower than 25 mV2 in each EEG channel in order to reject muscular artifacts. Frontopolar, frontal and temporal derivations had a higher threshold set to 50 mV2.
- Absolute power ratio of alpha (7.5 to 13 Hz) to delta (0.5 to 3.5 Hz) power higher than a variable threshold depending on the channel amplitude and global alpha activity present in the signal. This criterion was set to counterbalance a possible incomplete elimination of ocular artifacts, mainly indicated by an increase in delta activity.
2.3. Spectral Analysis
2.4. Transfer Entropy Analysis
2.5. Statistical Analysis
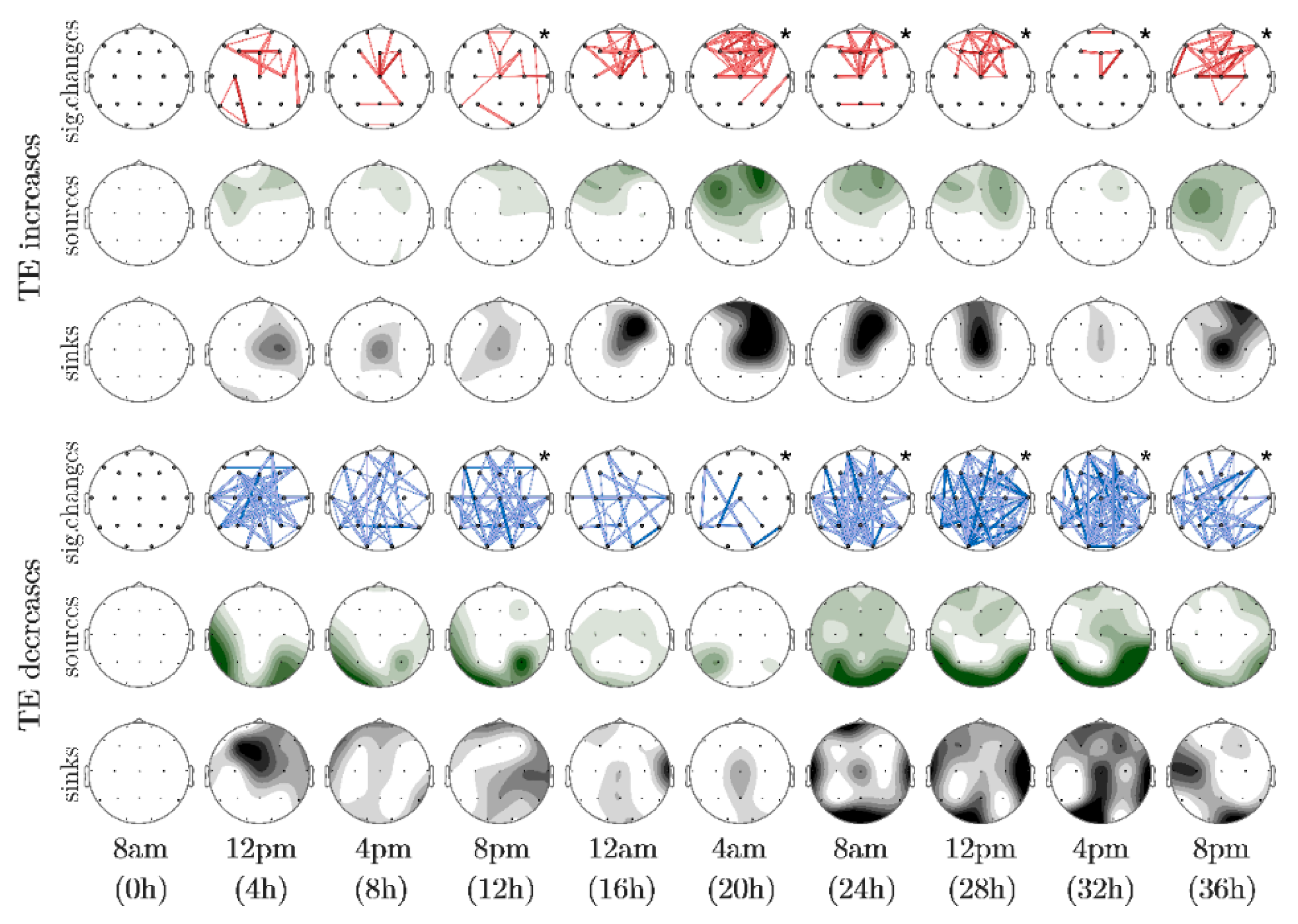
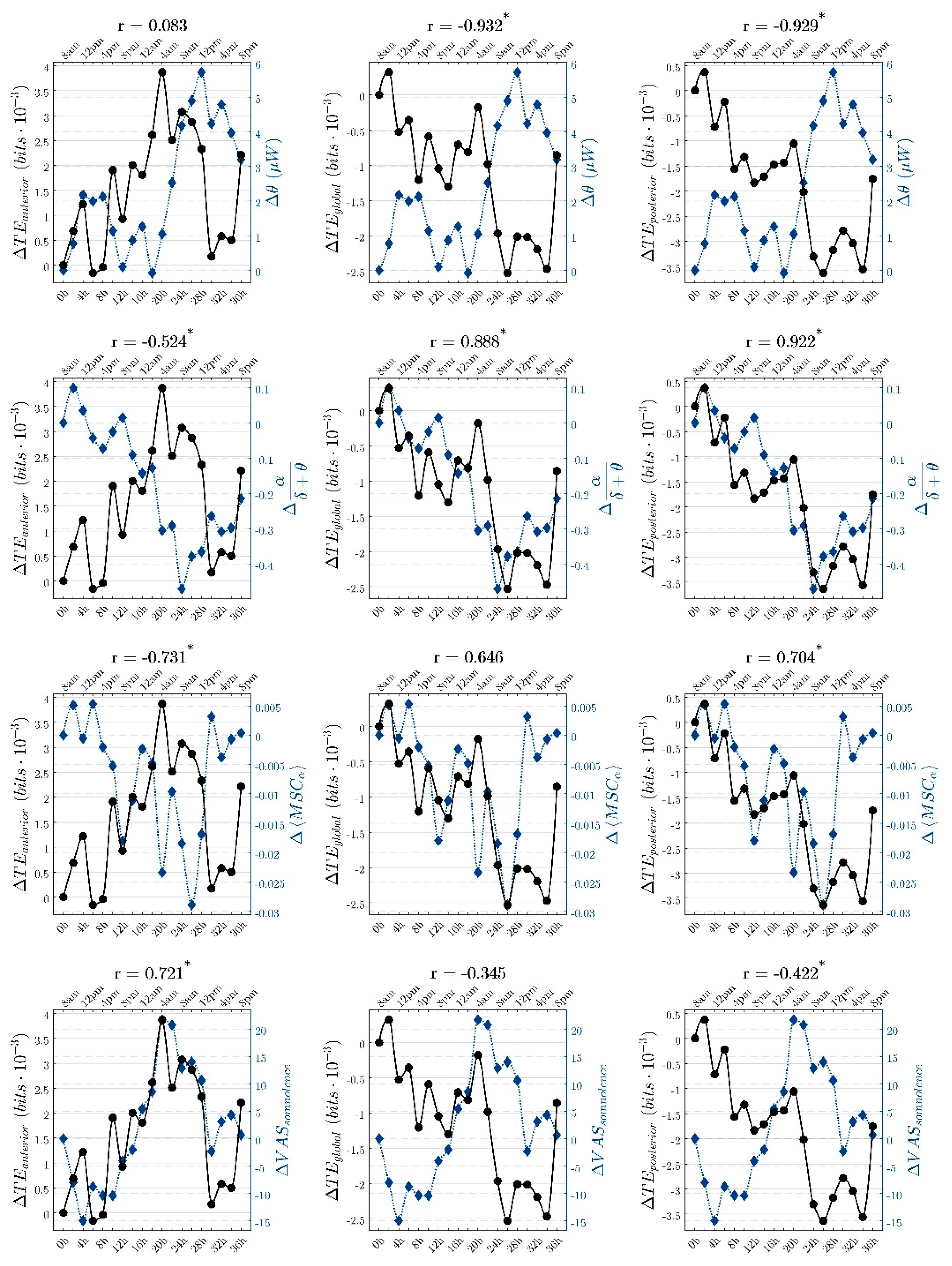
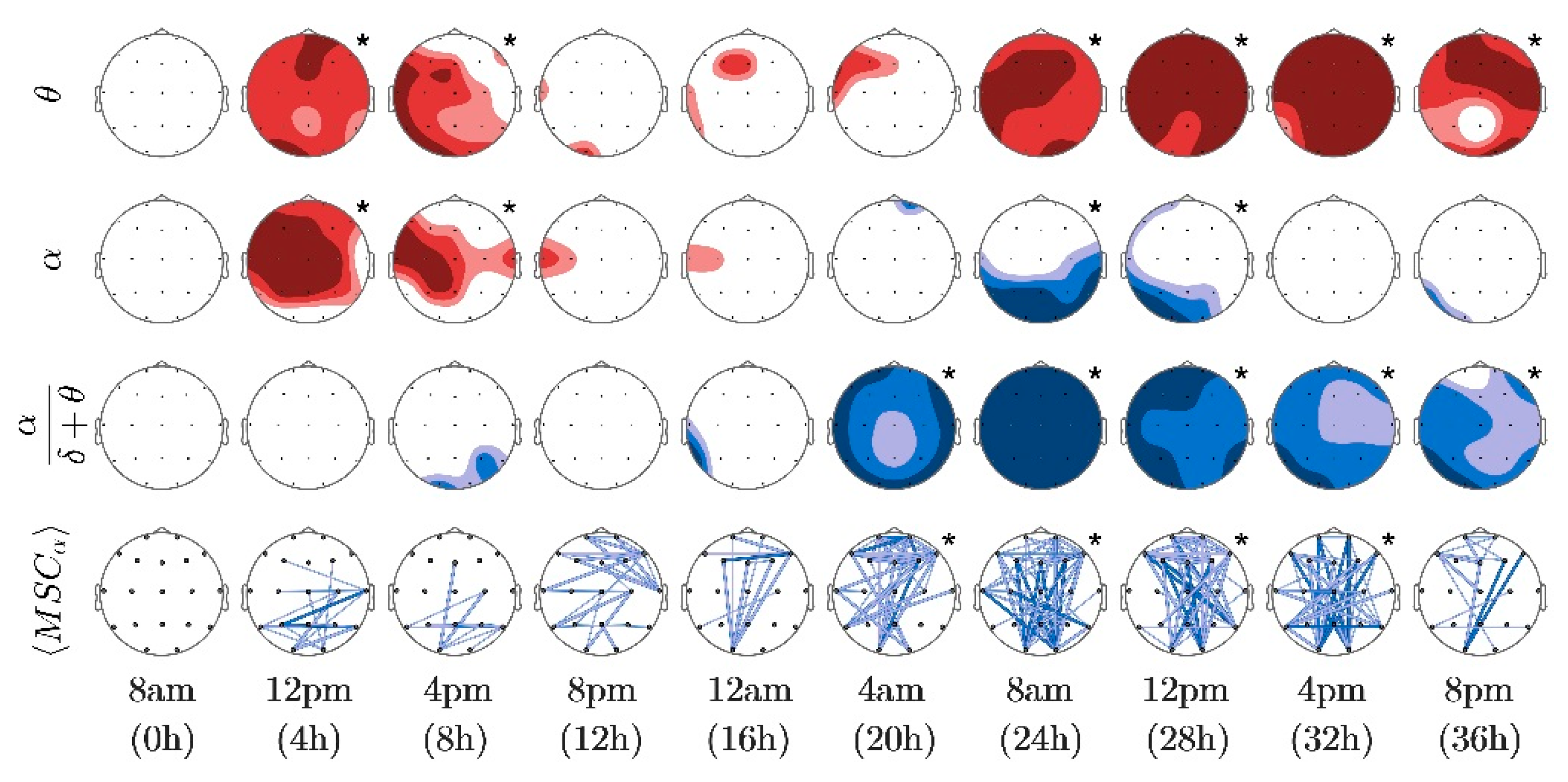
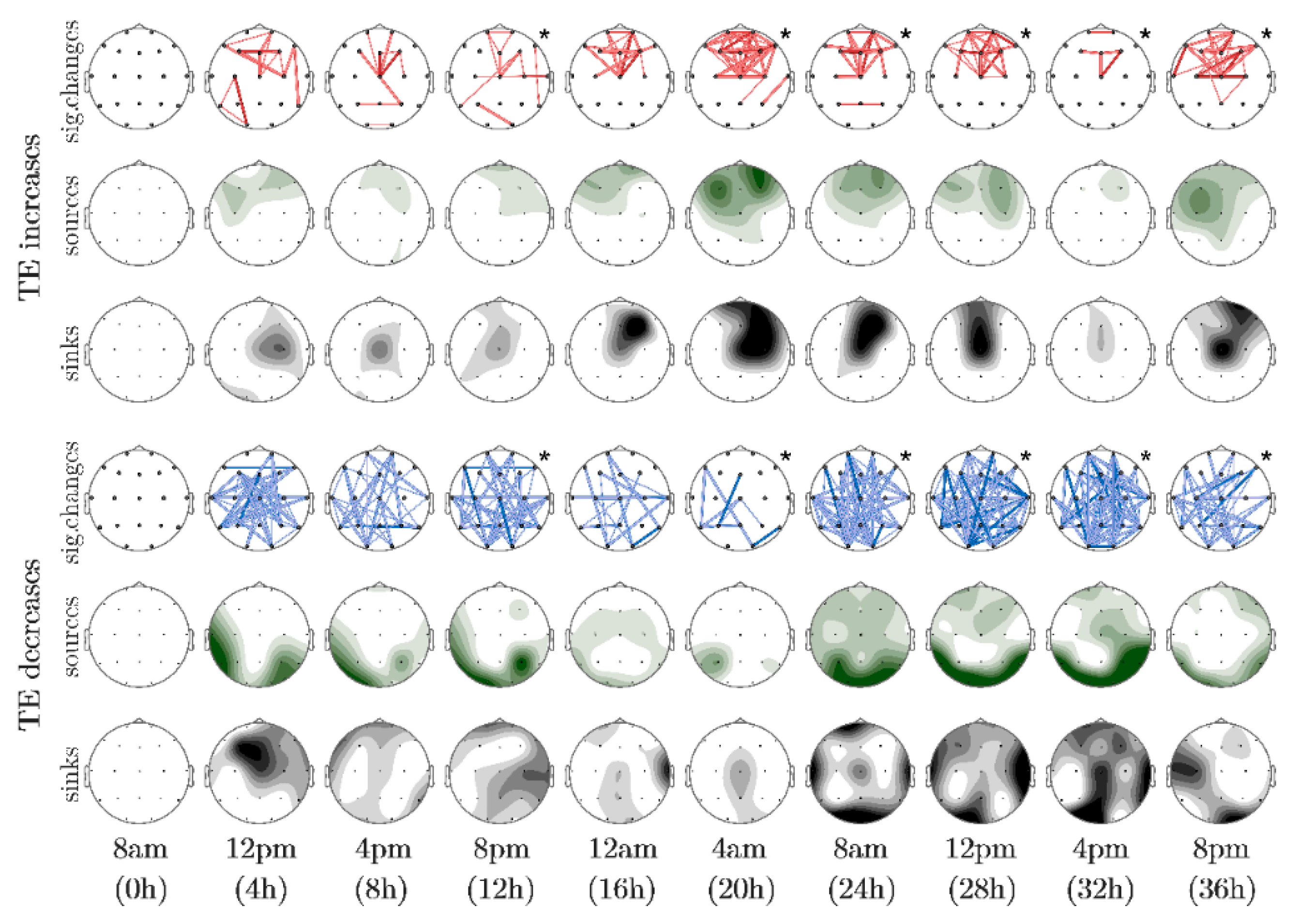
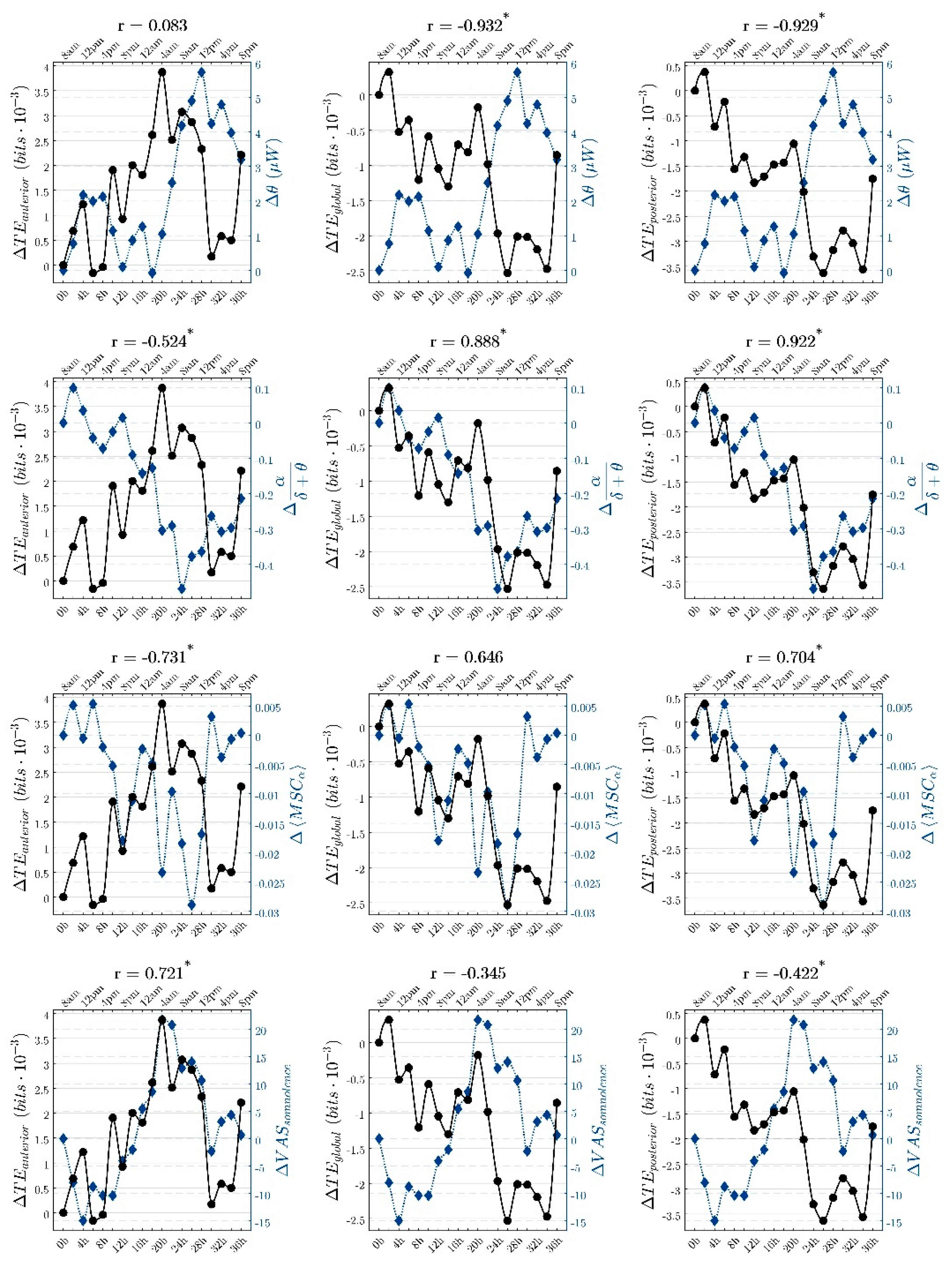
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ECG | Electrocardiogram |
| ECG | Electroencephalogram, electroencephalography, electroencephalographic |
| DMN | Default mode network |
| MSC | Magnitude squared coherence |
| P | probability value |
| SD | Sleep deprivation |
| TE | Transfer entropy |
References
- Alhola, P.; Polo-Kantola, P. Sleep deprivation: Impact on cognitive performance. Neuropsychiatr. Dis. Treat. 2007, 3, 553–567. [Google Scholar] [PubMed]
- Gast, H.; Schindler, K.; Rummel, C.; Herrmann, U.S.; Roth, C.; Hess, C.W.; Mathis, J. EEG correlation and power during maintenance of wakefulness test after sleep-deprivation. Clin. Neurophysiol. 2011, 122, 2025–2031. [Google Scholar] [CrossRef] [PubMed]
- Robillard, R.; Prince, F.; Boissonneault, M.; Filipini, D.; Carrier, J. Effects of increased homeostatic sleep pressure on postural control and their modulation by attentional resources. Clin. Neurophysiol. 2011, 122, 1771–1778. [Google Scholar] [CrossRef] [PubMed]
- Orzeł-Gryglewska, J. Consequences of sleep deprivation. Int. J. Occup. Med. Environ. Health 2010, 23, 95–114. [Google Scholar] [CrossRef] [PubMed]
- Borbély, A.A. A two process model of sleep regulation. Hum. Neurobiol. 1982, 1, 195–204. [Google Scholar] [PubMed]
- Borbély, A.A.; Baumann, F.; Brandeis, D.; Strauch, I.; Lehmann, D. Sleep deprivation: Effect on sleep stages and EEG power density in man. Electroencephalogr. Clin. Neurophysiol. 1981, 51, 483–493. [Google Scholar] [CrossRef]
- Aeschbach, D.; Matthews, J.R.; Postolache, T.T.; Jackson, M.A.; Giesen, H.A.; Wehr, T.A. Dynamics of the human EEG during prolonged wakefulness: Evidence for frequency-specific circadian and homeostatic influences. Neurosci. Lett. 1997, 239, 121–124. [Google Scholar] [CrossRef]
- Brunner, D.P.; Dijk, D.J.; Borbély, A.A. Repeated partial sleep deprivation progressively changes in EEG during sleep and wakefulness. Sleep 1993, 16, 100–113. [Google Scholar] [PubMed]
- Corsi-Cabrera, M.; Arce, C.; Ramos, J.; Lorenzo, I.; Guevara, M.A. Time course of reaction time and EEG while performing a vigilance task during total sleep deprivation. Sleep 1996, 19, 563–569. [Google Scholar] [PubMed]
- Ferreira, C.; Deslandes, A.; Moraes, H.; Cagy, M.; Pompeu, F.; Basile, L.F.; Piedade, R.; Ribeiro, P. Electroencephalographic changes after one night of sleep deprivation. Arq Neuropsiquiatr. 2006, 64, 388–393. [Google Scholar] [CrossRef] [PubMed]
- Strijkstra, A.M.; Beersma, D.G.M.; Drayer, B.; Halbesma, N.; Daan, S. Subjective sleepiness correlates negatively with global alpha (8–12 Hz) and positively with central frontal theta (4–8 Hz) frequencies in the human resting awake electroencephalogram. Neurosci. Lett. 2003, 340, 17–20. [Google Scholar] [CrossRef]
- Finelli, L.A.; Baumann, H.; Borbély, A.A.; Achermann, P. Dual electroencephalogram markers of human sleep homeostasis: Correlation between theta activity in waking and slow-wave activity in sleep. Neuroscience 2000, 101, 523–529. [Google Scholar] [CrossRef]
- Ehlen, J.C.; Jefferson, F.; Brager, A.J.; Benveniste, M.; Paul, K.N. Period-amplitude analysis reveals wake-dependent changes in the electroencephalogram during sleep deprivation. Sleep 2013, 36, 1723–1735. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Wyatt, J.; Czeisler, C.; Dijk, D. Separation of circadian and wake duration-dependent modulation of EEG activation during wakefulness. Neuroscience 2002, 114, 1047–1060. [Google Scholar] [CrossRef]
- Vyazovskiy, V.V.; Tobler, I. Theta activity in the waking EEG is a marker of sleep propensity in the rat. Brain Res. 2005, 1050, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Deboer, T.; Détári, L.; Meijer, J.H. Long term effects of sleep deprivation on the mammalian circadian pacemaker. Sleep 2007, 30, 257–262. [Google Scholar] [PubMed]
- Kar, S.; Routray, A.; Nayak, B.P. Functional network changes associated with sleep deprivation and fatigue during simulated driving: validation using blood biomarkers. Clin. Neurophysiol. 2011, 122, 966–974. [Google Scholar] [CrossRef] [PubMed]
- Kar, S.; Routray, A. Effect of sleep deprivation on functional connectivity of EEG channels. IEEE Trans. Syst. Man Cybern. Part A Syst. Hum. 2013, 43, 666–672. [Google Scholar] [CrossRef]
- Na, S.H.; Jin, S.-H.; Kim, S.Y. The effects of total sleep deprivation on brain functional organization: Mutual information analysis of waking human EEG. Int. J. Psychophysiol. 2006, 62, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Verweij, I.M.; Romeijn, N.; Smit, D.J.; Piantoni, G.; Van Someren, E.J.; van der Werf, Y.D. Sleep deprivation leads to a loss of functional connectivity in frontal brain regions. BMC Neurosci. 2014, 15. [Google Scholar] [CrossRef] [PubMed]
- Bosch, O.G.; Rihm, J.S.; Scheidegger, M.; Landolt, H.-P.; Stämpfli, P.; Brakowski, J.; Esposito, F.; Rasch, B.; Seifritz, E. Sleep deprivation increases dorsal nexus connectivity to the dorsolateral prefrontal cortex in humans. Proc. Natl. Acad. Sci. USA 2013, 110, 19597–19602. [Google Scholar] [CrossRef] [PubMed]
- Sämann, P.G.; Tully, C.; Spoormaker, V.I.; Wetter, T.C.; Holsboer, F.; Wehrle, R.; Czisch, M. Increased sleep pressure reduces resting state functional connectivity. Magn. Reson. Mater. Physics. Biol. Med. 2010, 23, 375–389. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Wang, L.; Ye, E.; Jin, X.; Ni, W.; Yang, Y.; Wen, B.; Hu, D.; Yang, Z. Decreased thalamocortical functional connectivity after 36 hours of total sleep deprivation: Evidence from resting state fMRI. PLoS ONE 2013, 8. [Google Scholar] [CrossRef]
- Cantero, J.L.; Atienza, M.; Salas, R.M.; Gómez, C.M. Alpha EEG coherence in different brain states: An electrophysiological index of the arousal level in human subjects. Neurosci. Lett. 1999, 271, 167–170. [Google Scholar] [CrossRef]
- Corsi-Cabrera, M.; Ramos, J.; Arce, C.; Guevara, M.A.; Ponce-de León, M.; Lorenzo, I. Changes in the waking EEG as a consequence of sleep and sleep deprivation. Sleep 1992, 15, 550–555. [Google Scholar] [PubMed]
- Koenis, M.M.G.; Romeijn, N.; Piantoni, G.; Verweij, I.; Van Der Werf, Y.D.; Van Someren, E.J.W.; Stam, C.J. Does sleep restore the topology of functional brain networks. Hum. Brain Mapp. 2013, 34, 487–500. [Google Scholar] [CrossRef] [PubMed]
- De Havas, J.A.; Parimal, S.; Soon, C.S.; Chee, M.W.L. Sleep deprivation reduces default mode network connectivity and anti-correlation during rest and task performance. Neuroimage 2012, 59, 1745–1751. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, T. Measuring information transfer. Phys. Rev. Lett. 2000, 85, 461–464. [Google Scholar] [CrossRef] [PubMed]
- Paluš, M.; Komárek, V.; Hrnčíř, Z.; Štěrbová, K. Synchronization as adjustment of information rates: Detection from bivariate time series. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2001, 63. [Google Scholar] [CrossRef] [PubMed]
- Wibral, M.; Vicente, R.; Lizier, J. Directed Information Measures in Neuroscience; Wibral, M., Vicente, R., Lizier, J.T., Eds.; Springer: Berlin, Germany, 2014. [Google Scholar]
- Lee, U.; Ku, S.; Noh, G.; Baek, S.; Choi, B.; Mashour, G.A. Disruption of Frontal-Parietal Communication by Ketamine, Propofol, and Sevoflurane. Anesthesiology 2013, 118, 1264–1275. [Google Scholar] [CrossRef] [PubMed]
- Untergehrer, G.; Jordan, D.; Kochs, E.F.; Ilg, R.; Schneider, G. Fronto-Parietal connectivity is a non-static phenomenon with characteristic changes during unconsciousness. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Blain-Moraes, S.; Mashour, G.A.; Lee, H.; Huggins, J.E.; Lee, U. Altered cortical communication in amyotrophic lateral sclerosis. Neurosci. Lett. 2013, 543, 172–176. [Google Scholar] [CrossRef] [PubMed]
- McBride, J.; Zhao, X.; Munro, N.; Jicha, G.; Smith, C.; Jiang, Y. Discrimination of mild cognitive impairment and alzheimer’s disease using transfer entropy measures of scalp EEG. J. Healthc. Eng. 2015, 6, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Blain-Moraes, S.; Lee, U.; Ku, S.; Noh, G.; Mashour, G.A. Electroencephalographic effects of ketamine on power, cross-frequency coupling, and connectivity in the alpha bandwidth. Front. Syst. Neurosci. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.F.; Romero, S.; Mananas, M.A.; Riba, J. Serotonergic psychedelics temporarily modify information transfer in humans. Int. J. Neuropsychopharmacol. 2015, 18, pyv039–pyv039. [Google Scholar] [CrossRef] [PubMed]
- Babkoff, H.; Caspy, T.; Mikulincer, M. Subjective sleepiness ratings: The effects of sleep deprivation, circadian rhythmicity and cognitive performance. Sleep 1991, 14, 534–539. [Google Scholar] [PubMed]
- Curcio, G.; Casagrande, M.; Bertini, M. Sleepiness: Evaluating and quantifying methods. Int. J. Psychophysiol. 2001, 41, 251–263. [Google Scholar] [CrossRef]
- Johns, M.W. The subjective measurement of excessive daytime sleepiness. In Sleep Disorders: Diagnosis and Therapeutics; Pandi-Perumal, S.R., Verster, J.C., Monti, J., Lader, M., Langer, S.Z., Eds.; CRC Press: London, UK, 2008; pp. 643–657. [Google Scholar]
- Belouchrani, A.; Abed-Meraim, K.; Cardoso, J.F.; Moulines, E. A blind source separation technique using second-order statistics. IEEE Trans. Signal Process. 1997, 45, 434–444. [Google Scholar] [CrossRef]
- Romero, S.; Mañanas, M.A.; Barbanoj, M.J. A comparative study of automatic techniques for ocular artifact reduction in spontaneous EEG signals based on clinical target variables: a simulation case. Comput. Biol. Med. 2008, 38, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Anderer, P.; Semlitsch, H.V.; Saletu, B.; Barbanoj, M.J. Artifact processing in topographic mapping of electroencephalographic activity in neuropsychopharmacology. Psychiatry Res. 1992, 45, 79–93. [Google Scholar] [CrossRef]
- Oken, B.S.; Salinsky, M.C.; Elsas, S.M. Vigilance, alertness, or sustained attention: physiological basis and measurement. Clin. Neurophysiol. 2006, 117, 1885–1901. [Google Scholar] [CrossRef] [PubMed]
- Olbrich, E.; Landolt, H.P.; Achermann, P. Effect of prolonged wakefulness on electroencephalographic oscillatory activity during sleep. J. Sleep Res. 2014, 23, 255–262. [Google Scholar] [CrossRef] [PubMed]
- Matousek, M.; Petersén, I. A method for assessing alertness fluctuations from EEG spectra. Electroencephalogr. Clin. Neurophysiol. 1983, 55, 108–113. [Google Scholar] [CrossRef]
- Vejmelka, M.; Paluš, M. Inferring the directionality of coupling with conditional mutual information. Phys. Rev. E Stat. Nonlinear. Soft Matter Phys. 2008, 77, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Nemati, S.; Silva, I.; Edwards, B.A.; Butler, J.P.; Malhotra, A. Transfer entropy estimation and directional coupling change detection in biomedical time series. Biomed. Eng. Online 2012, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Cross, E.M.; Chaffin, W.W. Use of the binomial theorem in interpreting results of multiple tests of Significance. Educ. Psychol. Meas. 1982, 42, 25–34. [Google Scholar] [CrossRef]
- Saletu, M.T.; Anderer, P.; Saletu-Zyhlarz, G.M.; Mandl, M.; Arnold, O.; Nosiska, D.; Zeitlhofer, J.; Saletu, B. EEG-mapping differences between narcolepsy patients and controls and subsequent double-blind, placebo-controlled studies with modafinil. Eur. Arch. Psychiatry Clin. Neurosci. 2005, 255, 20–32. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.F.; Mañanas, M.A.; Romero, S.; Hoyer, D.; Riba, J.; Barbanoj, M.J. Drug effect on EEG connectivity assessed by linear and nonlinear couplings. Hum. Brain Mapp. 2010, 31, 487–497. [Google Scholar] [CrossRef] [PubMed]
- Alonso, J.F.; Romero, S.; Ballester, M.R.; Antonijoan, R.M.; Mañanas, M.A. Stress assessment based on EEG univariate features and functional connectivity measures. Physiol. Meas. 2015, 36, 1351–1365. [Google Scholar] [CrossRef] [PubMed]
- Painold, A.; Anderer, P.; Holl, A.K.; Letmaier, M.; Saletu-Zyhlarz, G.M.; Saletu, B.; Bonelli, R.M. EEG low-resolution brain electromagnetic tomography (LORETA) in Huntington’s disease. J. Neurol. 2011, 258, 840–854. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Khalsa, S.B.; Wyatt, J.K.; Czeisler, C.A.; Dijk, D.J. EEG and ocular correlates of circadian melatonin phase and human performance decrements during sleep loss. Am. J. Physiol. 1999, 277, R640–R649. [Google Scholar] [PubMed]
- Cantero, J.L.; Atienza, M.; Salas, R.M. State-modulation of cortico-cortical connections underlying normal EEG alpha variants. Physiol. Behav. 2000, 71, 107–115. [Google Scholar] [CrossRef]
- Münch, M.; Knoblauch, V.; Blatter, K.; Schröder, C.; Schnitzler, C.; Kräuchi, K.; Wirz-Justice, A.; Cajochen, C. The frontal predominance in human EEG delta activity after sleep loss decreases with age. Eur. J. Neurosci. 2004, 20, 1402–1410. [Google Scholar] [CrossRef] [PubMed]
- Cajochen, C.; Dijk, D. Electroencephalographic activity during wakefulness, rapid eye movement and non-rapid eye movement sleep in humans: Comparison of their circadian and. Sleep Biol. Rhythm. 2003, 85–96. [Google Scholar] [CrossRef]
- Durmer, J.S.; Dinges, D.F. Neurocognitive consequences of sleep deprivation. Semin. Neurol. 2005, 25, 320–339. [Google Scholar] [CrossRef] [PubMed]
- Drummond, S.P.A.; Gillin, J.C.; Brown, G.G. Increased cerebral response during a divided attention task following sleep deprivation. J. Sleep Res. 2001, 10, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Warden, M.R.; Miller, E.K. Task-dependent changes in short-term memory in the prefrontal cortex. J. Neurosci. 2010, 30, 15801–15810. [Google Scholar] [CrossRef] [PubMed]
- Gujar, N.; Yoo, S.-S.; Hu, P.; Walker, M.P. The unrested resting brain: sleep deprivation alters activity within the default-mode network. J. Cogn. Neurosci. 2010, 22, 1637–1648. [Google Scholar] [CrossRef] [PubMed]
- Drummond, S.P.; Brown, G.G.; Gillin, J.C.; Stricker, J.L.; Wong, E.C.; Buxton, R.B. Altered brain response to verbal learning following sleep deprivation. Nature 2000, 403, 655–657. [Google Scholar] [PubMed]
- Horovitz, S.G.; Braun, A.R.; Carr, W.S.; Picchioni, D.; Balkin, T.J.; Fukunaga, M.; Duyn, J.H. Decoupling of the brain’s default mode network during deep sleep. Proc. Natl. Acad. Sci. USA 2009, 106, 11376–11381. [Google Scholar] [CrossRef] [PubMed]
- Benedetti, F.; Bernasconi, A.; Blasi, V.; Cadioli, M.; Colombo, C.; Falini, A.; Lorenzi, C.; Radaelli, D.; Scotti, G.; Smeraldi, E. Neural and genetic correlates of Antidepressant response to sleep deprivation. Arch. Gen. Psychiatry 2007, 64, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.C.; Gillin, J.C.; Buchsbaum, M.S.; Schachat, C.; Darnall, L.A.; Keator, D.B.; Fallon, J.H.; Bunney, W.E. Sleep deprivation PET correlations of Hamilton symptom improvement ratings with changes in relative glucose metabolism in patients with depression. J. Affect. Disord. 2008, 107, 181–186. [Google Scholar] [CrossRef] [PubMed]
- Marinazzo, D.; Gosseries, O.; Boly, M.; Ledoux, D.; Rosanova, M.; Massimini, M.; Noirhomme, Q.; Laureys, S. Directed information transfer in scalp electroencephalographic recordings: Insights on disorders of consciousness. Clin. EEG Neurosci. 2014, 45, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Zubler, F.; Gast, H.; Abela, E.; Rummel, C.; Hauf, M.; Wiest, R.; Pollo, C.; Schindler, K. Detecting functional hubs of ictogenic networks. Brain Topogr. 2015, 28, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, H.; Wang, Y.; Lei, X. Enhanced brain small-worldness after sleep deprivation: A compensatory effect. J. Sleep Res. 2014, 23, 554–563. [Google Scholar] [CrossRef] [PubMed]
- Aeschbach, D.; Matthews, J.R.; Postolache, T.T.; Jackson, M.A.; Giesen, H.A.; Wehr, T.A. Two circadian rhythms in the human electroencephalogram during wakefulness. Am. J. Physiol. 1999, 277, R1771–R1779. [Google Scholar] [PubMed]
- Scheeringa, R.; Bastiaansen, M.C.M.; Petersson, K.M.; Oostenveld, R.; Norris, D.G.; Hagoort, P. Frontal theta EEG activity correlates negatively with the default mode network in resting state. Int. J. Psychophysiol. 2008, 67, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Niessing, J.; Ebisch, B.; Schmidt, K.E.; Niessing, M.; Singer, W.; Galuske, R.A.W. Hemodynamic signals correlate tightly with synchronized gamma oscillations. Science 2005, 309, 948–951. [Google Scholar] [CrossRef] [PubMed]
- Ward, A.M.; McLaren, D.G.; Schultz, A.P.; Chhatwal, J.; Boot, B.P.; Hedden, T.; Sperling, R.A. Daytime sleepiness is associated with decreased default mode network connectivity in both young and cognitively intact elderly subjects. Sleep 2013, 36, 1609–1615. [Google Scholar] [CrossRef] [PubMed]
- Fisher, S.P.; Vyazovskiy, V.V. Local sleep taking care of high-maintenance cortical circuits under sleep restriction. Sleep 2014, 37, 1727–1730. [Google Scholar] [CrossRef] [PubMed]
- Poudel, G.R.; Innes, C.R.H.; Jones, R.D. Cerebral perfusion differences between drowsy and nondrowsy individuals after acute sleep restriction. Sleep 2012, 35, 1085–1096. [Google Scholar] [CrossRef] [PubMed]
- Lückmann, H.C.; Jacobs, H.I.L.; Sack, A.T. The cross-functional role of frontoparietal regions in cognition: Internal attention as the overarching mechanism. Prog. Neurobiol. 2014, 116, 66–86. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Probability Value (p) | Color Intensityof the Electrode | Color and Thickessof the Connection |
|---|---|---|
| p ≤ 0.01 | dark | dark, thick |
| 0.01 < p ≤ 0.05 | medium | light, thick |
| 0.05 < p ≤ 0.10 | light | light thin |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alonso, J.F.; Romero, S.; Mañanas, M.A.; Alcalá, M.; Antonijoan, R.M.; Giménez, S. Acute Sleep Deprivation Induces a Local Brain Transfer Information Increase in the Frontal Cortex in a Widespread Decrease Context. Sensors 2016, 16, 540. https://doi.org/10.3390/s16040540
Alonso JF, Romero S, Mañanas MA, Alcalá M, Antonijoan RM, Giménez S. Acute Sleep Deprivation Induces a Local Brain Transfer Information Increase in the Frontal Cortex in a Widespread Decrease Context. Sensors. 2016; 16(4):540. https://doi.org/10.3390/s16040540
Chicago/Turabian StyleAlonso, Joan F., Sergio Romero, Miguel A. Mañanas, Marta Alcalá, Rosa M. Antonijoan, and Sandra Giménez. 2016. "Acute Sleep Deprivation Induces a Local Brain Transfer Information Increase in the Frontal Cortex in a Widespread Decrease Context" Sensors 16, no. 4: 540. https://doi.org/10.3390/s16040540
APA StyleAlonso, J. F., Romero, S., Mañanas, M. A., Alcalá, M., Antonijoan, R. M., & Giménez, S. (2016). Acute Sleep Deprivation Induces a Local Brain Transfer Information Increase in the Frontal Cortex in a Widespread Decrease Context. Sensors, 16(4), 540. https://doi.org/10.3390/s16040540