

Skin Anatomy, Bone Histology and Taphonomy of a Toarcian (Lower Jurassic) Ichthyosaur (Reptilia: Ichthyopterygia) from Luxembourg, with Implications for Paleobiology

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Geological Setting
3. Materials and Methods
4. Systematic Paleontology
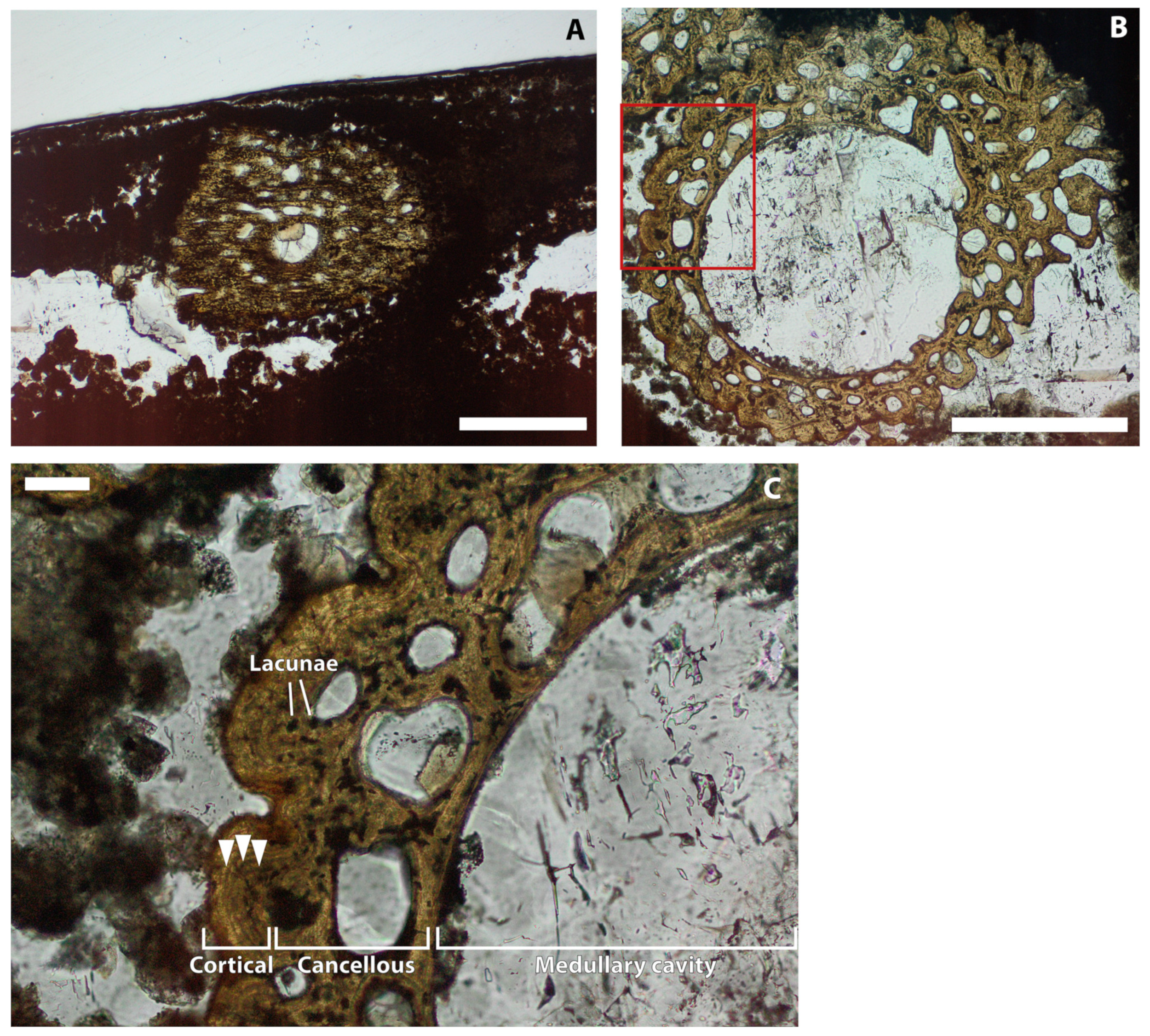
5. Results
5.1. General Description
5.2. Vertebral Column
5.3. Ribs
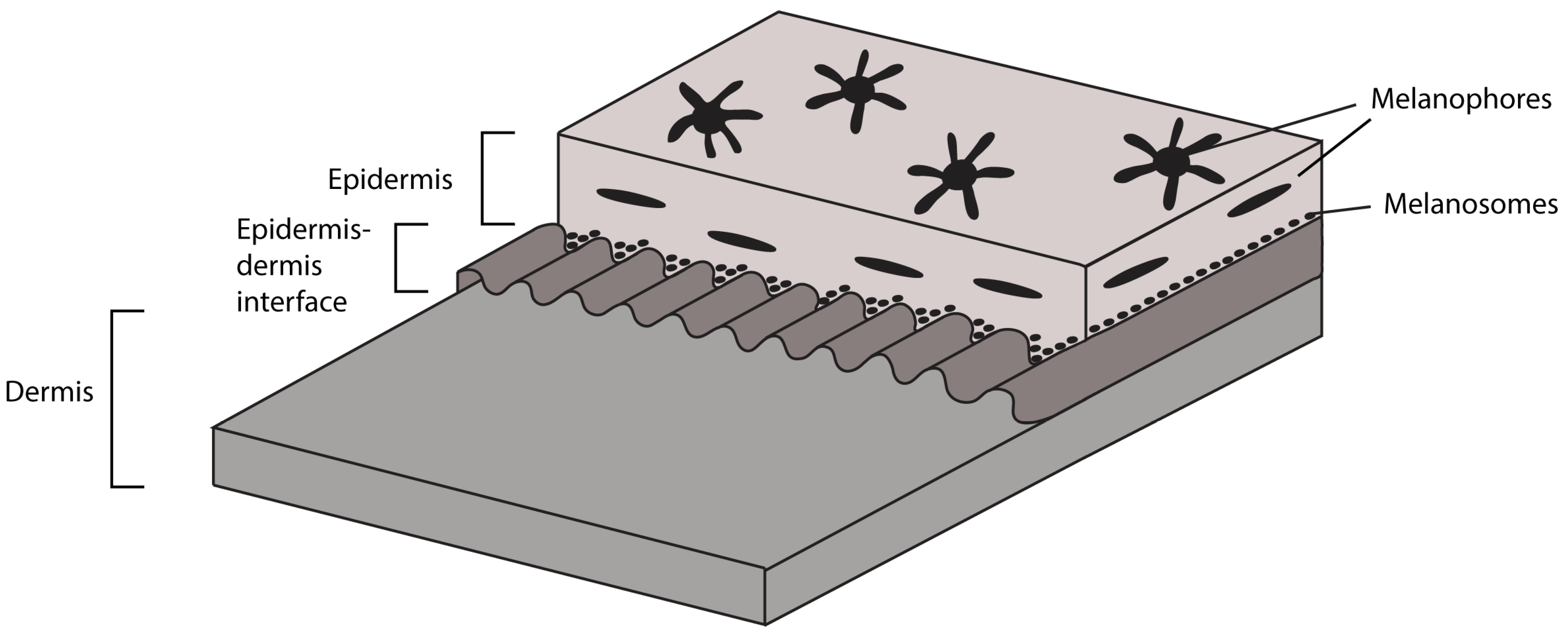
5.4. Soft Tissues
6. Discussion
6.1. Integument of TV344
6.2. Ontogenetic Stage
6.3. Implications of Rib Anatomy
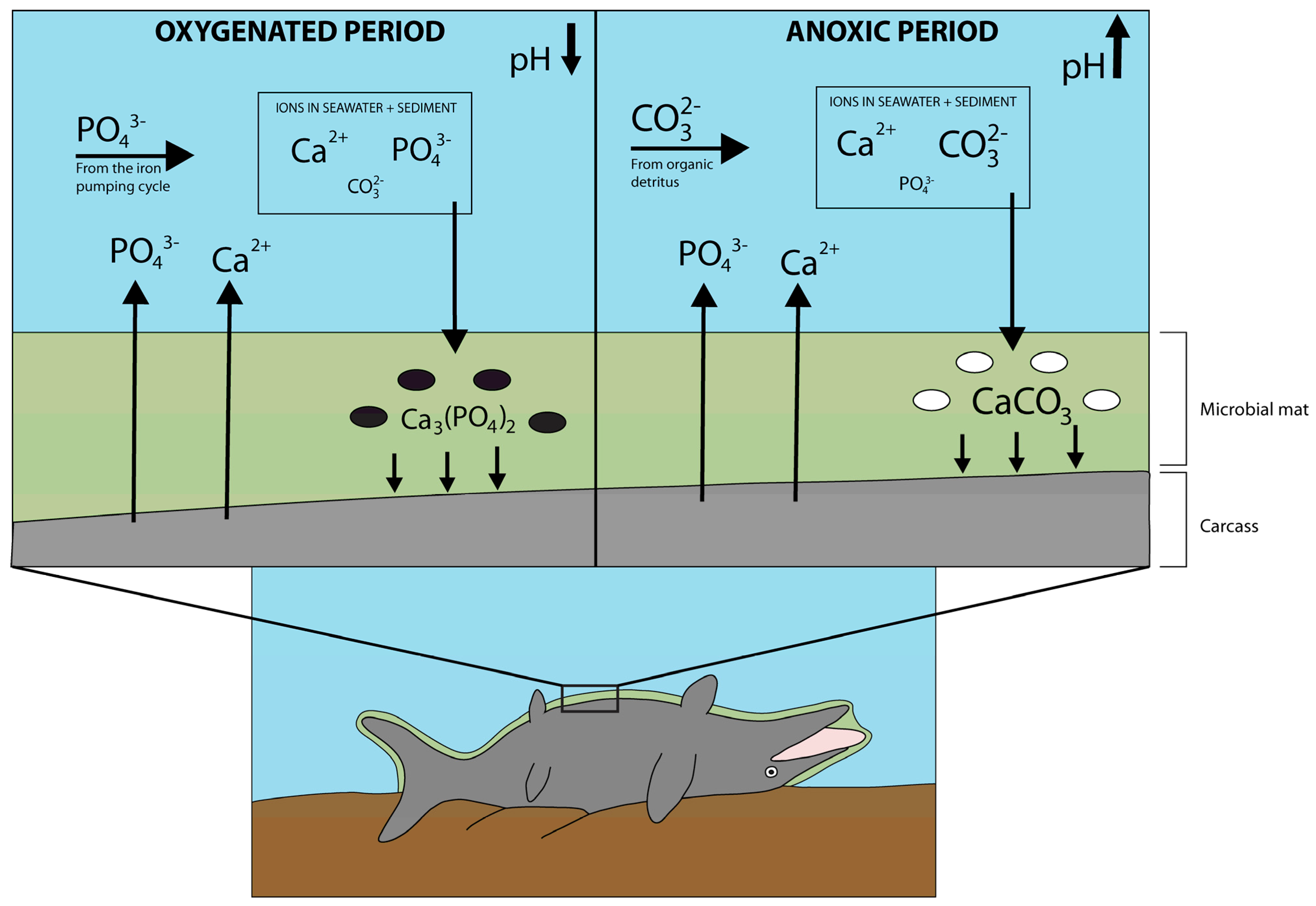
6.4. Taphonomy
7. Conclusions
- TV344 represents an incomplete juvenile individual belonging to the clade Neoichthyosauria, and possibly the genus Stenopterygius;
- The fossil comprises a consecutive series of 54 vertebral centra, associated ribs, 20 bone fragments, as well as extensive soft-tissue remains;
- The soft tissues surrounding the vertebral column are largely integumentary in nature. In addition, there is a distinct layering of the skin, where the epidermis, dermis and epidermis–dermis interface are preserved. Furthermore, the epidermis contains melanophores and melanosomes;
- Concentric ring-like structures observed in sectioned ribs of TV344 may represent annual LAGs, to suggest that the animal was at least three years old at the time of death;
- The taphonomic conditions that enabled the exceptional preservation of TV344 included a combination of seafloor dysoxia/anoxia with periodical oxygenation, thus facilitating rapid phosphatization of the soft tissues during oxygenated periods and the formation of the carbonate nodule during dysoxic/anoxic intervals. Microbial activity aided by lowering the pH during the oxygenated phases, which further enhanced phosphate precipitation.
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A




References
- Lhuyd, E. Lithophylacii Brittannici Ichnographia, Sive Lapidum Aliorumque Fossilium Brittanicorum Singulari Figura Insignium; Gleditsch & Weidmann: London, UK, 1699. [Google Scholar]
- Eriksson, M.E.; De La Garza, R.; Horn, E.; Lindgren, J. A review of ichthyosaur (Reptilia, Ichthyopterygia) soft tissues with implications for life reconstructions. Earth-Sci. Rev 2022, 226, 103965. [Google Scholar] [CrossRef]
- Lindgren, J.; Sjövall, P.; Thiel, V.; Zheng, W.; Ito, S.; Wakamatsu, K.; Hauff, R.; Kear, B.P.; Engdahl, A.; Alwmark, C.; et al. Soft-tissue evidence for homeothermy and crypsis in a Jurassic ichthyosaur. Nature 2018, 564, 359–365. [Google Scholar] [CrossRef] [PubMed]
- Motani, R. Evolution of fish-shaped reptiles (Reptilia: Ichthyopterygia) in their physical environments and constraints. Annu. Rev. Earth Planet. Sci. 2005, 33, 395–420. [Google Scholar] [CrossRef]
- Lingham-Soliar, T.; Plodowski, G. Taphonomic evidence for high-speed adapted fins in thunniform ichthyosaurs. Sci. Nat. 2007, 94, 65–70. [Google Scholar] [CrossRef] [PubMed]
- Lindgren, J.; Sjövall, P.; Carney, R.M.; Uvdal, P.; Gren, J.A.; Dyke, G.; Schultz, B.P.; Shawkey, M.D.; Barnes, K.R.; Polcyn, M.J. Skin pigmentation provides evidence of convergent melanism in extinct marine reptiles. Nature 2014, 506, 484–488. [Google Scholar] [CrossRef] [PubMed]
- Renesto, S.; Dal Sasso, C.; Fogliazza, F.; Ragni, C. New findings reveal that the Middle Triassic ichthyosaur Mixosaurus cornalianus is the oldest amniote with a dorsal fin. Acta Palaeontol. Pol. 2020, 65, 511–522. [Google Scholar] [CrossRef]
- Martill, D.M. An ichthyosaur with preserved soft tissue from the Sinemurian of southern England. Paleontology 1995, 38, 897–903. [Google Scholar]
- Jacobs, M.L.; Martill, D.M. A new ophthalmosaurid ichthyosaur from the Upper Jurassic (Early Tithonian) Kimmeridge Clay of Dorset, UK, with implications for Late Jurassic ichthyosaur diversity. PLoS ONE 2020, 15, e0241700. [Google Scholar] [CrossRef]
- Whitear, M. XCVII.—On the colour of an ichthyosaur. Ann. Mag. Nat. Hist. 1956, 9, 742–744. [Google Scholar] [CrossRef]
- Schintgen, T.; Förster, A. Geology and basin structure of the Trier-Luxembourg Basin—Implications for the existence of a buried Rotliegend graben. ZDGG 2013, 164, 615–637. [Google Scholar] [CrossRef]
- Muscente, A.D.; Vinnes, O.; Sinha, S.; Schiffbauer, J.D.; Maxwell, E.; Schweigert, G.; Martindale, R.C. What role does anoxia play in exceptional fossil preservation? Lessons from the taphonomy of the Posidonia Shale (Germany). Earth-Sci. Rev. 2023, 238, 104323. [Google Scholar] [CrossRef]
- Fantasia, A.; Föllmi, K.B.; Adatte, T.; Spangenberg, J.E.; Montero-Serrano, J.-C. The Early Toarcian oceanic anoxic event: Paleoenvironmental and paleoclimatic change across the Alpine Tethys (Switzerland). Glob. Planet. Chang. 2018, 162, 53–68. [Google Scholar] [CrossRef]
- Ruebsam, W.; Schmid-Röhl, W.; Al-Husseini, M. Astronomical timescale for the early Toarcian (Early Jurassic) Posidonia Shale and global environmental changes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2023, 623, 111619. [Google Scholar] [CrossRef]
- Hermoso, M.; Delsate, D.; Baudin, F.; Le Callonnec, L.; Minoletti, F.; Renard, M.; Faber, A. Record of Early Toarcian carbon cycle perturbations in a nearshore environment: The Bascharage section (easternmost Paris Basin). Solid Earth 2014, 5, 793–804. [Google Scholar] [CrossRef]
- Bechly, G. First record and a new species of the fossil dragonfly genus Proinogomphus (Odonata: Liassogomphidae) from the Early Jurassic of Bascharage in the Grand Duchy of Luxembourg. Zootaxa 2018, 4450, 108–114. [Google Scholar] [CrossRef] [PubMed]
- Heads, S.W.; Thuy, B.; Tamarri, J. Two new species of Archelcana (Sharov (Orthoptera: Elcanidae) from the Lower Jurassic of Luxembourg. Kentiana 2022, 1, 1–13. [Google Scholar] [CrossRef]
- Hanzo, M. A propos de nodules carbonatés du Toarcien inférieur de la région de Bettembourg (Grand-Duché de Luxembourg). In Proceedings of the 103e Congrès National de Sociétés Savants, Nancy-Metz, Nancy, France, 10-15 April 1978. [Google Scholar]
- Hanzo, M. Milieu de dépôt et évolution diagénétique des argilites toarciennes d’après l’etude nodules carbonatés des “Schistescarton” de Bettembourg (Grand-Duché de Luxembourg). Mém. Sci. Terre Nancy 1979, 23, 45–49. [Google Scholar]
- Henrotay, M.; Marques, D.; Paicheler, J.-C.; Gall, J.-C.; Nel, A. Le Toarcien inferieur des régions de Bascharage et de Bettembourg (Grand-Duché du Luxembourg): Évidences paléontologiques et sédimentologiques d’environnements restreints proches de l’émersion. Geodiversitas 1998, 20, 263–284. [Google Scholar]
- Johnson, M.M.; Young, M.T.; Brusatte, S.L.; Thuy, B.; Weis, R. A catalogue of teleosauroids (Crocodylomorpha: Thalattosuchia) from the Toarcian and Bajocian (Jurassic) of southern Luxembourg. Hist. Biol. 2019, 31, 1179–1194. [Google Scholar] [CrossRef]
- Riegraf, W.; Werner, G.; Lörcher, F. Der Posidonienschiefer. Biostratigraphie, Fauna und Fazies des südwestdeutschen Untertoarciums (Lias epsilon); Enke Verlag: Stuttgart, Germany, 1984; 195p. [Google Scholar]
- Frimmel, A. Hochauflösende Untersuchungen von Biomarkern an epikontinentalen Schwarzschiefern des Unteren Toarciums (Posidonienschiefer, Lias ε) von SW-Deutschland. Ph.D. Thesis, Eberhard-Karls-Universität Tübingen, Metzingen, Germany, 2003. [Google Scholar]
- Thibault, N.; Ruhl, M.; Ullmann, C.V.; Korte, C.; Kemp, D.B.; Gröcke, D.R.; Hesselbo, S.P. The wider context of the Lower Jurassic Toarcian oceanic anoxic event in Yorkshire coastal outcrops, UK. Proc. Geol. Assoc. 2018, 129, 372–391. [Google Scholar] [CrossRef]
- Crabb, P. The use of Polarised light in photography of macrofossils. Palaeontology 2001, 44, 659–664. [Google Scholar] [CrossRef]
- Tischlinger, H.; Arratia, G. Ultraviolet light as a tool of investigating Mesozoic fishes with a focus on the ichthyofauna of the Solnhofen Limestone. In Mesozoic Fishes 5. Global Diversity and Evolution, Proceedings of the International Meeting, Saltillo, Mexico, 2010; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2010; pp. 549–560. [Google Scholar]
- de Blainville, H.M.D. Description of some species of reptiles of California, preceded by the analysis of a general system of Herpetology and amphibiology. Ann. Mag. Nat. Hist. 1835, 4, 236–296. [Google Scholar]
- Sander, P.M. Ichthyosauria: Their diversity, distribution, and phylogeny. Palaontol. Z. 2000, 74, 1–35. [Google Scholar] [CrossRef]
- Romer, A.S. Osteology of the Reptiles; University of Chicago Press: Chicago, IL, USA, 1956; p. 772. [Google Scholar]
- Cozzi, B.; Huggenberger, S.; Oelschläger, H. Anatomy of Dolphins: Insights into Body Structure and Function; Academic: Amsterdam, The Netherlands, 2017. [Google Scholar]
- De La Garza, R.; Sjövall, P.; Hauff, R.; Lindgren, J. Preservational modes of some ichthyosaur soft tissues (Reptilia, Ichthyopterygia) from the Jurassic Posidonia Shale of Germany. Palaeontology 2023, 66, e12668. [Google Scholar] [CrossRef]
- Smithwick, F. A Taphonomic and Paleoecological Approach to the Study of Paleocolour. Ph.D. Thesis, University of Bristol, Bristol, UK, 2019. [Google Scholar]
- Delair, J.B. Unusual preservation of fibrous elements in an ichthyosaur skull. Nature 1966, 212, 575–576. [Google Scholar] [CrossRef]
- Lingham-Soliar, T. Rare soft tissue preservation showing fibrous structures in an ichthyosaur from the Lower Lias (Jurassic) of England. Proc. Biol. Sci. 1999, 266, 2367–2373. [Google Scholar] [CrossRef]
- Lingham-Soliar, T. The ichthyosaur integument: Skin fibers, a means for a strong, flexible and smooth skin. Lethaia 2001, 34, 287–302. [Google Scholar] [CrossRef]
- Houssaye, A.; Nakajima, Y.; Sander, P.M. Structural, functional, and physiological signals in ichthyosaur vertebral centrum microanatomy and histology. Geodiversitas 2018, 40, 161–170. [Google Scholar] [CrossRef]
- Kolb, C.; Sánchez-Villagra, M.R.; Scheyer, T.M. The palaeohistology of the basal ichthyosaur Mixosaurus Baur, 1887 (Ichthyopterygia, Mixosauridae) from the Middle Triassic: Palaeobiological implications. Comptes Rendus Palevol 2011, 10, 403–411. [Google Scholar] [CrossRef]
- Seitz, A.L.L. Vergleichende Studien über den Mikroskopischen Knochenbau Fossiler und Rezenter Reptilien und Dessen Bedeutung für das Wachstum und Umbildung des Knochengewebes im Allgemeinen; Druck von E. Karras: Halle, Germany, 1907; p. 172. [Google Scholar]
- Gross, W. Die Typen des mikroskopischen Knochenbaues bei fossilen Stegoccphalen und Reptilien. Z. Anat. Entwickl. 1934, 103, 731–764. [Google Scholar] [CrossRef]
- Anderson, K.L.; Druckenmiller, P.S.; Erickson, G.M.; Maxwell, E.E. Skeletal microstructure of Stenopterygius quadriscissus (Reptilia, Ichthyosauria) from the Posidonienschiefer (Posidonia Shale, Lower Jurassic) of Germany. Palaeontology 2019, 62, 433–449. [Google Scholar] [CrossRef]
- Houssaye, A. Bone histology of aquatic reptiles: What does it tell us about secondary adaptation to an aquatic life? Biol. J. Linn. Soc. 2012, 108, 3–21. [Google Scholar] [CrossRef]
- Horner, J.R.; Padian, K. Age and growth dynamics of Tyrannosaurus rex. Proc. Biol. Sci. 2004, 271, 1875–1880. [Google Scholar] [CrossRef] [PubMed]
- Talevi, M.; Fernandez, M. Unexpected skeletal histology of an ichthyosaur from the Middle Jurassic of Patagonia: Implications for evolution of bone microstructure among secondary aquatic tetrapods. Naturwissenschaften 2012, 99, 241–244. [Google Scholar] [CrossRef] [PubMed]
- Talevi, M.; Fernandez, M.; Salgado, L. Variación Ontogenética en la Histología Ósea de Caypullisaurus Bonapartei Fernández, 1997 (Ichthyosauria: Ophthalmosauridae). Ameghiniana 2012, 49, 38–46. [Google Scholar] [CrossRef]
- Briggs, D.E.G. The Role of Decay and Mineralization in the Preservation of Soft-Bodied Fossils. Annu. Rev. Earth Planet. Sci. 2003, 31, 275–301. [Google Scholar] [CrossRef]
- Briggs, D.E.G.; Wilby, P.R. The role of the calcium carbonate-calcium phosphate switch in the mineralization of soft-bodied fossils. J. Geol. Soc. 1996, 153, 665–668. [Google Scholar] [CrossRef]
- Iniesto, M.; Buscalioni, Á.D.; Carmen Guerrero, M.; Benzerara, K.; Moreira, D.; López-Archilla, A.I. Involvement of microbial mats in early fossilization by decay delay and formation of impressions and replicas of vertebrates and invertebrates. Sci. Rep. 2016, 6, 25716. [Google Scholar] [CrossRef]
- Peterson, J.E.; Lenczewski, M.E.; Scherer, R.P. Influence of microbial biofilms on the preservation of primary soft tissue in fossil and extant archosaurs. PLoS ONE 2010, 5, e13334. [Google Scholar] [CrossRef]
- Brock, J.; Schulz-Vogt, H.N. Sulfide induces phosphate release from polyphosphate in cultures of a marine Beggiatoa strain. ISME J. 2011, 5, 497–506. [Google Scholar] [CrossRef]
- Bomou, B.; Suan, G.; Schlögl, J.; Grosjean, A.-S.; Suchéras-Marx, B.; Adatte, T.; Spangenberg, J.E.; Fouché, S.; Zacaï, A.; Gibert, C.; et al. The paleoenvironmental context of Toarcian vertebrate-yielding shales of southern France (Hérault). Geol. Soc. Spec. Publ. 2021, 514, 121–152. [Google Scholar] [CrossRef]
- Muscente, A.D.; Martindale, R.C.; Schiffbauer, J.D.; Creighton, A.L.; Bogan, B.A. Taphonomy of the Lower Jurassic Konservat-Lagerstätte at Ya Ha Tinda (Alberta, Canada) and its significance for exceptional fossil preservation during oceanic anoxic events. Palaios 2019, 34, 515–541. [Google Scholar] [CrossRef]
- Berner, R.A. Calcium carbonate concretions formed by the decomposition of organic matter. Science 1968, 159, 195–197. [Google Scholar] [CrossRef]
- Briggs, D.E.G.; Kear, A.J. Fossilization of Soft Tissue in the Laboratory. Science 1993, 259, 1439–1442. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonnevier Wallstedt, I.; Sjövall, P.; Thuy, B.; De La Garza, R.G.; Eriksson, M.E.; Lindgren, J. Skin Anatomy, Bone Histology and Taphonomy of a Toarcian (Lower Jurassic) Ichthyosaur (Reptilia: Ichthyopterygia) from Luxembourg, with Implications for Paleobiology. Diversity 2024, 16, 492. https://doi.org/10.3390/d16080492
Bonnevier Wallstedt I, Sjövall P, Thuy B, De La Garza RG, Eriksson ME, Lindgren J. Skin Anatomy, Bone Histology and Taphonomy of a Toarcian (Lower Jurassic) Ichthyosaur (Reptilia: Ichthyopterygia) from Luxembourg, with Implications for Paleobiology. Diversity. 2024; 16(8):492. https://doi.org/10.3390/d16080492
Chicago/Turabian StyleBonnevier Wallstedt, Ida, Peter Sjövall, Ben Thuy, Randolph G. De La Garza, Mats E. Eriksson, and Johan Lindgren. 2024. "Skin Anatomy, Bone Histology and Taphonomy of a Toarcian (Lower Jurassic) Ichthyosaur (Reptilia: Ichthyopterygia) from Luxembourg, with Implications for Paleobiology" Diversity 16, no. 8: 492. https://doi.org/10.3390/d16080492
APA StyleBonnevier Wallstedt, I., Sjövall, P., Thuy, B., De La Garza, R. G., Eriksson, M. E., & Lindgren, J. (2024). Skin Anatomy, Bone Histology and Taphonomy of a Toarcian (Lower Jurassic) Ichthyosaur (Reptilia: Ichthyopterygia) from Luxembourg, with Implications for Paleobiology. Diversity, 16(8), 492. https://doi.org/10.3390/d16080492