
Man-Made Changes in the Flora and Vegetation of Poland: Current Review

Abstract
1. Introduction
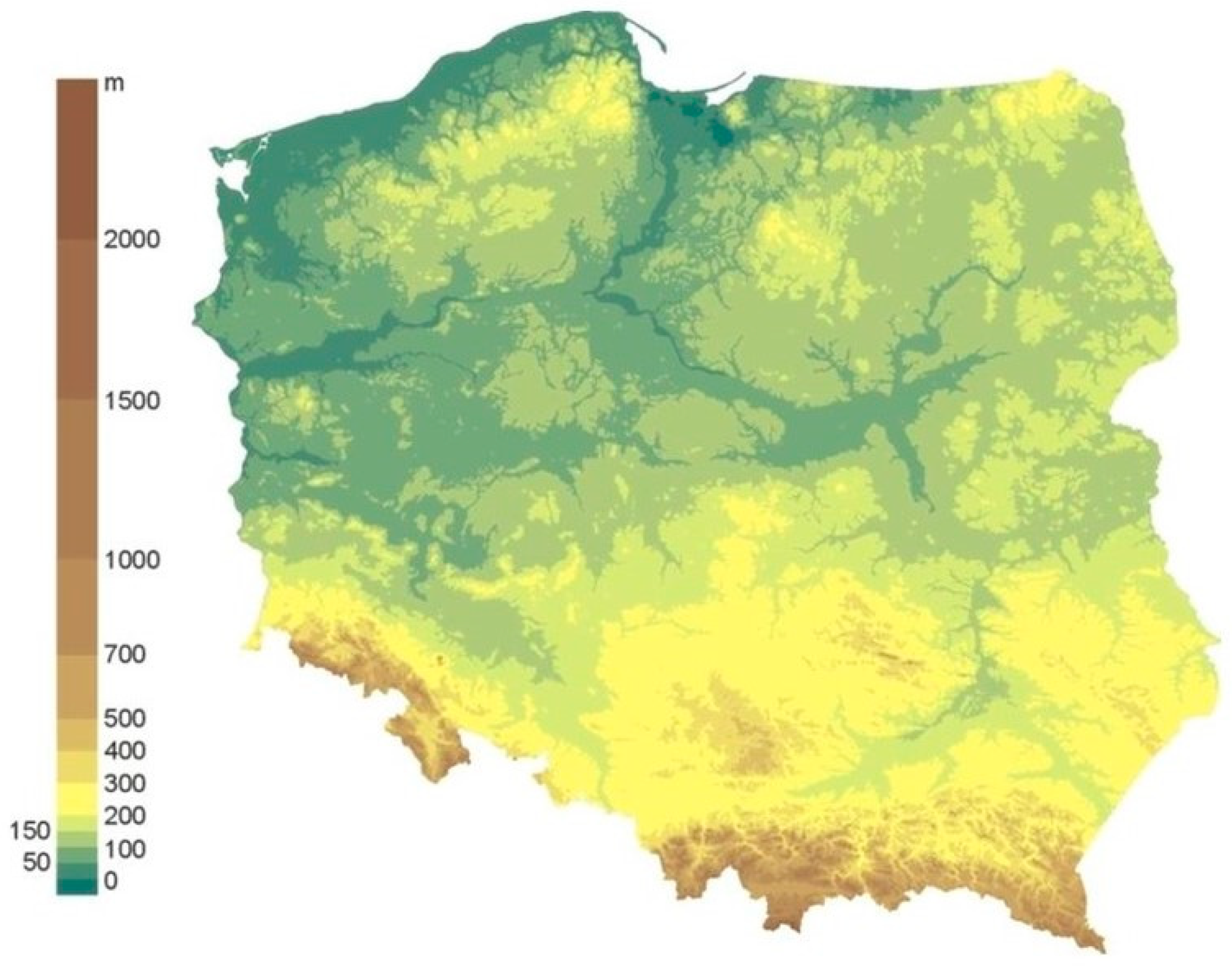
1.1. Study Area: The Location and Factors Influencing Both the Flora and the Vegetation
1.1.1. Natural Factors
1.1.2. Anthropogenic Factors
2. Material and Methods
2.1. Data Sources and Database
2.2. Data Analysis Methods
2.2.1. Geographic-Historical Classification
2.2.2. Classification of Extinct and Declining Species
2.2.3. Synecological Classification
2.3. Methods of Developing and Presenting the Results
- SynI (%)—synanthropization index of flora: the percentage of native species found in habitats created or disturbed by humans (apophytes) and alien species (anthropophytes) in the flora.
- ApI (%)—apophytization index of flora: the percentage of apophytes in the flora.
- AnI (%)—anthropophytization index of flora: the percentage of alien species (anthropophytes) in the flora.
- ArI (%)—archaeophytization index of flora: the percentage of archaeophytes in the flora.
- NeI (%)—neophytization index of flora: the percentage of neophytes in the flora.
- ExI (%)—extinction index of flora: the percentage of extinct and endangered species in the flora.
3. Results
3.1. Geographic-Historical Structure of the Flora
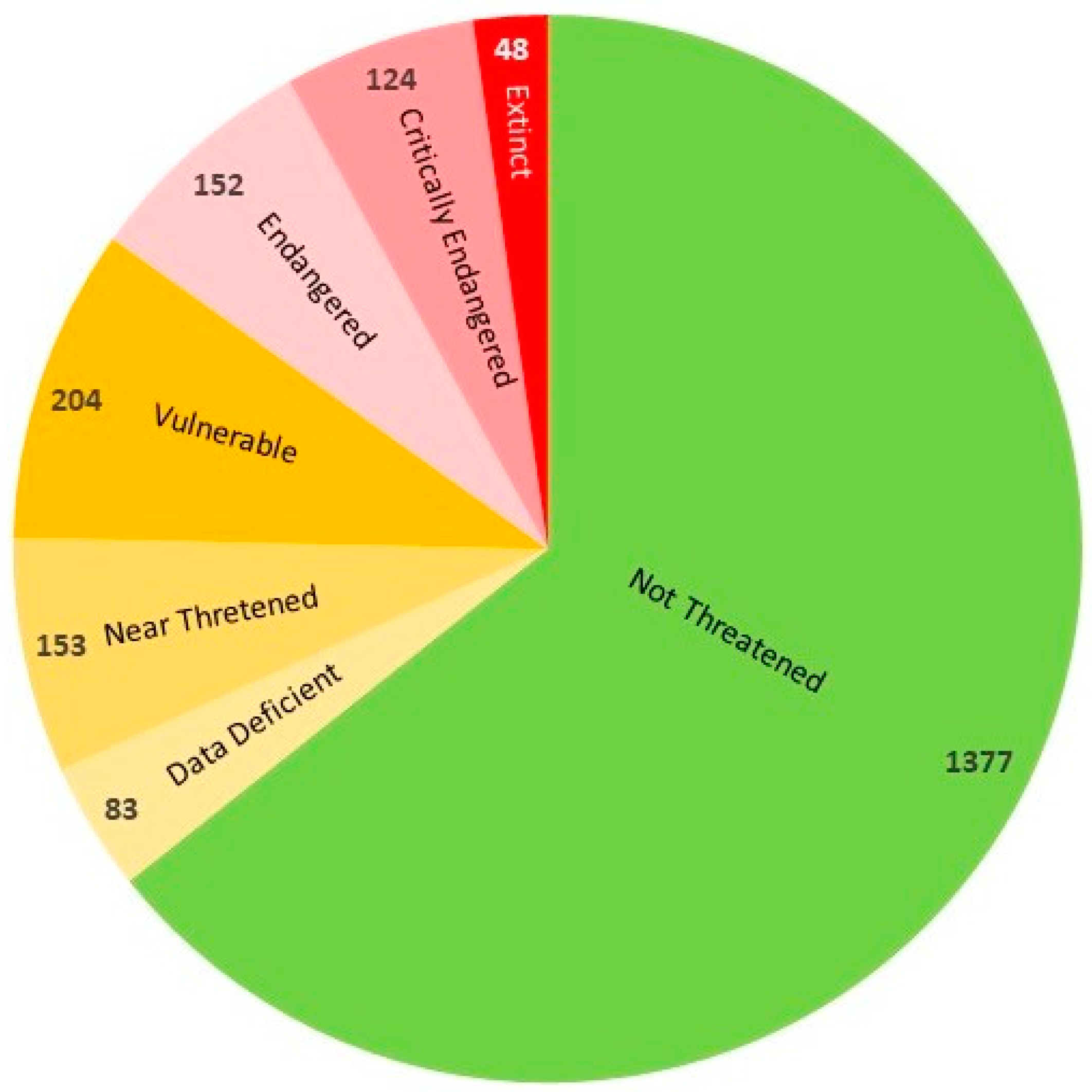
3.2. Species Decline under Human Impact
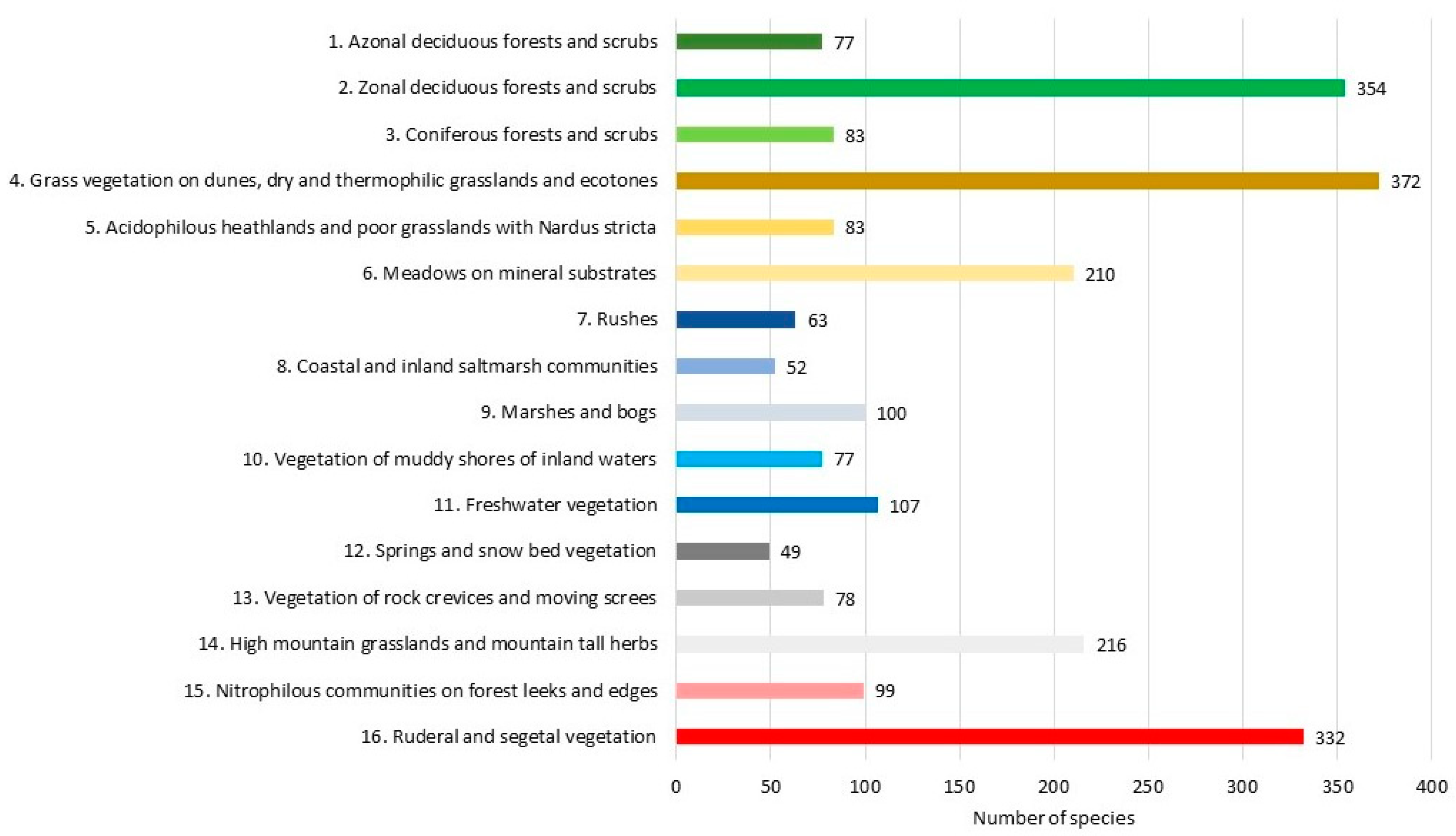
3.3. The Species Composition of Plant Communities
3.4. Process of Human-Influenced Extinction and Expansion of Species
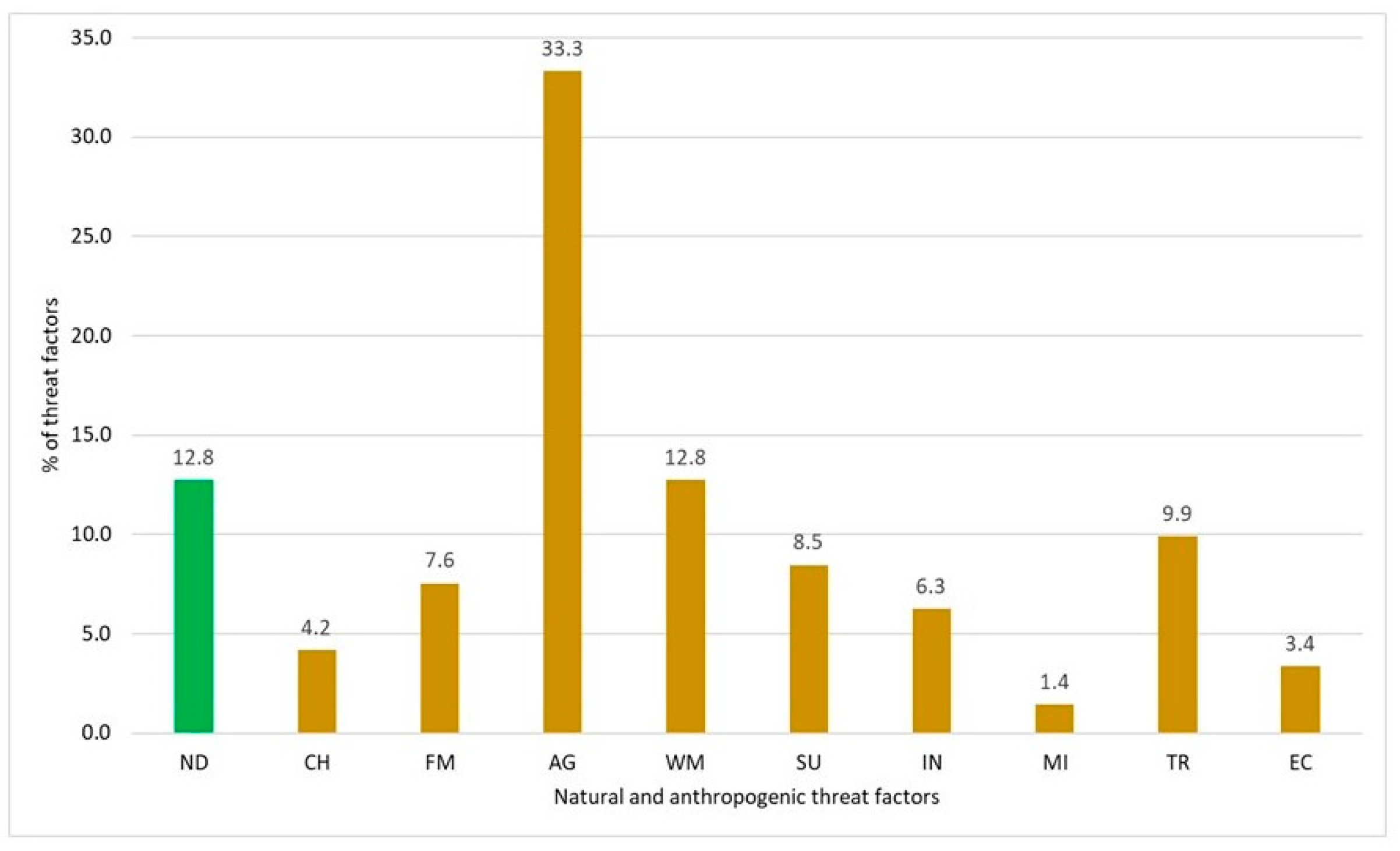
3.4.1. Natural and Anthropogenic Extinction Factors
3.4.2. The Process of Human-Induced Spread of Native Species
3.4.3. Establishment and Naturalization of Alien Species
4. Discussion
4.1. Main Directions of Anthropogenic Flora Changes
4.2. Native Species Adaptation to Man-Made Habitats
4.3. Introduction and Naturalization of Alien Species
4.4. Human-Induced Species Decline
4.5. Human-Influenced Vegetation Changes
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Roberts, N. The Holocene. An Environmental History, 3rd ed.; Wiley-Blackwell: Oxford, UK, 2014; 372p. [Google Scholar]
- Steffen, W.; Crutzen, P.J.; McNeill, J.R. The Anthropocene: Are Humans Now Overwhelming the Great Forces of Nature? In Environment and Society: A Reader; Schlottmann, C., Jamieson, D., Colin Jerolmack, C., Anne Rademacher, A., Eds.; New York University Press: New York, NY, USA, 2017; pp. 12–31. [Google Scholar] [CrossRef]
- Behre, K.E. The role of man in European vegetation history. In Vegetation History. Handbook of Vegetation Science; Huntley, B., Webb, T., Eds.; Springer: Dordrecht, The Netherlands, 1998; Volume 7, pp. 633–672. [Google Scholar]
- Watson, H.C. Cybelle Brittanica: Or British Plants and Their Geographical Relations, I; Longmans: London, UK, 1847; 560p. [Google Scholar]
- Candolle, M.A. De, Géographie Botanique Raisonnée ou Exposition des Faits Principaux et des lois Concentrant La Distribution Geographieque des Plantes de l’epoque Actuelle; Librarie de Victor Masson: Paris, France; Geneva, Switzerland, 1855; 1366p. [Google Scholar]
- Thellung, A. Einteilung der Ruderal und Adventivflora in genetische Gruppen. In Die Flora des Kanton Zürich. I. Teil. Die Ruderal- und Adventivflora des Kanton Zürich; Naegeli, O.O., Thellung, A., Eds.; Vierteljahresschr. Naturforsch. Ges.: Zürich, Switzerland, 1905; Volume 50, pp. 232–236. [Google Scholar]
- Thellung, A. Pflanzenwanderungen unter dem Einfluss des Menschen. Englers Bot. Jb. Leipzig. 1915, 53, 37–66. [Google Scholar]
- Thellung, A. Zur Terminologie der Adventiv und Ruderalflora. Allg. Bot. Zeitschr. Karlsr. 1919, 24, 36–43. [Google Scholar]
- Thellung, A. Kulturpflanzen-Eigenschaften bei Unkräutern. Veröff. Geobot. Inst. Rübel Zürich 1925, 3, 745–762. [Google Scholar]
- Linkola, K. Studien über den Einfluss der Kultur auf die Flora in den Gegenden nördlichem Ladegasee. I. Allgemeiner Teil. Acta Soc. Faun. Flor. Fenn. 1916, 45, 1–432. [Google Scholar]
- Zerbe, S. Restoration of natural broad-leaved woodland in Central Europe on sites with coniferous forest plantations. For. Ecol. Manag. 2002, 167, 27–42. [Google Scholar] [CrossRef]
- Augusto, L.; Dupouey, J.-L.; Ranger, J. Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann. For. Sci. 2003, 60, 823–831. [Google Scholar] [CrossRef]
- Tockner, K.; Stanford, J. Riverine flood plains: Present state and future trends. Environ. Conserv. 2002, 29, 308–330. [Google Scholar] [CrossRef]
- Nobis, A.; Żmihorski, M.; Kotowska, D. Linking the diversity of native flora to land cover heterogeneity and plant invasions in a river valley. Biol. Conserv. 2016, 203, 17–24. [Google Scholar] [CrossRef]
- Gołdyn, H. Changes in plant species diversity of aquatic ecosystems in the agricultural landscape in West Poland in the last 30 years. Biodivers. Conserv. 2010, 19, 61–80. [Google Scholar] [CrossRef]
- Olson, E.R.; Doherty, J.M. Macrophyte diversity–abundance relationship with respect to invasive and native dominants. Aquat. Bot. 2014, 119, 111–119. [Google Scholar] [CrossRef]
- Pavlů, L.; Pavlů, V.; Gaisler, J.; Hejcman, M.; Mikulka, J. Effect of long-term cutting versus abandonment on the vegetation of a mountain hay meadow (Polygono-Trisetion) in Central Europe. Flora 2011, 206, 1020–1029. [Google Scholar] [CrossRef]
- Wesche, K.; Krause, B.; Culmsee, H.; Leuschner, C. Fifty years of change in Central European grassland vegetation: Large losses in species richness and animal-pollinated plants. Biol. Conserv. 2012, 150, 76–85. [Google Scholar] [CrossRef]
- Baessler, C.; Klotz, S. Effects of changes in agricultural land-use on landscape structure and arable weed vegetation over the last 50 years. Agric. Ecosyst. Environ. 2006, 115, 43–50. [Google Scholar] [CrossRef]
- Rühl, A.T.; Eckstein, R.L.; Otte, A.; Donath, T.W. Future challenge for endangered arable weed species facing global warming: Low temperature optima and narrow moisture requirements. Biol. Conserv. 2015, 182, 262–269. [Google Scholar] [CrossRef]
- Kowarik, I. On the role of alien species in urban flora and vegetation. In Plant Invasions-General Aspects and Special Problems; Pyšek, P., Prach, K., Rejmánek, M., Wade, M., Eds.; SPB Academic Publishing: Amsterdam, The Netherlands, 1995; pp. 85–103. Available online: https://link.springer.com/content/pdf/10.1007/978-0-387-73412-5.pdf#page=333 (accessed on 1 March 2023).
- Jackowiak, B. Dynamik der Gefäßpflanzenflora einer Großstadt am Beispiel von Poznan/Polen. Braun-Blanquetia 1989, 3, 89–98. [Google Scholar]
- Pyšek, P. Alien and native species in Central European urban floras: A quantitative comparison. J. Biogeogr. 1998, 25, 155–163. [Google Scholar] [CrossRef]
- Kühn, I.; Brandl, R.; Klotz, S. The flora of German cities is naturally species rich. Evol. Ecol. Res. 2004, 6, 749–764. Available online: https://www.evolutionary-ecology.com/abstracts/v06/1629.html (accessed on 1 March 2023).
- Jehlîk, V.; Dostálek, J.; Frantík, T. Alien plants in Central European river ports. NeoBiota 2019, 45, 93–115. [Google Scholar] [CrossRef]
- Jehlîk, V. Beitrag zur synanthropen (besonders Adventiv-) Flora des Hamburger Hafens. Tuexenia 1981, 1, 81–97. [Google Scholar]
- Brandes, D. Flora und Vegetation der Bahnhöfe Mitteleuropas. Phytocoenologia 1983, 11, 31–115. [Google Scholar] [CrossRef]
- Wittig, R. Ferns in a new role as a frequent constituent of railway flora in Central Europe. Flora 2002, 197, 341–350. [Google Scholar] [CrossRef]
- Wrzesień, M. Kenophytes chorologically related to the habitats of railway grounds in central eastern Poland. Biodiver. Res. Conserv. 2006, 1, 92–94. Available online: http://brc.amu.edu.pl/Kenophytes-chorologically-related-to-the-habitats-of-railway-grounds-in-central-eastern,121552,0,2.html (accessed on 1 March 2023).
- Jehlík, V.; Zaliberová, M.; Májeková, J. The influence of the Eastern migration route on the Slovak flora—A comparison after 40 years. Tuexenia 2017, 37, 313–332. [Google Scholar]
- Lippe, M.; Kowarik, I. Long-Distance Dispersal of Plants by Vehicles as a Driver of Plant Invasions. Conserv. Biol. 2007, 21, 986–996. [Google Scholar] [CrossRef]
- Kornaś, J. Influence of man and his economic activities on the vegetation of Poland. The synanthropic flora. In The Vegetation of Poland; Szafer, W., Ed.; Pergamon Press-PWN: Oxford, UK; Warszawa, Poland, 1966; pp. 93–137. [Google Scholar]
- Kornaś, J. Man’s impact upon the flora: Processes and effects. Memorab. Zool. 1982, 37, 11–30. Available online: https://rcin.org.pl/miiz/dlibra/publication/edition/38769 (accessed on 1 March 2023).
- Kornaś, J. Man’s impact upon the flora and vegetation in Central Europe. In Man’s Impact on Vegetation; Holzner, W., Werger, M.J.A., Ikusima, I., Eds.; Dr. W. Junk Publishers: The Hague, The Netherlands; Boston, MA, USA; London, UK, 1983; pp. 277–286. [Google Scholar]
- Sukopp, H. Der Einfluß des Menschen auf die Vegetation. Vegetatio 1969, 17, 360–371. [Google Scholar] [CrossRef]
- Sukopp, H. Wandel von Flora und Vegetation in Mitteleuropa unter dem Einfluß des Menschen. Ber. Ldw. 1972, 50, 112–139. [Google Scholar]
- Faliński, J.B. Synantropizacja szaty roślinnej—Próba określenia istoty procesu i głównych kierunków badań. Phytocoenosis 1972, 1, 157–170. (In Polish) [Google Scholar]
- Sukopp, H.; Trepl, L. Extinction and Naturalization of Plant Species as Related to Ecosystem Structure and Function. In Ecological Studies; Schulze, E.D., Zwolfer, H., Eds.; Springer: Berlin/Heidelberg, Germany, 1987; Volume 61, pp. 245–276. Available online: https://link.springer.com/chapter/10.1007/978-3-642-71630-0_12 (accessed on 1 March 2023).
- Faliński, J.B. The interpretation of Contemporary Vegetation Transformations on the Basis of the Theories of Synanthropisation and Syndynamics. In Mechanism of Anthropogenic Changes of the Plant Cover; Jackowiak, B., Żukowski, W., Eds.; Publ. Dep. Plant Taxon. AMU: Poznań, Poland, 2000; Volume 10, pp. 9–30. [Google Scholar]
- Daru, B.H.; Davies, T.J.; Willis, C.G.; Meineke, E.K.; Ronk, A.; Zobel, M.; Pärtel, M.; Antonelli, A.; Davis, C.C. Widespread homogenization of plant communities in the Anthropocene. Nat. Commun. 2021, 12, 6983. [Google Scholar] [CrossRef]
- Bartolucci, F.; Galasso, G.; Peruzzi, L.; Conti, F. Report 2020 on plant biodiversity in Italy: Native and alien vascular flora. Nat. Hist. Sci. 2021, 8, 41–54. [Google Scholar] [CrossRef]
- Scherrer, D.; Bürgi, M.; Gessler, A.; Kessler, M.; Nobis, M.P.; Wohlgemuth, T. Abundance changes of neophytes and native species indicate a thermophilisation and eutrophisation of the Swiss flora during the 20th century. Ecol. Indic. 2022, 135, 108558. [Google Scholar] [CrossRef]
- Web of Science Platform. 2023. Available online: https://www-1webofscience-1com-1lbxc51ax2c09.han.amu.edu.pl/wos/woscc/analyze-results/10a4a97c-e438-47e5-8d4f-5cd4b82052df-86701a09 (accessed on 1 March 2023).
- Statistical Yearbook of the Republic of Poland. Statistics Poland. 2022. Available online: https://stat.gov.pl/ (accessed on 1 March 2023).
- Geography of Poland. Available online: https://en.wikipedia.org/wiki/Geography_of_Poland (accessed on 1 March 2023).
- File:Polska Relief.jpg. (27 September 2020). Wikimedia Commons, the Free Media Repository. Retrieved. 1 March 2023. Available online: https://commons.wikimedia.org/w/index.php?title=File:Polska_relief.jpg&oldid=473655244 (accessed on 1 March 2023).
- Climate-Data-org. Available online: https://pl.climate-data.org/europa/polska/masovian-voivodeship/warszawa-4560/ (accessed on 1 March 2023).
- Kornaś, J. Wpływ człowieka i jego gospodarki na szatę roślinną Polski. Flora synantropijna. In Szata roślinna Polski; Szafer, W., Zarzycki, K., Eds.; PWN: Warszawa, Poland, 1977; Volume 1, pp. 95–128. (In Polish) [Google Scholar]
- Symonides, E. Ochrona Przyrody; Wydawnictwo Uniwersytetu Warszawskiego: Warszawa, Poland, 2008; 766p. (In Polish) [Google Scholar]
- Mirek, Z.; Piękoś-Mirkowa, H.; Zając, A.; Zając, M. Vascular Plants of Poland. An Annotated Checklist; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2020; 526p. [Google Scholar]
- Zając, A.; Zając, M. (Eds.) Distribution Atlas of Vascular Plants in Poland; Laboratory of Computer Chorology, Institute of Botany, Jagiellonian University: Kraków, Poland, 2001; 716p. [Google Scholar]
- Zając, A.; Zając, M. (Eds.) Distribution Atlas of Vascular Plants in Poland. Appendix; Institute of Botany, Jagiellonian University: Kraków, Poland, 2019; 321p. [Google Scholar]
- Zarzycki, K.; Trzcińska-Tacik, H.; Różański, W.; Szeląg, Z.; Wołek, J.; Korzeniak, U. Ecological indicator values of vascular plants of Poland. In Biodiversity of Poland; Mirek, Z., Ed.; W. Szafer Institute of Botany, Polish Academy of Sciences: Kraków, Poland, 2002; Volume 1, 183p. [Google Scholar]
- Matuszkiewicz, W. Przewodnik do oznaczania zbiorowisk roślinnych Polski. In Vademecum Geobotanicum; Faliński, J., Ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2001; 537p. (In Polish) [Google Scholar]
- Kaźmierczakowa, R.; Bloch-Orłowska, J.; Celka, Z.; Cwener, A.; Dajdok, Z.; Michalska-Hejduk, D.; Pawlikowski, P.; Szczęśniak, E.; Ziarnek, K. Polska Czerwona Lista Paprotników i Roślin Kwiatowych; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2016; 44p. [Google Scholar]
- Zając, A. Pochodzenie Archeofitów Występujących w Polsce; Rozpr. Habil. Uniw. Jagiell: Kraków, Poland, 1979; Volume 29, 213p. [Google Scholar]
- Zając, M.; Zając, A.; Tokarska-Guzik, B. Extinct and endangered archaeophytes and the dynamics of their diversity in Poland. Biodivers. Res. Conserv. 2009, 13, 17–24. [Google Scholar] [CrossRef]
- Tokarska-Guzik, B. The Establishment and Spread of Alien Plant Species (Kenophytes) in the Flora of Poland; Prace Naukowe Uniw. Śląskiego w Katowicach: Katowice, Poland, 2005; Volume 2372, pp. 1–192. [Google Scholar]
- Tokarska-Guzik, B.; Dajdok, Z.; Zając, M.; Zając, A.; Urbisz, A.; Danielewicz, W.; Hołdyński, C. Rośliny Obcego Pochodzenia w Polsce ze Szczególnym Uwzględnieniem Gatunków Inwazyjnych; Generalna Dyrekcja Ochrony Środowiska: Warszawa, Poland, 2012; 197p. (In Polish) [Google Scholar]
- Urbisz, A. Occurrence of Temporarily-Introduced Alien Plant Species (Ephemerophytes) in Poland—Scale and Assessment of the Phenomenon; Wyd. Uniw. Śląskiego: Katowice, Poland, 2011; 199p. [Google Scholar]
- Kaźmierczakowa, R.; Zarzycki, K.; Mirek, Z. (Eds.) Polska Czerwona Księga Roślin. Paprotniki i Rośliny Kwiatowe, 3rd ed.; Instytut Ochrony Przyrody PAN: Kraków, Poland, 2014; 895p. (In Polish) [Google Scholar]
- Holub, J.; Jirásek, V. Zur Vereinheitlichung der terminologie in der phytogeographie. Folia Geobot. Phytotaxon. 1967, 2, 69–113. [Google Scholar] [CrossRef]
- Kornaś, J. Geograficzno-historyczna klasyfikacja roślin synantropijnych. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1968, 25, 33–41. (In Polish) [Google Scholar]
- Kornaś, J. Oddziaływanie człowieka na florę: Mechanizmy i konsekwencje. Wiadomości Bot. 1981, 25, 165–182. (In Polish) [Google Scholar]
- Schroeder, F.G. Zur Klassifizierung der Antropochoren. Vegetatio 1969, 16, 225–238. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien plants in checklists and floras: Towards better communication between taxonomists and ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Trzcińska-Tacik, H. Flora Synantropijna Krakowa; Rozprawy habilitacyjne UJ: Kraków, Poland, 1979; Volume 22, 278p. (In Polish) [Google Scholar]
- Mirek, Z. Problemy klasyfikacji roślin synantropijnych. Wiadomości Bot. 1981, 25, 45–54. [Google Scholar]
- Guidelines for Application of IUCN Red List Criteria at Regional and National Levels: Version 4.0; IUCN: Gland, Switzerland; Cambridge, UK, 2012; 41p.
- Braun-Blanquet, J. Pflanzensoziologie. Grundzüge der Vegetationskunde; Springer: New York, NY, USA, 1964; 865p. [Google Scholar]
- Jackowiak, B. Antropogeniczne przemiany flory roślin naczyniowych Poznania; Wyd. UAM, Seria Biologia: Poznań, Poland, 1990; Volume 42, 232p. (In Polish) [Google Scholar]
- Jackowiak, B. Methodological proposals for studies on the structure and dynamics of urban flora. Pol. Bot. Stud. 2006, 22, 251–260. [Google Scholar]
- Sudnik-Wójcikowska, B. Studies on Indices of Flora Synanthropization. Flora 1992, 187, 37–50. [Google Scholar] [CrossRef]
- Fukarek, F. Der Mensch beeinflusst die Pflanzenwelt. In Pflanzenwelt der Erde; Fukarek, F., Ed.; Urania Verlag: Jena, Germany; Berlin, Germany; Leipzig, Germany, 1979; pp. 65–77. [Google Scholar]
- Kornaś, J. Wymieranie flory europejskiej—Fakty, interpretacje, prognozy. Phytocoenosis 1976, 5, 173–185. (In Polish) [Google Scholar]
- Ellenberg, H. Vegetation Mitteleuropas mit den Alpen; 5. Stark Veränd. u. Verb. Aufl.; Ulmer: Stuttgart, Germany, 1996; 989p. [Google Scholar]
- Faliński, J.B. Antropogeniczna Roślinność Puszczy Białowieskiej Jako Wynik Synantropizacji Naturalnego Kompleksu Leśnego; Rozpr. Uniw. Warsz.: Warszawa, Poland, 1966; Volume 13, 256p. (In Polish) [Google Scholar]
- Faliński, J.B. Flora i roślinność synantropijna wsi i miast. Próba analizy porównawczej. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1971, 27, 15–38. [Google Scholar]
- Sukopp, H. Apophytes in the flora of Central Europe. Pol. Bot. Stud. 2006, 22, 473–485. Available online: https://www.cabdirect.org/cabdirect/abstract/20083013267 (accessed on 1 March 2023).
- Sukopp, H. Apophyten in der Flora von Mitteleuropa. Braunschw. Geobot. Arb. 2008, 9, 443–458. [Google Scholar]
- Kopecký, K. Der Apophytisierungsprozess und die Apophytengesellschaften der Galio-Urticetea mit einigen Beispielen aus der südwestlichen Umgebung von Praha. Fol. Geobot. Phytotax. 1984, 19, 113–138. [Google Scholar] [CrossRef]
- Kopecký, K. Der Apophytisierungsprozess am Beispiel der Saumgesellschaften mit Chaerophyllum aromaticum L. in der Tschechoslowakei. Tuexenia 1985, 5, 127–130. [Google Scholar]
- Kopecký, K. Einfluss der Strassen auf die Synanthropisierung der Flora und Vegetation nach Beobachtungen in der Tschechoslovakei. Folia Geobot. Phytotaxon. 1988, 23, 145–172. [Google Scholar] [CrossRef]
- Wittig, R. The origin and development of the urban flora of Central Europe. Urban Ecosyst. 2004, 7, 323–339. [Google Scholar] [CrossRef]
- Jackowiak, B. Modele Ekspansji Roślin Synantropijnych i Transgenicznych; Wydawn. Uniw. Warszawskiego: Warsaw, Poland, 1999; Volume 6, pp. 3–16. (In Polish) [Google Scholar]
- Trzcińska-Tacik, H.; Wasylikowa, K. History of the synanthropic changes of flora and vegetation of Poland. Memorab. Zool. 1982, 3, 47–69. [Google Scholar]
- Latałowa, M.; Jarosińska, J.; Badura, M. Elbląg średniowieczny w świetle dotychczasowych materiałów archeobotanicznych. Archeol. Pol. 1998, 43, 147–166. (In Polish) [Google Scholar]
- Woch, M.W. Antropofity znalezione w trakcie badań archeobotanicznych średniowiecznego Krakowa. In Rośliny w Życiu Codziennym Mieszkańców Średniowiecznego Krakowa; Mueller-Bieniek, A., Ed.; Instytut Botaniki im. W. Szafera PAN: Kraków, Poland, 2012; pp. 185–210. (In Polish) [Google Scholar]
- Tokarska-Guzik, B. Invasive ability of kenophytes occurring in Poland: A tentative assessment. In Biological Invasions—From Ecology to Control; Nentwig, N., Bacher, S., Cock, M., Dietz, H.J., Gigon, A., Wittenberg, R., Eds.; Neobiota; Pensoft Publishers: Sofia, Bulgaria, 2005; Volume 6, pp. 47–65. [Google Scholar]
- Faliński, J.B. Stadia neofityzmu i stosunek neofitów do innych komponentów zbiorowiska. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1968, 25, 15–31. (In Polish) [Google Scholar]
- Faliński, J.B. Zbiorowiska autogeniczne i antropogeniczne. Próba określenia i klasyfikacji. Dyskusje fitosocjologiczne. Ekol. Pol. Ser. B 1969, 15, 173–182. (In Polish) [Google Scholar]
- Faliński, J.B. Invasive alien plants and vegetation dynamics. In Plant Invasions: Ecological Mechanisms and Human Responses; Starfinger, U., Edwards, K., Kowarik, I., Williamson, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1998; pp. 3–21. [Google Scholar]
- Udvardy, M.D.F. Dynamic Zoogeography with Special Reference to Land Animals; Litton Educational Publishing, Inc.: New York, NY, USA; Van Nostrand Reinhold Company: New York, NY, USA, 1969; 445p. [Google Scholar]
- Danihelka, J.; Chrtek Jr, J.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. Available online: https://www.cabdirect.org/cabdirect/abstract/20123395489 (accessed on 1 March 2023).
- Kühn, I.; Klotz, S. The alien flora of Germany—Basics from a new German database. In Plant Invasions: Ecological Threats and Management Solutions; Child, L.E., Ed.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 90–100. [Google Scholar]
- Medvecká, J.; Kliment, J.; Májeková, J.; Halada, L.; Zaliberová, M.; Gojdičová, E.; Feráková, V.; Jarolimek, L. Inventory of the alien flora of Slovakia. Preslia 2012, 84, 257–309. Available online: https://www.cabdirect.org/cabdirect/abstract/20123299140 (accessed on 1 March 2023).
- Piotrowska, H.; Żukowski, W.; Jackowiak, B. Rośliny Naczyniowe Słowińskiego Parku Narodowego; Prace Zakładu Taksonomii Roślin UAM w Poznaniu, Bogucki Wyd. Nauk.: Poznań, Poland, 1997; Volume 6, 216p. (In Polish) [Google Scholar]
- Żukowski, W.; Latowski, K.; Jackowiak, B.; Chmiel, J. Rośliny naczyniowe Wielkopolskiego Parku Narodowego; Prace Zakładu Taksonomii Roślin UAM w Poznaniu, Bogucki Wyd. Nauk.: Poznań, Poland, 1995; Volume 4, 231p. (In Polish) [Google Scholar]
- Sudnik-Wójcikowska, B. Flora Miasta Warszawy i jej Przemiany w Ciągu XIX i XX Wieku; Wyd. UW: Warszawa, Poland, 1987; Volume 1, 242p, Volume 2, 435p. (In Polish) [Google Scholar]
- Chmiel, J. Flora Roślin Naczyniowych Wschodniej Części Pojezierza Gnieźnieńskiego i jej Antropogeniczne Przemiany; Prace Zakładu Taksonomii Roślin UAM w Poznaniu, Wyd. Sorus: Poznań, Poland, 1993; 202p. (In Polish) [Google Scholar]
- Urbisz, A. Flora naczyniowa Płaskowyżu Rybnickiego na tle antropogenicznych przemian tego obszaru. Scr. Rudensia 1996, 6, 174. (In Polish) [Google Scholar]
- Lambdon, P.W.; Pyšek, P.; Basnou, C.; Delipetrou, P.; Essl, F.; Hejda, M.; Jarošík, V.; Pergl, J.; Winter, M.; Andriopoulos, P.; et al. Alien flora of Europe: Species diversity, geographical pattern and state of the art of research. Preslia 2008, 80, 101–149. Available online: http://www.preslia.cz/P082Lam.pdf (accessed on 1 March 2023).
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Panetta, F.D.; West, C.C. Naturalization and invasion of alien plants: Concepts and definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Protopopova, V.V.; Shevera, M.; Fedoronchuk, M.M. The geographical analysis of apophytes fraction of urban floras of Ukraine. Thaiszhia J. Bot. Kosice 2012, 22, 181–189. Available online: https://www.upjs.sk/public/media/7803/181-189-protopopova-et-al-upr.pdf (accessed on 1 March 2023).
- Zając, A. Studies on the origin of archaeophytes in Poland. Part I. Methodical consideration. Zesz. Nauk. Uniw. Jagiellońskiego Prace Bot. 1983, 11, 87–107. [Google Scholar]
- Zając, A. Studies on the origin of archaeophytes in Poland. Part II. Taxa of Mediterranean and Atlantic-Mediterranean origin. Zesz. Nauk. Uniw. Jagiellońskiego Prace Bot. 1987, 14, 7–50. [Google Scholar]
- Zając, A. Studies on the origin of archaeophytes in Poland. Part III. Taxa of Irano-Turanian, Euro-Siberian-Irano-Turanian and Mediterranean-Irano-Turanian origin. Zesz. Nauk. Uniw. Jagiellońskiego Pr. Bot. 1987, 15, 93–129. [Google Scholar]
- Zając, A. Studies on the origin of archaeophytes in Poland. Part IV. Taxa of Pontic-Pannonian, Mediterraneo-South Asiatic, South Asiatic and Middle European origin, archaeophyta anthropogena, Archaeophyta resistentia, Archaeophytes of unknown origin. Zesz. Nauk. Uniw. Jagiellońskiego Pr. Bot. 1988, 11, 87–107. [Google Scholar]
- Kornaś, J. Prowizoryczna liczba nowszych przybyszów synantropijnych (kenofitów) zadomowionych w Polsce. Mater. Zakł. Fitosoc. Stos. Uniw. Warsz. 1968, 25, 43–53. (In Polish) [Google Scholar]
- Eliáš, P.; Dítě, D.; Kliment, J.; Hrivnák, R.; Feráková, V. Red list of ferns and flowering plants of Slovakia, 5th edition (October 2014). Biologia 2015, 70, 218–228. [Google Scholar] [CrossRef]
- Grulich, V. Red List of vascular plants of the Czech Republic. Preslia 2012, 84, 631–645. Available online: https://www.cabdirect.org/cabdirect/abstract/20123395488 (accessed on 1 March 2023).
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M.; et al. Red list of threatened vascular plants in Italy. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- Moreno, J.C. Lista Roja 2008 de la Flora Vascular Española. Dirección General de Medio Natural y Política Forestal; Ministerio de Medio Ambiente, y Medio Rural y Marino, y Sociedad Española de Biología de la Conservación de Plantas: Madrid, Spain, 2008; 86p. [Google Scholar]
- Stroh, P.; Leach, S.J.; August, T.A.; Walker, K.J.; Pearman, D.A.; Rumsey, F.J.; Harrower, C.A.; Fay, M.F.; Martin, J.P.; Pankhurst, T. A vascular plant red list for England. BSBI News 2014, 9, 1–193. [Google Scholar]
- Jäger, E.J. Veränderungen des Artenbestandes von Floren unter dem Einfluß des Menschen. Biol. Rundsch. 1977, 15, 287–300. [Google Scholar]
- Jäger, E.J. Möglichkeiten der Prognose synanthroper Pflanzenausbreitungen. Flora 1988, 180, 101–131. Available online: https://www.sciencedirect.com/science/article/abs/pii/S036725301730302X (accessed on 1 March 2023). [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011; 130p. [Google Scholar]
- Le Roux, J.J.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent Anthropogenic Plant Extinctions Differ in Biodiversity Hotspots and Coldspots. Curr. Biol. 2019, 29, 2912–2918. Available online: https://www.sciencedirect.com/science/article/pii/S0960982219309431 (accessed on 1 March 2023). [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category Name | Classification Criteria | Explanation |
|---|---|---|
| 1 Native plants | Taxa area of origin | Taxa that have originated in a given area without human involvement or that have arrived there without any intentional or unintentional intervention of humans from an area in which they are native [66]. |
| 1.1 Native non-synanthropic plants | Habitats occupied in the native area | Taxa whose occurrence is limited to natural and seminatural habitats. |
| 1.2 Native synanthropic plants (Apophytes) | Taxa found exclusively in man-made habitats (especially ruderal and segetal habitats) or occupying them permanently alongside natural and seminatural habitats. | |
| 2 Alien plants (Anthropophytes) | Taxa area of origin | Taxa in a given area (see below) whose presence there is due to intentional or unintentional human involvement, or which have arrived there without the help of people from an area in which they are alien [66]. |
| 2.1 Naturalized (established) plants (Metaphytes of the Trzcińska-Tacik [67] and Kornaś [64]) sense | Persistence of establishment | Permanently established anthropophytes [64,67]. Alien taxa that sustain self-replacing populations for at least 10 years without direct intervention by people (or in spite of human intervention) by recruitment from the seed or ramets (tillers, tubers, bulbs, fragments, etc.) capable of independent growth [66]. |
| 2.1.1 Archaeophytes | Time of arrival (introduction) in the study area | Alien taxa introduced more or less before the year 1500, both intentionally and accidentally, regardless of the degree of establishment (naturalization). |
| 2.1.2 Neophytes (Kenophytes of the Kornaś [64]) sense | Alien taxa introduced more or less after the year 1500, both intentionally and accidentally, regardless of the degree of establishment (naturalization). | |
| 2.1.2.1 Epecophytes | Degree of establishment (naturalization) | Neophytes established in human-made habitats. |
| 2.1.2.2 Hemiagriofity | Neophytes established in seminatural habitats. | |
| 2.1.2.3 Holoagriophytes | Neophytes established in natural habitats. | |
| 2.2 Casual alien plants (Diaphytes of the Trzcińska-Tacik [67] and Kornaś [64]) sense | Persistence of establishment | Alien plants that may flourish and even reproduce occasionally outside of cultivation in an area, but that eventually die out, because they do not form self-replacing populations and rely on repeated introductions for their persistence [66]. |
| 2.2.1 Ephemerophytes | Source and way of importation | Plants introduced into an area without intentional human involvement. |
| 2.2.2 Ergasiophytes | Alien plants escaping from crops grown in the area. |
| Number and Name of the Synecological Group |
|---|
| Description of the Synecological Group Class Names |
| 1. Azonal deciduous forests and scrubs |
| Willow-poplar forests and thickets, found in river valleys on sandy, gravelly, or stony alluviums, with poplar (Populus alba L., P. nigra L.) and willow (Salix alba L.) and deciduous forests with the prevailing alder (Alnus glutinosa (L.) Gaertn.) developing on wet peaty and peaty-mineral soils. Salicetea purpureae Moor 1958; Alnetea glutinosae Br.-Bl. et R. Tx. 1943. |
| 2. Zonal deciduous forest and scrubs Deciduous forests and scrubs shedding their leaves for the winter, found on mineral soils of varying nutrient content, moisture content and reaction. The forest stand is made up of elm (Ulmus laevis Pall., U. minor Mill., U. glabra Huds.), ash (Fraxinus excelsior L.), oak (Quercus robur L., Q. petraea (Matt.) Liebl.), lime (Tilia cordata Mill., T. platyphyllos Scop.), and beech (Fagus sylvatica L.). Querco-Fagetea Br.-Bl. et Vlieg. 1937; Quercetea robori-petraeae Br.-Bl. et R. Tx. 1943; Rhamno-Prunetea Moor 1958. |
| 3. Coniferous forests and scrubs |
| Acidophilous, oligo-, and mesotrophic communities with a predominance of coniferous species (Pinus L., Picea A. Dietr., Abies (L.) H. Karst), shrubs and abundant mosses; xerothermic forest and scrub communities dominated by pine (Pinus sylvestris L.). Vaccinio-Piceetea Br.-Bl. 1939; Erico-Pinetea Horvat 1959. |
| 4. Grass vegetation on dunes, dry and thermophilic grasslands and ecotones |
| The group encompasses a wide spectrum of vegetation; from communities of grand grasses on the coastal dunes, through low grasslands overgrowing soils with an elevated heavy metal content, dry grasslands on a mineral substrate to thermophilic grasslands, and communities forming an ecotone at the forest boundary. Ammophiletea Br.-Bl. et R. Tx. 1943; Violetea calaminariae Br.-Bl. et R.Tx. 1943; Koelerio-Corynephoretea canescentis Klika in Klika et Novak 1941; Festuco-Brometea Br.-Bl. et R.Tx. 1943; Trifolio-Geranietea sanguinei Th. Müller 1962. |
| 5. Acidophilous heathlands and poor grasslands with Nardus stricta |
| Semi-natural and anthropogenic communities, which in the primary landscape occupied small areas under specific environmental conditions and as a result of human activity have spread significantly. These include shrub communities with Calluna vulgaris (L.) Hull as well as acidophilous low grasslands and meadows with the dominant grass Nardus stricta L. Nardo-Callunetea Prsg 1949. |
| 6. Meadows on mineral substrates |
| Semi-natural and anthropogenic turf meadow and pasture communities on meso- and eutrophic undrained mineral soils. Meadows of this type also occupy organic-mineral soils and desiccated low peats. Molinio-Arrhenatheretea R.Tx. 1937. |
| 7. Rushes |
| Communities of high grasses and sedges with dicotyledonous herbaceous perennials, found in the littoral and riparian zone of the inland reservoirs of standing and flowing water. Phragmitetea R. Tx. et Prsg 1942. |
| 8. Coastal and inland saltmarsh communities |
| Extremely poor “submarine meadows” found in the littoral zone of the Baltic Sea; therophyte communities developing along the Baltic coastline; communities of the initial stages of the coastal dune formation, communities of muddy, highly saline habitats (both coastal and inland); rush-meadow communities found in the supralittoral of salt water bodies. Zosteretea marinae Pign. 1953; Rupietea maritimae J. Tx. 1960; Cakiletea maritimae R. Tx. et Prsg 1950; Thero-Salicornietea Pign. 1953 em. R. Tx. 1954 in R. Tx. et Oberd. 1958; Asteretea tripolium Westh. et Beeft. ap. Beeft 1962; Ammophiletea Br.-Bl. et R. Tx. 1943 (Agropyro-Honckenyion R. Tx. 1952). |
| 9. Marshes and bogs |
| Wet heath and raised bogs on acidic and dystrophic habitats, fed exclusively or predominantly by rainwater, as well as low-sedge swamp meadow communities, transitional and low bogs. Oxycocco-Sphagnetea Br.-Bl. et R. Tx. 1943; Scheuchzerio-Caricetea nigrae (Nordh. 1937) R. Tx. 1937. |
| 10. Vegetation of muddy shores of the inland waters |
| Moderately nitrophilous, mostly natural summer and autumn therophyte communities found on the drying shores of the inland water bodies. Bidentetea tripartiti R. Tx., Lohm. et Prsg. 1950; Isoëto-Nanojuncetea Br.-Bl. et R. Tx. 1943. |
| 11. Freshwater vegetation |
| Communities of water reefs (Lemna L.) forming aggregations on the surface of still and slow-moving waters, as well as communities of small perennials and macrophytes found in the waters with a wide range of nutrients. Lemnetea R. Tx. 1955; Potametea R. Tx. et Prsg; Littorelletea Br.-Bl. et R. Tx. 1943; Utricularietea intermedio-minoris Den Hartog et Segal 1964 em. Pietsch 1965. |
| 12. Springs and snow bed vegetation |
| Spring communities, often with a significant proportion of bryophytes, as well as high-mountain snow beds communities, i.e., places with a shortened growing season due to prolonged snow. Montio-Cardaminetea Br.-Bl. et R. Tx. 1943; Salicetea herbaceae Br.-Bl. et al. 1947. |
| 13. Vegetation of rock crevices and moving screes |
| Natural communities of rock crevices in the mountains and anthropogenic communities in the secondary habitats (walls, ruins), as well as pioneer communities of mobile or poorly consolidated scree in the rocky parts of mountains or in the mountainous sections of river valleys. Asplenietea rupestria Br.-Bl. 1934 in Meier et Br.-Bl. 1934; Thlaspietea rotundifolii Br.-Bl. in Br.-Bl. et Jenny 1926 em. Seibert 1977. |
| 14. High mountain grasslands and mountain tall herbs |
| High mountain grasslands on calciophilic and acidophilic substrates, as well as high mountain herb and grassland. The latter are the communities of tall dicotyledonous perennials or grasses found in relatively fertile moist or wet habitats. |
| Seslerietea variae Br.-Bl. 1948 em. Oberd. 1978; Juncetea trifidi Hadač in Klika et Hadač 1944; Betulo-Adenostyletea Br.-Bl. 1948 |
| 15. Nitrophilous communities on forest leeks and edges |
| Nitrophilous communities of therophytes, perennials, and shrubs that initiate forest regeneration after the removal or destruction of a forest stand, as well as natural and semi-natural nitrophilous communities in fertile fresh, moist or wet habitats, forming ecotones at the contact between woody and herbaceous communities. Epilobietea angustifolii R. Tx. et Prsg. 1950; Artemisietea vulgaris Lohm., Prsg et R. Tx. in R. Tx. 1950; (Galio-Urticenea Pass. 1967). |
| 16. Ruderal and segetal vegetation |
| Anthropogenic weed communities of cultivated fields and annual and perennial plants overgrowing the ruderal habitats (roadsides, balks, trackways, rubble pits, etc.), as well as thermophilic rhizomatous plant communities found on the mineral grounds. Stellarietea mediae R. Tx., Lohm. et Prsg. 1950; Agropyretea intermedio-repentis (Oberd. et al. 1967) Müller et Görs 1969; Artemisietea vulgaris Lohm., Prsg. et R. Tx. in R. Tx. 1950 (Artemisienea vulgaris Th. Müller 1981 in Oberd. 1983). |
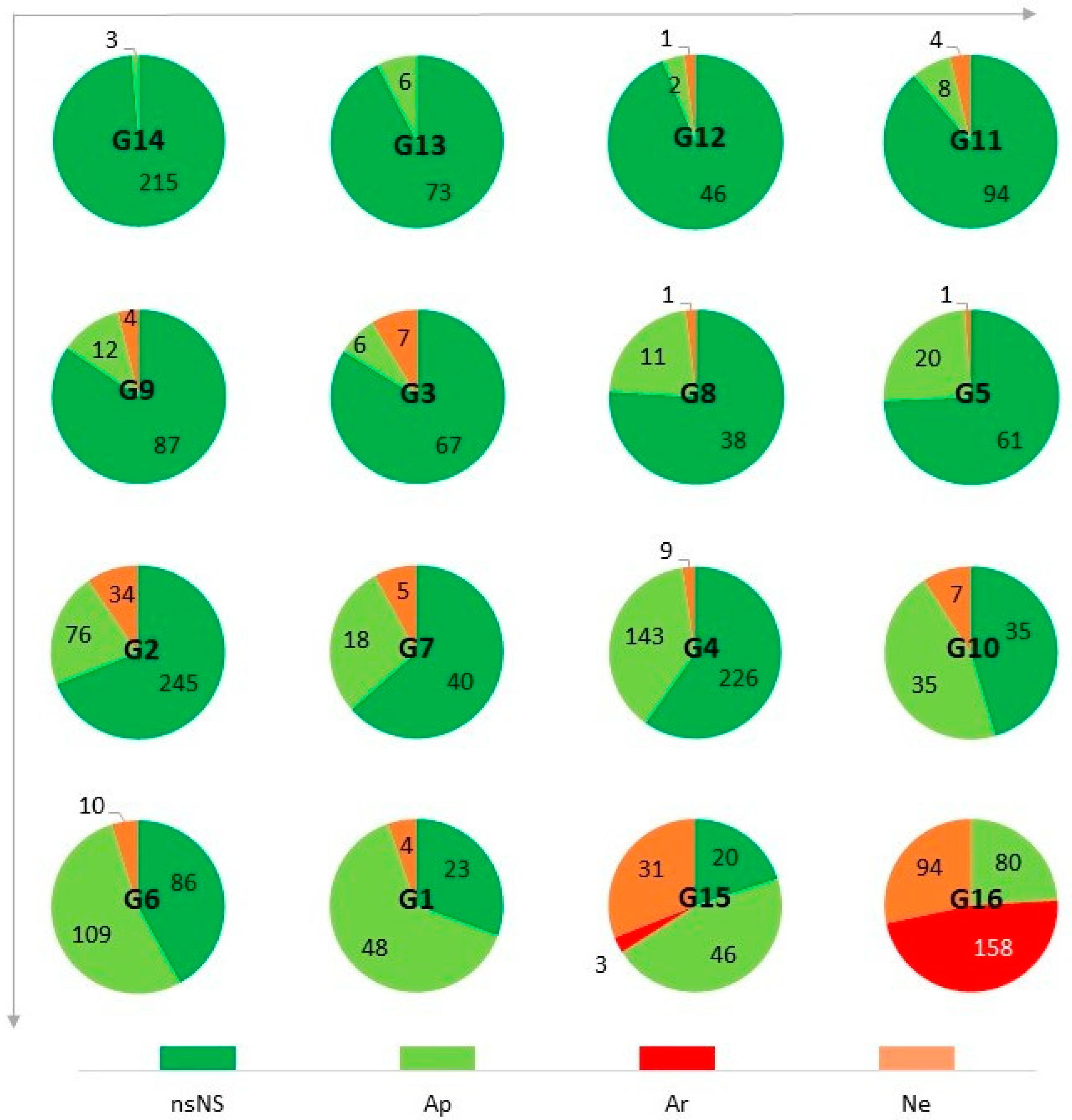
| SynI (%) | ApI (%) | AnI (%) | ArI (%) | NeI (%) | ExI (%) | |
|---|---|---|---|---|---|---|
| G1 | 69.3 | 64.0 | 5.3 | 0.0 | 5.3 | 4.2 |
| G2 | 31.0 | 21.4 | 9,6 | 0.0 | 9.6 | 20.9 |
| G3 | 16.3 | 7.5 | 8.8 | 0.0 | 8.8 | 37.0 |
| G4 | 40.2 | 37.8 | 2.4 | 0.0 | 2.4 | 49.1 |
| G5 | 25.6 | 24.4 | 1.2 | 0.0 | 1.2 | 37.0 |
| G6 | 58.0 | 53.2 | 4.9 | 0.0 | 4.9 | 27.7 |
| G7 | 36.5 | 28.6 | 7.9 | 0.0 | 7.9 | 31.0 |
| G8 | 24.0 | 22.0 | 2.0 | 0.0 | 2.0 | 79.6 |
| G9 | 15.5 | 11.7 | 3.9 | 0.0 | 3.9 | 69.7 |
| G10 | 54.5 | 45.5 | 9.1 | 0.0 | 9.1 | 51.4 |
| G11 | 11.3 | 7.5 | 3.8 | 0.0 | 3.8 | 53.9 |
| G12 | 6.1 | 4.1 | 2.0 | 0.0 | 2.0 | 31.3 |
| G13 | 7.6 | 7.6 | 0.0 | 0.0 | 0.0 | 40.5 |
| G14 | 1.4 | 1.4 | 0.0 | 0.0 | 0.0 | 32.1 |
| G15 | 80.0 | 46.0 | 34.0 | 3.0 | 31.0 | 11.6 |
| G16 | 100.0 | 24.1 | 75.9 | 47.6 | 28.3 | 25.1 |
| AVG | 36.1 | 25.4 | 10.7 | 3.2 | 7.5 | 37.6 |
| STD | 29.0 | 19.1 | 19.2 | 11.9 | 9.2 | 19.7 |
| Example Species | Threat Category | Freq. Class | Synecol. Group | Geo-Hist. Status |
|---|---|---|---|---|
| A. Species occurring only in natural and semi-natural communities | ||||
| Osmunda regalis L. | VU | 3 | 1 | nsNS |
| Corydalis pumila (Host) Rchb. | VU | 2 | 2 | nsNS |
| Neottianthe cucullata (L.) Schltr. | CR | 1 | 3 | nsNS |
| Hypericum elegans Stephan ex Willd. | CR | 1 | 4 | nsNS |
| Spiranthes spiralis (L.) Chevall. | CR | 1 | 5 | nsNS |
| Gladiolus paluster Gaudin | CR | 1 | 6 | nsNS |
| Apium nodiflorum (L.) Lag. | CR | 1 | 7 | nsNS |
| Ruppia maritima L. | VU | 1 | 8 | nsNS |
| Schoenus nigricans L. | EN | 1 | 9 | nsNS |
| Apium repens (Jacq.) Lag. | CR | 1 | 10 | nsNS |
| Isoëtes echinospora Durieu | CR | 1 | 11 | nsNS |
| Cochlearia polonica E. Fröhl. | EW | 1 | 12 | nsNS |
| Asplenium adiantum-nigrum L. | EN | 1 | 13 | nsNS |
| Androsace obtusifolia All. | EN | 1 | 14 | nsNS |
| Bromus ramosus Huds. | VU | 2 | 15 | nsNS |
| Carex elongata L. | NotT | 4 | 1 | nsNS |
| Pulmonaria officinalis L. s. s. | NotT | 2 | 2 | nsNS |
| Luzula sylvatica (Huds.) Gaudin | NotT | 3 | 3 | nsNS |
| Eryngium campestre L. | NotT | 2 | 4 | nsNS |
| Galium saxatile L. | NotT | 2 | 5 | nsNS |
| Cirsium canum (L.) All. | NotT | 3 | 6 | nsNS |
| Carex cespitosa L. | NotT | 4 | 7 | nsNS |
| Festuca salina Natho & Stohr | NotT | 1 | 8 | nsNS |
| Carex flava L. | NotT | 4 | 9 | nsNS |
| Peplis portula L. | NotT | 4 | 10 | nsNS |
| Najas major All. | NotT | 1 | 11 | nsNS |
| Stellaria uliginosa Murray | NotT | 4 | 12 | nsNS |
| Cystopteris sudetica A. Braun & Milde | NotT | 2 | 13 | nsNS |
| Gentiana frigida Haenke | NotT | 1 | 14 | nsNS |
| Bromus benekenii (Lange) Trimen | NotT | 3 | 15 | nsNS |
| B. Species occurring in natural and semi-natural communities and sporadically found in anthropogenic habitats | ||||
| Artemisia pontica L. | CR | 1 | 4 | nsNS |
| Asplenium septentrionale (L.) Hoffm. | VU | 2 | 13 | nsNS |
| Carex buekii Wimm. | NT | 1 | 7 | nsNS |
| Carex secalina Wahlenb. | CR | 1 | 8 | nsNS |
| Cirsium decussatum Janka | VU | 1 | 6 | nsNS |
| Orchis pallens L. | VU | 2 | 2 | nsNS |
| Phelipanche arenaria (Borkh.) Pomel | CR | 1 | 4 | nsNS |
| Sagina ciliata Fr. | CR | 1 | 10 | nsNS |
| C. Species occurring in natural and semi-natural communities and permanently established in anthropogenic habitats | ||||
| Plantago coronopus L. | CR | 1 | 8 | Ap |
| Spergularia salina J. Presl & C. Presl | VU | 3 | 8 | Ap |
| Triglochin maritimum L. | VU | 2 | 8 | Ap |
| Aegopodium podagraria L. | NotT | 5 | 2 | Ap |
| Asplenium ruta-muraria L. | NotT | 3 | 13 | Ap |
| Bidens tripartita L. | NotT | 5 | 10 | Ap |
| Calamagrostis epigejos (L.) Roth | NotT | 5 | 15 | Ap |
| Campanula rapunculoides L. | NotT | 5 | 4 | Ap |
| Deschampsia caespitosa (L.) P. Beauv. | NotT | 5 | 6 | Ap |
| Epilobium hirsutum L. | NotT | 5 | 7 | Ap |
| Epilobium obscurum Schreb. | NotT | 3 | 12 | Ap |
| Juncus articulatus L. em. K. Richt. | NotT | 5 | 9 | Ap |
| Lemna minor L. | NotT | 5 | 11 | Ap |
| Luzula campestris (L.) DC. | NotT | 5 | 5 | Ap |
| Solanum dulcamara L. | NotT | 5 | 1 | Ap |
| Sorbus aucuparia L. em. Hedl. | NotT | 5 | 3 | Ap |
| Thalictrum aquilegiifolium L. | NotT | 4 | 14 | Ap |
| Verbascum densiflorum Bertol. | NotT | 4 | 16 | Ap |
| D. Species occurring exclusively in man-made habitats, lacking contact with natural and semi-natural communities | ||||
| Arabidopsis thaliana (L.) Heynh. | NotT | 4 | 16 | Ap-AF |
| Arnoseris minima (L.) Schweigg. & Körte | NotT | 4 | 16 | Ap-AF |
| Berteroa incana (L.) DC. | NotT | 4 | 16 | Ap-RC |
| Melilotus officinalis (L.) Pall | NotT | 5 | 16 | Ap-RC |
| Example Species | Threat Category | Freq. Class | Synecol. Group | Geo-Hist. Status |
|---|---|---|---|---|
| A. Casual alien species | ||||
| Aegilops ligustica Asch. & Graebn. | nc | 1 | nc | Cas-Eh |
| Amaranthus deflexus L. | nc | 1 | nc | Cas-Eh |
| Bidens pilosa L. | nc | 1 | nc | Cas-Eh |
| Citrus aurantium L. | nc | 1 | nc | Cas-Eh |
| Rumex triangulivalvis (Danser) Rech.f. | nc | 1 | nc | Cas-Eh |
| Sideritis montana L. | nc | 1 | nc | Cas-Eh |
| Consolida orientalis (J. Gay) Schrödinger | nc | 1 | nc | Cas-Er |
| Corylus colurna L. | nc | 1 | nc | Cas-Er |
| Cynosurus echinatus L. | nc | 1 | nc | Cas-Er |
| Daucus aureus Desf. | nc | 1 | nc | Cas-Er |
| Glaucium flavum Crantz | nc | 1 | nc | Cas-Er |
| B. Established alien species | ||||
| B1. Archaeophytes | ||||
| Camelina alyssum (Mill.) Thell. | EX | 2 | 16 | Ar-AF |
| Caucalis platycarpos L. | CR | 2 | 16 | Ar-AF |
| Ranunculus arvensis L. | EN | 3 | 16 | Ar-AF |
| Atriplex rosea L. | VU | 2 | 16 | Ar-RC |
| Chenopodium murale L. | EN | 3 | 16 | Ar-RC |
| Chenopodium vulvaria L. | EN | 3 | 16 | Ar-RC |
| Apera spica-venti (L.) P. Beauv. | NotT | 5 | 16 | Ar-AF |
| Consolida regalis Gray | NotT | 4 | 16 | Ar-AF |
| Papaver rhoeas L. | NotT | 4 | 16 | Ar-AF |
| Lamium album L. | NotT | 5 | 16 | Ar-RC |
| Artemisia absinthium L. | NotT | 4 | 16 | Ar-RC |
| Verbena officinalis L. | NotT | 4 | 16 | Ar-RC |
| B2. Neophytes | ||||
| Cochlearia danica L. | nc | nc | nc | Ep-21 |
| Duchesnea indica (Andrews) Focke | nc | nc | nc | Ep-21 |
| Senecio inaequidens DC. | nc | nc | nc | Ep-21 |
| Anthoxanthum aristatum Boiss. | nc | 3 | 16 | Ep-AF |
| Oxalis dillenii Jacq. | nc | 2 | 16 | Ep-AF |
| Silene dichotoma Ehrh. | nc | 3 | 16 | Ep-AF |
| Vicia dasycarpa Ten. | nc | 3 | 16 | Ep-AF |
| Vicia grandiflora Scop. | nc | 4 | 16 | Ep-AF |
| Amaranthus blitoides S. Watson | nc | 2 | 16 | Ep-RC |
| Atriplex tatarica L. | nc | 2 | 16 | Ep-RC |
| Iva xanthiifolia Nutt. | nc | 3 | 16 | Ep-RC |
| Lepidium virginicum L. | nc | 3 | 16 | Ep-RC |
| Reseda luteola L. | nc | 3 | 16 | Ep-RC |
| Amaranthus chlorostachys Willd. | nc | 2 | 16 | Ep-AR |
| Amaranthus retroflexus L. | nc | 5 | 16 | Ep-AR |
| Galinsoga ciliata (Raf.) S. F. Blake | nc | 4 | 16 | Ep-AR |
| Galinsoga parviflora Cav. | nc | 5 | 16 | Ep-AR |
| Oxalis corniculata L. | nc | 3 | 16 | Ep-AR |
| Aronia x prunifolia (Marshall) Rehder | nc | 1 | 9 | He-Ag |
| Clematis vitalba L. | nc | 3 | 2 | He-Ag |
| Genistella sagittalis (L.) Gams in Hegi | nc | 2 | 5 | He-Ag |
| Lactuca tatarica (L.) C. A. Mey. | nc | 1 | 8 | He-Ag |
| Lemna turionifera Landolt | nc | 2 | 11 | He-Ag |
| Onobrychis viciifolia Scop. | nc | 3 | 4 | He-Ag |
| Quercus rubra L. | nc | 4 | 3 | He-Ag |
| Rumex longifolius DC. | nc | 1 | 7 | He-Ag |
| Solidago canadensis L. | nc | 4 | 15 | He-Ag |
| Veronica filiformis Sm. | nc | 3 | 6 | He-Ag |
| Xanthium albinum (Widder) H. Scholz | nc | 3 | 10 | He-Ag |
| Acer negundo L. | nc | 4 | 1 | Ho-Ag |
| Acorus calamus L. | nc | 5 | 7 | Ho-Ag |
| Bidens frondosa L. | nc | 3 | 10 | Ho-Ag |
| Elodea canadensis Michx. | nc | 5 | 11 | Ho-Ag |
| Impatiens parviflora DC. | nc | 4 | 2 | Ho-Ag |
| Mimulus moschatus Douglas ex Lindl. | nc | 2 | 12 | Ho-Ag |
| Padus serotina (Ehrh.) Borkh. | nc | 4 | 3 | Ho-Ag |
| Spiraea tomentosa L. | nc | 2 | 6 | Ho-Ag |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jackowiak, B. Man-Made Changes in the Flora and Vegetation of Poland: Current Review. Diversity 2023, 15, 618. https://doi.org/10.3390/d15050618
Jackowiak B. Man-Made Changes in the Flora and Vegetation of Poland: Current Review. Diversity. 2023; 15(5):618. https://doi.org/10.3390/d15050618
Chicago/Turabian StyleJackowiak, Bogdan. 2023. "Man-Made Changes in the Flora and Vegetation of Poland: Current Review" Diversity 15, no. 5: 618. https://doi.org/10.3390/d15050618
APA StyleJackowiak, B. (2023). Man-Made Changes in the Flora and Vegetation of Poland: Current Review. Diversity, 15(5), 618. https://doi.org/10.3390/d15050618