
Bat Activity in South Mediterranean Forests: A Seasonal Acoustic Survey

 , , ,
, , ,
Abstract
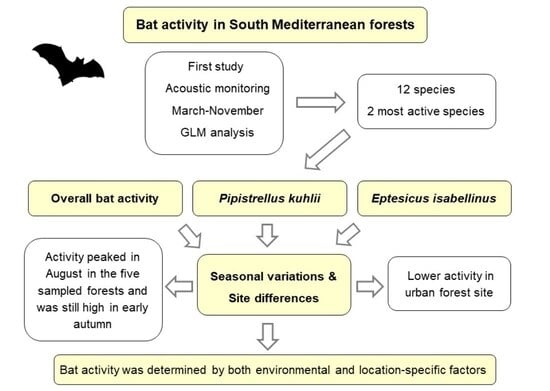
1. Introduction
2. Material and Methods
2.1. Study Areas
2.2. Acoustic Sampling
2.3. Data Analysis
3. Results
4. Discussion
4.1. Bat Assemblage Composition
4.2. Spatial and Temporal Variation in Bat Activity
4.3. Spatial and Temporal Activity of P. kuhlii and E. isabellinus
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- FAO and Plan Bleu. State of Mediterranean Forests 2018; Food and Agriculture Organization of the United Nations: Roma, Italy; Plan Bleu: Marseille, France, 2018. [Google Scholar]
- Martín-Ortega, P.; Picard, N.; García-Montero, L.G.; Del Rio, S.; Penas, A.; Marchetti, M.; Lasserre, B.; Özdemir, E.; García-Robredo, F.; Pascual, C. Importance of Mediterranean forests. In State of Mediterranean Forests; Bourlion, N., Garavaglia, V., Picard, N., Eds.; Food and Agriculture Organization of the United Nations: Roma, Italy; Plan Bleu: Marseille, France, 2018; pp. 31–50. [Google Scholar]
- FAO. State of Mediterranean Forests 2013; Food and Agriculture Organization of the United Nations: Roma, Italy; Plan Bleu: Marseille, France, 2013. [Google Scholar]
- Bentouati, A.; Oudjehih, B.; Alatou, D. Croissance en hauteur dominante et classes de fertilité du pin d’Alep (Pinus halepensis Mill.) dans le massif de Ouled-Yakoub et des Benioudjana (Khenchela–Aures). Sci. Technol. C 2005, 23, 57–62. [Google Scholar]
- Kadik, B. Contribution à l’étude du pin d’Alep (Pinus halepensis mill.) en Algérie: Écologie, dendrométrie, morphologie. Ph.D. Thesis, Université Paul Cézanne, Aix-Marseille, France, 1983. [Google Scholar]
- Dahmani-Megrerouche, M. Typologie et dynamique des chênaies vertes en Algérie. Forêt Méditerranéenne 2002, 23, 117–132. [Google Scholar]
- Simmons, N.B.; Cirranello, A.L. Bats of the World. A Taxonomic and Geographic Database. Version 1.9. 2025. Available online: https://batnames.org/ (accessed on 10 March 2025).
- Kehal, L.; Zouidi, M.; Keddari, D.; Hadjout, S.; Borsali, A.H. Comparative analysis of forest soil properties from sub-humid and semi-arid areas of Djebel El Ouahch biological reserve in Algeria. S. Asian J. Exp. Biol. 2022, 12, 349–356. [Google Scholar] [CrossRef]
- Rodríguez-San Pedro, A.; Simonetti, J.A. The relative influence of forest loss and fragmentation on insectivorous bats: Does the type of matrix matter? Landsc. Ecol. 2015, 30, 1561–1572. [Google Scholar] [CrossRef]
- Russo, D.; Billington, G.; Bontadina, F.; Dekker, J.; Dietz, M.; Gazaryan, S.; Jones, G.; Meschede, A.; Rebelo, H.; Reiter, G. Identifying key research objectives to make European forests greener for bats. Front. Ecol. Evol. 2016, 4, 87. [Google Scholar] [CrossRef]
- Langridge, J.; Pisanu, B.; Laguet, S.; Archaux, F.; Tillon, L. The role of complex vegetation structures in determining hawking bat activity in temperate forests. For. Ecol. Manag. 2019, 448, 559–571. [Google Scholar] [CrossRef]
- Brigham, R.M. Bats in forests: What we know and what we need to learn. In Bats in Forests. Conservation and Management; Lacki, M.J., Hayes, J.P., Kurta, A., Eds.; The Johns Hopkins University Press: Baltimore, MD, USA, 2007; pp. 1–15. [Google Scholar]
- Kunz, T.H.; Braun de Torrez, E.; Bauer, D.; Lobova, T.; Fleming, T.H. Ecosystem services provided by bats. Ann. N. Y. Acad. Sci. 2011, 1223, 1–38. [Google Scholar] [CrossRef]
- Lacki, M.J.; Johnson, J.S.; Dodd, L.E.; Baker, M.D. Prey consumption of insectivorous bats in coniferous forests of North-Central Idaho. Northwest Sci. 2007, 81, 199–205. [Google Scholar] [CrossRef]
- Böhm, S.M.; Wells, K.; Kalko, E.K.V. Top-down control of herbivory by birds and bats in the canopy of temperate broad-leaved oaks (Quercus robur). PLoS ONE 2011, 6, e17857. [Google Scholar] [CrossRef]
- Aulagnier, S.; Haffner, P.; Mitchell-Jones, A.J.; Moutou, F.; Zima, J. Mammifères d’Europe, d’Afrique du Nord et du Moyen-Orient, 6th ed.; Delachaux et Niestlé: Paris, France, 2024. [Google Scholar]
- Ahmim, M. Écologie et biologie de la conservation des Chiroptères de la région de la Kabylie des Babors (Algérie). Ph.D. Thesis, Université Abderrahmane Mira, Bejaia, Algeria, 2014. [Google Scholar]
- Dalhoumi, R. Les Chiroptères du Parc National de Bou Hedma: Cycle annuel et utilisation de l’habitat. Ph.D. Thesis, Université de Carthage, Bizerte, Tunisia, 2016. [Google Scholar]
- Loumassine, H.-E. Écologie des Chiroptères dans quelques biotopes en Algérie Occidentale. Ph.D. Thesis, Université Ibn Khaldoun, Tiaret, Algeria, 2018. [Google Scholar]
- Russo, D.; Jones, G. Use of foraging habitats by bats in a Mediterranean area determined by acoustic surveys: Conservation implications. Ecography 2003, 26, 197–209. [Google Scholar] [CrossRef]
- Rainho, A. Summer foraging habitats of bats in a Mediterranean Region of the Iberian Peninsula. Acta Chiropterol. 2007, 9, 171–181. [Google Scholar] [CrossRef]
- Novella-Fernandez, R.; Juste, J.; Ibañez, C.; Nogueras, J.; Osborne, P.E.; Razgour, O. The role of forest structure and composition in driving the distribution of bats in Mediterranean Regions. Sci. Rep. 2022, 12, 3224. [Google Scholar] [CrossRef] [PubMed]
- Dalhoumi, R.; Hedfi, A.; Aissa, P.; Aulagnier, S. Bats of Jebel Mghilla National Park (Central Tunisia): First survey and habitat-related activity. Trop. Zool. 2014, 27, 53–62. [Google Scholar] [CrossRef]
- Allegrini, C.; Korine, C.; Krasnov, B.R. Insectivorous bats in Eastern Mediterranean Planted pine forests—Effects of forest structure on foraging activity, diversity, and implications for management practices. Forests 2022, 13, 1411. [Google Scholar] [CrossRef]
- Dbiba, Y.; Dakki, M.; Mansouri, I.; El Mouden, E.H. Bats of the Safi-Essaouira Provinces (Morocco): New Inputs to the knowledge of bat populations of the Atlantic littoral. Int. J. Zool. 2023, 2023, 6624548. [Google Scholar] [CrossRef]
- Kaci, K.; Ahmim, M.; Dalhoumi, R. Winter habitat use and nocturnal activity of the Kuhl’s pipistrelle (Pipistrellus kuhlii) and the European free-tailed bat (Tadarida teniotis) in a Mediterranean region (Algeria). Zool. Ecol. 2024, 34, 169–179. [Google Scholar] [CrossRef]
- Ait Abdesselam, M.; Dalhoumi, R.; Ouabed, A.; Farid, B.; Aulagnier, S. Bat Species richness and activity in a Mediterranean area of Northwest Africa: Urban–rural shifts. Anim. Taxon. Ecol. 2025, 71, 36–50. [Google Scholar] [CrossRef]
- Zamora-Gutierrez, V.; MacSwiney, G.M.C.; Martínez Balvanera, S.; Robredo Esquivelzeta, E. The evolution of acoustic methods for the study of bats. In 50 Years of Bat Research: Foundations and New Frontiers; Springer: Berlin/Heidelberg, Germany, 2021; pp. 43–59. [Google Scholar]
- Neuweiler, G. The Biology of Bats; Oxford University Press: New York, NJ, USA, 2000. [Google Scholar]
- Ziouche, S.; Baali, F.; Moutassem, D.; Djazouli, Z. Stratégies de choix de l’emplacement des nids d’hiver de Thaumetopoea pityocampa (Denis & Schiffermüller, 775) au niveau de trois pinèdes dans la région de Bordj Bou Arreridj (Algérie). Agrobiologia 2017, 7, 412–426. [Google Scholar]
- Bouttaba, C.; Nouibat, B.; Benmechiche, M. Peri-urban forests: An exploratory study of users’ recreational activities: The case of the El Meridj-Est recreational forest in Constantine, Algeria. J. Degrad. Min. Lands Manag. 2024, 11, 5695–5706. [Google Scholar] [CrossRef]
- Bourenane, H.; Bouhadad, Y. Impact of land use changes on landslides occurrence in urban area: The case of the Constantine City (NE Algeria). Geotech. Geol. Eng. 2021, 39, 1–21. [Google Scholar] [CrossRef]
- Lemouissi, S. Approche phytosociologique de la végétation dans le massif forestier de Chettabah (Constantine). Master’s Thesis, Université Frères Mentouri, Constantine, Algeria, 2014. [Google Scholar]
- Kanouni, M.R.; Hani, I.; Bousba, R.; Beldjazia, A.; Khamar, H. Structural variability of Aleppo pine stands in two forests in Northeastern Algeria. Biodiversitas J. Biol. Divers. 2020, 21, 2848–2853. [Google Scholar] [CrossRef]
- Russ, J. Bat Calls of Britain and Europe; Pelagic Publishing: Exeter, UK, 1999. [Google Scholar]
- Barataud, M. Écologie Acoustique des Chiroptères d’Europe; Biotope: Mèze, France; Muséum National d’Histoire Naturelle: Paris, France, 2012. [Google Scholar]
- Disca, T.; Allegrini, B.; Prié, V. Caractéristiques acoustiques des cris d’écholocation de 16 espèces de Chiroptères (Mammalia, Chiroptera) du Maroc. Vespère 2014, 3, 209–229. [Google Scholar]
- Dalhoumi, R.; Aïssa, P.; Aulagnier, S. Bat species richness and activity in Bou Hedma National Park (Central Tunisia). Turk. J. Zool. 2016, 40, 864–875. [Google Scholar] [CrossRef]
- Ahmim, M.; Dalhoumi, R.; Măntoiu, D.S. First data on the acoustic characteristics of some chiropteran species from Northern Algeria. Bioacoustics 2020, 29, 499–517. [Google Scholar] [CrossRef]
- The R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2024; Available online: http://www.R-project.org/ (accessed on 18 November 2024).
- Brooks, M.E.; Kristensen, K.; Van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Mächler, M.; Bolker, B.M. glmm TMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- Sugiura, N. Further analysis of the data by Akaike’s information criterion and the finite corrections: Further analysis of the data by Akaike’s. Commun. Stat.-Theory Methods 1978, 7, 13–26. [Google Scholar] [CrossRef]
- Hurvich, C.M.; Tsai, C.-L. Regression and time series model selection in small samples. Biometrika 1989, 76, 297–307. [Google Scholar] [CrossRef]
- Mazerolle, M.J.; Mazerolle, M.M.J. Package ‘AICcmodavg’. 2020. Available online: https://cran.uib.no/web/packages/AICcmodavg/AICcmodavg.pdf (accessed on 14 January 2025).
- Hartig, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Available online: https://cran.r-project.org/web/packages/DHARMa/vignettes/DHARMa.html (accessed on 14 January 2025).
- Lüdecke, D.; Ben-Shachar, M.S.; Patil, I.; Waggoner, P.; Makowski, D. Performance: An R package for assessment, comparison and testing of statistical models. J. Open Source Softw. 2021, 6, 3139. [Google Scholar] [CrossRef]
- Gori, M.; Betti, A.; Melacci, S. Machine Learning. A Constraint-Based Approach, 2nd ed.; Morgan Kaufmann Publishers: Cambridge, MA, USA, 2024. [Google Scholar]
- Lisón, F.; Calvo, J.F. The significance of water infrastructures for the conservation of bats in a semiarid Mediterranean landscape. Anim. Conserv. 2011, 14, 533–541. [Google Scholar] [CrossRef]
- Dalhoumi, R.; Nefla, A.; Bedoui, W.; Ouni, R.; Aissa, P.; Aulagnier, S. Preliminary study of habitat-related bat fauna of Mastouta-Bishshouk region (Northwest Tunisia). Zoodiversity 2019, 53, 23–30. [Google Scholar] [CrossRef]
- Tillon, L.; Barataud, M.; Giosa, S.; Aulagnier, S. Acoustic detection of radiotracked foraging bats in temperate lowland forests. Mamm. Biol. 2019, 95, 155–159. [Google Scholar] [CrossRef]
- Dekker, J.; Steen, W.; Bouman, H.B.; van der Vliet, R.E. Differences in acoustic detectability of bat species hamper environmental impact assessment studies. Eur. J. Wildl. Res. 2022, 68, 14. [Google Scholar] [CrossRef]
- Teets, K.D.; Loeb, S.C.; Jachowski, D.S. Detection probability of bats using active versus passive monitoring. Acta Chiropterol. 2019, 21, 205–213. [Google Scholar] [CrossRef]
- Perks, S.J.; Goodenough, A.E. Comparing acoustic survey data for European bats: Do walked transects or automated fixed-point surveys provide more robust data? Wildl. Res. 2021, 49, 314–323. [Google Scholar] [CrossRef]
- Fisher-Phelps, M.; Schwilk, D.; Kingston, T. Mobile acoustic transects detect more bat activity than stationary acoustic point counts in a semi-arid and agricultural landscape. J. Arid. Environ. 2017, 136, 38–44. [Google Scholar] [CrossRef]
- Fischer, J.; Stott, J.; Law, B.S.; Adams, M.D.; Forrester, R.I. Designing effective habitat studies: Quantifying multiple sources of variability in bat activity. Acta Chiropterol. 2009, 11, 127–137. [Google Scholar] [CrossRef]
- Skalak, S.L.; Sherwin, R.E.; Brigham, R.M. Sampling period, size and duration influence measures of bat species richness from acoustic surveys. Methods Ecol. Evol. 2012, 3, 490–502. [Google Scholar] [CrossRef]
- Kubista, C.E.; Bruckner, A. Within-site variability of field recordings from stationary, passively working detectors. Acta Chiropterol. 2017, 19, 189–197. [Google Scholar] [CrossRef]
- Froidevaux, J.S.; Zellweger, F.; Bollmann, K.; Obrist, M.K. Optimizing passive acoustic sampling of bats in forests. Ecol. Evol. 2014, 4, 4690–4700. [Google Scholar] [CrossRef]
- Tena, E.; Tellería, J. Modelling the distribution of bat activity areas for conservation in a Mediterranean mountain range. Anim. Conserv. 2022, 25, 65–76. [Google Scholar] [CrossRef]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Aulagnier, S.; Cuzin, F.; Thévenot, M. Mammifères Sauvages du Maroc. Peuplement, Répartition, Écologie; Société Française pour l’Étude et la Protection des Mammifères: Paris, France, 2017. [Google Scholar]
- Dalhoumi, R.; Aissa, P.; Aulagnier, S. Taxonomie et répartition des Chiroptères de Tunisie. Rev. Suisse Zool. 2011, 118, 265–292. [Google Scholar] [CrossRef]
- Puechmaille, S.J.; Hizem, W.M.; Allegrini, B.; Abiadh, A. Bat fauna of Tunisia: Review of records and new records, morphometrics and echolocation data. Vespertilio 2012, 16, 211–239. [Google Scholar]
- Ahmim, M.; Oubaziz, B. New data on the distribution of bats (Mammalia: Chiroptera) in Algeria. Acta Soc. Zool. Bohemicae 2017, 81, 17–30. [Google Scholar]
- Ancillotto, L.; Tomassini, A.; Russo, D. The fancy city life: Kuhl’s pipistrelle, Pipistrellus kuhlii, benefits from urbanisation. Wildl. Res. 2015, 42, 598–606. [Google Scholar] [CrossRef]
- Davy, C.M.; Russo, D.; Fenton, M.B. Use of native woodlands and traditional olive groves by foraging bats on a Mediterranean Island: Consequences for conservation. J. Zool. 2007, 273, 397–405. [Google Scholar] [CrossRef]
- Di Salvo, I.; Russo, D.; Sarà, M. Habitat preferences of bats in a rural area of Sicily determined by acoustic surveys. Hystrix 2009, 20, 137–146. [Google Scholar]
- Georgiakakis, P.; Vasilakopoulos, P.; Mylonas, M.; Russo, D. Bat species richness and activity over an elevation gradient in Mediterranean shrublands of Crete. Hystrix 2010, 21, 43–56. [Google Scholar]
- Herrera, J.M.; Costa, P.; Medinas, D.; Marques, J.T.; Mira, A. Community composition and activity of insectivorous bats in Mediterranean olive farms. Anim. Conserv. 2015, 18, 557–566. [Google Scholar] [CrossRef]
- Ancillotto, L.; Ariano, A.; Nardone, V.; Budinski, I.; Rydell, J.; Russo, D. Effects of free-ranging cattle and landscape complexity on bat foraging: Implications for bat conservation and livestock management. Agric. Ecosyst. Environ. 2017, 241, 54–61. [Google Scholar] [CrossRef]
- Tzortzakaki, O.; Papadatou, E.; Kati, V.; Giokas, S. Winners and losers in an urban bat community: A case study from Southeastern Europe. Hystrix 2019, 30, 134–140. [Google Scholar]
- Loumassine, H.E.; Bonnot, N.; Allegrini, B.; Bendjeddou, M.L.; Bounaceur, F.; Aulagnier, S. How arid environments affect spatial and temporal activity of bats. J. Arid. Environ. 2020, 180, 104206. [Google Scholar] [CrossRef]
- Dalhoumi, R.; Aïssa, P.; Hamouda, B.; Aulagnier, S. Bat species richness and activity in Dghoumes National Park (Southwest Tunisia): A preliminary survey. Arx. Miscel-Lània Zool. 2020, 18, 89–100. [Google Scholar] [CrossRef]
- Dalhoumi, R.; El Mokni, R.; Ouni, R.; Hamouda, B.; Aulagnier, S. Bats of the Tunisian desert: Preliminary data using acoustic identification and first record of Taphozous nudiventris in the country. Diversity 2023, 15, 1108. [Google Scholar] [CrossRef]
- Lisón, F.; Calvo, J. Bat activity over small ponds in dry Mediterranean forests: Implications for conservation. Acta Chiropterol. 2014, 16, 95–101. [Google Scholar] [CrossRef]
- Kipson, M.; Gazaryan, S.; Horáček, I. Savi’s pipistrelle Hypsugo savii (Bonaparte, 1837). In Handbook of the Mammals of Europe; Russo, D., Ed.; Springer Nature: Cham, Switzerland, 2025; pp. 95–112. [Google Scholar]
- Lisón, F.; Calvo, J.F. Ecological niche modelling of three pipistrelle bat species in semiarid Mediterranean Landscapes. Acta Oecol. 2013, 47, 68–73. [Google Scholar] [CrossRef]
- Froidevaux, J.; Zellweger, F.; Bollmann, K.; Jones, G.; Obrist, M.K. From field surveys to LiDAR: Shining a light on how bats respond to forest structure. Remote Sens. Environ. 2016, 175, 242–250. [Google Scholar] [CrossRef]
- Froidevaux, J.S.P.; Barbaro, L.; Vinet, O.; Larrieu, L.; Bas, Y.; Molina, J.; Calatayud, F.; Brin, A. Bat responses to changes in forest composition and prey abundance depend on landscape matrix and stand structure. Sci. Rep. 2021, 11, 10586. [Google Scholar] [CrossRef]
- Bender, M.J.; Perea, S.; Castleberry, S.B.; Miller, D.A.; Wigley, T.B. Influence of Insect abundance and vegetation structure on site-occupancy of bats in managed pine forests. For. Ecol. Manag. 2021, 482, 118839. [Google Scholar] [CrossRef]
- Perea, S.; Vicente-Santos, A.; Larsen-Gray, A.L.; Gandhi, K.J.; Greene, D.U.; Barnes, B.F.; Castleberry, S.B. Disentangling winter relationships: Bat responses to forest stand structure, environmental conditions, and prey composition. For. Ecol. Manag. 2025, 578, 122484. [Google Scholar] [CrossRef]
- Charbonnier, Y.M.; Barbaro, L.; Barnagaud, J.-Y.; Ampoorter, E.; Nezan, J.; Verheyen, K.; Jactel, H. Bat and bird diversity along independent gradients of latitude and tree composition in European forests. Oecologia 2016, 182, 529–537. [Google Scholar] [CrossRef]
- Tibbels, A.E.; Kurta, A. Bat activity is low in thinned and unthinned stands of red pine. Can. J. For. Res. 2003, 33, 2436–2442. [Google Scholar] [CrossRef]
- Węgiel, A.; Grzywiński, W.; Ciechanowski, M.; Jaros, R.; Kalcounis-Rüppell, M.; Kmiecik, A.; Kmiecik, P.; Węgiel, J. The foraging activity of bats in managed pine forests of different ages. Eur. J. For. Res. 2019, 138, 383–396. [Google Scholar] [CrossRef]
- Charbonnier, Y.; Barbaro, L.; Theillout, A.; Jactel, H. Numerical and functional responses of forest bats to a major insect pest in pine plantations. PLoS ONE 2014, 9, e109488. [Google Scholar] [CrossRef] [PubMed]
- Morris, A.D.; Miller, D.A.; Kalcounis-Rueppell, M.C. Use of forest edges by bats in a managed pine forest landscape. J. Wildl. Manag. 2010, 74, 26–34. [Google Scholar] [CrossRef]
- Kalcounis, M.C.; Brigham, R.M. Intraspecific variation in wing loading affects habitat use by little brown bats (Myotis lucifugus). Can. J. Zool. 1995, 73, 89–95. [Google Scholar] [CrossRef]
- Menzel, J.M.; Menzel, M.A., Jr.; Kilgo, J.C.; Ford, W.M.; Edwards, J.W.; McCracken, G.F. Effect of habitat and foraging height on bat activity in the coastal plain of South Carolina. J. Wildl. Manag. 2005, 69, 235–245. [Google Scholar] [CrossRef]
- Müller, J.; Mehr, M.; Bässler, C.; Fenton, M.B.; Hothorn, T.; Pretzsch, H.; Klemmt, H.-J.; Brandl, R. Aggregative response in bats: Prey abundance versus habitat. Oecologia 2012, 169, 673–684. [Google Scholar] [CrossRef]
- Dodd, L.E.; Lacki, M.J.; Britzke, E.R.; Buehler, D.A.; Keyser, P.D.; Larkin, J.L.; Rodewald, A.D.; Wigley, T.B.; Wood, P.B.; Rieske, L.K. Forest structure affects trophic linkages: How silvicultural disturbance impacts bats and their insect prey. For. Ecol. Manag. 2012, 267, 262–270. [Google Scholar] [CrossRef]
- Titchenell, M.A.; Williams, R.A.; Gehrt, S.D. Bat response to shelterwood harvests and forest structure in oak-hickory forests. For. Ecol. Manag. 2011, 262, 980–988. [Google Scholar] [CrossRef]
- Stone, E.L.; Harris, S.; Jones, G. Impacts of artificial lighting on bats: A review of challenges and solutions. Mamm. Biol. 2015, 80, 213–219. [Google Scholar] [CrossRef]
- Gaston, K.J.; Bennie, J.; Davies, T.W.; Hopkins, J. The ecological impacts of nighttime light pollution: A mechanistic appraisal. Biol. Rev. 2013, 88, 912–927. [Google Scholar] [CrossRef] [PubMed]
- Schaub, A.; Ostwald, J.; Siemers, B.M. Foraging bats avoid noise. J. Exp. Biol. 2008, 211, 3174–3180. [Google Scholar] [CrossRef]
- Florencio, M.; Serrano, L.; Gómez-Rodríguez, C.; Millán, A.; Díaz-Paniagua, C. Inter-and intra-annual variations of macroinvertebrate assemblages are related to the hydroperiod in Mediterranean temporary ponds. Hydrobiologia 2009, 634, 167–183. [Google Scholar] [CrossRef]
- Florencio, M.; Díaz-Paniagua, C.; Serrano, L.; Bilton, D.T. Spatio-temporal nested patterns in macroinvertebrate assemblages across a pond network with a wide hydroperiod range. Oecologia 2011, 166, 469–483. [Google Scholar] [CrossRef]
- Gaisler, J.; Madkour, G.; Pelikán, J. On the bats (Chiroptera) of Egypt. Československá akademie věd v Brně. Přírodovědné práce Ústavů Československé akademie vĕd v Brnĕ Acta scientiarum naturalium Academiae scientiarum Bohemoslovacae Brno. Ser. Nova 1972, 6, 1–40. [Google Scholar]
- Dalhoumi, R.; Morellet, N.; Aissa, P.; Aulagnier, S. Seasonal activity pattern and habitat use by the Kuhl’s pipistrelle (Pipistrellus kuhlii) in an arid environment. Eur. J. Wildl. Res. 2018, 64, 36. [Google Scholar] [CrossRef]
- Yekwayo, I.; Pryke, J.S.; Roets, F.; Samways, M.J. Surrounding vegetation matters for arthropods of small, natural patches of indigenous forest. Insect Conserv. Divers. 2016, 9, 224–235. [Google Scholar] [CrossRef]
- Uhler, J.; Redlich, S.; Zhang, J.; Hothorn, T.; Tobisch, C.; Ewald, J.; Thorn, S.; Seibold, S.; Mitesser, O.; Morinière, J. Relationship of insect biomass and richness with land use along a climate gradient. Nat. Commun. 2021, 12, 5946. [Google Scholar] [CrossRef]
- van Deventer, H.D.; Gaigher, R.; Pryke, J.S. Grassland arthropod assemblages differ between a variety of neighbouring biotopes and small features in a timber production landscape. Biodivers. Conserv. 2025, 34, 2939–2955. [Google Scholar] [CrossRef]
- Kalko, E.K.; Schnitzler, H.-U. Plasticity in echolocation signals of European pipistrelle bats in search flight: Implications for habitat use and prey detection. Behav. Ecol. Sociobiol. 1993, 33, 415–428. [Google Scholar] [CrossRef]
- Ancillotto, L.; Santini, L.; Ranc, N.; Maiorano, L.; Russo, D. Extraordinary range expansion in a common bat: The potential roles of climate change and urbanisation. Sci. Nat. 2016, 103, 15. [Google Scholar] [CrossRef]
- Korine, C.; Pinshow, B. Guild structure, foraging space use, and distribution in a community of insectivorous bats in the Negev Desert. J. Zool. 2004, 262, 187–196. [Google Scholar] [CrossRef]
- Razgour, O.; Korine, C.; Saltz, D. Does interspecific competition drive patterns of habitat use in desert bat communities? Oecologia 2011, 167, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.; Whitaker, J.O.; Yom-Tov, Y. Dietary composition and habitat use in a desert insectivorous bat community in Israel. Acta Chiropterol. 2000, 2, 15–22. [Google Scholar]
- Speakman, J.R.; Thomas, D.W. Physiological ecology and energetics of bats. In Bat Ecology; Kunz, T.H., Fenton, M.B., Eds.; The University of Chicago Press: Chicago, MI, USA, 2003; pp. 430–490. [Google Scholar]
- Sharifi, M.; Vaissi, S.; Javanbakht, H.; Akmali, V. Postnatal growth and wing development in Kuhl’s pipistrelle Pipistrellus kuhlii (Chiroptera: Vespertilionidae) in captivity. Zool. Stud. 2012, 51, 1235–1247. [Google Scholar]
- Benda, P.; Lucan, R.K.; Obuch, J.; Reiter, A.; Andreas, M.; Backor, P.; Bohnenstengel, T.; Eid, E.K.; Sevcik, M.; Vallo, P. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 8. Bats of Jordan: Fauna, ecology, echolocation, ectoparasites. Acta Soc. Zool. Bohemicae 2010, 74, 185–353. [Google Scholar]
- Benda, P.; Spitzenberger, F.; Hanák, V.; Andreas, M.; Reiter, A.; Ševčík, M.; Šmíd, J.; Uhrin, M. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean and Middle East. Part 11. On the bat fauna of Libya II. Acta Soc. Zool. Bohemicae 2014, 78, 1–162. [Google Scholar]
- Benda, P.; Andreas, M.; Kock, D.; Lucan, R.K.; Munclinger, P.; Nova, P.; Obuch, J.; Ochman, K.; Reiter, A.; Uhrin, M. Bats (Mammalia: Chiroptera) of the Eastern Mediterranean. Part 4. Bat Fauna of Syria: Distribution, systematics, ecology. Acta Soc. Zool. Bohemicae 2006, 70, 1–329. [Google Scholar]
- Lewis, R.E.; Harrison, D.L. Notes on bats from the Republic of Lebanon. Proc. Zool. Soc. Lond. 1962, 138, 473–486. [Google Scholar] [CrossRef]
- Alagaili, A. Biological, Ecological, and Conservational Study of Kuhl’s Bat (Pipistrellus kuhlii) from Unizah Province, Saudi Arabia. Ph.D. Thesis, University of Arkansas, Fayetteville, NC, USA, 2008. [Google Scholar]
- Alagaili, A.N.; James, D.A.; Mohammed, O.B. Timing and pattern of molt in Kuhl’s bat, Pipistrellus kuhlii, in Saudi Arabia. Acta Chiropterol. 2011, 13, 465–470. [Google Scholar] [CrossRef]
- Kelm, D.H.; Popa-Lisseanu, A.G.; Dehnhard, M.; Ibáñez, C. Non-invasive monitoring of stress hormones in the bat Eptesicus isabellinus–Do fecal glucocorticoid metabolite concentrations correlate with survival? Gen. Comp. Endocrinol. 2016, 226, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Lisón, F.; Haz, Á.; Calvo, J.F. Preferencia de hábitat del murciélago hortelano meridional Eptesicus isabellinus (Temminck, 1840) en ambientes mediterráneos semiáridos. Anim. Biodivers. Conserv. 2014, 37, 59–67. [Google Scholar] [CrossRef]
- Pérez Jordá, J.L. Ecología del murciélago hortelano, Eptesicus serotinus, en Andalucía. Ph.D. Thesis, Universidad de Sevilla, Sevilla, Spain, 1994. [Google Scholar]
- Lisón, F.; Espinosa, J.A.L.; Calvo, J.F.; Jones, G. Diet of the meridional serotine Eptesicus isabellinus in an urban semiarid Mediterranean landscape. Acta Chiropterol. 2015, 17, 371–378. [Google Scholar] [CrossRef]
- Dalhoumi, R.; Morellet, N.; Aissa, P.; Aulagnier, S. Seasonal activity pattern and habitat use by the isabelline serotine bat (Eptesicus isabellinus) in an arid environment of Tunisia. Acta Chiropterol. 2017, 19, 141–153. [Google Scholar] [CrossRef]
- Baker, R.J.; Davis, B.L.; Jordan, R.G.; Binous, A. Karyotypic and morphometric studies of Tunisian mammals: Bats. Mammalia 1974, 38, 695–705. [Google Scholar] [CrossRef]
- Loeb, S.C.; Waldrop, T.A. Bat activity in relation to fire and fire surrogate treatments in southern pine stands. For. Ecol. Manag. 2008, 255, 3185–3192. [Google Scholar] [CrossRef]
- López-Baucells, A.; Flaquer, C.; Mas, M.; Puig-Montserrat, X. Recurring fires in Mediterranean habitats and their impact on bats. Biodivers. Conserv. 2021, 30, 385–402. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Coordinates | Characteristics |
|---|---|---|
| Bordj Zemoura | 36°17′15.8″ N 4°49′29.5″ E | Open forest, with a density of 0.08 tree/m2 at an altitude of 930 m, including villages, and agricultural lands. Tree height ranges from 2 to 6 m. The forest covers an area of 658.99 km2 composed in majority of Aleppo pines, and less substantially of holm oaks. |
| Boumergad | 36°3′32.46″ N 4°48′41.51″ E | Moderately dense forest (0.14 tree/m2) at an altitude of 1009 m, in the midst of the city, surrounded by urban habitats and agricultural lands. Tree height ranges from 2 to 7 m. A small forest of 7 km2, composed of Aleppo pines. |
| Djaafra | 36°16′49.75″ N 4°38′18.51″ E | Dense forest (0.2 tree/m2) at an altitude of 1200 m, including some villages, roads and agricultural lands. Tree height ranges from 3 and 13 m. The forest covers an area of 1132.77 km2 composed in majority of Aleppo pines and less substantially Atlas cedars and holm oaks. |
| Chettaba Aleppo Pine Forest | 36°17′57.2″ N 6°27′22.1″ E | Dense forest (0.18 tree per m2) at an altitude of 866 m, including agricultural lands. Tree height ranges from 1 to 3 m. Composed of Aleppo pine trees. |
| Chettaba Holm Oak Forest | 36°19′16.9″ N 6°27′34.7″ E | Dense maquis of holm oak trees (0.22 tree per m2) at an altitude of 991 m. Tree height ranges from 5 to 13 m. |
| Species | Bordj Zemoura | Boumergad | Djaafra | Chettaba Aleppo Pine | Chettaba Holm Oak |
|---|---|---|---|---|---|
| Rhinolophus blasii | 1.14 | 0.00 | 0.00 | 0.00 | 0.42 |
| Rhinolophus hipposideros | 1.14 | 0.00 | 0.00 | 0.70 | 1.27 |
| Tadarida teniotis | 0.38 | 4.61 | 6.16 | 19.16 | 15.25 |
| Miniopterus schreibersii | 0.38 | 0.00 | 1.31 | 5.92 | 2.12 |
| Eptesicus isabellinus | 37.57 | 2.63 | 34.89 | 6.97 | 4.24 |
| Hypsugo savii | 5.12 | 0.66 | 25.00 | 4.18 | 5.93 |
| Pipistrellus kuhlii | 49.53 | 90.13 | 30.04 | 50.17 | 63.14 |
| Pipipistrellus pipipistrellus | 2.85 | 1.32 | 0.37 | 6.97 | 1.27 |
| Plecotus gaisleri | 1.71 | 0.00 | 0.37 | 0.00 | 0.00 |
| Myotis capaccinii | 0.19 | 0.66 | 1.87 | 0.00 | 0.42 |
| Myotis emarginatus | 0.00 | 0.00 | 0.00 | 0.57 | 0.00 |
| Myotis punicus | 0.00 | 0.00 | 0.00 | 2.66 | 2.66 |
| Models (Overall Bat Activity) | K | AICc | ΔAICc | AICcWt |
|---|---|---|---|---|
| Site + Month | 15 | 1114.33 | 0.00 | 0.85 |
| Site × Temperature | 12 | 1117.82 | 3.50 | 0.15 |
| Region × Month + Period | 23 | 1123.57 | 9.24 | 0.01 |
| Region × Month | 20 | 1127.21 | 12.88 | 0.00 |
| Region + Month | 12 | 1129.59 | 15.26 | 0.00 |
| Site + Temperature | 8 | 1131.26 | 16.93 | 0.00 |
| Site × Month + Period | 50 | 1134.91 | 20.58 | 0.00 |
| Site × Month | 47 | 1135.58 | 21.25 | 0.00 |
| Region × Temperature | 6 | 1136.14 | 21.82 | 0.00 |
| Region + Temperature | 5 | 1140.67 | 26.34 | 0.00 |
| Site × Hygrometry | 12 | 1151.02 | 36.70 | 0.00 |
| Site × Hygrometry | 8 | 1152.10 | 37.77 | 0.00 |
| Site | 7 | 1155.73 | 41.40 | 0.00 |
| Region + Hygrometry | 5 | 1159.21 | 44.88 | 0.00 |
| Region × Hygrometry | 6 | 1160.26 | 45.94 | 0.00 |
| Region | 4 | 1164.63 | 50.31 | 0.00 |
| Null | 2 | 1170.57 | 56.24 | 0.00 |
| Models (P. kuhlii) | K | AICc | ΔAICc | AICcWt |
|---|---|---|---|---|
| Region × Month + Period | 23 | 865.37 | 0.00 | 0.55 |
| Site + Month | 15 | 866.18 | 0.82 | 0.37 |
| Region × Month | 20 | 869.86 | 4.49 | 0.06 |
| Region + Month | 12 | 872.09 | 6.72 | 0.02 |
| Site × Temperature | 12 | 885.06 | 19.69 | 0.00 |
| Region × Temperature | 6 | 887.77 | 22.40 | 0.00 |
| Site × Month + Period | 50 | 889.72 | 24.36 | 0.00 |
| Site × Month | 47 | 891.28 | 25.92 | 0.00 |
| Region + Temperature | 5 | 891.88 | 26.52 | 0.00 |
| Site + Temperature | 8 | 893.99 | 28.63 | 0.00 |
| Region + Hygrometry | 5 | 913.59 | 48.23 | 0.00 |
| Site × Hygrometry | 12 | 915.44 | 50.07 | 0.00 |
| Region × Hygrometry | 6 | 915.68 | 50.32 | 0.00 |
| Region | 4 | 916.60 | 51.23 | 0.00 |
| Hygrometry + Site | 8 | 916.72 | 50.35 | 0.00 |
| Site | 7 | 918.32 | 52.96 | 0.00 |
| Null | 2 | 918.99 | 53.63 | 0.00 |
| Models | K | AICc | ΔAICc | AICcWt |
|---|---|---|---|---|
| Site + Month | 15 | 479.14 | 0.00 | 0.92 |
| Site + Temperature | 8 | 484.04 | 4.91 | 0.08 |
| Site | 7 | 498.75 | 19.61 | 0.00 |
| Region × Temperature | 6 | 507.98 | 28.84 | 0.00 |
| Region × Month | 20 | 514.33 | 35.19 | 0.00 |
| Region + Month | 12 | 514.36 | 35.23 | 0.00 |
| Region + Temperature | 5 | 516.66 | 37.52 | 0.00 |
| Site × Month | 47 | 520.78 | 41.64 | 0.00 |
| Region | 4 | 524.28 | 45.15 | 0.00 |
| Null | 2 | 542.85 | 63.72 | 0.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boukhari, A.; Ait-Abdesselam, M.; Bissaad, F.Z.; Bounaceur, F.; Khazar, I.; Bouhani, M.; Aulagnier, S. Bat Activity in South Mediterranean Forests: A Seasonal Acoustic Survey. Diversity 2025, 17, 750. https://doi.org/10.3390/d17110750
Boukhari A, Ait-Abdesselam M, Bissaad FZ, Bounaceur F, Khazar I, Bouhani M, Aulagnier S. Bat Activity in South Mediterranean Forests: A Seasonal Acoustic Survey. Diversity. 2025; 17(11):750. https://doi.org/10.3390/d17110750
Chicago/Turabian StyleBoukhari, Afaf, Moncef Ait-Abdesselam, Fatima Zohra Bissaad, Farid Bounaceur, Inès Khazar, Mouna Bouhani, and Stéphane Aulagnier. 2025. "Bat Activity in South Mediterranean Forests: A Seasonal Acoustic Survey" Diversity 17, no. 11: 750. https://doi.org/10.3390/d17110750
APA StyleBoukhari, A., Ait-Abdesselam, M., Bissaad, F. Z., Bounaceur, F., Khazar, I., Bouhani, M., & Aulagnier, S. (2025). Bat Activity in South Mediterranean Forests: A Seasonal Acoustic Survey. Diversity, 17(11), 750. https://doi.org/10.3390/d17110750