Invasion of Eastern Texas Forestlands by Chinese Privet: Efficacy of Alternative Management Strategies



Abstract
:1. Introduction
2. Methods
2.1. Study Area and Data Sources
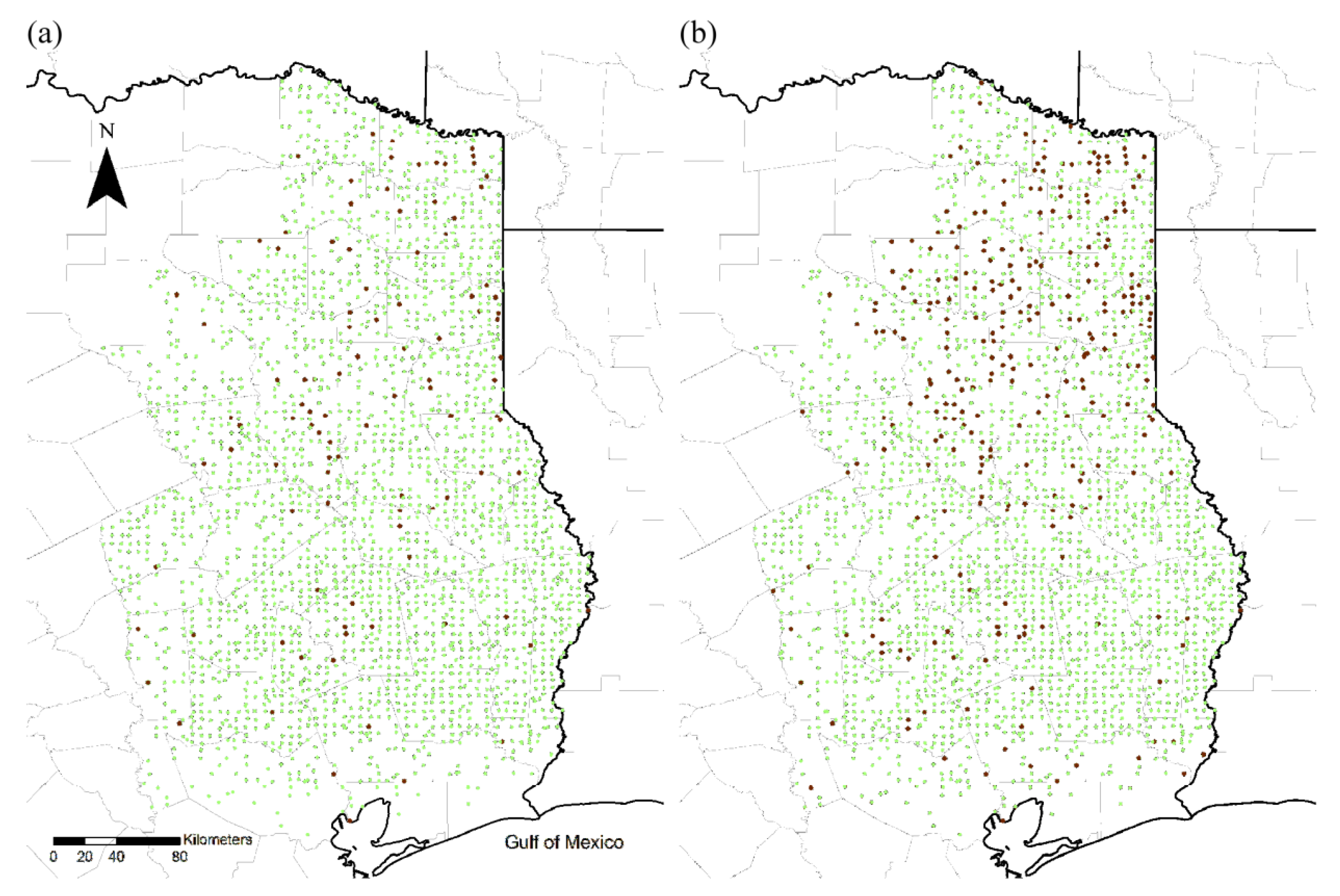
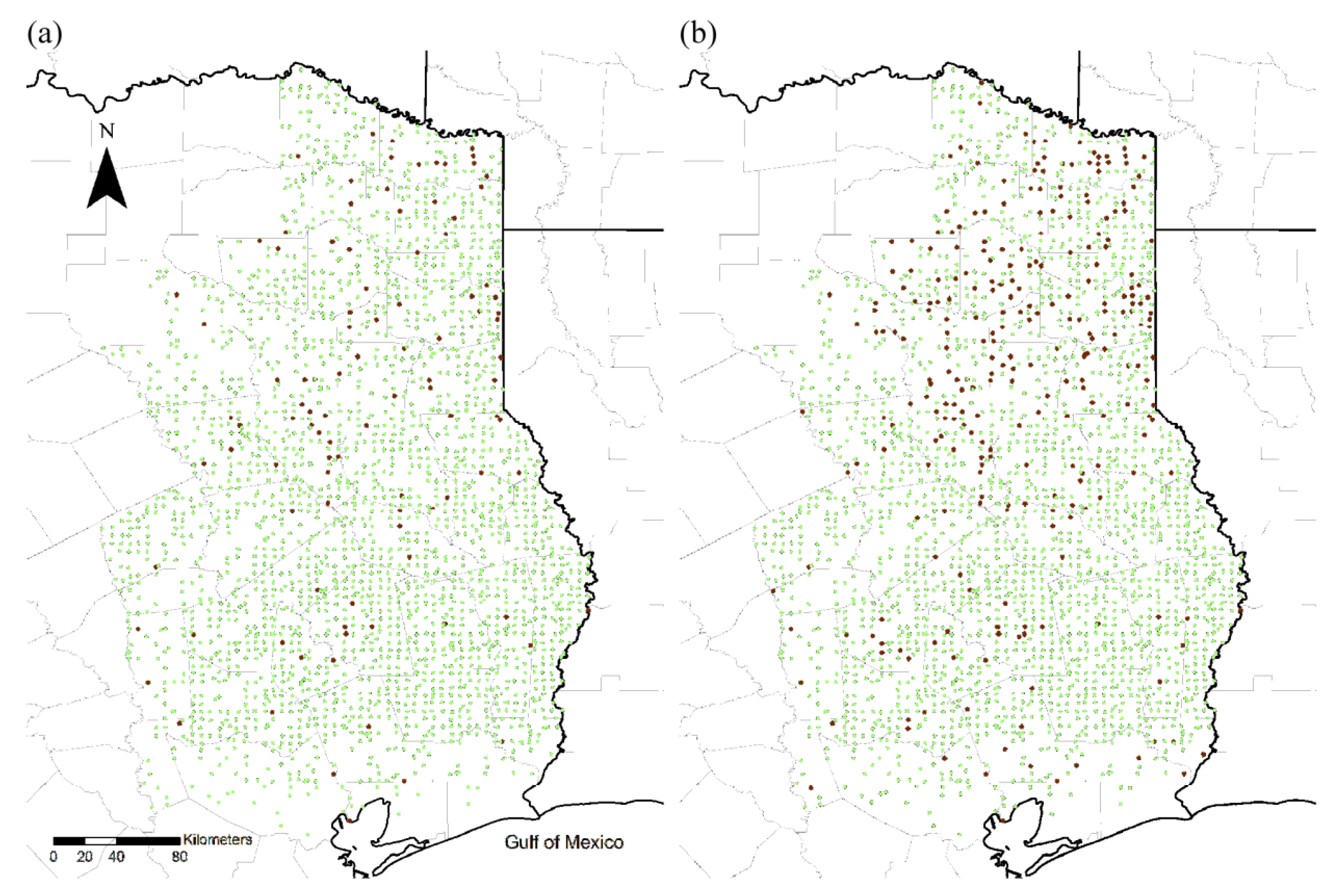
2.2. Quantification Spread
2.3. Identification of Invasion Determinants

{kind=link}
{kind=link}
{kind=link}
| Variable | Value or Unit of Measure | Mean (Range) or Frequency |
|---|---|---|
| Landscape conditions | ||
| Elevation | m | 259.36 (−79 ~ 999) |
| Slope | degree | 2.07 (0 ~ 32.5) |
| Adjacency to water bodies within 300 m | 0: no 1:yes | 0: 1924 1: 473 |
| Forest features | ||
| Stand age | year | 35.73 (1 ~ 104) |
| Site productivity | 1: 0–1.39 2: 1.40–3.49 3: 3.50–5.94 4: 5.95–8.39 5: 8.40–11.54 6: 11.55–15.74 7: >15.74 m3ha-1year-1 | 1: 9 2: 154 3: 661 4: 933 5: 541 6: 95 7: 4 |
| Forest management activities | ||
| Site preparation a | 0: no 1:yes | 0: 2312 1: 85 |
| Artificial regeneration a | 0: no 1:yes | 0: 1712 1: 685 |
| Natural regeneration a | 0: no 1:yes | 0: 2351 1: 46 |
| Disturbances | ||
| Distance to the nearest road | 1: <30 2: 30–91 3: 92–152 4: 153–305 5: 306–805 6: 806–1609 7: 1610–4828 8: 4829–8047 9: >8047 m | 1: 179 2: 394 3: 283 4: 508 5: 685 6: 253 7: 72 8: 13 9: 10 |
| Fire disturbance a,b | 0: no 1:yes | 0: 2381 1: 16 |
| Animal disturbance a,b | 0: no 1:yes | 0: 2384 1: 13 |
| Wind disturbance a,b | 0: no 1:yes | 0: 2329 1: 68 |
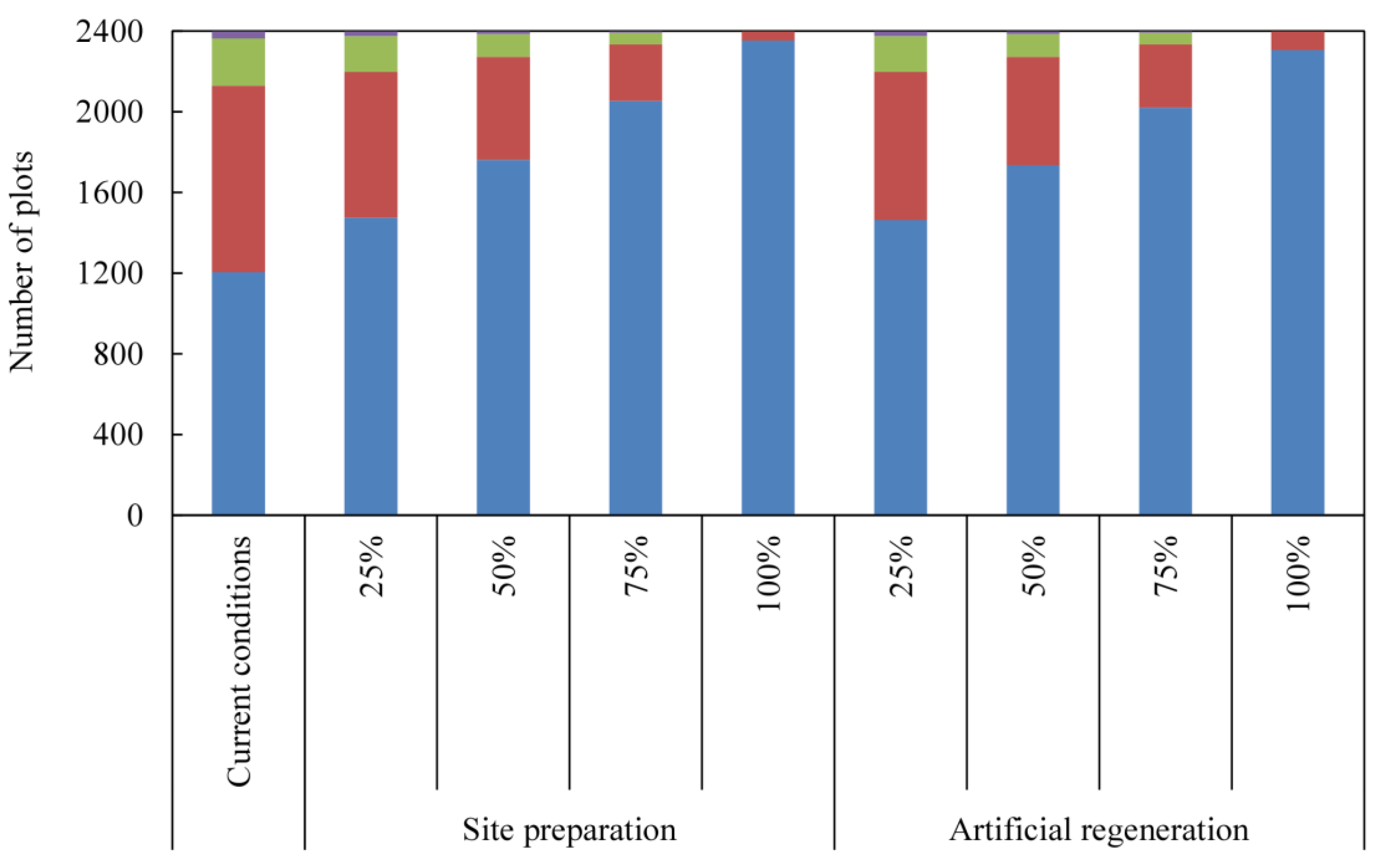
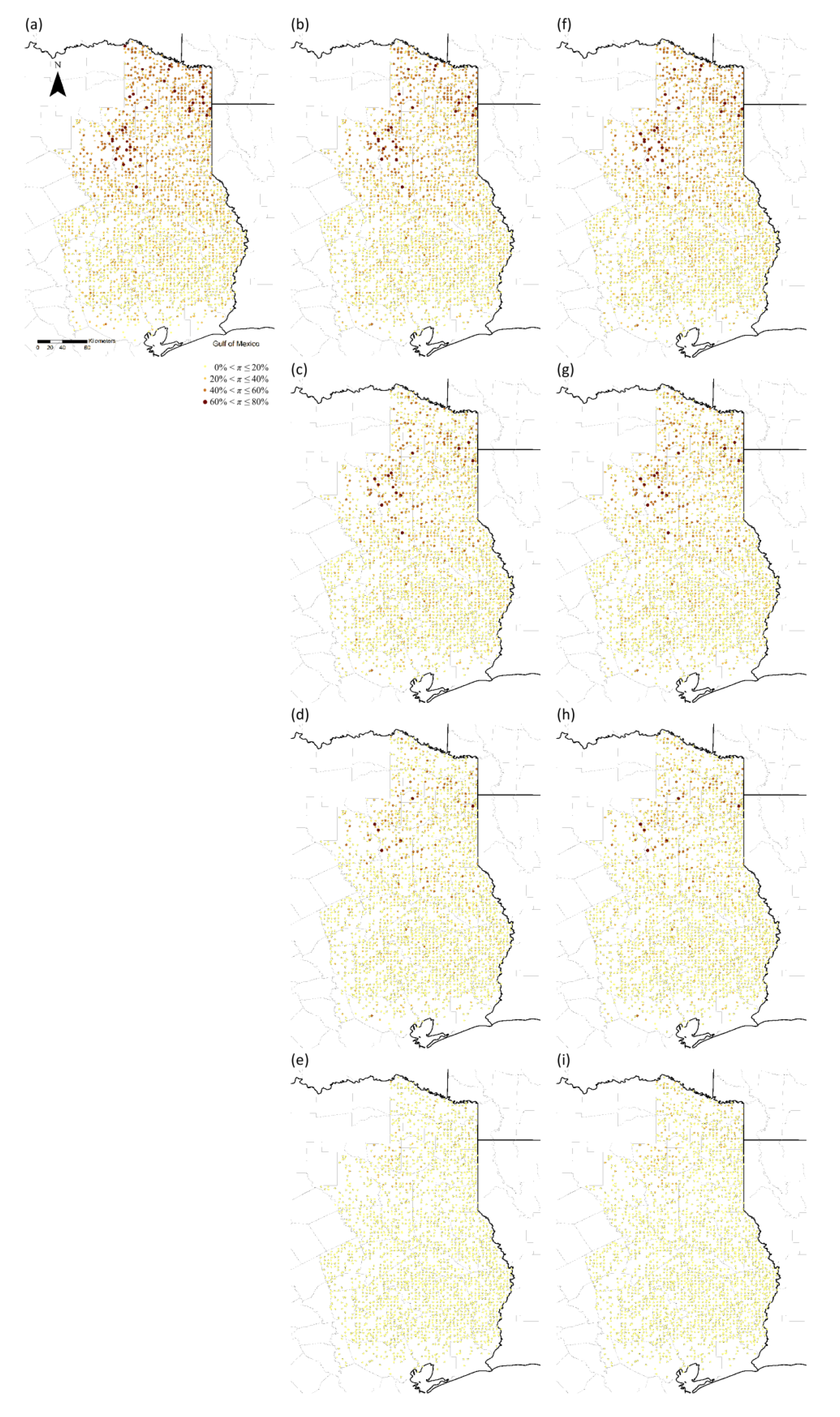
2.4. Probabilities of Further Invasion
3. Results
| Variable | Estimated Coefficient | Estimated Standard Error a | Estimated Odds Ratio b |
|---|---|---|---|
| Landscape conditions | |||
| Elevation | 0.0023 | 0.0006 | 1.0023 |
| Adjacency to water bodies within 300 m | 0.4746 | 0.1675 | 1.6073 |
| Forest features | |||
| Stand age | −0.0179 | 0.0037 | 0.9823 |
| Site productivity | 0.3144 | 0.0747 | 1.3694 |
| Forest management activities | |||
| Site preparation | −0.8501 | 0.3916 | 0.4273 |
| Artificial regeneration | −0.6436 | 0.1732 | 0.5254 |
| Disturbances | |||
| Distance to the nearest road | −0.1306 | 0.0436 | 0.8776 |
| Constant | 1.7972 | 0.8683 | — |
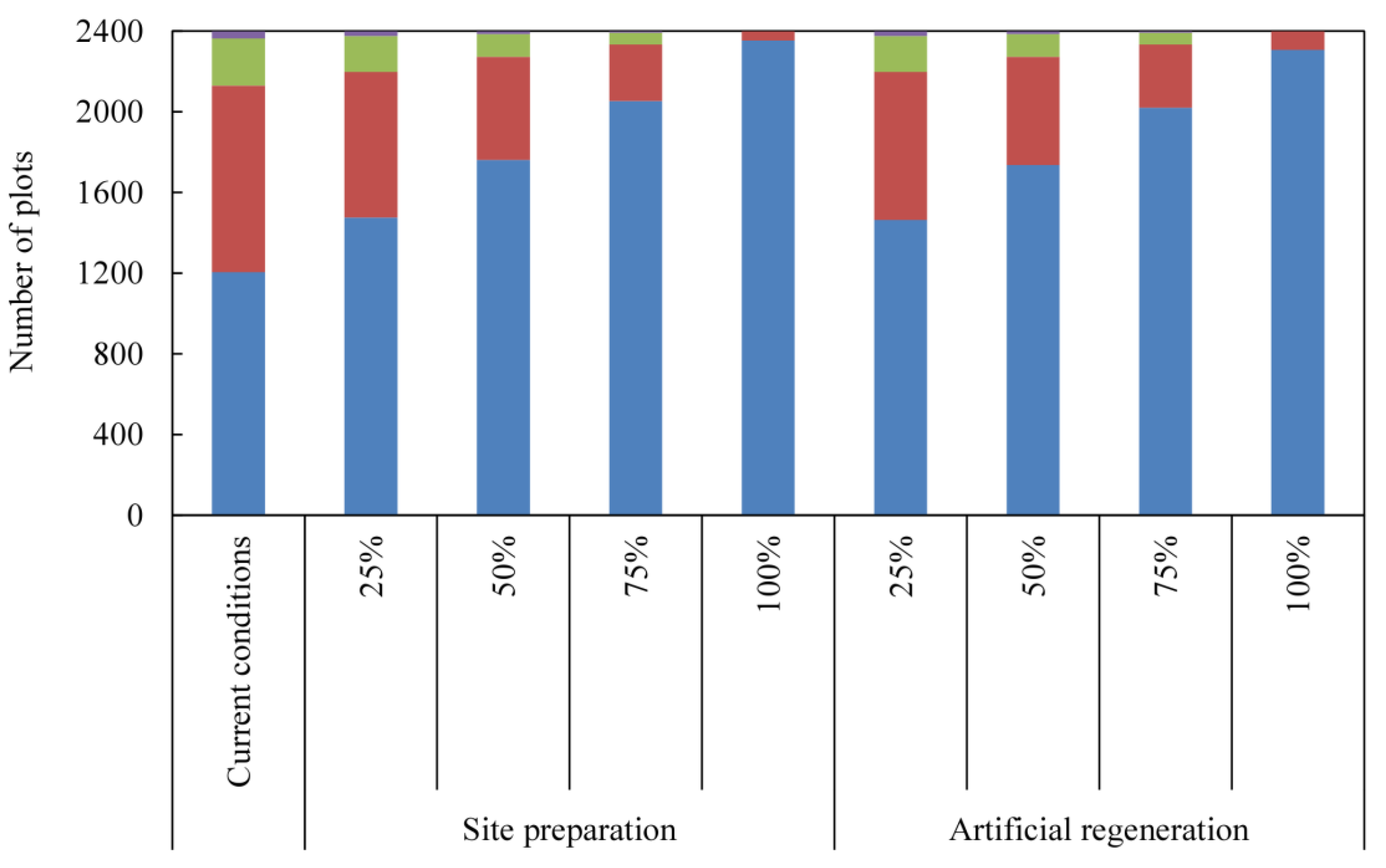
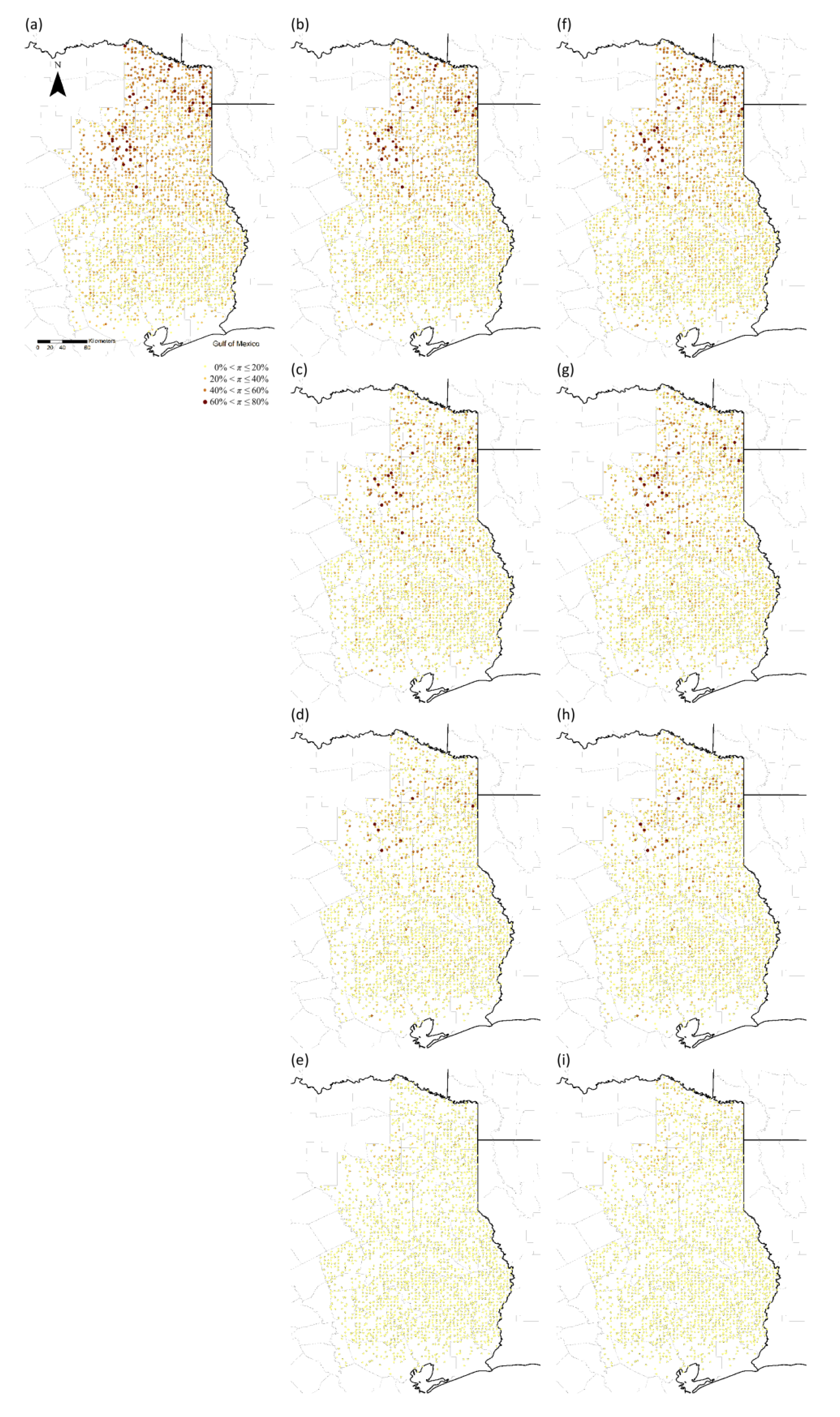
4. Discussion
Author Contributions
Conflicts of Interest
References
- USDA. Southern Nonnative Invasive Plant Data Extraction Tool (SNIPET). Available online: http://srsfia2.fs.fed.us/SNIPET/ (accessed on 1 October 2014).
- Camilli, K.S. Chinese and European pri4vet: A Threat to Texas’ Forests. Sixth of the “Dirty Dozen”. Available online: http://texasinvasives.org/resources/publications/06_Privet_TFA.pdf (accessed on 1 October 2014).
- Miller, J.H. Nonnative Invasive Plants of Southern Forests: A Field Guide for Identification and Control; Eastern Forest Environmental Threat Assessment Center: Asheville, NC, USA, 2003; p. 11. [Google Scholar]
- Merriam, R.W.; Feil, E. The potential impact of an introduced shrub on native plant diversity and forest regeneration. Biol. Invasions 2002, 4, 369–373. [Google Scholar] [CrossRef]
- Hanula, J.L.; Horn, S.; Taylor, J.W. Chinese privet (Ligustrum sinense) removal and its effect on native plant communities of riparian forests. Invasive Plant Sci. Manag. 2009, 2, 292–300. [Google Scholar] [CrossRef]
- Wilcox, J.; Beck, C.W. Effects of Ligustrum sinense Lour. (Chinese privet) on abundance and diversity of songbirds and native plants in a southeastern nature preserve. Southeast. Nat. 2007, 6, 535–550. [Google Scholar]
- Hanula, J.L.; Horn, S. Removing an invasive shrub (Chinese privet) increases native bee diversity and abundance in riparian forests of the southeastern United States. Insect Conserv. Divers. 2011, 4, 275–283. [Google Scholar] [CrossRef]
- Lobe, J.W.; Callaham, M.A., Jr.; Hendrix, P.F.; Hanula, J.L. Removal of an invasive shrub (Chinese privet: Ligustrum sinense Lour.) reduces exotic earthworm abundance and promotes recovery of native North American earthworms. Appl. Soil Ecol. 2014, in press. [Google Scholar]
- Ulyshen, M.D.; Horn, S.; Hanula, J.L. Response of beetles (Coleoptera) at three heights to the experimental removal of an invasive shrub, Chinese privet (Ligustrum sinense), from floodplain forests. Biol. Invasions 2010, 12, 1573–1579. [Google Scholar] [CrossRef]
- Brown, C.E.; Pezeshki, S.R. A study on waterlogging as a potential tool to control Ligustrum sinense populations in western Tennessee. Wetlands 2000, 20, 429–437. [Google Scholar] [CrossRef]
- Miller, J.H. Nonnative Invasive Plants of Southern Forests: A Field Guide for Identification and Control; USDA Forest Service, Southern Research Station: Asheville, NC, USA, 2003. [Google Scholar]
- Dirr, M.A. Manual of Woody Landscape Plants: Their Identification, Ornamental Characteristics, Culture, Propagation and Uses, 5th ed.; Stipes Publishing, LLC: Champaign, IL, USA, 1998; p. 1250. [Google Scholar]
- Young, J.A.; Young, C.G. Seeds of Woody Plants in North America; Timberpress Press Inc.: Portland, OR, USA, 1992. [Google Scholar]
- Rossell, C.R.; Patch, S.; Salmons, S. Effects of deer browsing on native and non-native vegetation in a mixed oak-beech forest on the Atlantic coastal plain. Northeast. Nat. 2007, 14, 61–72. [Google Scholar] [CrossRef]
- Christopher, C.C.; Barrett, G.W. Coexistence of white-footed mice (Peromyscus leucopus) and golden mice (Ochrotomys nuttalli) in a southeastern forest. J. Mammal. 2006, 87, 102–107. [Google Scholar] [CrossRef]
- USDA. The Forest Inventory and Analysis Database: Database Description and Users Manual Version 5.1; U.S. Department of Agriculture Forest Service: Arlington, VA, USA, 2011; pp. 12–15. [Google Scholar]
- Lodge, D.M.; Williams, S.; Macisaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: Recommendations for U.S. Policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar]
- Kronrad, G.D.; Huang, C.-H. Economic analysis of pruning and low-density management compared to traditional management of loblolly pine plantations in east Texas. South. J. Appl. For. 2004, 28, 12–20. [Google Scholar]
- Wang, H.-H.; Grant, W.E. Determinants of Chinese and European privet (Ligustrum sinense and Ligustrum vulgare) invasion and likelihood of further invasion in southern U.S. Forestlands. Invasive Plant Sci. Manag. 2012, 5, 454–463. [Google Scholar] [CrossRef]
- Streng, D.R.; Glitzenstein, J.S.; Harcombe, P.A. Woody seedling dynamics in an east Texas floodplain forest. Ecol. Monogr. 1989, 59, 177–204. [Google Scholar] [CrossRef]
- USDA. FIA Data and Tools. Available online: http://www.fia.fs.fed.us/tools-data/ (accessed on 1 October 2014).
- Bechtold, W.A.; Patterson, P.L. The Enhanced Forest Inventory and Analysis Program: National Sampling Design and Estimation Procedures; Gen. Tech. Rep. Srs-80; Southern Research Station, Forest Service, U.S. Department of agriculture: Asheville, NC, USA, 2005; p. 14. [Google Scholar]
- Rudis, V.A.; Gray, A.; McWilliams, W.; O’Brien, R.; Olson, C.; Oswalt, S.; Schulz, B. Regional Monitoring of Nonnative Plant Invasions with the Forest Inventory and Analysis Program. In Proceedings of the Sixth Annual FIA Symposium, Denver, CO, USA, 21–24 September 2006; McRoberts, R.E., Reams, G.A., Deusen, P.C.V., McWilliams, W.H., Eds.; USDA Forest Service: Denver, CO, USA; pp. 49–64.
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed.; John Wiley and Sons, Inc.: Hoboken, NJ, USA, 2007. [Google Scholar]
- Akaike, H. Information theory and an extension of the maximum likelihood principle. In Second International Symposium on Information Theory; Kotz, S., Johnson, N.L., Eds.; Akadémiai Kiadó: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied logistic regression. In Wiley Series in Probability and Statistics. Texts and References Section; John Wiley and Sons, Inc.: New York, NY, USA, 2000; pp. 147–156. [Google Scholar]
- Conner, W.; Stanturf, J.A.; Gardiner, E.S.; Schweitzer, C.J.; Ezell, A.W. Recognizing and overcoming difficult site conditions for afforestation of bottomland hardwoods. Ecol. Restor. 2004, 22, 183–193. [Google Scholar] [CrossRef]
- Merriam, R.W. The abundance, distribution and edge associations of six non-indigenous, harmful plants across North Carolina. J. Torrey Bot. Soc. 2003, 130, 283–291. [Google Scholar] [CrossRef]
- Stone, S.L. Privet removed from Austin nature preserves. Restor. Manag. Notes 1997, 15, 93–94. [Google Scholar]
- Greipsson, S.; DiTommaso, A. Invasive non-native plants alter the occurrence of arbuscular mycorrhizal fungi and benefit from this association. Ecol. Restor. 2006, 24, 236–241. [Google Scholar] [CrossRef]
- Mitchell, J.D.; Lockaby, B.G.; Brantley, E.F. Influence of Chinese privet (Ligustrum sinense) on decomposition and nutrient availability in riparian forests. Invasive Plant Sci. Manag. 2011, 4, 437–447. [Google Scholar] [CrossRef]
- Grove, E.; Clarkson, B.D. An Ecological Study of Chinese Privet (Ligustrum sinense Lour.) in the Waikato Region; University of Waikato: Hamilton, New Zealand, 2005. [Google Scholar]
- Wang, H.-H. Occupation, Dispersal, and Economic Impact of Major Invasive Plant Species in Southern U.S. Forests. Ph.D. Dissertation, Texas A&M University, College Station, TX, USA, 2009. [Google Scholar]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology; John Wiley and Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Wang, Z.; Nyland, R.D. Changes in the condition and species composition of developing even-aged northern hardwood stands in central New York. Northern J. Appl. For. 1996, 13, 189–194. [Google Scholar]
- Wang, H.-H.; Wonkka, C.L.; Grant, W.E.; Rogers, W.E. Potential range expansion of Japanese honeysuckle (Lonicera japonica Thunb.) in southern U.S. Forestlands. Forests 2012a, 3, 573–590. [Google Scholar]
- Flory, S.L.; Clay, K. Effects of roads and forest successional age on experimental plant invasions. Biol. Conserv. 2009, 142, 2531–2537. [Google Scholar] [CrossRef]
- Swarbrick, J.T.; Timmins, S.M.; Bullen, K.M.T. The biology of Australian weeds. 36. Ligustrum lucidum Aiton and Ligustrum sinense Lour. Plant Prot. Quart. 1999, 14, 122–130. [Google Scholar]
- Ward, R.W. Extent and dispersal rates of Chinese privet (Ligustrum sinense) invasion on the upper Oconee river floodplain, north Georgia. Southeast. Geogr. 2002, 42, 29–48. [Google Scholar] [CrossRef]
- Renne, I.J.; Gauthreaux, S.A.; Gresham, C.A. Seed dispersal of the Chinese tallow tree (Sapium sebiferum (L.) Roxb.) by birds in coastal South Carolina. Am. Midl. Nat. 2000, 144, 202–215. [Google Scholar]
- Wang, H.-H.; Grant, W.E.; Swannack, T.M.; Gan, J.; Rogers, W.E.; Koralewski, T.E.; Miller, J.H.; Taylor, J.W. Predicted range expansion of Chinese tallow tree (Triadica sebifera) in forestlands of the southern United States. Divers. Distrib. 2011, 17, 552–565. [Google Scholar] [CrossRef]
- Adams, S.B.; Hamel, P.B.; Connor, K.; Burke, B.; Gardiner, E.S.; Wise, D. Potential roles of fish, birds, and water in swamp privet (Forestiera acuminata) seed dispersal. Southeast. Nat. 2007, 6, 669–682. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting the geography of species’ invasions via ecological niche modeling. Quart. Rev. Biol. 2003, 78, 419–433. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.-H.; Grant, W.E. Invasion of Eastern Texas Forestlands by Chinese Privet: Efficacy of Alternative Management Strategies. Diversity 2014, 6, 652-664. https://doi.org/10.3390/d6040652
Wang H-H, Grant WE. Invasion of Eastern Texas Forestlands by Chinese Privet: Efficacy of Alternative Management Strategies. Diversity. 2014; 6(4):652-664. https://doi.org/10.3390/d6040652
Chicago/Turabian StyleWang, Hsiao-Hsuan, and William E. Grant. 2014. "Invasion of Eastern Texas Forestlands by Chinese Privet: Efficacy of Alternative Management Strategies" Diversity 6, no. 4: 652-664. https://doi.org/10.3390/d6040652
APA StyleWang, H.-H., & Grant, W. E. (2014). Invasion of Eastern Texas Forestlands by Chinese Privet: Efficacy of Alternative Management Strategies. Diversity, 6(4), 652-664. https://doi.org/10.3390/d6040652