
Ecological Factors Associated with Burrow System Occupancy by Great Desert Skinks (Liopholis kintorei)


{kind=link}
{kind=link}
Abstract
1. Introduction
2. Methods
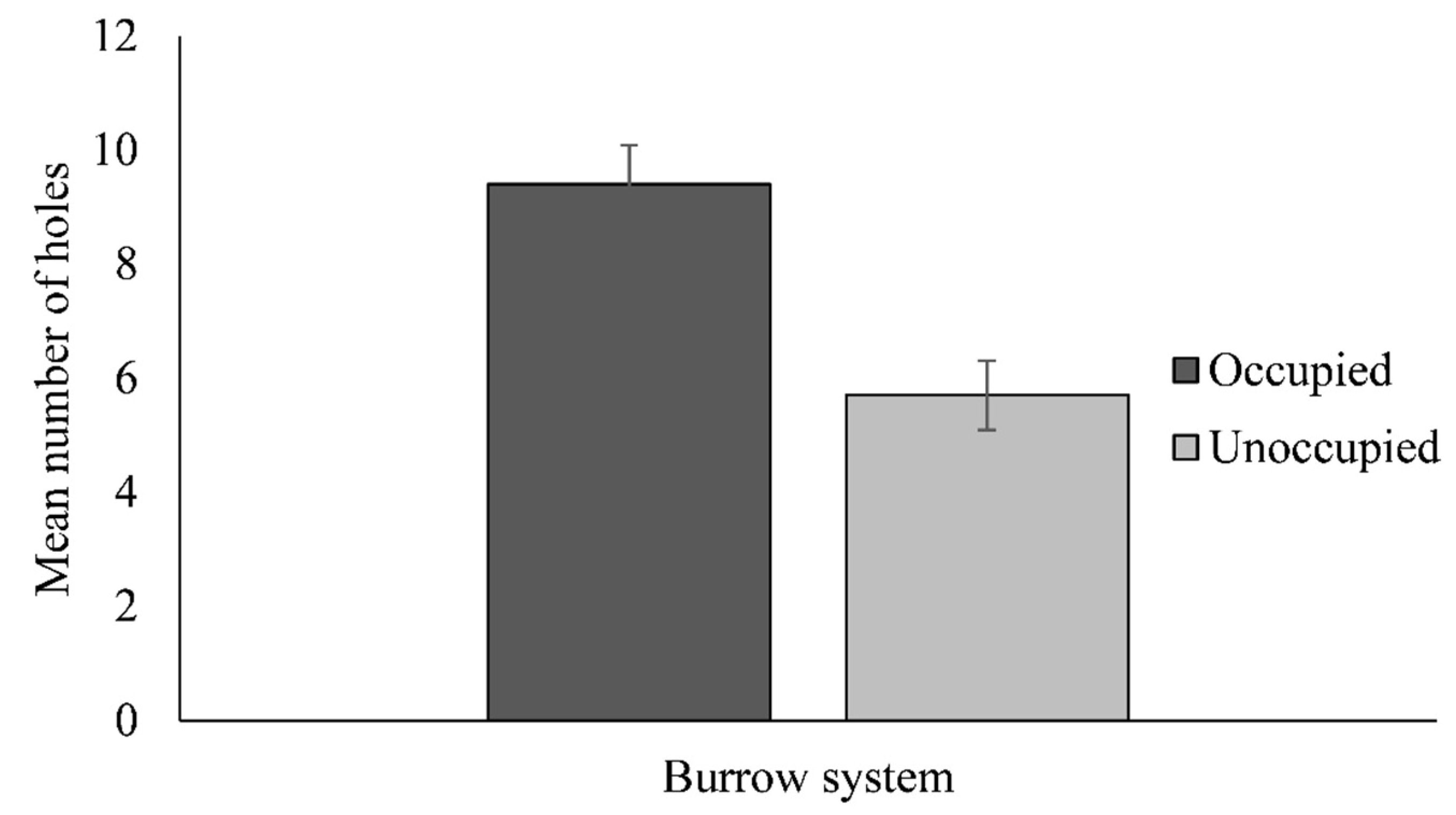
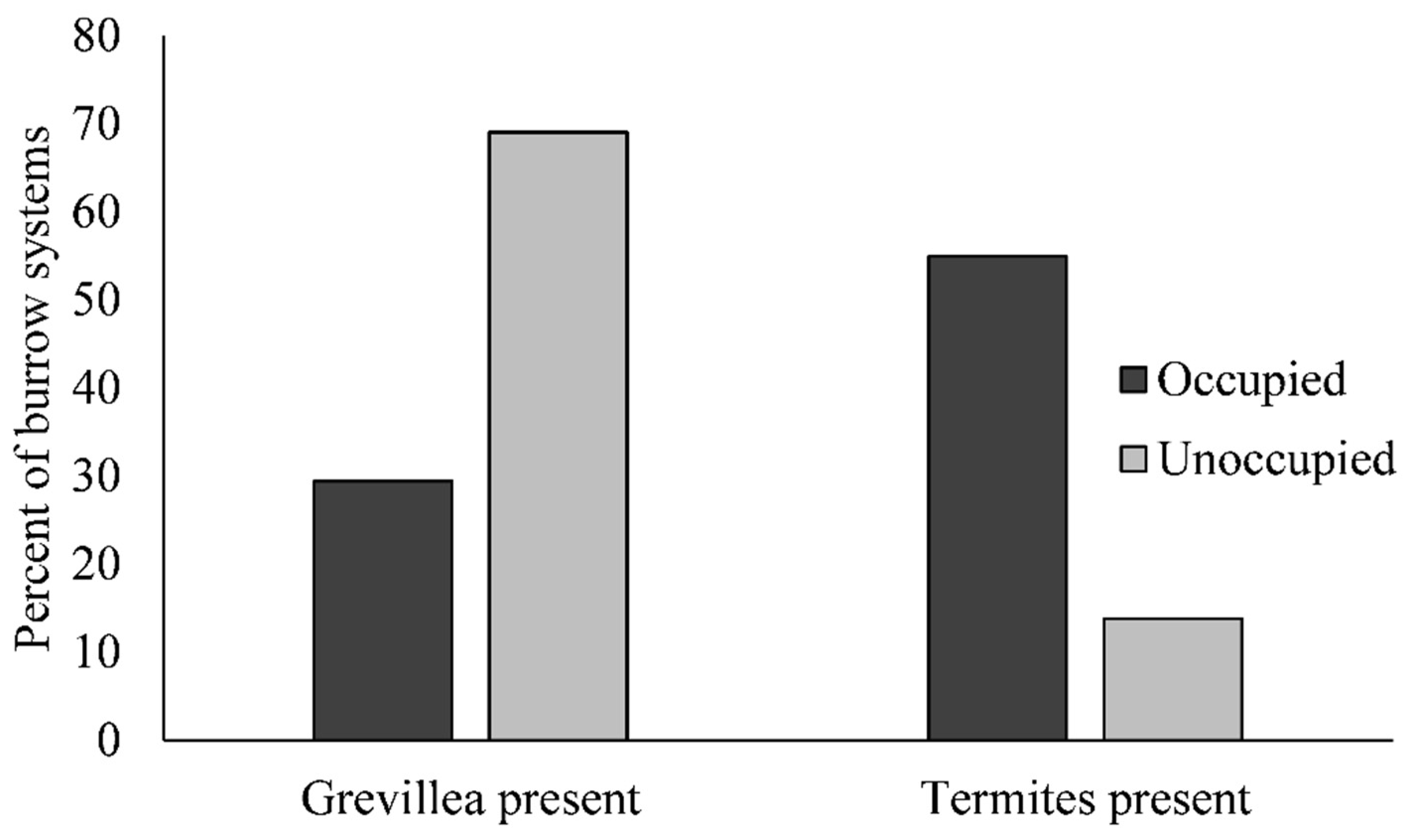
3. Results
3.1. Demographics
3.2. Habitat
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kinlaw, A. A review of borrowing by semi-fossorial vertebrates in arid environments. J. Arid. Environ. 1999, 41, 127–145. [Google Scholar] [CrossRef]
- Moore, D.; Kearney, M.R.; Paltridge, R.; McAlpin, S.; Stow, A. Is fire a threatening process for Liopholis kintorei, a nationally listed threatened skink? Wildl. Res. 2015, 42, 207–216. [Google Scholar] [CrossRef]
- Hodgson, M.J.; Ritchie, D. Strange bedfellows: Mammal burrow disturbances may provide thermoregulatory microsites for fossorial reptiles in densely vegetated dunes. Austral Ecol. 2023, 48, 1473–1478. [Google Scholar] [CrossRef]
- Morris, S.D.; Johnson, C.N.; Brook, B.W.; Kearney, M.R. Seasonal and depth-dependent thermoregulatory benefits of burrows for wombats—The largest burrowing marsupials. J. Therm. Biol. 2024, 125, 103961. [Google Scholar] [CrossRef] [PubMed]
- Cadenhead, N.C.R.; Kearney, M.R.; Moore, D.; McAlpin, S.; Wintle, B.A. Climate and fire scenario uncertainty dominate the evaluation of options for conserving the Great Desert Skink. Conserv. Lett. 2016, 9, 181–190. [Google Scholar] [CrossRef]
- Robb, G.N.; Harrison, A.; Woodborne, S.; Bennett, N.C. Diet composition of two common mole-rat populations in arid and mesic environments in South Africa as determined by stable isotope analysis. J. Zool. 2016, 300, 257–264. [Google Scholar] [CrossRef]
- Hoogland, J.L. The Black-Tailed Prairie Dog: Social Life of a Burrowing Mammal; Chicago University Press: Chicago, IL, USA, 1995. [Google Scholar]
- Spinks, A.C.; Bennett, N.C.; Jarvis, J.U.M. A comparison of the ecology of two populations of the common mole-rat, Cryptomys hottentotus hottentotus: The effect of aridity on food, foraging and body mass. Oecologia 2000, 125, 341–349. [Google Scholar] [CrossRef]
- Ebensperger, L.A.; Chesh, A.S.; Castro, R.A.; Tolhuysen, L.O.; Quirici, V.; Burger, J.R.; Sobrero, R.; Hayes, L.D. Burrow limitations and group living in the communally rearing rodent, Octodon degus. J. Mammal. 2011, 92, 21–30. [Google Scholar] [CrossRef]
- Chapple, D.G. Ecology, life-history, and behavior in the Australian scincid genus Egernia, with comments on the evolution of complex sociality in lizards. Herpetol. Monogr. 2003, 17, 145–180. [Google Scholar] [CrossRef]
- Moore, D.; Stow, A.; Kearney, M.R. Under the weather?—The direct effects of climate warming on a threatened desert lizard are mediated by their activity phase and burrow system. J. Anim. Ecol. 2017, 87, 660–671. [Google Scholar] [CrossRef]
- Du, W.-G.; Li, S.-R.; Sun, B.-J.; Shine, R. Can nesting behavior allow reptiles to adapt to climate change? Philos. Trans. R. Soc. B 2022, 278, 20220153. [Google Scholar]
- Hradsky, B.A.; Mildwaters, C.; Ritchie, E.G.; Christie, F.; Di Stefano, J. Responses of invasive predators and natural prey to a prescribed forest fire. J. Mammal. 2017, 98, 835–847. [Google Scholar] [CrossRef]
- Davidson, A.D.; Lightfoot, D.C.; McIntyre, J.L. Engineering rodents create key habitat for lizards. J. Arid. Environ. 2008, 72, 2142–2149. [Google Scholar] [CrossRef]
- Molyneux, J.; Pavey, C.R.; James, A.I.; Carthew, S.M. Habitat use by the brush-tailed mulgara (Dasycercus blythi). Aust. J. Zool. 2017, 65, 335–345. [Google Scholar] [CrossRef]
- Pianka, E.R. Convexity, desert lizards, and spatial heterogeneity. Ecology 1966, 47, 1055–1059. [Google Scholar] [CrossRef]
- Paltridge, R.; Catt, G.; Cowan, M.; Gaikhorst, G.; How, R.; Zichy-Woinarski, J.; Cogger, H.; Teale, R. Liopholis kintorei. IUCN Red List of Threatened Species; International Union for Conservation of Nature and Natural Resources: Gland, Switzerland, 2018. [Google Scholar] [CrossRef]
- Indigenous Desert Alliance. Looking After Tjakura, Tjalapa, Mulyamiji, Warrarna, Nampu: National Recovery Plan for the Great Desert Skink (Liopholis kintorei) 2023–2033; Department of Climate Change, Energy, the Environment and Water: Canberra, Australia, 2023.
- McAlpin, S.; Duckett, P.; Stow, A. Lizards cooperatively tunnel to construct a long-term home for family members. PLoS ONE 2011, 6, e19041. [Google Scholar] [CrossRef]
- McAlpin, S. A Recovery Plan for the Great Desert Skink (Egernia kintorei); Arid Lands Environment Centre: Alice Springs, Australia, 2001. [Google Scholar]
- Thuo, D.; Macgregor, N.A.; Merson, S.D.; Scopel, D.; Keogh, J.S.; Kenny, J.; Williams, J.L.; Guest, T.; Swan, S.; McAlpin, S.; et al. Metabarcoding clarifies the diet of the elusive and vulnerable Australian tjakuṟa (great desert skink, Liopholis kintorei). Front. Ecol. Evol. 2024, 12, 1354138. [Google Scholar] [CrossRef]
- Abensperg-Traun, M.; Steven, D. Ant- and termite-eating in Australian mammals and lizards: A comparison. Aust. J. Ecol. 1997, 22, 9–17. [Google Scholar] [CrossRef]
- McGregor, H.W.; Legge, S.; Jones, M.E.; Johnson, C.N. Landscape management of fire and grazing regimes alters fine-scale habitat utilisation by feral cats. PLoS ONE 2014, 9, e109097. [Google Scholar] [CrossRef]
- Moore, D.; Kearney, M.R.; Paltridge, R.; McAlpin, S.; Stow, A. Feeling the pressure at home: Predator activity at the burrow entrance of an endangered arid-zone skink. Austral. Ecol. 2018, 43, 102–109. [Google Scholar] [CrossRef]
- Ridley, J.C.H.; Schlesinger, C.A.; Bull, C.M. Location of long-term communal burrows of a threatened arid-zone lizard in relation to soil and vegetation. Austral. Ecol. 2020, 45, 444–453. [Google Scholar] [CrossRef]
- Ridley, J.C.H.; Schlesinger, C.A. Activity of tjakuṟa (great desert skinks) at burrows in relation to plant cover and predators. Ecol. Evol. 2023, 13, e10391. [Google Scholar] [CrossRef]
- Halliwell, B.; Uller, T.; Wapstra, E.; While, G.M. Resource distribution mediates social and mating behavior in a family living lizard. Behav. Ecol. 2017, 28, 145–153. [Google Scholar] [CrossRef]
- Halliwell, B.; Uller, T.; Chapple, D.G.; Gardner, M.G.; Wapstra, E.; While, G.M. Habitat saturation promotes delayed dispersal in a social reptile. Behav. Ecol. 2017, 28, 515–522. [Google Scholar] [CrossRef]
- Pavey, C.R.; Burwell, C.J.; Nano, C.E.M. Foraging ecology and habitat use of Slater’s skink (Egernia slateri): An endangered Australian desert skink. J. Herpetol. 2010, 44, 563–571. [Google Scholar] [CrossRef]
- Greenville, A.C.; Dickman, C.R. Factors affecting habitat selection in a specialist fossorial skink. Biol. J. Linn. Soc. 2009, 97, 531–544. [Google Scholar] [CrossRef]
- Masters, P.; Dickman, C.R.; Crowther, M. Effects of cover reduction on mulgara Dasycercus cristicauda (Marsupialia: Dasyuridae), rodent and invertebrate populations in central Australia: Implications for land management. Austral. Ecol. 2003, 28, 658–665. [Google Scholar] [CrossRef]
- Moseby, K.E.; McGregor, H.M. Feral cats use fine scale prey cues and microhabitat patches of dense vegetation when hunting prey in arid Australia. Glob. Ecol. Conserv. 2022, 35, e02093. [Google Scholar] [CrossRef]
- Griffiths, A.D.; Christian, K.A. Diet and habitat use of frillneck lizards in a seasonal tropical environment. Oecologia 1996, 106, 39–48. [Google Scholar] [CrossRef]
- Loveridge, J.P.; Moe, S.R. Termitaria as browsing hotspots for African megaherbivores in miombo woodland. J. Trop. Ecol. 2004, 20, 337–343. [Google Scholar] [CrossRef]
- Fleming, P.A.; Loveridge, J.P. Miombo woodland termite mounds: Resource islands for small vertebrates? J. Zool. Soc. Lond. 2003, 259, 161–168. [Google Scholar] [CrossRef]
- Vitt, L.J.; Shepard, D.B.; Caldwell, J.P.; Vieira, G.H.C.; França, F.G.R.; Colli, G.R. Living with your food: Geckos in termitaria in Cantão. J. Zool. 2007, 272, 321–328. [Google Scholar] [CrossRef]
- Moro, D.; Cullen, P.; Fletcher, J. Vertebrate fauna in termite mounds compared to surrounding vegetation on Barrow Island. Pac. Conserv. Biol. 2014, 20, 296–301. [Google Scholar] [CrossRef]
- Thompson, G.G.; Thompson, S.A. Termitaria are an important refuge for reptiles in the Pilbara of Western Australia. Pac. Conserv. Biol. 2015, 21, 226–233. [Google Scholar] [CrossRef]
- Whittingham, M.J.; Evans, K.L. The effects of habitat structure on predation risk of birds in agricultural landscapes. Ibis 2004, 146, 210–220. [Google Scholar] [CrossRef]
- Risbey, D.A.; Calver, M.C.; Short, J.; Bradley, J.S.; Wright, I.W. The impact of cats and foxes on the small vertebrate fauna of Heirisson Prong, Western Australia. II. A field experiment. Wildl. Res. 2000, 27, 223–235. [Google Scholar] [CrossRef]
- Denny, E.A.; Dickman, C.R. Review of Cat Ecology and Management Strategies in Australia; Invasive Animals Cooperative Research Centre: Canberra, Australia, 2010. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eifler, M.A.; Eifler, D.A. Ecological Factors Associated with Burrow System Occupancy by Great Desert Skinks (Liopholis kintorei). Diversity 2025, 17, 134. https://doi.org/10.3390/d17020134
Eifler MA, Eifler DA. Ecological Factors Associated with Burrow System Occupancy by Great Desert Skinks (Liopholis kintorei). Diversity. 2025; 17(2):134. https://doi.org/10.3390/d17020134
Chicago/Turabian StyleEifler, Maria A., and Douglas A. Eifler. 2025. "Ecological Factors Associated with Burrow System Occupancy by Great Desert Skinks (Liopholis kintorei)" Diversity 17, no. 2: 134. https://doi.org/10.3390/d17020134
APA StyleEifler, M. A., & Eifler, D. A. (2025). Ecological Factors Associated with Burrow System Occupancy by Great Desert Skinks (Liopholis kintorei). Diversity, 17(2), 134. https://doi.org/10.3390/d17020134