
The Population of the Glacial Relict Betula nana Surviving Anthropogenic Pressure (the Case of Šepeta Peatland in Northeastern Lithuania)

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
- (1)
- Cover and abundance of each plant species following the Braun–Blanquet scale [33].
- (2)
- Percentage cover of tree, shrub/subshrub, herb, and bryophyte layers and B. nana.
- (3)
- Physical and chemical parameters of peat (conductivity; pH; concentrations total nitrogen, NH4+, NO3−, P, K, Mg, Cl, organic material and organic carbon, peat decomposition degree, and peat moisture). Peat samples were selected from 10 randomly chosen sites, and a pooled sample (approximately 500 g of peat) was prepared. The analysis of peat samples was performed in the Agrochemical Research Laboratory of the Lithuanian Research Centre for Agriculture and Forestry. In the laboratory, the pH was measured using the potenciometrical method according to LSTEN 13037 [34]. Spectrophotometrical methods were used for the measurements of P, atomic emission spectral method for K, atomic absorption emission spectrometrical method for Ca and Mg, colourimetric method with FIA star for N-NO3−, N-NH4, and Cl was measured with the argentometrical method LSTEN 13652 [35]. The peat decomposition degree was ascertained according to LST 1957:2022 [36], organic material and peat moisture according to [37] LST EN 13040, and organic carbon according to ISO10694:19995 [38].
- (4)
- Water table. Free groundwater table before the ground surface was measured using 20 mm diameter PVC pipes perforated with 50 mm holes. The pipes were hammered down to a depth of 1 m from the surface of the peat at the centre of each study plot. After half an hour, the water table depth from the peat surface was measured with a stable measuring tape. If water did not appear at 1 m, its depth was estimated as >1 m.
- (1)
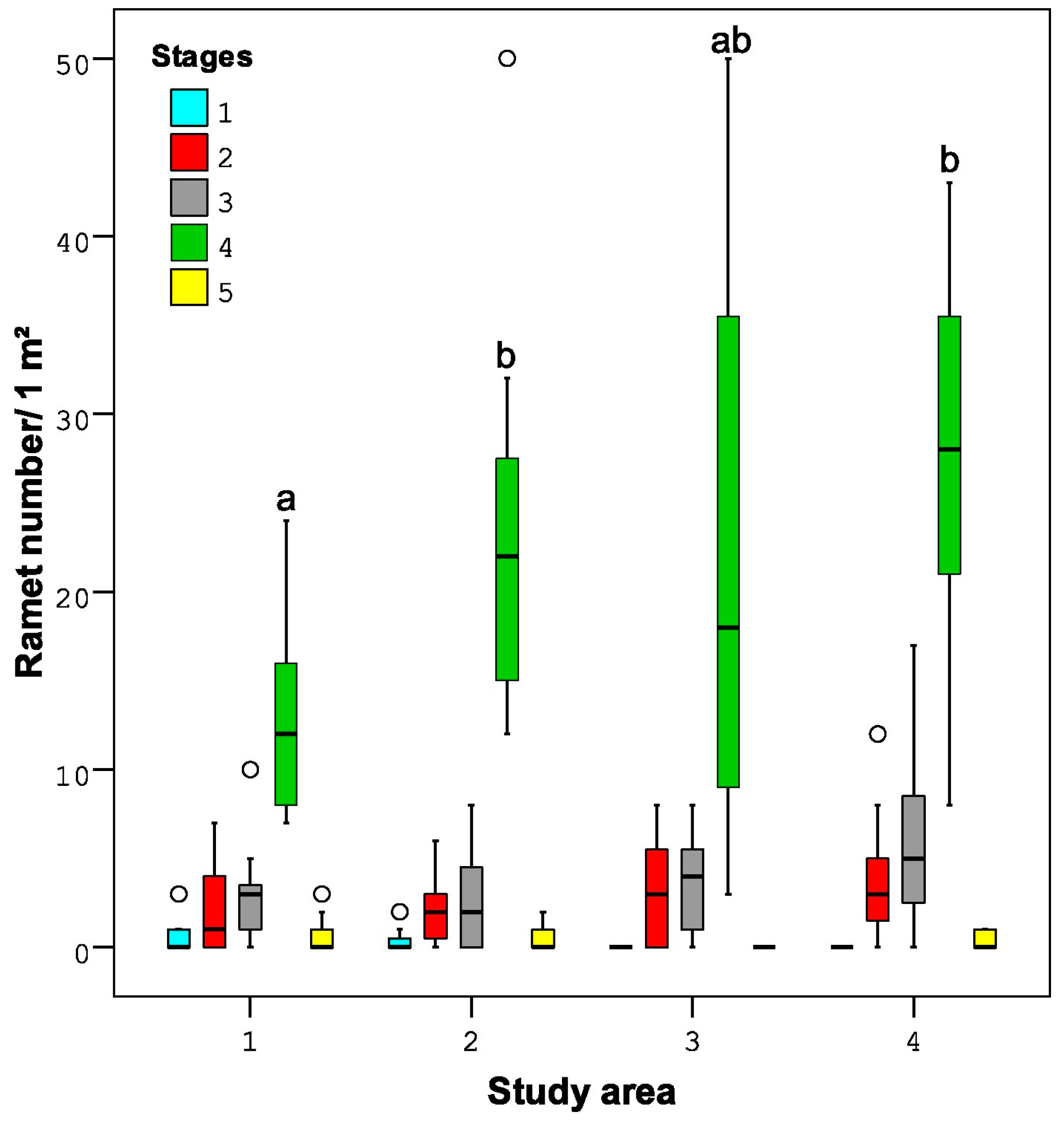
- The total number of ramets (sensu [39]) and the number of ramets in each of the five stages of development. The first four stages followed [15]: stage 1—one-year-old unbranched, stage 2—two-year-old unbranched, stage 3—two-year-old branched, stage 4—branched ramets that were three or more years old. Additionally, we distinguished the 5th stage for the ramets with signs of ageing and degradation (shoots with a grey-coloured main stem and long, thick, and stiff branches and with occasional thin leafy twigs).
- (2)
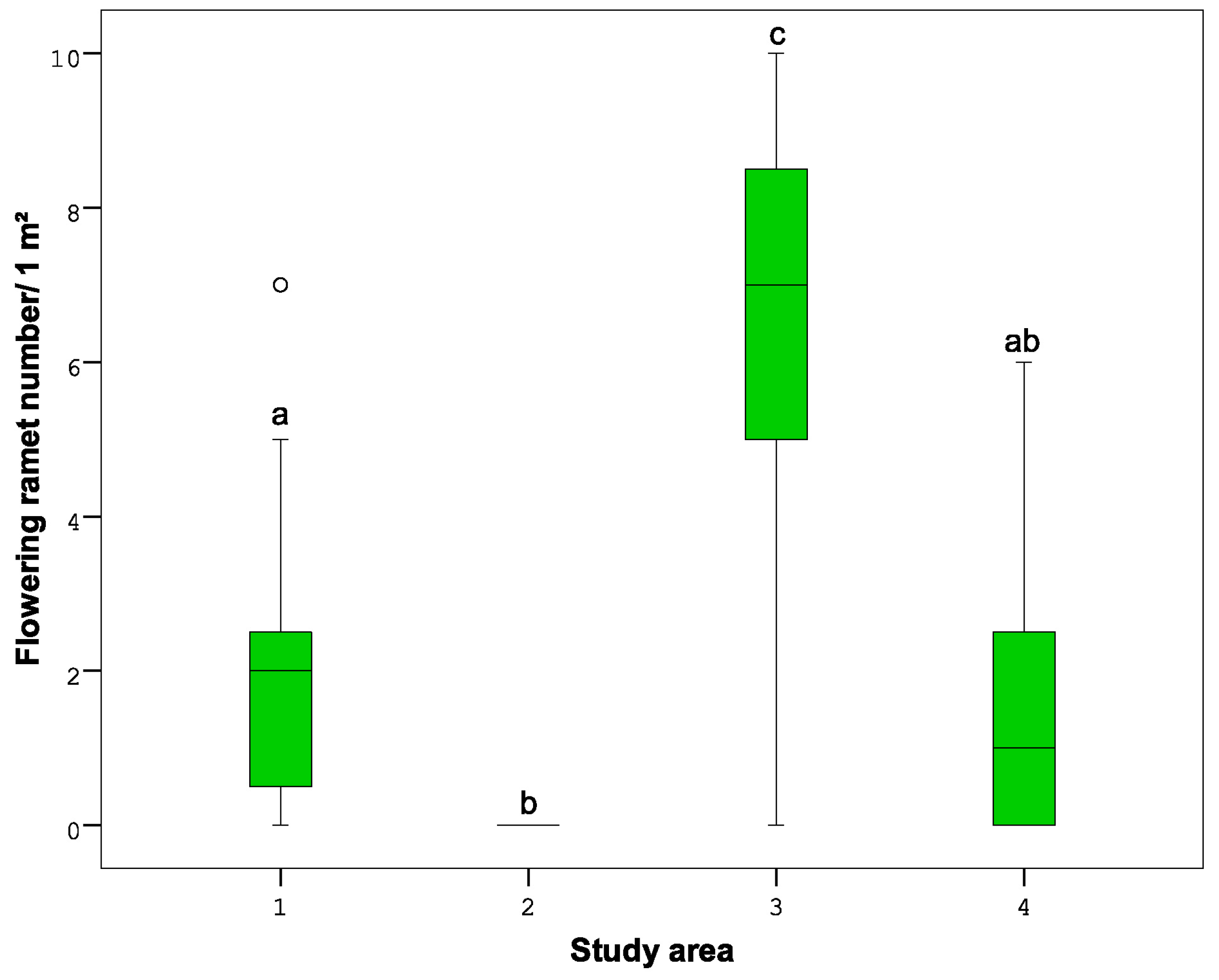
- The number of flowering ramets, followed by the number of male and female catkins on each.
3. Results
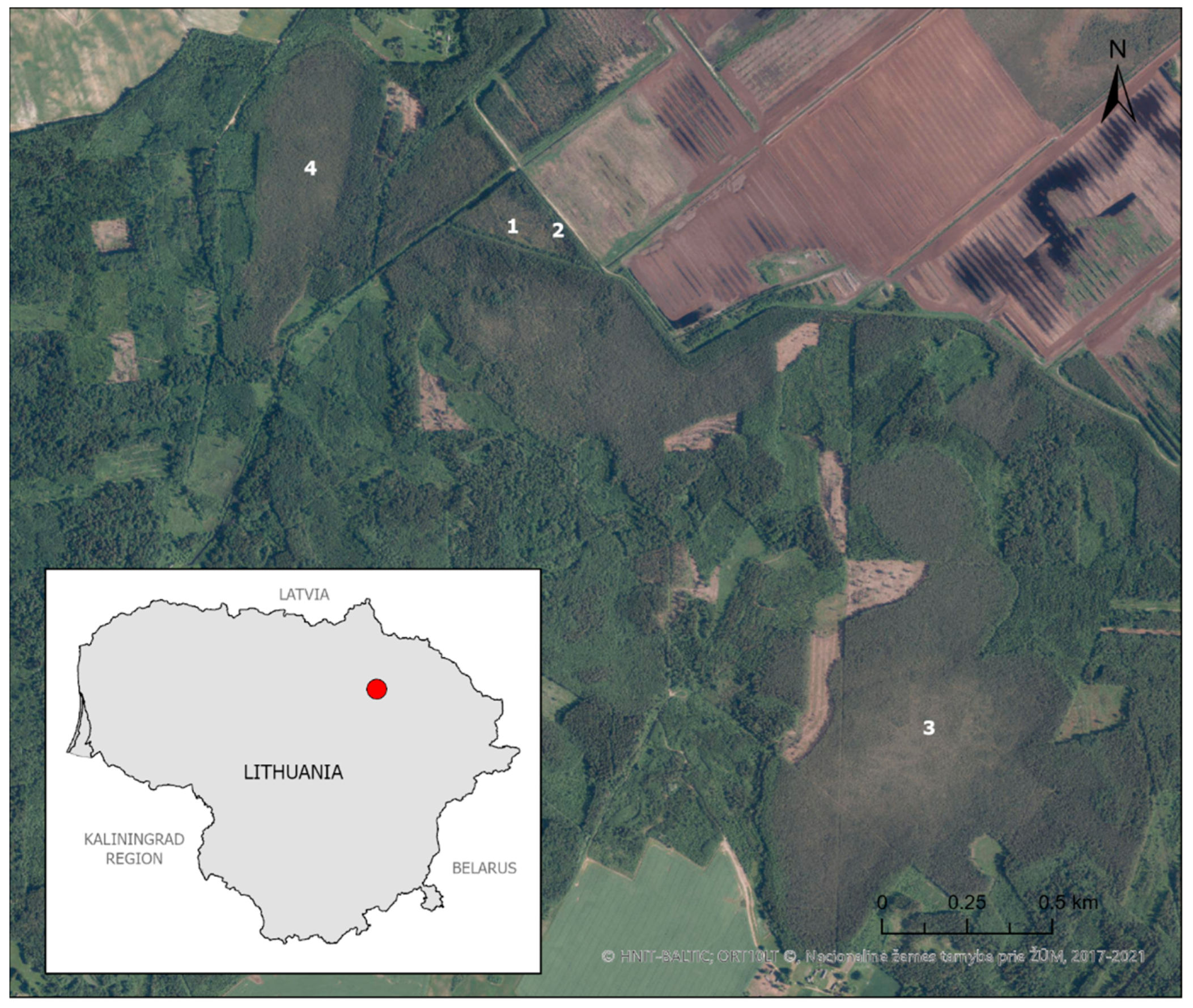
3.1. Characteristics of Betula nana Habitats
3.2. Characteristics Betula nana Population in Šepeta Peatland
3.2.1. Cover, Density, and Flowering of Betula nana
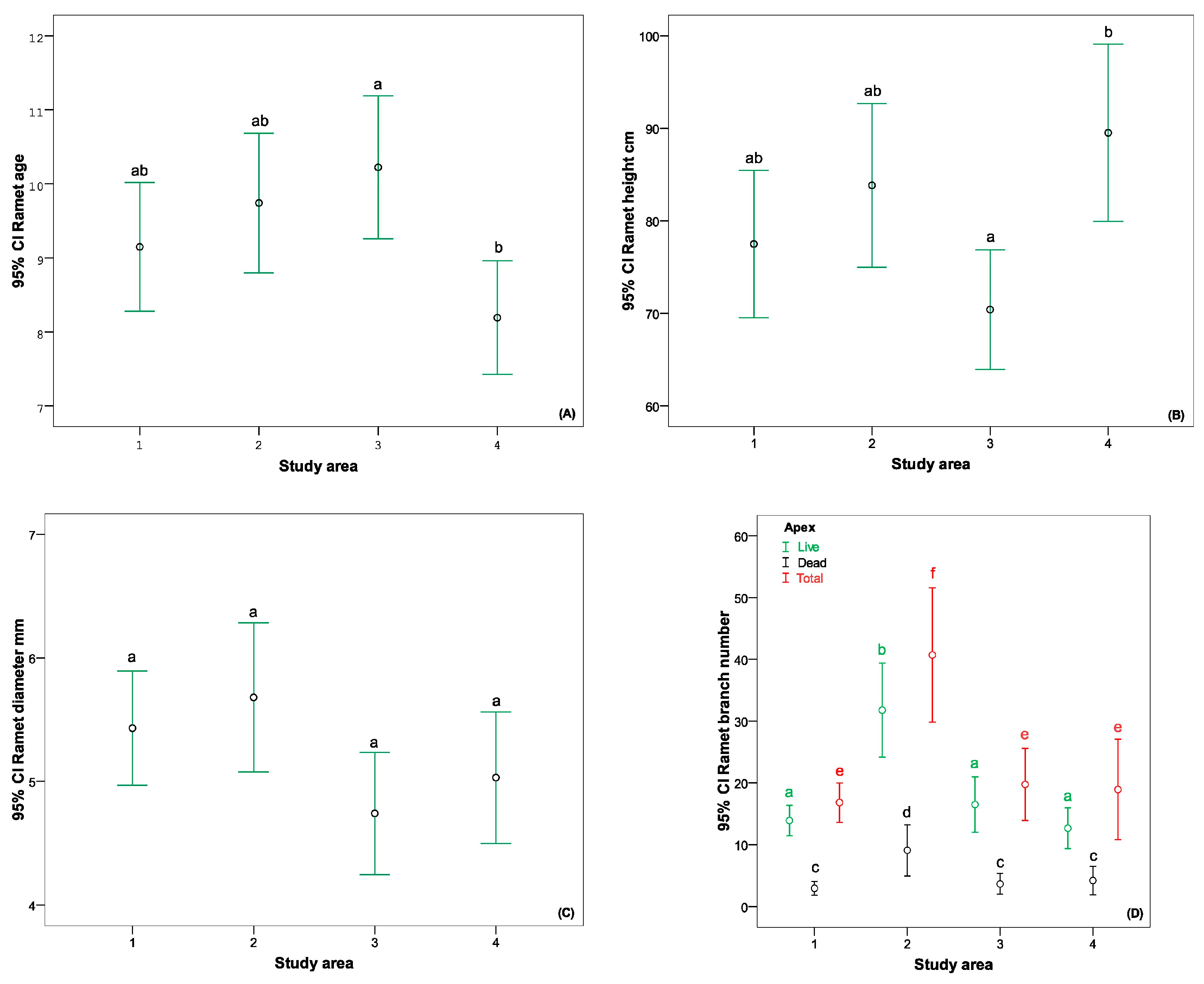
3.2.2. Age and Morphometric Parameters of the Ramets
4. Discussion
4.1. Environmental Differences in Betula nana Cenopopulations
4.2. Cover, Density, and Flowering of Betula nana in Different Cenopopulations
4.3. Age and Morphometric Parameters of the Ramets in Different Cenopopulations
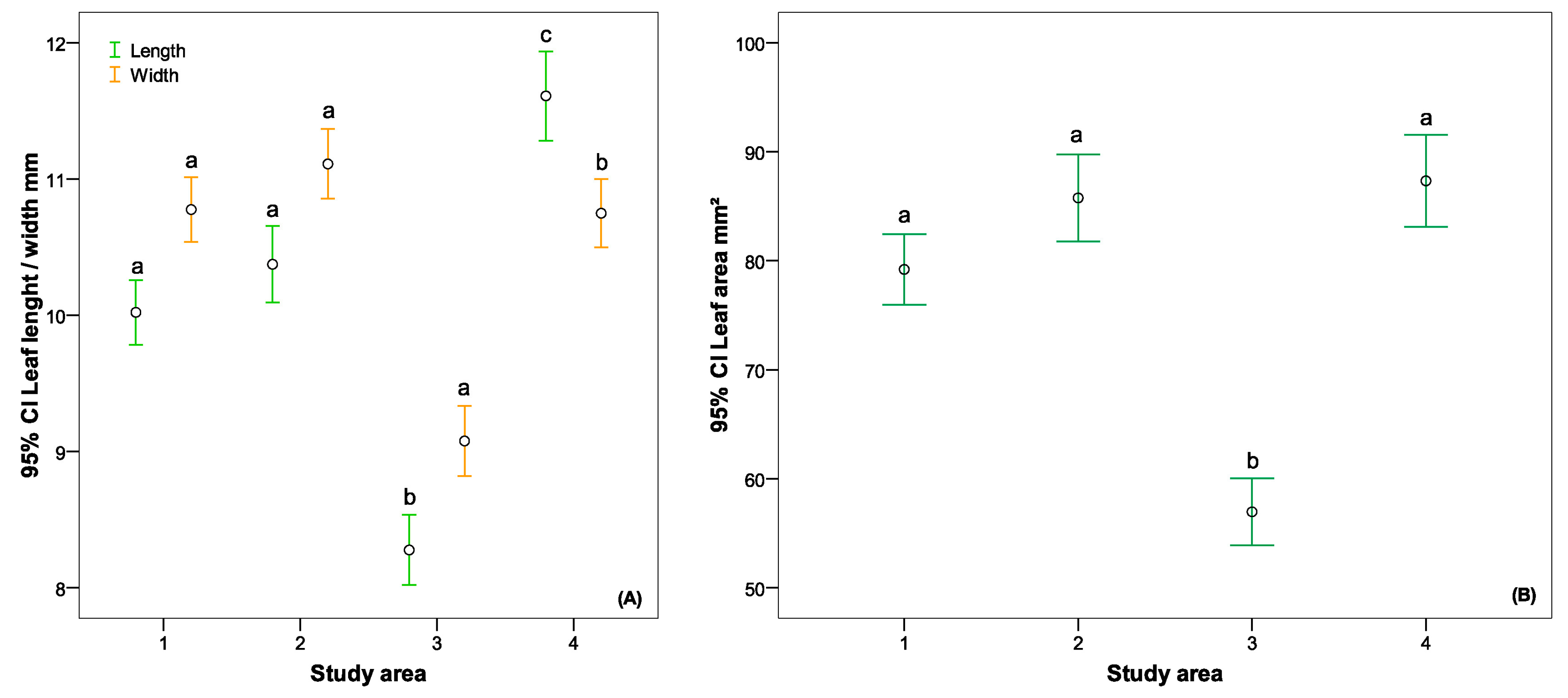
4.4. Variations in Leaf Sizes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Study Area | Minimum | Maximum | Mean | Std. Deviation |
|---|---|---|---|---|---|
| pH 1:5 (H2O) | 1a | 4.0 | 4.0 | 4.000 | 0.000 |
| 2b | 3.9 | 4.0 | 3.967 | 0.0488 | |
| 3b | 3.8 | 4.0 | 3.933 | 0.0976 | |
| 4c | 3.8 | 3.9 | 3.829 | 0.0469 | |
| N (N-NO3+ N-NH4) mg/L | 1a | 0.6 | 1.5 | 1.000 | 0.3873 |
| 2b | 1.2 | 2.5 | 1.800 | 0.5542 | |
| 3a | 0.8 | 1.3 | 1.000 | 0.2236 | |
| 4c | 0.3 | 1.0 | 0.607 | 0.2895 | |
| Ammonia (N-NH4) mg/L | 1a | 0.4 | 0.8 | 0.567 | 0.1759 |
| 2b | 0.9 | 2.1 | 1.367 | 0.5434 | |
| 3a | 0.4 | 1.0 | 0.600 | 0.2928 | |
| 4c | 0.2 | 0.6 | 0.386 | 0.1657 | |
| Nitrate (N-NO3) mg/L | 1a | 0.2 | 0.7 | 0.433 | 0.2127 |
| 2a | 0.3 | 0.6 | 0.433 | 0.1291 | |
| 3a | 0.3 | 0.5 | 0.400 | 0.0845 | |
| 4b | 0.1 | 0.4 | 0.221 | 0.1251 | |
| P mg/L | 1a | 0.3 | 0.4 | 0.367 | 0.488 |
| 2a | 0.3 | 0.4 | 0.367 | 0.488 | |
| 3b | 0.2 | 0.3 | 0.233 | 0.488 | |
| 4b | 0.2 | 0.3 | 0.264 | 0.497 | |
| K mg/L | 1a | 1.0 | 1.7 | 1.300 | 0.3047 |
| 2b | 1.2 | 2.5 | 1.933 | 0.5627 | |
| 3c | 1.5 | 1.9 | 1.633 | 0.1952 | |
| 4d | 0.8 | 1.2 | 0.986 | 0.1657 | |
| Ca mg/L | 1ab | 13 | 19 | 16.00 | 2.535 |
| 2a | 16 | 19 | 17.67 | 1.291 | |
| 3a | 17 | 18 | 17.33 | 0.488 | |
| 4b | 15 | 16 | 15.29 | 0.469 | |
| Mg mg/L | 1a | 6 | 8 | 7.33 | 0.976 |
| 2a | 7 | 9 | 8.00 | 0.845 | |
| 3a | 7 | 8 | 7.67 | 0.488 | |
| 4b | 6 | 7 | 6.29 | 0.469 | |
| Cl mg/L | 1a | 7 | 11 | 9.67 | 1.952 |
| 2b | 7 | 9 | 8.33 | 0.976 | |
| 3b | 7 | 9 | 8.33 | 0.976 | |
| 4c | 7 | 8 | 7.36 | 0.497 | |
| Electrical conductivity mS/m | 1ac | 2.08 | 2.86 | 2.5567 | 0.35318 |
| 2b | 2.58 | 3.14 | 2.9433 | 0.26623 | |
| 3a | 2.44 | 2.75 | 2.5533 | 0.14450 | |
| 4c | 2.39 | 2.58 | 2.4686 | 0.08708 | |
| Organic material % | 1a | 93.77 | 95.66 | 94.7533 | 0.80064 |
| 2b | 93.07 | 95.22 | 94.0700 | 0.91515 | |
| 3c | 92.62 | 94.03 | 93.5067 | 0.65249 | |
| 4b | 93.22 | 94.78 | 94.0857 | 0.69841 | |
| Organic carbon (C) % | 1a | 50.69 | 56.89 | 54.5600 | 2.85215 |
| 2b | 47.99 | 54.24 | 51.9767 | 2.92685 | |
| 3a | 52.76 | 57.84 | 54.9633 | 2.20254 | |
| 4a | 51.26 | 57.78 | 54.0429 | 2.96446 | |
| Peat decomposition % | 1a | 27. 46 | 29.88 | 28.3833 | 1.10539 |
| 2b | 29.50 | 32.04 | 31.1800 | 1.22975 | |
| 3c | 29.04 | 31.21 | 30.1733 | 0.91972 | |
| 4b | 30.61 | 32.43 | 31.3000 | 0.87596 | |
| Moisture % | 1a | 89.14 | 90.29 | 89.8067 | 0.50415 |
| 2b | 87.77 | 88.48 | 88.0933 | 0.30359 | |
| 3ac | 88.25 | 90.68 | 89.6233 | 1.05269 | |
| 4c | 88.28 | 89.86 | 89.2136 | 0.72834 | |
| Water level depth m | 1a | 37 | 45 | 42.33 | 3.904 |
| 2b | >1 | >1 | >1 | 0.000 | |
| 3a | 24 | 54 | 38.33 | 12.715 | |
| 4a | 44 | 49 | 45.43 | 2.344 | |
| Herb cover % | 1a | 0 | 60 | 15.00 | 16.583 |
| 2b | 0 | 40 | 4.40 | 10.253 | |
| 3a | 3 | 60 | 21.40 | 16.612 | |
| 4a | 0 | 35 | 10.36 | 12.475 | |
| Tree cover % | 1a | 20 | 60 | 43.33 | 17.593 |
| 2a | 30 | 60 | 43.33 | 12.910 | |
| 3b | 5 | 30 | 18.33 | 10.635 | |
| 4a | 0 | 40 | 37.33 | 10.328 | |
| Shrub and subshrub cover % (Betula nana excluded) | 1a | 0.00 | 70.00 | 30.3571 | 27.62832 |
| 2a | 2.00 | 40.00 | 20.8000 | 10.73845 | |
| 3b | 3.00 | 80.00 | 51.8667 | 23.03992 | |
| 4ac | 0.00 | 40.00 | 19.0000 | 11.5824 | |
| Moss cover % | 1a | 70 | 80 | 76.67 | 4.880 |
| 2b | 40 | 80 | 61.67 | 17.078 | |
| 3c | 80 | 90 | 83.33 | 4.880 | |
| 4ac | 70 | 85 | 78.21 | 6.682 |
| Study Area | 1 | 2 | 3 | 4 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number of study plot | 1.1 | 1.2 | 1.3 | 2.1 | 2.2 | 2.3 | 3.1 | 3.2 | 3.3 | 4.1 | 4.2 | 4.3 |
| Cover of tree layer [%]: | 50 | 20 | 60 | 40 | 60 | 30 | 20 | 5 | 30 | 40 | 40 | 40 |
| Cover of shrub and subshrub layers [%] | 80 | 60 | 80 | 85 | 60 | 60 | 80 | 80 | 80 | 80 | 40 | 70 |
| Cover of herb layer [%] | 2 | 50 | 5 | 8 | 5 | 2 | 1 | 50 | 60 | 5 | 10 | 10 |
| Cover of moss layer [%] | 80 | 80 | 70 | 40 | 65 | 80 | 80 | 95 | 80 | 80 | 70 | 85 |
| Trees | ||||||||||||
| Pinus sylvestris | 3 | 2 | 3 | 3 | 3 | 2 | 2 | 1 | 2 | 3 | 2 | 3 |
| Betula pubescens | + | 2 | + | |||||||||
| Betula pendula | 1 | 1 | ||||||||||
| B. pubescens × pendula | + | + | 1 | |||||||||
| Shrubs and subshrubs | ||||||||||||
| Betula nana | 2 | 3 | 3 | 4 | 4 | 3 | 3 | 3 | 3 | 3 | 3 | 4 |
| Andromeda polifolia | 2 | 1 | 2 | 1 | 1 | 2 | 1 | 2 | 1 | 1 | 2 | |
| Vaccinium oxycoccos | 2 | 3 | 1 | 3 | 2 | 2 | 2 | 2 | 1 | 1 | 2 | |
| Empetrum nigrum | 3 | 2 | 2 | 1 | 1 | 2 | 2 | 2 | 2 | 2 | ||
| Calluna vulgaris | 3 | 3 | 2 | 2 | 1 | 2 | 3 | 3 | 3 | |||
| Rhododendron tomentosum | 2 | 1 | 1 | 3 | 3 | 1 | 3 | 1 | ||||
| Vaccinium uliginosum | + | |||||||||||
| Herbs | ||||||||||||
| Eriophorum vaginatum | 3 | 1 | 1 | 1 | 3 | 4 | 1 | 2 | 2 | |||
| Melampyrum pratense | 1 | 1 | + | 1 | 1 | |||||||
| Betula pubescens juv. | + | 1 | 1 | |||||||||
| Pinus sylvestris juv. | + | + | + | |||||||||
| Rubus chamaemorus | + | + | ||||||||||
| Carex rostrata | + | + | ||||||||||
| Carex sp. | + | |||||||||||
| Phragmites australis | + | |||||||||||
| Mosses | ||||||||||||
| Sphagnum divinum | 2 | 1 | 2 | 1 | 2 | 3 | 3 | 2 | 3 | 2 | 3 | 3 |
| Polytrichum strictum | 2 | 2 | 2 | 1 | 2 | 2 | 1 | 1 | + | 1 | 2 | |
| Pleurozium schreberi | 2 | 2 | 3 | 2 | 1 | 2 | 3 | 2 | 3 | 1 | 3 | |
| Sphagnum cuspidatum | + | 3 | 2 | 1 | 1 | 1 | 1 | 1 | ||||
| Sphagnum fallax | 3 | 1 | + | 1 | 1 | 1 | 3 | 1 | ||||
| Dicranum polysetum | 2 | 1 | 2 | 1 | + | 1 | ||||||
| Sphagnum angustifolium | + | 1 | 1 | 1 | 1 | 1 | ||||||
| Sphagnum flexuosum | 1 | 1 | 1 | 1 | 1 | 1 | ||||||
| Sphagnum fuscum | 1 | 1 | 1 | |||||||||
| Pohlia sphagnicola | + | |||||||||||
| Calliergon stramineum | + | |||||||||||
| Sphagnum capillifolium | 1 | |||||||||||
| Sphagnum balticum | + | |||||||||||
| Sphagnum rubellum | + |
References
- Jalas, J.; Suominen, J. Atlas Florae Europaeae; Salicaceae to Balanophoracea; Committee for Mapping the Flora of Europe and Societas Biologica Fennica Vanamo: Helsinki, Finland, 1976; Volume 3, p. 57. [Google Scholar]
- de Groot, W.J.; Thomas, P.A.; Wein, R.W. Betula nana L. and Betula glandulosa Michx. J. Ecol 1997, 85, 241–264. [Google Scholar] [CrossRef]
- Drzymulska, D. Postglacial occurrence and decline of Betula nana L. (dwarf birch) in northeastern Poland. Est. J. Earth Sci. 2014, 63, 76–87. [Google Scholar] [CrossRef]
- Jadwiszczak, K.A.; Kłosowski, S.; Zalewska, I.; Banaszek, A.; Chrzanowska, A. Genetic diversity and sexual reproduction in relict populations of Betula nana. Silva Fenn. 2017, 51, id5643. [Google Scholar] [CrossRef]
- Gostyńska-Jakuszewska, M.; Lekavičius, A. Selected Boreal and Subboreal species in the flora of Poland and the Lithuanian SSR. Part I. Fragm. Flor. Geobot. 1989, 34, 299–314. [Google Scholar]
- Fredskild, B. The genus Betula in Greenland-Holocene history, present distribution and synecology. Nord. J. Bot. 2008, 11, 393–412. [Google Scholar] [CrossRef]
- Anamthawat-Jónsson, K.; Thór Thórsson, A. Natural hybridisation in birch: Triploid hybrids between Betula nana and B. pubescens. Plant Cell Tissue Organ Cult. 2003, 75, 99–107. [Google Scholar] [CrossRef]
- Anamthawat-Jónsson, K.; Karlsdóttir, L.; Thórsson, T.; Jóhannsson, M.H. Naturally occurring triploid birch hybrids from woodlands in Iceland are partially fertile. New For. 2021, 52, 659–678. [Google Scholar] [CrossRef]
- Alsos, I.G.; Engelskjøn, T. Conservation genetics and population history of Betula nana, Vaccinium uliginosum, and Campanula rotundifolia in the Arctic Archipelago of Svalbard. Arct. Antarct. Alp. Res. 2002, 34, 408–418. [Google Scholar] [CrossRef]
- Alsos, I.G.; Spjelkavik, S.; Engelskjøn, T. Seed bank size and composition of Betula nana, Vaccinium ulginosum, and Campanula rotundifolia habitats in Svalbard and Northern Norway. Can. J. Bot. 2003, 81, 220–231. [Google Scholar] [CrossRef]
- Hollesen, J.; Buchwal, A.; Rachlewicz, G.; Hansen, B.; Hansen, M.O.; Stecher, O.; Elberling, B. Winter warming as an important co-driver for Betula nana growth in western Greenland during the past century. Global Chang. Biol. 2015, 21, 2410–2423. [Google Scholar] [CrossRef]
- Dąbrowska, G.; Dzialuk, A.; Burnicka, O.; Ejankowski, W.; Gugnacka-Fiedor, W.; Goc, A. Genetic diversity of postglacial relict shrub Betula nana revealed by RAPD analysis. Dendrobiology 2006, 55, 19–23. [Google Scholar]
- Dąbrowska, G.B.; Henryk, P.; Dąbrowski, H.B.; Szyp-Borowska, I. Genetic diversity of Betula nana in Sweden and conservation implications for protection of relict Polish populations. Folia For. Pol. Ser. A-For. 2021, 63, 225–231. [Google Scholar] [CrossRef]
- Ejankowski, W. The influence of ground water level on the demography and population structure of the dwarf birch Betula nana L. Ecol. Quest. 2004, 6, 63–68. [Google Scholar]
- Ejankowski, W. Effect of waterlogging on regeneration in the dwarf birch (Betula nana). Biologia 2008, 3, 670–676. [Google Scholar] [CrossRef]
- Ejankowski, W. Demographic variation of dwarf birch (Betula nana) in communities dominated by Ledum palustre and Vaccinium uliginosum. Biologia 2010, 65, 248–253. [Google Scholar] [CrossRef]
- Ejankowski, W.; Kunz, M. Reconstructing of vegetation dynamics in “Linje” Peat-bog (N Poland) using remote sensing method. Biodiv. Res. Conserv. 2006, 1–2, 111–113. [Google Scholar]
- Jadwiszczak, K.A.; Drzymulska, D.; Banaszek, A.; Jadwiszczak, P. Population history, genetic variation and conservation status of the endangered birch species Betula nana L. in Poland. Silva Fenn. 2012, 46, 465–477. [Google Scholar] [CrossRef]
- Cinovskis, R. Betula nana L. In Latvijas Sarkanā grāmata. Vaskulārie aug; Andrušaitis, G., Ed.; Latvijas Universitātes Bioloģijas institūts: Riga, Latvia, 2003; Volume 3, p. 298. [Google Scholar]
- Patalauskaitė, D. Beržas keružis. Betula nana L. In Red Data Book of Lithuania. Animals, Plants, Fungi; Rašomavičius, V., Ed.; Aplinkos Ministerija: Vilnius, Lithuania, 2021; p. 466. [Google Scholar]
- Kruszelnicki, J.; Fabiszewski, J. Betula nana L. Brzoza karłowata [Betula nana L. Dwarf birch]. In Polska Czerwona Księga Roślin [Polish Red Data Book of Plants]; Kaźmierczakowa, R., Zarzycki, K., Eds.; Instytut Botaniki im. W. Szafera PAN, Instytut Ochrony Przyrody PAN: Krakow, Poland, 2001; pp. 82–83. [Google Scholar]
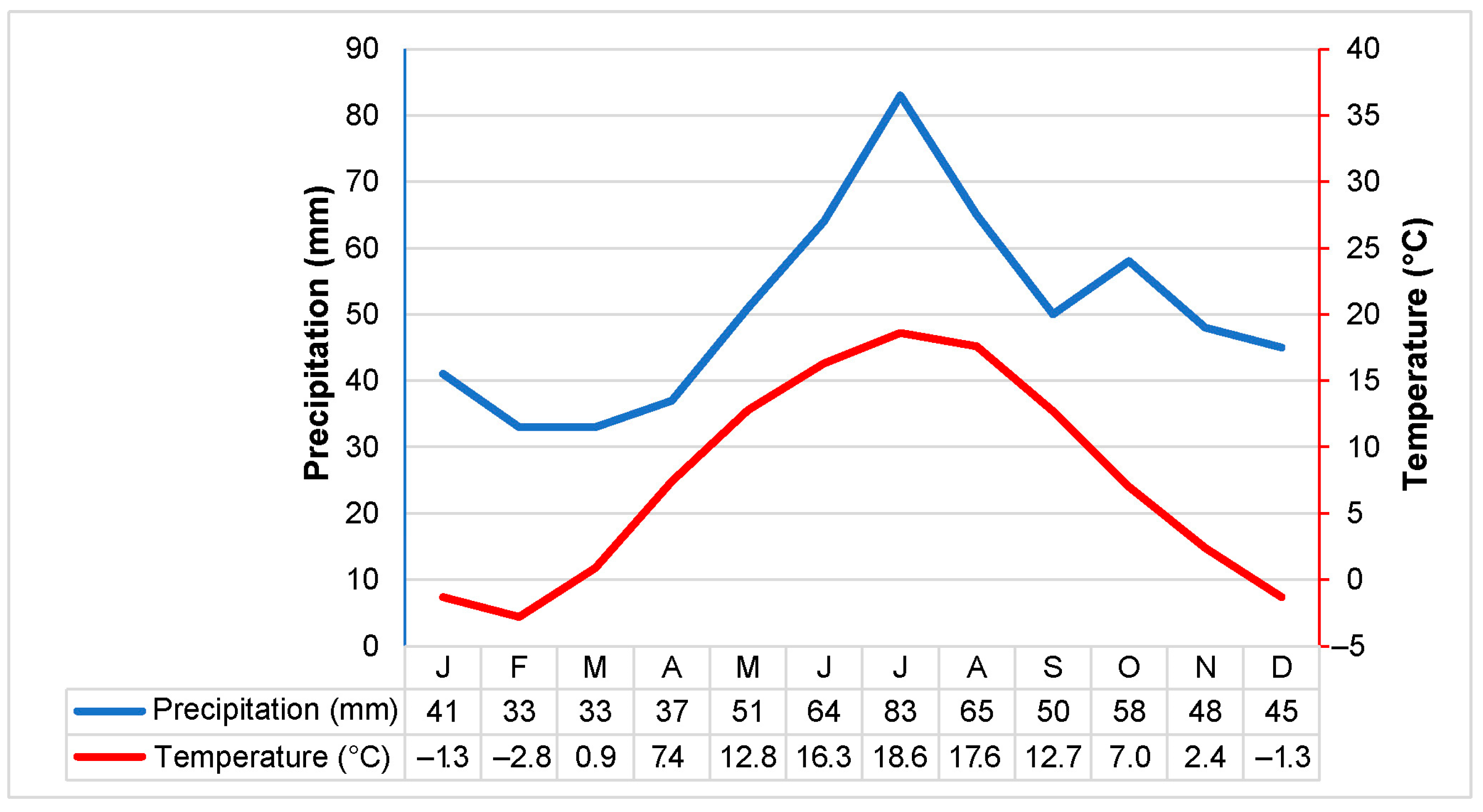
- Bukantis, A.; Gulbinas, Z.; Kazakevičius, S.; Kilkus, K.; Mikelinskienė, A.; Morkūnaitė, R.; Rimkus, E.; Samuila, M.; Stankūnavičius, G.; Valiuškevičius, G.; et al. Klimato Svyravimų Poveikis Fiziniams Geografiniams Procesams Lietuvoje; Geografijos Institutas, Vilniaus Universitetas: Vilnius, Lithuania, 2001; pp. 1–280. [Google Scholar]
- Povilaitis, A.; Taminskas, J.; Gulbinas, Z.; Linkevičienė, R.; Pileckas, M. Lietuvos Šlapynės ir jų Vandensauginė Reikšmė; Apyaušris: Vilnius, Lithuania, 2011; p. 52. [Google Scholar]
- LHT. Lietuvos Hidrometeorologijos Tarnyba. Available online: https://www.meteo.lt/en/ (accessed on 1 August 2024).
- Liužinas, R. Lietuvos Durpynų Kadastras I, 1st ed.; Lietuvos Respublikos Aplinkos Apsaugos Ministerija: Vilnius, Lithuania, 1995; p. 302. [Google Scholar]
- Brundza, K. Šepeta. Aukštapelkio monografija. Žemės Akad. Metraštis 1940, 13, 1–208. [Google Scholar]
- Ivanauskas, T. Gamtos paminklai ir jų klausimas Lietuvoje. Švietimo Darbas 1921, 1–2, 13–33. [Google Scholar]
- Rabotnov, T.A. Dynamics of Plant Coenotic Populations. In The Population Structure of Vegetation. Handbook of Vegetation Science; White, J., Ed.; Springer: Dordrecht, The Netherlands, 1985; Volume 3, pp. 121–142. [Google Scholar] [CrossRef]
- Jonsell, B. Betula L. In Flora Nordica; Lycopodiaceae–Polygonaceae; Jonsel, B., Karlssson, T., Eds.; Flora Nordica: Stockholm, Sweden, 2000; Volume 1, pp. 197–203. [Google Scholar]
- Falińska, K. Osobnik, Populacja, Fitocenoza; PWN: Warszawa, Poland, 1990; pp. 1–309. [Google Scholar]
- Scrosati, R. An updated definition of genet applicable to clonal seaweeds, bryophytes, and vascular plants. Basic Appl. Ecol. 2002, 3, 97–99. [Google Scholar] [CrossRef]
- Tsujimoto, M.; Araki, K.S.; Honjo, M.N.; Yasugi, M.; Nagano, A.J.; Akama, S.; Hatakeyama, M.; Shimizu-Inatsugi, R.; Sese, J.; Shimizu, K.K.; et al. Genet assignment and population structure analysis in a clonal forest-floor herb, Cardamine leucantha, using RAD-seq. AoB Plants 2019, 11, plz080. [Google Scholar] [CrossRef] [PubMed]
- Braun-Blanquet, J. Pflanzensoziologie, Grundzüge der Vegetationskunde, 3rd ed.; Springer: Wien, Austria; New York, NY, USA, 1964; p. 866. [Google Scholar]
- LST EN 13037; Dirvožemio Gerinimo Medžiagos ir Auginimo Terpės. pH Nustatymas. Lietuvos Standartizacijos Departamentas: Vilnius, Lithuania, 2012; pp. 1–8.
- LST EN 13652:2006; Dirvožemio Gerinimo Medžiagos ir Auginimo Terpės. Vandenyje Tirpių Maisto Medžiagų ir Elementų Eekstrahavimas. Lietuvos Standartizacijos Departamentas: Vilnius, Lithuania, 2006; pp. 1–15.
- LST 1957:2022; Durpės, Skirtos Daržininkystei, Sodininkystei ir Dekoratyvinei Sodininkystei. Savybės, Tyrimo Metodai, Techninės Tiekimo Sąlygos. Lietuvos Standartizacijos Departamentas: Vilnius, Lithuania, 2022; pp. 1–26.
- LST EN 13040:2008; Dirvožemio Gerinimo Medžiagos ir Auginimo Terpės. Mėginių Paruošimas Cheminiams ir Fizikiniams Tyrimams, Sausųjų Medžiagų Kiekio, Drėgnio ir Laboratorijoje Tankinto Piltinio Tankio Nustatymas. Lietuvos Standartizacijos departamentas: Vilnius, Lithuania, 2009; pp. 1–17.
- ISO 10694:1995; Soil Quality Determination of Organic and Total Carbon after Dry Combustion (Elementary Analysis). International Organization for Standardization: Geneva, Switzerland, 1995; pp. 1–18.
- Harper, J.L. Population Biology of Plants; Academic Press: London, UK, 1977; pp. 1–892. [Google Scholar]
- Gärtner, H.; Schweingruber, F.H. Microscopic Preparation Techniques for Plant Stem Analysis; Verlag Dr. Kessel: Remagen, Germany, 2013; pp. 19–29. [Google Scholar]
- SPSS Inc. SPSS for Windows, Version 16.0; SPSS Inc.: Chicago, IL, USA, 2007; pp. 1–165. [Google Scholar]
- McGarigal, K.; Stafford, S.; Cushman, S. Ordination: Principal components analysis. In Multivariate Statistics for Wildlife and Ecology Research; McGarigal, K., Stafford, S., Cushman, S., Eds.; Springer: New York, NY, USA, 2000; pp. 19–80. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Szajdak, L.W.; Jezierski, A.; Wegner, K.; Meysner, T.; Szczepański, M. Influence of Drainage on Peat Organic Matter: Implications for Development, Stability, and Transformation. Molecules 2020, 25, 2587. [Google Scholar] [CrossRef]
- Koeselman, W.; Verhoeven, J.T.A. Eutrophication of fen ecosystems: External and internal nutrient sources and restoration strategies. In Restoration of Temperate Wetlands; Wheeler, B.D., Shaw, S.C., Fojt, W.J., Robertson, R.A., Eds.; John Wiley & Sons: Chichester, UK, 1995; pp. 91–112. [Google Scholar]
- Laine, J.; Vanha-Majamaa, I. Vegetation ecology along a trophic gradient on drained pine mires in southern Finland. Ann. Bot. Fenn 1992, 29, 213–233. [Google Scholar]
- Laine, J.; Vasander, H.; Laiho, R. Long-term effects of water level drawdown on the vegetation of drained pine mires in southern Finland. J. Appl. Ecol 1995, 32, 785–802. [Google Scholar] [CrossRef]
- Harris, L.I.; Moore, T.R.; Roulet, N.T.; Pinsonneault, A.J. Limited effect of drainage on peat properties, porewater chemistry, and peat decomposition proxies in a boreal peatland. Biogeochemistry 2020, 151, 43–62. [Google Scholar] [CrossRef]
- Edvardsson, J.; Šimanauskienė, R.; Taminskas, J.; Baužienė, I.; Stoffel, M. Increased tree establishment in Lithuanian peat bogs detected using a combination of field and remotely sensed approaches. Sci. Total Environ. 2015, 505, 113–120. [Google Scholar] [CrossRef]
- Dudová, L.; Hájek, M.; Hájková, P. The origin and vegetation development of the Rejvíz pine bog and the history of the surrounding landscape during the holocene. Preslia 2010, 82, 223–246. [Google Scholar]
- Felèchoux, F.; Buttler, A.; Gillet, F. Dynamics of bog-pine-dominated mires in the Jura Mountains, Switzerland: A tentative scheme based on synusial phytosociology. Folia Geobot 2000, 35, 273–288. [Google Scholar] [CrossRef]
- Paal, J.; Jürjendal, I.; Suija, A.; Kull, A. Impact of drainage on vegetation of transitional mires in Estonia. Mires Peat 2016, 18, 1–19. [Google Scholar]
- Ebert, T.A.; Ebert, C.A. A method for studying vegetation dynamics when there are no obvious individuals: Virtual-population analysis applied to the tundra shrub Betula nana L. Vegetatio 1989, 85, 33–44. [Google Scholar] [CrossRef]
- Nowakowska, J.; Gazda, A.; Tomski, A.; Szwagrzyk, J. Drainage ditches enhance forest succession in a raised bog but do not affect the spatial pattern of tree encroachment. PLoS ONE 2021, 16, e0247760. [Google Scholar] [CrossRef]
- Myers-Smith, I.H.; Hallinger, M.; Blok, D.; Sass-Klaassen, U.; Rayback, S.A.; Weijers, S.; Trant, A.J.; Tape, K.D.; Naito, A.T.; Wipf, S.; et al. Methods for measuring arctic and alpine shrub growth: A review. Earth-Sci. Rev. 2015, 140, 1–13. [Google Scholar] [CrossRef]
- Breshears, D.D.; Rich, P.M.; Barnes, F.J.; Campbell, K. Overstory-imposed heterogeneity in solar radiation and soil moisture in a semiarid woodland. Ecol. Appl. 1997, 7, 1201–1215. [Google Scholar] [CrossRef]
- Breshears, D.D.; Nyhan, J.W.; Heil, C.E.; Wilcox, B.P. Effects of woody plants on microclimate in a semiarid woodland: Soil temperature and evaporation in canopy and intercanopy patches. Int. J. Plant Sci. 1998, 159, 1010–1017. [Google Scholar] [CrossRef]
- Wen, B.; Zhang, Y.; Hussain, S.; Wang, S.; Zhang, X.; Yang, J.; Xu, M.; Qin, S.; Yang, W.; Liu, W. Slight Shading Stress at Seedling Stage Does not Reduce Lignin Biosynthesis or Affect Lodging Resistance of Soybean Stems. Agronomy 2020, 10, 544. [Google Scholar] [CrossRef]
- Franklin, K.A. Shade avoidance. New Phytol. 2008, 179, 930–944. [Google Scholar] [CrossRef]
- Casal, J.J. Shade avoidance. Arab. Book Am. Soc. Plant Biol. 2012, 10, e0157. [Google Scholar] [CrossRef]
- Casal, J.J. Photoreceptor signaling networks in plant responses to shade. Annu. Rev. Plant Biol 2013, 64, 403–427. [Google Scholar] [CrossRef]
- Hutchinson, T.C. Comparative studies of the ability of species to withstand prolonged periods of darkness. J. Ecol. 1967, 55, 291–299. [Google Scholar] [CrossRef]
- Schmitt, J.; McCormac, A.C.; Smith, H. A test of the adaptive plasticity hypothesis using transgenic and mutant plants disabled inphytochrome-mediated elongation responses to neighbors. Am. Nat. 1995, 146, 937–953. [Google Scholar] [CrossRef]
- Wein, R.W.; Bliss, L.C. Primary production in arctic cottongrass tussock tundra communities. Arct. Alp. Res. 1974, 6, 261–274. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Kempf, J.S. Branching patterns in forest shrubs and understorey trees in relation to habitat. New Phytol. 1980, 86, 219–228. [Google Scholar] [CrossRef]
- Bret-Harte, M.S.; Shaver, G.R.; Zoerner, J.P.; Johnstone, J.F.; Wagner, J.L.; Chavez, A.S.; Gunkelman, R.F.; Lippert, S.C.; Laundre, J.A. Developmental plasticity allows Betula nana to dominate tundra subjected to an alternative environment. Ecology 2001, 82, 18–32. [Google Scholar] [CrossRef]
- Gommers, C.M.; Visser, E.J.; St Onge, K.R.; Voesenek, L.A.; Pierik, R. Shade tolerance: When growing tall is not an option. Trends Plant Sci. 2013, 18, 65–71. [Google Scholar] [CrossRef]
- Furlow, J.J. Betulaceae Gray. Betula L. In Flora of North America North of Mexico; Flora of North America Editorial Committee: New York, NY, USA; Oxford, UK, 1997; Volume 3, Available online: http://www.efloras.org/florataxon.aspx?flora_id=1&taxon_id=10101 (accessed on 27 May 2024).
- Lekavičius, A. Vadovas Augalams Pažinti; Mokslas: Vilnius, Lithuania, 1989; pp. 54–56. [Google Scholar]
- Gostyńska-Jakuszewska, M. Betulaceae, Brzozowate. In Flora Polski. Rośliny Naczyniowe, 3. Dwuliścienne Wolnopłatkowe-Jednookwiatowe, 2nd ed.; Jasiewicz, W.A., Ed.; Instytut Botaniki im. W. Szafera, Polska Akademia Nauk: Kraków, Poland, 1992; pp. 7–18. [Google Scholar]
- Hodgetts, N.G.; Söderström, L.; Blockeel, T.L.; Caspari, S.; Ignatov, M.S.; Konstantinova, N.A.; Lockhart, N.; Papp, B.; Schröck, C.; Sim-Sim, M.; et al. An annotated checklist of bryophytes of Europe, Macaronesia and Cyprus. J. Bryol. 2020, 42, 1–116. [Google Scholar] [CrossRef]
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/ (accessed on 8 May 2024).


| Environmental Variables | PC1 | PC2 |
|---|---|---|
| NH4+ | −0.128 | 0.574 |
| Ca | −0.030 | 0.062 |
| Cl | −0.035 | −0.023 |
| K | −0.113 | 0.202 |
| Mg | −0.035 | 0.089 |
| N | −0.152 | 0.512 |
| NO3− | −0.155 | 0.363 |
| P | 0.072 | 0.128 |
| pH | −0.003 | 0.006 |
| Organic carbon | −0.019 | −0.010 |
| Organic material | 0.001 | 0.001 |
| Peat decomposition degree | 0.018 | 0.006 |
| Peat moisture | −0.003 | −0.002 |
| Electric conductivity | 0.008 | 0.083 |
| Water level | 0.096 | 0.338 |
| Herb cover | −0.102 | 0.049 |
| Moss cover | −0.051 | −0.117 |
| Shrub cover | −0.361 | −0.226 |
| Tree cover | 0.505 | 0.064 |
| Tree cover in the surrounding | 0.710 | 0.108 |
| Eigenvalue | 1.24 | 0.97 |
| % variance | 43.42 | 33.84 |
| Cumulative explanation % | 43.42 | 77.26 |
| Study Area | Minimum | Maximum | Mean ± Std. Deviation |
|---|---|---|---|
| ♀ 1a | 1.0 | 9.0 | 2.5217 ± 2.44707 |
| ♀ 2ac | 0.0 | 2.0 | 1.0000 ± 1.41421 |
| ♀ 3b | 0.0 | 63.0 | 6.3656 ± 10.55399 |
| ♀ 4c | 0.0 | 1.0 | 0.0526 ± 0.22942 |
| ♂ 1a | 1.0 | 10.0 | 3.2609 ± 2.63227 |
| ♂ 2a | 1.0 | 1.0 | 1.0000 ± 1.41421 |
| ♂ 3b | 0.0 | 37.0 | 5.2258 ± 7.18563 |
| ♂ 4a | 0.0 | 4.0 | 1.4583 ± 0.93153 |
| Ramet Age | Ramet Height | Ramet Diameter | |
|---|---|---|---|
| Ramet height Ramet diameter Number of twigs | 0.339 | ||
| 0.658 | 0.596 | ||
| 0.196 | 0.253 | 0.430 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jukonienė, I.; Bagušinskaitė, A.; Kalvaitienė, M.; Ričkienė, A.; Sinkevičienė, Z. The Population of the Glacial Relict Betula nana Surviving Anthropogenic Pressure (the Case of Šepeta Peatland in Northeastern Lithuania). Diversity 2024, 16, 512. https://doi.org/10.3390/d16090512
Jukonienė I, Bagušinskaitė A, Kalvaitienė M, Ričkienė A, Sinkevičienė Z. The Population of the Glacial Relict Betula nana Surviving Anthropogenic Pressure (the Case of Šepeta Peatland in Northeastern Lithuania). Diversity. 2024; 16(9):512. https://doi.org/10.3390/d16090512
Chicago/Turabian StyleJukonienė, Ilona, Agnė Bagušinskaitė, Monika Kalvaitienė, Aurika Ričkienė, and Zofija Sinkevičienė. 2024. "The Population of the Glacial Relict Betula nana Surviving Anthropogenic Pressure (the Case of Šepeta Peatland in Northeastern Lithuania)" Diversity 16, no. 9: 512. https://doi.org/10.3390/d16090512
APA StyleJukonienė, I., Bagušinskaitė, A., Kalvaitienė, M., Ričkienė, A., & Sinkevičienė, Z. (2024). The Population of the Glacial Relict Betula nana Surviving Anthropogenic Pressure (the Case of Šepeta Peatland in Northeastern Lithuania). Diversity, 16(9), 512. https://doi.org/10.3390/d16090512