
Annual Dynamics of Phytoplankton Communities in Relation to Environmental Factors in Saline–Alkaline Lakes of Northwest China

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Site
2.2. Sample Collection and Processing
2.3. Water Quality Parameter Measurements
2.4. Data Processing
2.4.1. Density and Biomass of Phytoplankton
2.4.2. Dominant Species and Biodiversity Index of Phytoplankton
2.5. Data Analysis
3. Results
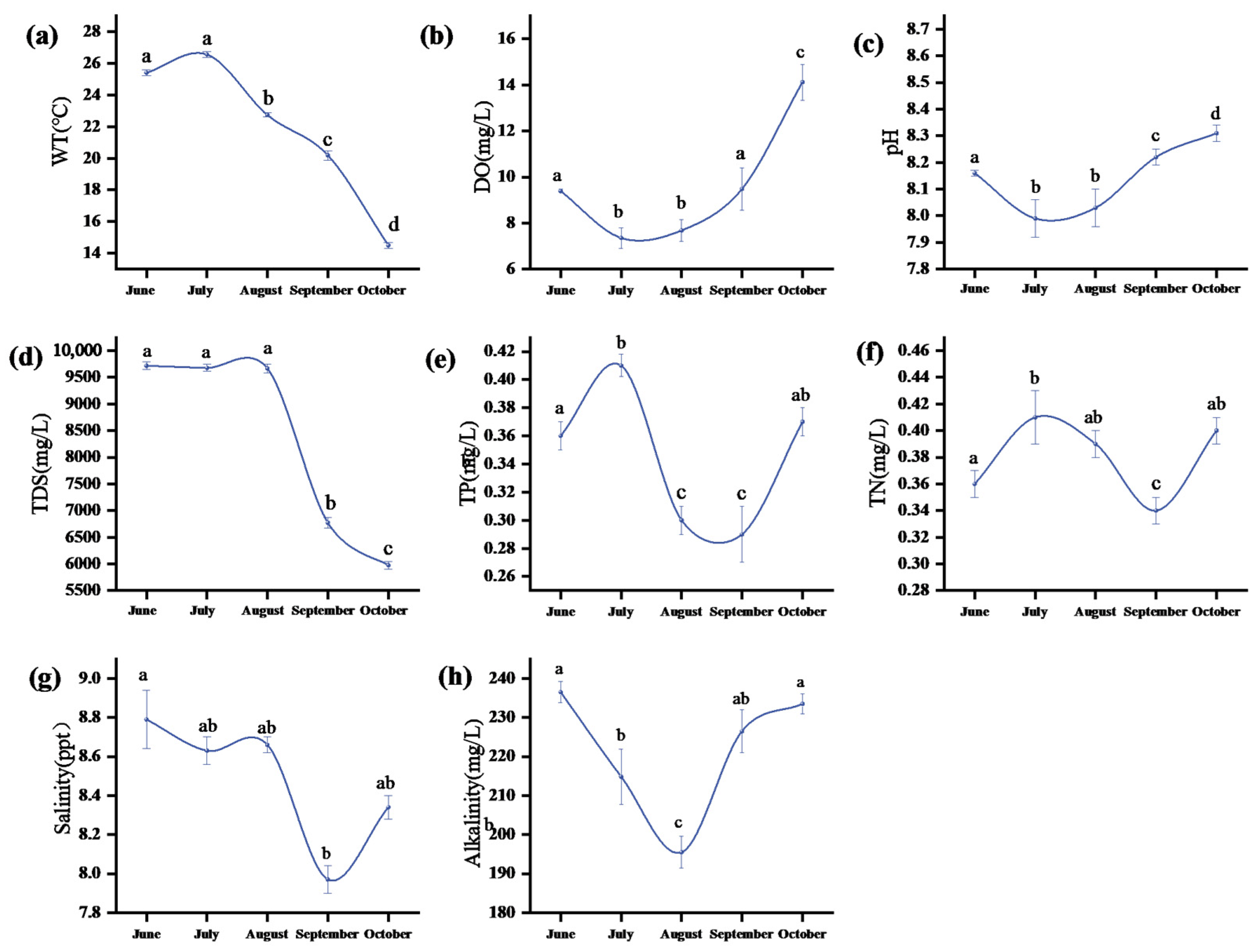
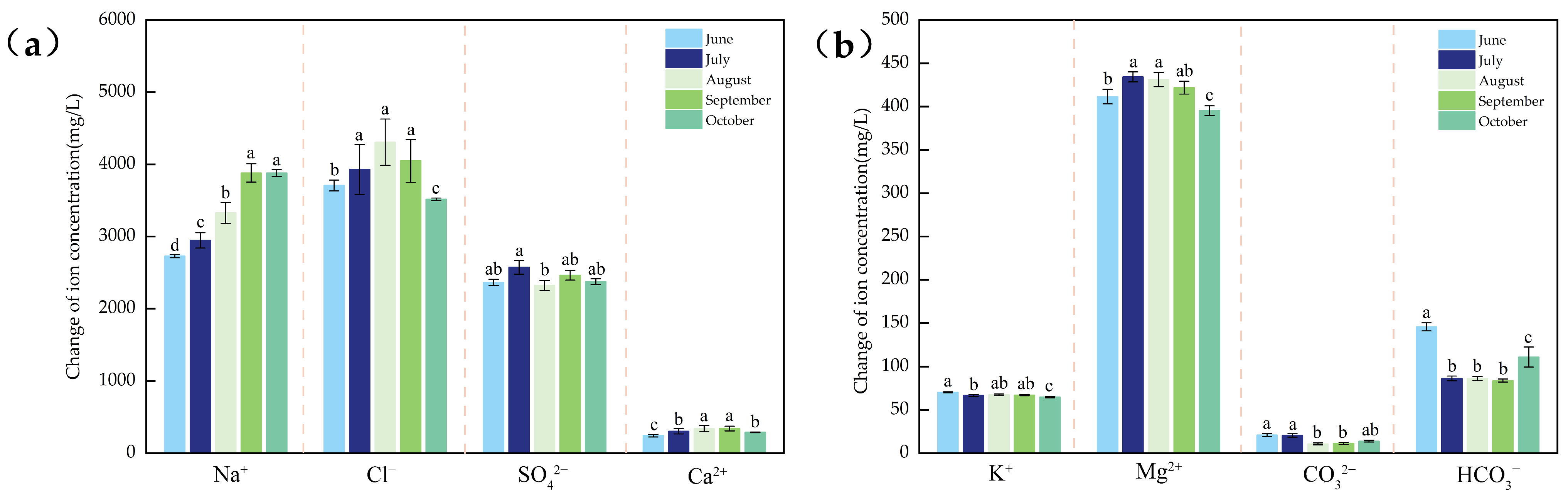
3.1. Physical and Chemical Indicators of Water Body
3.2. Taxonomic Composition of Phytoplankton
3.3. The Relative Density and Relative Biomass of Phytoplankton
3.4. Diversity Index Analysis
3.5. Dominant Phytoplankton Taxa
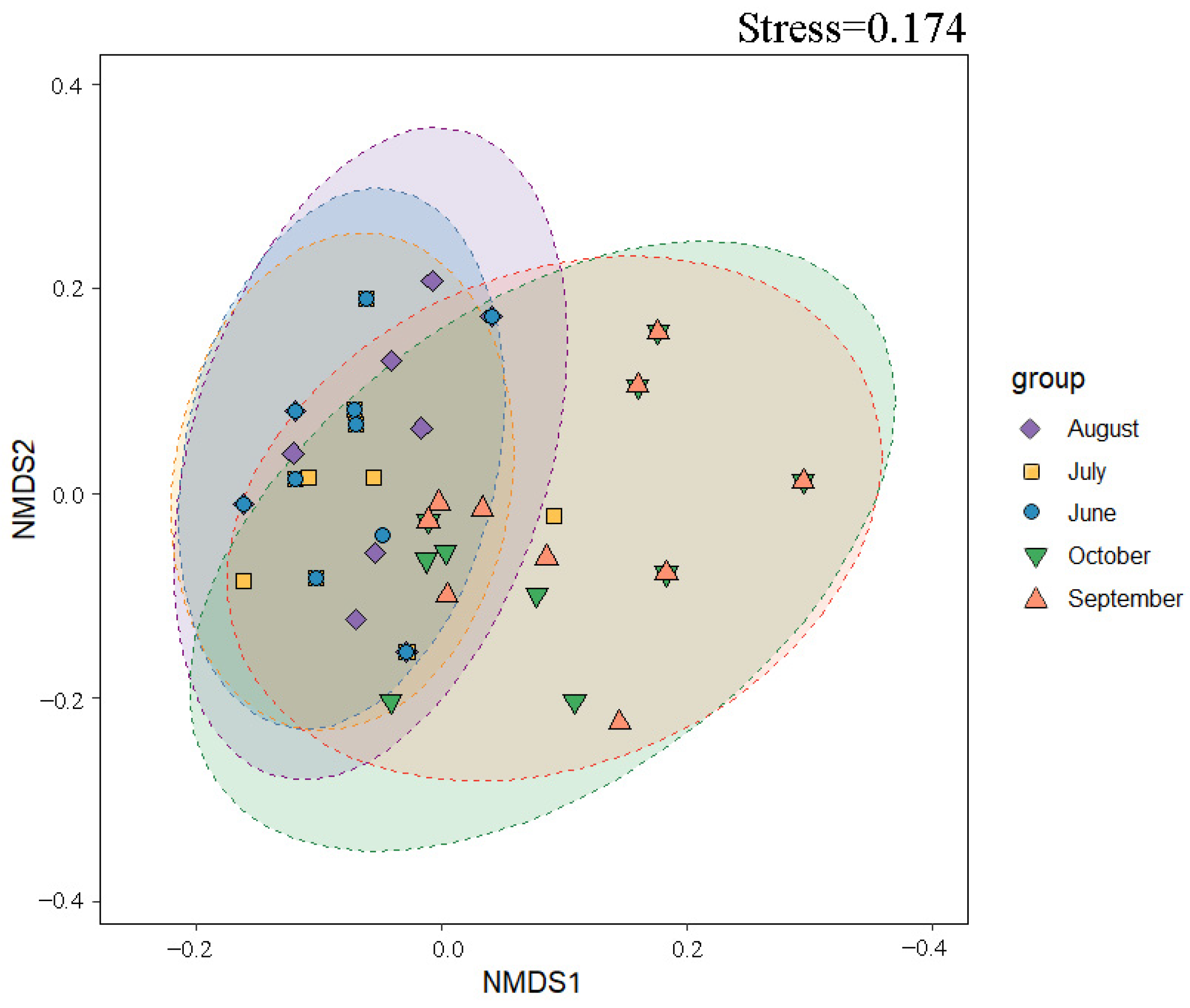
3.6. Analysis of Phytoplankton Community Structure Characteristics
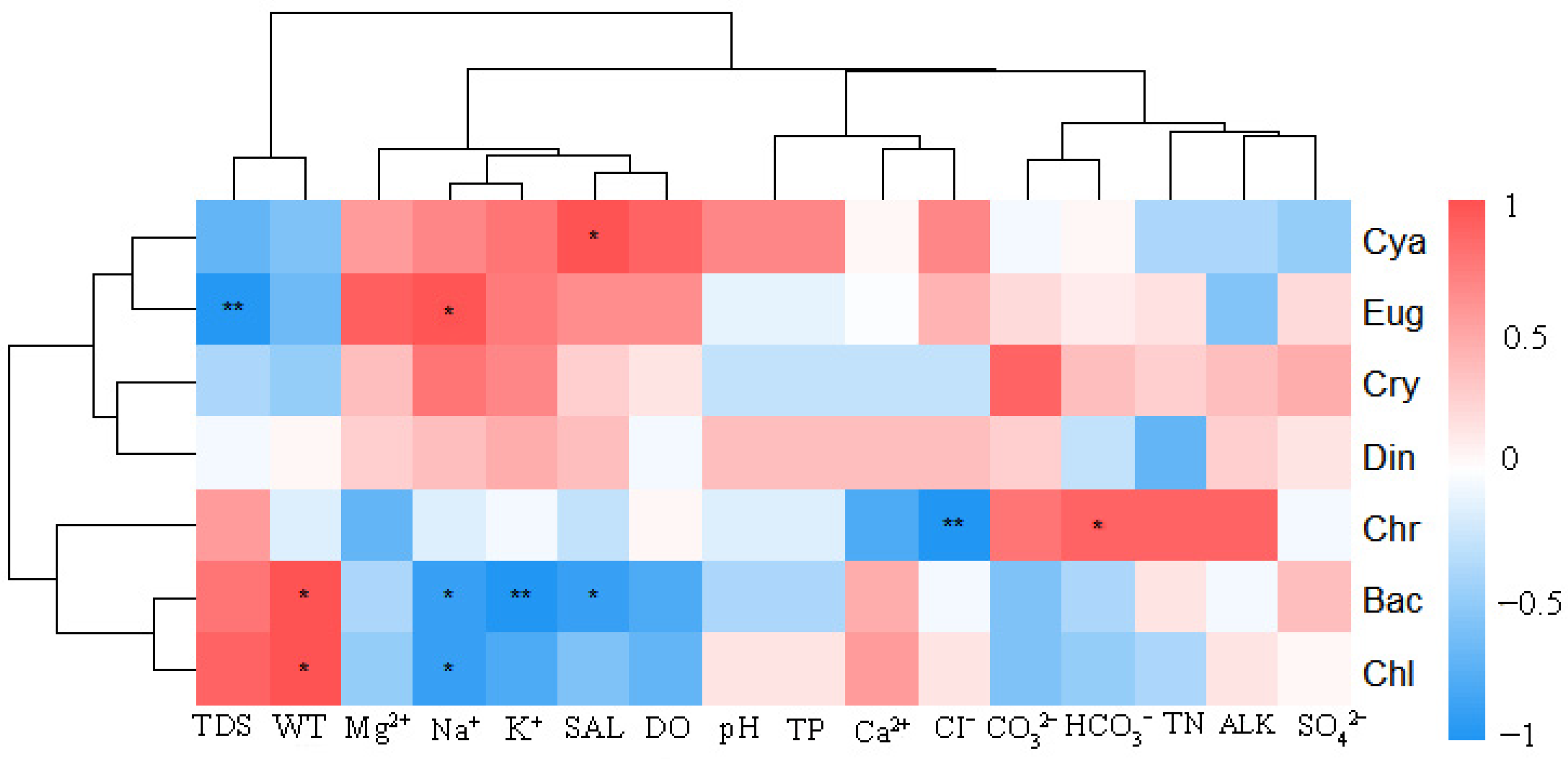
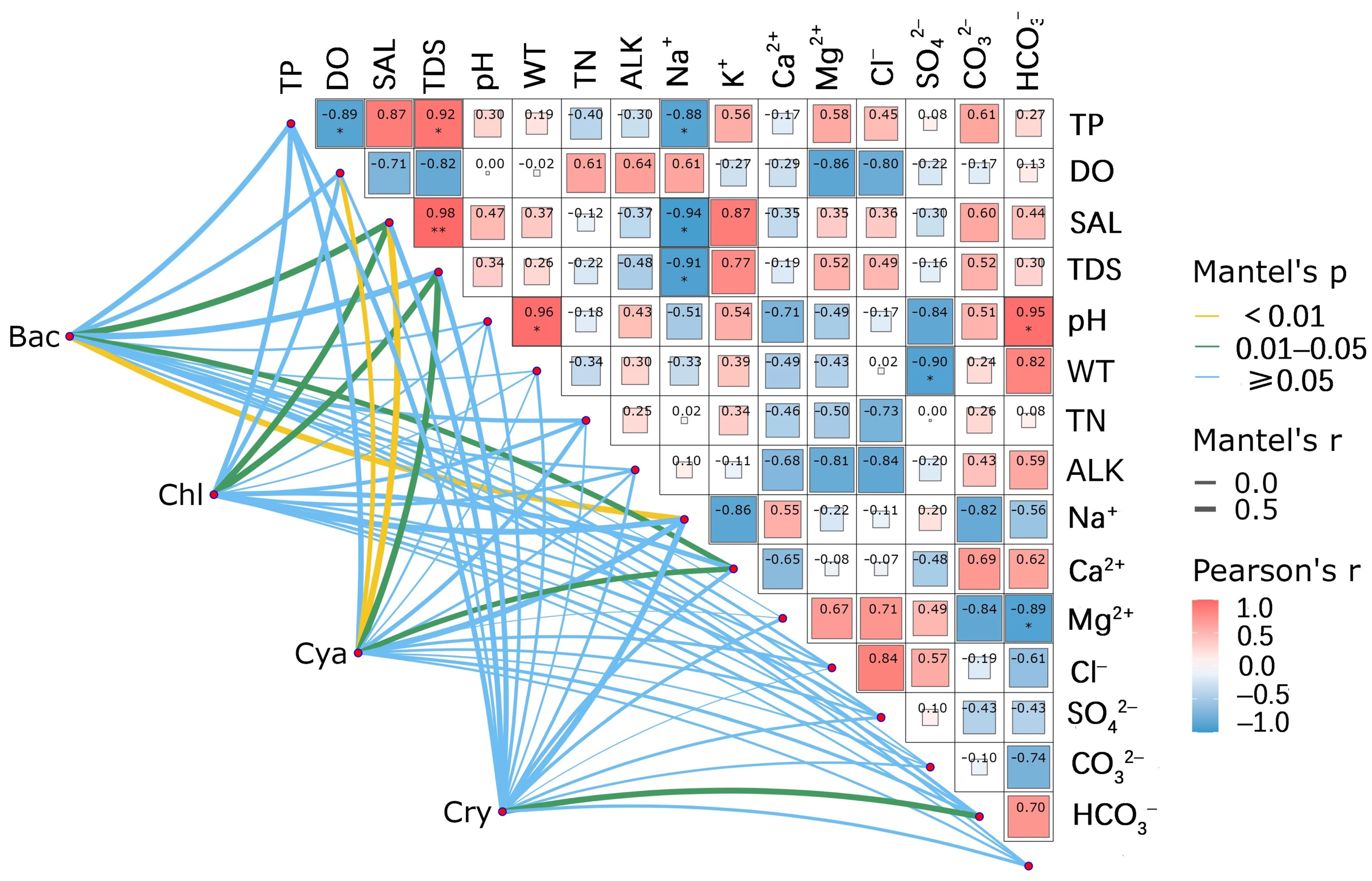
3.7. Correlation Analysis Between the Phytoplankton Community and Environmental Factors
4. Discussion
4.1. Characteristics of the Phytoplankton Community Structure
4.2. Relationships Between Phytoplankton and Environmental Factors
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yan, H.U.; Fan, Y.; Ning, Y.; Wei, J.; Yong, C. Analysis and prospects of saline-alkali land in China from the perspective of utilization. Chin. J. Soil Sci. 2023, 54, 489–494. [Google Scholar]
- Yang, J.; Zhang, S.; Li, Y.; Bu, K.; Zhang, Y.; Chang, L.; Zhang, Y. Dynamics of saline-alkali land and its ecological regionalization in western Songnen plain, China. Chin. Geogr. Sci. 2010, 20, 159–166. [Google Scholar] [CrossRef]
- Zhou, Y.; Li, Y.; Li, W.; Li, F.; Xin, Q. Ecological responses to climate change and human activities in the arid and semi-arid regions of Xinjiang in China. Remote Sens. 2022, 14, 3911. [Google Scholar] [CrossRef]
- Wang, F.; Chen, X.; Luo, G.; Han, Q. Mapping of regional soil salinities in Xinjiang and strategies for amelioration and management. Chin. Geogr. Sci. 2015, 25, 321–336. [Google Scholar] [CrossRef]
- Cuevas, J.; Daliakopoulos, I.N.; Del Moral, F.; Hueso, J.J.; Tsanis, I.K. A review of soil-improving cropping systems for soil salinization. Agronomy 2019, 9, 295. [Google Scholar] [CrossRef]
- Fang, S.; Tu, W.; Mu, L.; Sun, Z.; Hu, Q.; Yang, Y. Saline alkali water desalination project in southern Xinjiang of China: A review of desalination planning, desalination schemes and economic analysis. Renew. Sustain. Energy Rev. 2019, 113, 109268. [Google Scholar] [CrossRef]
- Wang, X.; Zhu, H.; Hou, S.; Cui, H.; Yan, B. Environmental impacts of fertilization during rice production in saline-alkali paddy fields based on life cycle assessment. J. Clean. Prod. 2024, 467, 142947. [Google Scholar] [CrossRef]
- Yu, Y.; Yu, R.; Chen, X.; Yu, G.; Gan, M.; Disse, M. Agricultural water allocation strategies along the oasis of Tarim River in northwest China. Agric. Water Manag. 2017, 187, 24–36. [Google Scholar] [CrossRef]
- Li, Q.; Zhou, J.; Zhou, Y.; Bai, C.; Tao, H.; Jia, R.; Ji, Y.; Yang, G. Variation of groundwater hydrochemical characteristics in the plain area of the Tarim Basin, Xinjiang region, China. Environ. Earth Sci. 2014, 72, 4249–4263. [Google Scholar] [CrossRef]
- Belovsky, G.E.; Stephens, D.; Perschon, C.; Birdsey, P.; Paul, D.; Naftz, D.; Baskin, R.; Larson, C.; Mellison, C.; Luft, J.; et al. The Great Salt Lake ecosystem (Utah, USA): Long term data and a structural equation approach. Ecosphere 2011, 2, art33. [Google Scholar] [CrossRef]
- Pálmai, T.; Szabó, B.; Kotut, K.; Krienitz, L.; Padisák, J. Ecophysiology of a successful phytoplankton competitor in the African flamingo lakes: The green alga Picocystis salinarum (picocystophyceae). J. Appl. Phycol. 2020, 32, 1813–1825. [Google Scholar] [CrossRef]
- Laplace-Treyture, C.; Derot, J.; Prévost, E.; Le Mat, A.; Jamoneau, A. Phytoplankton morpho-functional trait dataset from French water-bodies. Sci. Data 2021, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Hallegraeff, G.M. Ocean climate change, phytoplankton community responses, and harmful algal blooms: A formidable predictive challenge. J. Phycol. 2010, 46, 220–235. [Google Scholar] [CrossRef]
- Deppeler, S.L.; Davidson, A.T. Southern Ocean phytoplankton in a changing climate. Front. Mar. Sci. 2017, 4, 40. [Google Scholar] [CrossRef]
- Xiao, R.; Su, S.; Ghadouani, A.; Wu, J. Spatial analysis of phytoplankton patterns in relation to environmental factors across the southern Taihu Basin, China. Stoch Environ. Res. Risk Assess. 2013, 27, 1347–1357. [Google Scholar] [CrossRef]
- Çelekli, A.; Öztürk, B.; Kapı, M. Relationship between phytoplankton composition and environmental variables in an artificial pond. Algal Res. 2014, 5, 37–41. [Google Scholar] [CrossRef]
- Sun, W.; Dong, S.; Jie, Z.; Zhao, X.; Zhang, H.; Li, J. The impact of net-isolated polyculture of tilapia (Oreochromis niloticus) on plankton community in saline–alkaline pond of shrimp (Penaeus vannamei). Aquacult. Int. 2011, 19, 779–788. [Google Scholar] [CrossRef]
- de Melo Soares, R.H.; de Oliveira Fernandes, F.; de Assunção, C.A.; Borburema, H.D.; do Amaral Carneiro, M.A.; Marinho-Soriano, E. Macroalgal diversity along an environmental gradient in a saltwork. Estuar. Coast. Shelf Sci. 2023, 288, 108377. [Google Scholar] [CrossRef]
- Zhou, B.; Liang, C.; Chen, X.; Ye, S.; Peng, Y.; Yang, L.; Duan, M.; Wang, X. Magnetically-treated brackish water affects soil water-salt distribution and the growth of cotton with film mulch drip irrigation in Xinjiang, China. Agric. Water Manag. 2022, 263, 107487. [Google Scholar] [CrossRef]
- Kumar, M.; Chadha, N.K.; Prakash, S.; Pavan-Kumar, A.; Harikrishna, V.; Gireesh-Babu, P.; Krishna, G. Salinity, stocking density, and their interactive effects on growth performance and physiological parameters of white-leg shrimp, Penaeus vannamei (Boone, 1931), reared in inland ground saline water. Aquacult. Int. 2024, 32, 675–690. [Google Scholar] [CrossRef]
- Anufriieva, E.V. How can saline and hypersaline lakes contribute to aquaculture development? A review. J. Oceanol. Limnol. 2018, 36, 2002–2009. [Google Scholar] [CrossRef]
- Lymbery, A.J.; Kay, G.D.; Doupé, R.G.; Partridge, G.J.; Norman, H.C. The potential of a salt-tolerant plant (Distichlis spicata cv. nypa forage) to treat effluent from inland saline aquaculture and provide livestock feed on salt-affected farmland. Sci. Total Environ. 2013, 445–446, 192–201. [Google Scholar] [CrossRef]
- Salunke, M.; Kalyankar, A.; Khedkar, C.D.; Shingare, M.; Khedkar, G.D. A review on shrimp aquaculture in India: Historical perspective, constraints, status and future implications for impacts on aquatic ecosystem and biodiversity. Rev. Fish. Sci. Aquacult. 2020, 28, 283–302. [Google Scholar] [CrossRef]
- Fatima, A.; Abbas, G.; Kasprzak, R. Assessment of hydrobiological and soil characteristics of non-fertilized, earthen fish ponds in Sindh (Pakistan), supplied with seawater from tidal creeks. Water 2022, 14, 2115. [Google Scholar] [CrossRef]
- Palmer, P.J.; Burke, M.J.; Palmer, C.J.; Burke, J.B. Developments in controlled green-water larval culture technologies for estuarine fishes in Queensland, Australia and elsewhere. Aquaculture 2007, 272, 1–21. [Google Scholar] [CrossRef]
- Zhang, R.; Luo, L.; Wang, S.; Guo, K.; Xu, W.; Zhao, Z. Screening and characteristics of ammonia nitrogen removal bacteria under alkaline environments. Front. Microbiol. 2022, 13, 969722. [Google Scholar] [CrossRef]
- Shang, X.; Geng, L.; Yang, J.; Zhang, Y.; Xu, W. Transcriptome analysis reveals the mechanism of alkalinity exposure on spleen oxidative stress, inflammation and immune function of Luciobarbus capito. Ecotoxicol. Environ. Saf. 2021, 225, 112748. [Google Scholar] [CrossRef]
- Li, K.; Zhao, S.; Guan, W.; Li, K.J. Planktonic bacteria in white shrimp (Litopenaeus vannamei) and channel catfish (Letalurus punetaus) aquaculture ponds in a salt-alkaline region. Lett. Appl. Microbiol. 2022, 74, 212–219. [Google Scholar] [CrossRef]
- Xu, C.; Su, G.; Zhao, K.; Kong, X.; Wang, H.; Xu, X.; Li, Z.; Zhang, M.; Xu, J. Societal benefits and environmental performance of Chinese aquaculture. J. Clean. Prod. 2023, 422, 138645. [Google Scholar] [CrossRef]
- Wang, X.-N.; Gu, Y.-G.; Wang, Z.-H. Fingerprint characteristics and health risks of trace metals in market fish species from a large aquaculture producer in a typical arid province in northwestern China. Environ. Technol. Innov. 2020, 19, 100987. [Google Scholar] [CrossRef]
- Chang, Z.-Q.; Neori, A.; He, Y.-Y.; Li, J.-T.; Qiao, L.; Preston, S.I.; Liu, P.; Li, J. Development and current state of seawater shrimp farming, with an emphasis on integrated multi-trophic pond aquaculture farms, in China—A review. Rev. Aquacult. 2020, 12, 2544–2558. [Google Scholar] [CrossRef]
- Qiu, J.; Zhang, C.; Lv, Z.; Zhang, Z.; Chu, Y.; Shang, D.; Chen, Y.; Chen, C. Analysis of changes in nutrient salts and other water quality indexes in the pond water for largemouth bass (Micropterus salmoides) farming. Heliyon 2024, 10, e24996. [Google Scholar] [CrossRef] [PubMed]
- Yu, R.; Wang, J. Evaluation of the environmental sustainability of farmers’ land use decisions in the saline-alkaline areas. Environ. Monit. Assess. 2015, 187, 182. [Google Scholar] [CrossRef]
- Wang, D.; Zhang, L.; Zhang, J.; Li, W.; Li, H.; Liang, Y.; Han, Y.; Luo, P.; Wang, Z. Effect of magnetized brackish water drip irrigation on water and salt transport characteristics of sandy soil in southern Xinjiang, China. Water 2023, 15, 577. [Google Scholar] [CrossRef]
- Zhang, J.M.; He, Z.H. Handbook of Natural Resources Survey of Inland Waters Fisheries; Agricultural Press: Beijing, China, 1991. [Google Scholar]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China: System, Ecology and Classification; Science Press: Beijing, China, 2006. [Google Scholar]
- Qi, Y.Z.; Li, J.Y. Annals of Freshwater Algae in China; Science Press: Beijing, China, 2004. [Google Scholar]
- Administration, S.E.P. Monitoring and Analysis Methods of Water and Wastewater, 4th ed.; Environmental Science Press: Beijing, China, 2002. [Google Scholar]
- Chen, G.M. Ammonium molybdate spectrophotometric method for determination of total phosphorus in municipal sewage sludge. China Water Wastewater 2006, 22, 85–86. [Google Scholar]
- Jouanneau, S.; Recoules, L.; Durand, M.; Boukabache, A.; Picot, V.; Primault, Y.; Lakel, A.; Sengelin, M.; Barillon, B.; Thouand, G. Methods for assessing biochemical oxygen demand (BOD): A review. Water Res. 2014, 49, 62–82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W. Hydrobiology, 2nd ed.; China Agriculture Press: Beijing, China, 2016. [Google Scholar]
- Grigorszky, I.; Kétyi, T.; Borics, I.; Borbély, G.; Padisák, J.; Bácsi, I.; Bánkuti, K. The effects of temperature, nitrogen, and phosphorus on the encystment of Peridinium cinctum, Stein (Dinophyta). Hydrobiologia 2006, 563, 527–535. [Google Scholar] [CrossRef]
- Wang, X.; Sun, M.; Wang, J.; Yang, L.; Luo, L.; Li, P.; Kong, F. Microcystis genotype succession and related environmental factors in Lake Taihu during cyanobacterial blooms. Microb. Ecol. 2012, 64, 986–999. [Google Scholar] [CrossRef]
- Gamito, S. Caution is needed when applying margalef diversity index. Ecol. Indic. 2010, 10, 550–551. [Google Scholar] [CrossRef]
- Li, X.; Yu, H.; Wang, H.; Ma, C. Phytoplankton community structure in relation to environmental factors and ecological assessment of water quality in the upper reaches of the Genhe River in the Greater Hinggan Mountains. Environ. Sci. Pollut. Res. 2019, 26, 17512–17519. [Google Scholar] [CrossRef]
- Li, X.; Song, Y.; Nian, F. Characteristics of water transformation and its effects on environment in the arid region. Chin. Geogr. Sci. 2000, 10, 52–60. [Google Scholar] [CrossRef]
- Zhang, D.; Zhao, W.; Zhang, G. Soil moisture and salt ionic composition effects on species distribution and diversity in semiarid inland saline habitats, Northwestern China. Ecol. Res. 2018, 33, 505–515. [Google Scholar] [CrossRef]
- Sui, F.; Zang, S.; Fan, Y.; Ye, H. Effects of different saline-alkaline conditions on the characteristics of phytoplankton communities in the lakes of Songnen Plain, China. PLoS ONE 2016, 11, e0164734. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, Y.; Wang, S.; Lu, Y.; Sun, K.; Jia, J.; Wang, Y. Phytoplankton community response to nutrients along lake salinity and altitude gradients on the Qinghai-Tibet Plateau. Ecol. Indic. 2021, 128, 107848. [Google Scholar] [CrossRef]
- Zhao, Z.; Song, T.; Zhang, M.; Tong, S.; An, Y.; Zhang, P.; Sang, B.; Cao, G. Benefits of morphology-based functional group classification to study dynamic changes in phytoplankton in saline-alkali wetlands, taking typical saline-alkali wetlands in Northeast China as an example. Diversity 2023, 15, 1175. [Google Scholar] [CrossRef]
- Xu, F.; Li, P.; Du, Q.; Yang, Y.; Yue, B. Seasonal hydrochemical characteristics, geochemical evolution, and pollution sources of Lake Sha in an arid and semiarid region of Northwest China. Expo. Health 2023, 15, 231–244. [Google Scholar] [CrossRef]
- Wei, D.; Yan, H.; Xin-shan, S.; Bai-xing, Y. Hydrochemical characteristics of salt marsh wetlands in western Songnen Plain. J. Geogr. Sci 2001, 11, 217–223. [Google Scholar] [CrossRef]
- Astorg, L.; Gagnon, J.-C.; Lazar, C.S.; Derry, A.M. Effects of freshwater salinization on a salt-naïve planktonic eukaryote community. Limnol. Oceanogr. Lett. 2023, 8, 38–47. [Google Scholar] [CrossRef]
- Chakraborty, P.; Acharyya, T.; Babu, P.V.R.; Bandyopadhyay, D. Impact of salinity and pH on phytoplankton communities in a tropical freshwater system: An investigation with pigment analysis by HPLC. J. Environ. Monit. 2011, 13, 614–620. [Google Scholar] [CrossRef]
- Liu, Z.; Gao, M.; Sun, Q.; Hou, G.; Zhao, Y. Formation and evolution of soil salinization based on multivariate statistical methods in Ningxia Plain, China. Front. Earth Sci. 2023, 11, 1186779. [Google Scholar] [CrossRef]
- Huang, Y.; Shen, Y.; Zhang, S.; Li, Y.; Sun, Z.; Feng, M.; Li, R.; Zhang, J.; Tian, X.; Zhang, W. Characteristics of phytoplankton community structure and indication to water quality in the lake in agricultural areas. Front. Environ. Sci. 2022, 10, 833409. [Google Scholar] [CrossRef]
- Greco, D.A.; Arnott, S.E.; Fournier, I.B.; Schamp, B.S. Effects of chloride and nutrients on freshwater plankton communities. Limnol. Oceanogr. Lett. 2023, 8, 48–55. [Google Scholar] [CrossRef]
- Duan, L.; Wang, W.; Zhou, L.; Cheng, Z. The formation of shallow fresh groundwater in the north of Yanchi County, Ningxia, China: Main influencing factors and mechanism. Environ. Earth Sci. 2016, 75, 461. [Google Scholar] [CrossRef]
- Mo, Y.; Peng, F.; Gao, X.; Xiao, P.; Logares, R.; Jeppesen, E.; Ren, K.; Xue, Y.; Yang, J. Low shifts in salinity determined assembly processes and network stability of microeukaryotic plankton communities in a subtropical urban reservoir. Microbiome 2021, 9, 128. [Google Scholar] [CrossRef]
- Znachor, P.; Zapomělová, E.; Řeháková, K.; Nedoma, J.; Šimek, K. The effect of extreme rainfall on summer succession and vertical distribution of phytoplankton in a lacustrine part of a eutrophic reservoir. Aquat. Sci. 2008, 70, 77–86. [Google Scholar] [CrossRef]
- Zhou, L.; Wu, S.; Gu, W.; Wang, L.; Wang, J.; Gao, S.; Wang, G. Photosynthesis acclimation under severely fluctuating light conditions allows faster growth of diatoms compared with dinoflagellates. BMC Plant Biol. 2021, 21, 164. [Google Scholar]
- Deng, J.; Qin, B.; Paerl, H.W.; Zhang, Y.; Wu, P.; Ma, J.; Chen, Y. Effects of nutrients, temperature and their interactions on spring phytoplankton community succession in Lake Taihu, China. PLoS ONE 2014, 9, e113960. [Google Scholar] [CrossRef]
- Thomas, M.K.; Litchman, E. Effects of temperature and nitrogen availability on the growth of invasive and native cyanobacteria. Hydrobiologia 2016, 763, 357–369. [Google Scholar] [CrossRef]
- Bakker, E.S.; Hilt, S. Impact of water-level fluctuations on cyanobacterial blooms: Options for management. Aquat Ecol. 2016, 50, 485–498. [Google Scholar] [CrossRef]
- Li, C.; Feng, W.; Chen, H.; Li, X.; Song, F.; Guo, W.; Giesy, J.P.; Sun, F. Temporal variation in zooplankton and phytoplankton community species composition and the affecting factors in Lake Taihu—A large freshwater lake in China. Environ. Pollut. 2019, 245, 1050–1057. [Google Scholar] [CrossRef]
- Bryanskaya, A.V.; Malup, T.K.; Lazareva, E.V.; Taran, O.P.; Rozanov, A.S.; Efimov, V.M.; Peltek, S.E. The role of environmental factors for the composition of microbial communities of saline lakes in the Novosibirsk Region (Russia). BMC Microbiol. 2016, 16, S4. [Google Scholar] [CrossRef]
- Trombetta, T.; Vidussi, F.; Mas, S.; Parin, D.; Simier, M.; Mostajir, B. Water temperature drives phytoplankton blooms in coastal waters. PLoS ONE 2019, 14, e0214933. [Google Scholar] [CrossRef]
- Kholssi, R.; Lougraimzi, H.; Moreno-Garrido, I. Effects of global environmental change on microalgal photosynthesis, growth and their distribution. Mar. Environ. Res. 2023, 184, 105877. [Google Scholar] [CrossRef]
- Kutlu, B.; Aydın, R.; Danabas, D.; Serdar, O. Temporal and seasonal variations in phytoplankton community structure in Uzuncayir Dam Lake (Tunceli, Turkey). Environ. Monit. Assess. 2020, 192, 105. [Google Scholar] [CrossRef] [PubMed]
- Nalley, J.O.; O’Donnell, D.R.; Litchman, E. Temperature effects on growth rates and fatty acid content in freshwater algae and cyanobacteria. Algal Res. 2018, 35, 500–507. [Google Scholar] [CrossRef]
- Mitrovic, S.M.; Hitchcock, J.N.; Davie, A.W.; Ryan, D.A. Growth responses of Cyclotella meneghiniana (Bacillariophyceae) to various temperatures. J. Plankton Res. 2010, 32, 1217–1221. [Google Scholar] [CrossRef]
- Dai, Y.; Yang, S.; Zhao, D.; Hu, C.; Xu, W.; Anderson, D.M.; Li, Y.; Song, X.-P.; Boyce, D.G.; Gibson, L.; et al. Coastal phytoplankton blooms expand and intensify in the 21st century. Nature 2023, 615, 280–284. [Google Scholar] [CrossRef]
- Pulsifer, J.; Laws, E. Temperature dependence of freshwater phytoplankton growth rates and zooplankton grazing rates. Water 2021, 13, 1591. [Google Scholar] [CrossRef]
- Ayiti, O.E.; Babalola, O.O. Factors influencing soil nitrification process and the effect on environment and health. Front. Sustain. Food Syst. 2022, 6, 821994. [Google Scholar] [CrossRef]
- Yang, J.; Wang, C.; Wang, Z.; Li, Y.; Yu, H.; Feng, J.; Xie, S.; Li, X. Distribution patterns and co-occurrence network of eukaryotic algae in different salinity waters of Yuncheng Salt Lake, China. Sci. Rep. 2024, 14, 8340. [Google Scholar] [CrossRef]
- Hinga, K.R. Effects of pH on coastal marine phytoplankton. Mar. Ecol. Prog. Ser. 2002, 238, 281–300. [Google Scholar] [CrossRef]
- Moschonas, G.; Gowen, R.J.; Paterson, R.F.; Mitchell, E.; Stewart, B.M.; McNeill, S.; Glibert, P.M.; Davidson, K. Nitrogen dynamics and phytoplankton community structure: The role of organic nutrients. Biogeochemistry 2017, 134, 125–145. [Google Scholar] [CrossRef]
- Hagemann, M. Molecular biology of cyanobacterial salt acclimation. FEMS Microbiol. Rev. 2011, 35, 87–123. [Google Scholar] [CrossRef]
- Wang, C.; Sun, B.; Zhang, X.; Huang, X.; Zhang, M.; Guo, H.; Chen, X.; Huang, F.; Chen, T.; Mi, H.; et al. Structural mechanism of the active bicarbonate transporter from cyanobacteria. Nat. Plants 2019, 5, 1184–1193. [Google Scholar] [CrossRef]
- Xu, Y.; Shi, D.; Aristilde, L.; Morel, F.M.M. The effect of pH on the uptake of zinc and cadmium in marine phytoplankton: Possible role of weak complexes. Limnol. Oceanogr. 2012, 57, 293–304. [Google Scholar] [CrossRef]
- Brewer, P.G.; Goldman, J.C. Alkalinity changes generated by phytoplankton growth. Limnol. Oceanogr. 1976, 21, 108–117. [Google Scholar] [CrossRef]
- Ma, J.; Wang, P. Effects of rising atmospheric CO2 levels on physiological response of cyanobacteria and cyanobacterial bloom development: A review. Sci. Total Environ. 2021, 754, 141889. [Google Scholar] [CrossRef]
- Domingues, R.B.; Anselmo, T.P.; Barbosa, A.B.; Sommer, U.; Galvão, H.M. Nutrient limitation of phytoplankton growth in the freshwater tidal zone of a turbid, Mediterranean estuary. Estuar. Coast. Shelf Sci. 2011, 91, 282–297. [Google Scholar] [CrossRef]
- Piatka, D.R.; Frank, A.H.; Köhler, I.; Castiglione, K.; van Geldern, R.; Barth, J.A.C. Balance of carbon species combined with stable isotope ratios show critical switch towards bicarbonate uptake during cyanobacteria blooms. Sci. Total Environ. 2022, 807, 151067. [Google Scholar] [CrossRef]
- Rydzyński, D.; Piotrowicz-Cieślak, A.I.; Grajek, H.; Wasilewski, J. Investigation of chlorophyll degradation by tetracycline. Chemosphere 2019, 229, 409–417. [Google Scholar] [CrossRef]
- Yan, S.; Zhang, T.; Zhang, B.; Feng, H. A revised saline water quality assessment method considering Mg2+/Na+ as a new indicator for arid irrigated areas. J. Hydrol. 2024, 639, 131619. [Google Scholar] [CrossRef]
- Zhu, Y.; Qi, Q.; Lu, X.; Fan, Y.; Liu, Y.; Tan, X. Local environmental variables outperform spatial and land use pattern in the maintenance and assembly of phytoplankton communities in the wetland cluster. J. Clean. Prod. 2023, 419, 138275. [Google Scholar] [CrossRef]

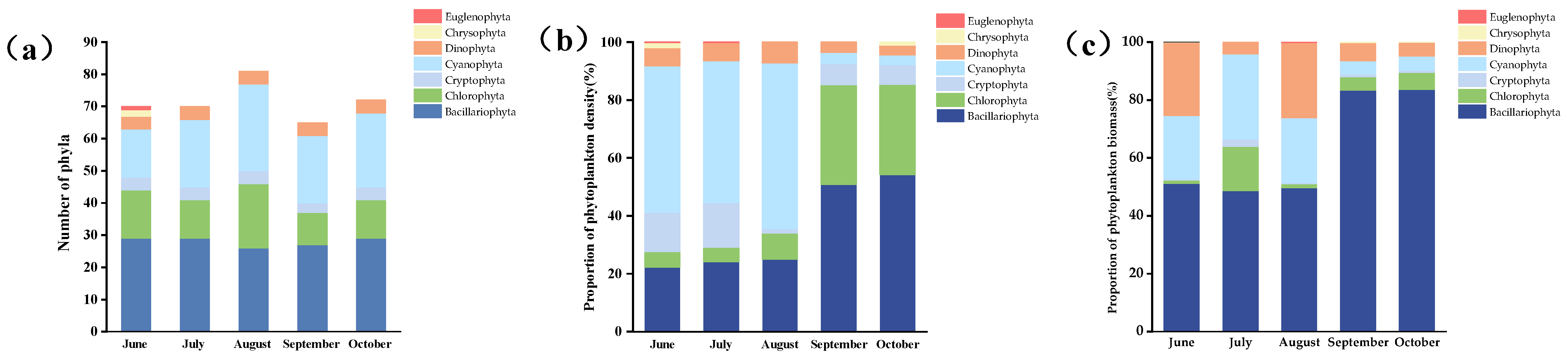



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial Number | Physical and Chemical Indicators | Analytical Method | Reference |
|---|---|---|---|
| 1 | Total Phosphorus (TP) (mg/L) | Ammonium molybdate spectrophotometric method | [39] |
| 2 | Total nitrogen (TN) (mg/L) | Alkaline potassium persulfate ultraviolet spectrophotometric method | [40] |
| 3 | Alkalinity (ALK) (mg/L) | Acid-base indicator titration method | [38] |
| 4 | Na+, K+ (mg/L) | Flame atomic absorption spectrophotometry | [38] |
| 5 | Mg2+, Ca2+ (mg/L) | Volumetric method | [38] |
| 6 | Cl−, SO42−, CO32−, HCO3− (mg/L) | Potentiometric titration | [38] |
| Phylum | Genus | Dominant Taxa | June | July | August | September | October |
|---|---|---|---|---|---|---|---|
| Bacillariophyta | Cyclotella | Cyclotella spp. | 0.04 | 0.03 | - | - | 0.03 |
| Cyclotella | Cyclotella meneghiniana | 0.04 | - | 0.04 | 0.04 | 0.06 | |
| Cyanophyta | Merismopedia | Merismopedia sp. | 0.12 | 0.03 | - | - | - |
| Microcystis | Microcystis spp. | 0.15 | 0.14 | 0.12 | 0.04 | - | |
| Chlorophyta | Chlorella | Chlorella sp. | 0.04 | 0.04 | 0.04 | - | 0.03 |
| Cryptophyta | Chroomonas | Chroomonas acuta | 0.05 | - | - | - | 0.03 |
| Navicula | Navicula cryptocephala | - | - | 0.03 | - | 0.05 | |
| Dinophyta | Peridinium | Gymnodinium aeruginosum | 0.05 | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Y.; Hu, L.; Ma, R.; Yang, L.; Huo, Q.; Song, Y.; Lin, X.; Sun, Z.; Chen, S.; Ren, D. Annual Dynamics of Phytoplankton Communities in Relation to Environmental Factors in Saline–Alkaline Lakes of Northwest China. Diversity 2025, 17, 328. https://doi.org/10.3390/d17050328
Ma Y, Hu L, Ma R, Yang L, Huo Q, Song Y, Lin X, Sun Z, Chen S, Ren D. Annual Dynamics of Phytoplankton Communities in Relation to Environmental Factors in Saline–Alkaline Lakes of Northwest China. Diversity. 2025; 17(5):328. https://doi.org/10.3390/d17050328
Chicago/Turabian StyleMa, Yuying, Linghui Hu, Ruomei Ma, Liting Yang, Qiang Huo, Yong Song, Xuyuan Lin, Zhen Sun, Sheng’ao Chen, and Daoquan Ren. 2025. "Annual Dynamics of Phytoplankton Communities in Relation to Environmental Factors in Saline–Alkaline Lakes of Northwest China" Diversity 17, no. 5: 328. https://doi.org/10.3390/d17050328
APA StyleMa, Y., Hu, L., Ma, R., Yang, L., Huo, Q., Song, Y., Lin, X., Sun, Z., Chen, S., & Ren, D. (2025). Annual Dynamics of Phytoplankton Communities in Relation to Environmental Factors in Saline–Alkaline Lakes of Northwest China. Diversity, 17(5), 328. https://doi.org/10.3390/d17050328