

Bone Connectivity and the Evolution of Ichthyosaur Fins


Abstract
1. Introduction
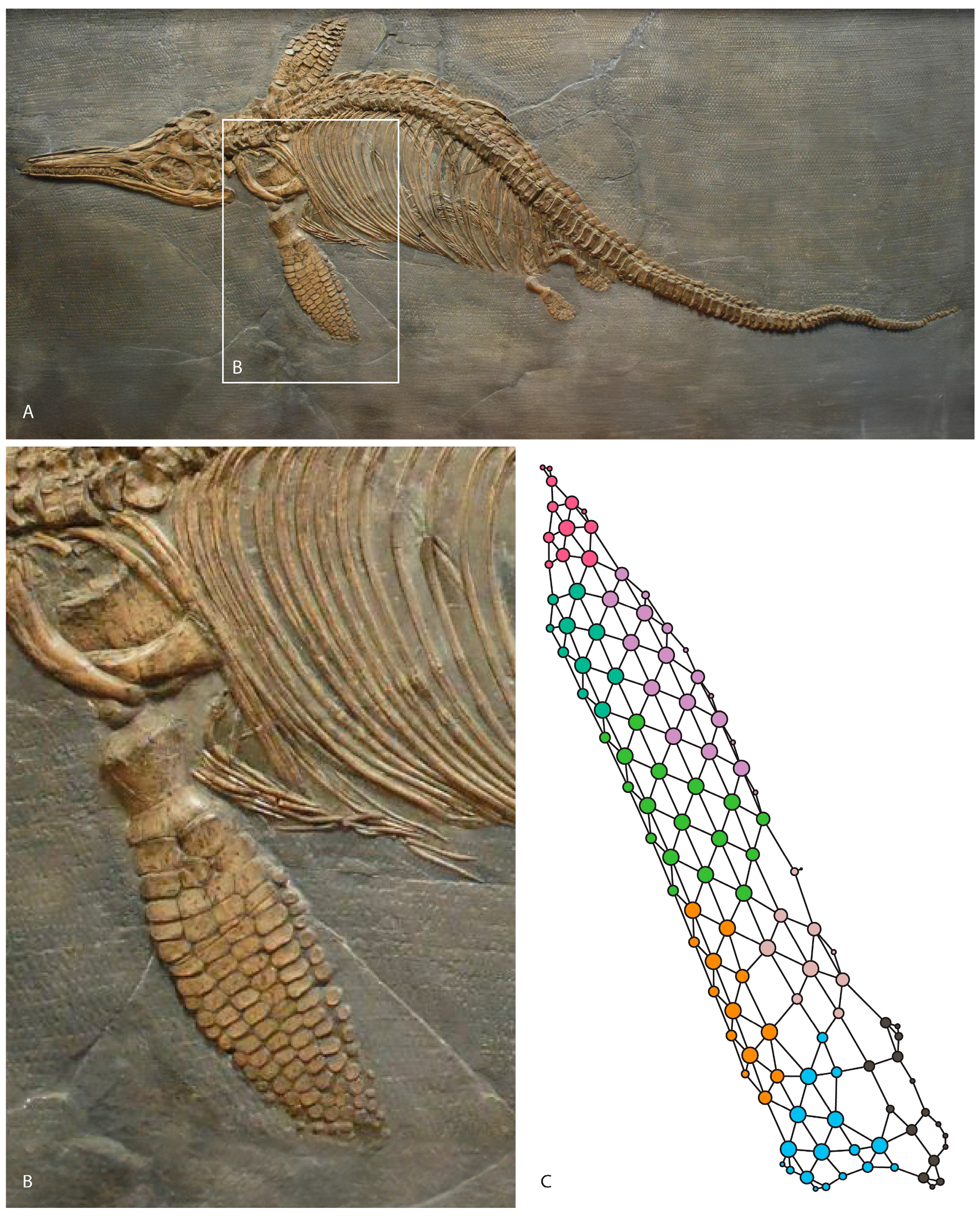
2. Materials and Methods
3. Results
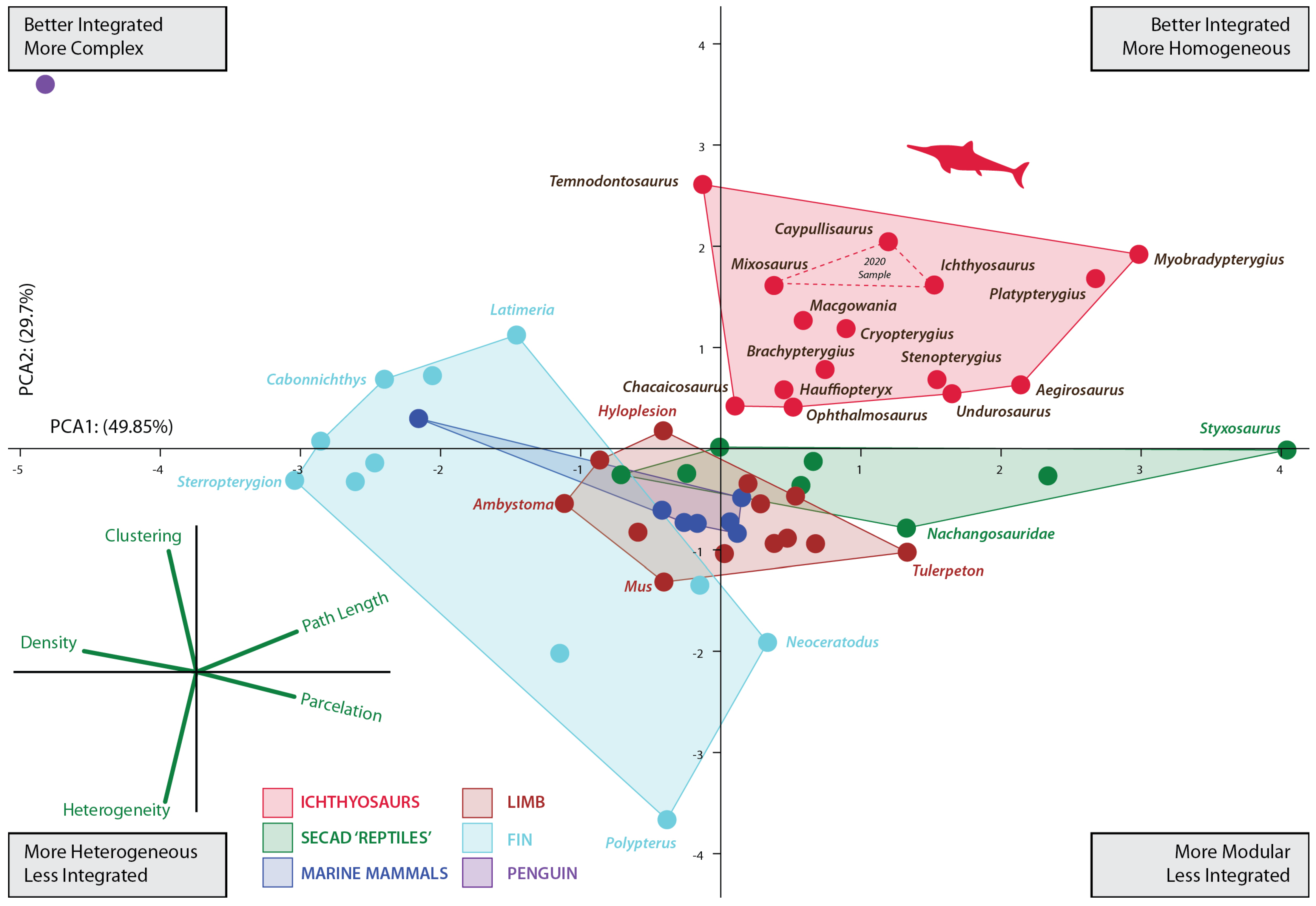
3.1. Morphospace Analyses
3.2. Connectivity Changes in the Forefins across Phylogeny
4. Discussion
4.1. Morphospace Occupation
4.2. Fin Connectivity and Functional Disparity
4.3. Stepwise Evolution of Ichthyosaur Hydrofoils
5. Conclusions and Future Directions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Motani, R.; Jiang, D.; Tintori, A.; Ji, C.; Huang, J. Pre- versus Post-Mass Extinction Divergence of Mesozoic Marine Reptiles Dictated by Time-Scale Dependence of Evolutionary Rates. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170241. [Google Scholar] [CrossRef] [PubMed]
- Moon, B.C.; Stubbs, T.L. Early High Rates and Disparity in the Evolution of Ichthyosaurs. Commun. Biol. 2020, 3, 68. [Google Scholar] [CrossRef]
- Kelley, N.P.; Pyenson, N.D. Evolutionary Innovation and Ecology in Marine Tetrapods from the Triassic to the Anthropocene. Science 2015, 348, aaa3716. [Google Scholar] [CrossRef] [PubMed]
- Renesto, S.; Dal Sasso, C.; Fogliazza, F.; Ragni, C. New Findings Reveal That the Middle Triassic Ichthyosaur Mixosaurus cornalianus Is the Oldest Amniote with a Dorsal Fin. Acta Palaeontol. Pol. 2020, 65, 511–522. [Google Scholar] [CrossRef]
- Massare, J.A. Swimming Capabilities of Mesozoic Marine Reptiles: Implications for Method of Predation. Paleobiology 1988, 14, 187–205. [Google Scholar] [CrossRef]
- Massare, J.A. Faunas, Behavior, and Evolution. In Ancient Marine Reptiles, 1st ed.; Callaway, J.M., Nicholls, E.L., Eds.; Elsevier: Amsterdam, The Netherlands, 1997; pp. 401–421. ISBN 978-0-12-155210-7. [Google Scholar] [CrossRef]
- Webb, P.W. Body Form, Locomotion and Foraging in Aquatic Vertebrates. Am. Zool. 1984, 24, 107–120. [Google Scholar] [CrossRef]
- Webb, P.W. Simple Physical Principles and Vertebrate Aquatic Locomotion. Am. Zool. 1988, 28, 709–725. [Google Scholar] [CrossRef]
- Motani, R. Scaling Effects in Caudal Fin Propulsion and the Speed of Ichthyosaurs. Nature 2002, 415, 309–312. [Google Scholar] [CrossRef]
- Motani, R.; Shimada, K. Skeletal Convergence in Thunniform Sharks, Ichthyosaurs, Whales, and Tunas, and Its Possible Ecological Links through the Marine Ecosystem Evolution. Sci. Rep. 2023, 13, 16664. [Google Scholar] [CrossRef]
- Caldwell, M.W. Modified Perichondral Ossification and the Evolution of Paddle-Like Limbs in Ichthyosaurs and Plesiosaurs. J. Vertebr. Paleontol. 1997, 17, 534–547. [Google Scholar] [CrossRef]
- Caldwell, M.W. From Fins to Limbs to Fins: Limb Evolution in Fossil Marine Reptiles. Am. J. Med. Genet. 2002, 112, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, E.E. Unraveling the Influences of Soft-Tissue Flipper Development on Skeletal Variation Using an Extinct Taxon. J. Exp. Zoolog. B Mol. Dev. Evol. 2012, 318, 545–554. [Google Scholar] [CrossRef]
- DeBlois, M.C.; Motani, R. Flipper Bone Distribution Reveals Flexible Trailing Edge in Underwater Flying Marine Tetrapods. J. Morphol. 2019, 280, 908–924. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.S.; Vlachos, E.; Buono, M.R.; Alzugaray, L.; Campos, L.; Sterli, J.; Herrera, Y.; Paolucci, F. Fingers Zipped up or Baby Mittens? Two Main Tetrapod Strategies to Return to the Sea. Biol. Lett. 2020, 16, 20200281. [Google Scholar] [CrossRef]
- Fischer, V.; Bardet, N.; Benson, R.B.J.; Arkhangelsky, M.S.; Friedman, M. Extinction of Fish-Shaped Marine Reptiles Associated with Reduced Evolutionary Rates and Global Environmental Volatility. Nat. Commun. 2016, 7, 10825. [Google Scholar] [CrossRef] [PubMed]
- Gasparini, Z. A New Pliosaur from the Bajocian of the Neuquen Basin, Argentina. Palaeontology 1997, 40, 135–147. [Google Scholar]
- Fernández, M.S.; Talevi, M. Ophthalmosaurian (Ichthyosauria) Records from the Aalenian–Bajocian of Patagonia (Argentina): An Overview. Geol. Mag. 2014, 151, 49–59. [Google Scholar] [CrossRef]
- Miedema, F.; Bastiaans, D.; Scheyer, T.M.; Klug, C.; Maxwell, E.E. A Large New Middle Jurassic Ichthyosaur Shows the Importance of Body Size Evolution in the Origin of the Ophthalmosauria. BMC Ecol. Evol. 2024, 24, 34. [Google Scholar] [CrossRef]
- Campos, L.; Fernández, M.S.; Herrera, Y.; Garrido, A. Morphological Disparity in the Evolution of the Ophthalmosaurid Forefin: New Clues from the Upper Jurassic of Argentina. Pap. Palaeontol. 2021, 7, 1995–2020. [Google Scholar] [CrossRef]
- Lomax, D.R.; Massare, J.A. Two New Species of Ichthyosaurus from the Lowermost Jurassic (Hettangian) of Somerset, England. Pap. Palaeontol. 2017, 3, 1–20. [Google Scholar] [CrossRef]
- Esteve-Altava, B.; Pierce, S.E.; Molnar, J.L.; Johnston, P.; Diogo, R.; Hutchinson, J.R. Evolutionary Parallelisms of Pectoral and Pelvic Network-Anatomy from Fins to Limbs. Sci. Adv. 2019, 5, eaau7459. [Google Scholar] [CrossRef] [PubMed]
- Bastian, M.; Heymann, S.; Jacomy, M. Gephi: An Open Source Software for Exploring and Manipulating Networks. Proc. Int. AAAI Conf. Web Soc. Media 2009, 3, 361–362. [Google Scholar] [CrossRef]
- Campos, L.; Fernández, M.S.; Bosio, V.; Herrera, Y.; Manzo, A. Revalidation of Myobradypterygius hauthali Huene, 1927 and the Phylogenetic Signal within the Ophthalmosaurid (Ichthyosauria) Forefins. Cretac. Res. 2024, 157, 105818. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Morales, M.E. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 2023, 39, 144–153. [Google Scholar] [CrossRef]
- Thorne, P.M.; Ruta, M.; Benton, M.J. Resetting the Evolution of Marine Reptiles at the Triassic-Jurassic Boundary. Proc. Natl. Acad. Sci. USA 2011, 108, 8339–8344. [Google Scholar] [CrossRef] [PubMed]
- Dick, D.G.; Maxwell, E.E. The Evolution and Extinction of the Ichthyosaurs from the Perspective of Quantitative Ecospace Modelling. Biol. Lett. 2015, 11, 20150339. [Google Scholar] [CrossRef]
- Stubbs, T.L.; Benton, M.J. Ecomorphological Diversifications of Mesozoic Marine Reptiles: The Roles of Ecological Opportunity and Extinction. Paleobiology 2016, 42, 547–573. [Google Scholar] [CrossRef]
- Reeves, J.C.; Moon, B.C.; Benton, M.J.; Stubbs, T.L. Evolution of Ecospace Occupancy by Mesozoic Marine Tetrapods. Palaeontology 2021, 64, 31–49. [Google Scholar] [CrossRef]
- Houssaye, A.; Martin Sander, P.; Klein, N. Adaptive Patterns in Aquatic Amniote Bone Microanatomy—More Complex than Previously Thought. Integr. Comp. Biol. 2016, 56, 1349–1369. [Google Scholar] [CrossRef]
- Talevi, M.; Fernández, M.S. Unexpected Skeletal Histology of an Ichthyosaur from the Middle Jurassic of Patagonia: Implications for Evolution of Bone Microstructure among Secondary Aquatic Tetrapods. Naturwissenschaften 2012, 99, 241–244. [Google Scholar] [CrossRef]
- Anderson, P.S.L.; Friedman, M.; Brazeau, M.D.; Rayfield, E.J. Initial Radiation of Jaws Demonstrated Stability despite Faunal and Environmental Change. Nature 2011, 476, 206–209. [Google Scholar] [CrossRef] [PubMed]
- Massare, J.A. Tooth Morphology and Prey Preference of Mesozoic Marine Reptiles. J. Vertebr. Paleontol. 1987, 7, 121–137. [Google Scholar] [CrossRef]
- Kelley, N.P.; Motani, R. Trophic Convergence Drives Morphological Convergence in Marine Tetrapods. Biol. Lett. 2015, 11, 20140709. [Google Scholar] [CrossRef] [PubMed]
- Foffa, D.; Young, M.T.; Stubbs, T.L.; Dexter, K.G.; Brusatte, S.L. The Long-Term Ecology and Evolution of Marine Reptiles in a Jurassic Seaway. Nat. Ecol. Evol. 2018, 2, 1548–1555. [Google Scholar] [CrossRef]
- Delsett, L.L.; Pyenson, N.; Miedema, F.; Hammer, Ø. Is the Hyoid a Constraint on Innovation? A Study in Convergence Driving Feeding in Fish-Shaped Marine Tetrapods. Paleobiology 2023, 49, 684–699. [Google Scholar] [CrossRef]
- Houssaye, A.; De Buffrénil, V. Bone Histology and the Adaptation to Aquatic Life in Tetrapods. In Vertebrate Skeletal Histology and Paleohistology; CRC Press: Boca Raton, FL, USA, 2021; pp. 744–756. ISBN 978-1-351-18959-0. [Google Scholar]
- Houssaye, A.; Fish, F.E. Functional (Secondary) Adaptation to an Aquatic Life in Vertebrates: An Introduction to the Symposium. Integr. Comp. Biol. 2016, 56, 1266–1270. [Google Scholar] [CrossRef] [PubMed]
- Gutarra, S.; Moon, B.C.; Rahman, I.A.; Palmer, C.; Lautenschlager, S.; Brimacombe, A.J.; Benton, M.J. Effects of Body Plan Evolution on the Hydrodynamic Drag and Energy Requirements of Swimming in Ichthyosaurs. Proc. R. Soc. B Biol. Sci. 2019, 286, 20182786. [Google Scholar] [CrossRef] [PubMed]
- Kear, B.P.; Boles, W.E.; Smith, E.T. Unusual Gut Contents in a Cretaceous Ichthyosaur. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, S206–S208. [Google Scholar] [CrossRef]
- Gerstner, C.L. Maneuverability of Four Species of Coral-Reef Fish That Differ in Body and Pectoral-Fin Morphology. Can. J. Zool. 1999, 77, 1102–1110. [Google Scholar] [CrossRef]
- Fish, F.E.; Lauder, G.V. Control Surfaces of Aquatic Vertebrates: Active and Passive Design and Function. J. Exp. Biol. 2017, 220, 4351–4363. [Google Scholar] [CrossRef]
- Fish, F.E. Aquatic locomotion: Environmental constraints that drive convergent evolution. In Convergent Evolution: Animal Form and Function; Springer International Publishing: Cham, Switzerland, 2023; pp. 477–522. [Google Scholar]
- Fish, F.E.; Shannahan, L.D. The role of the pectoral fins in body trim of sharks. J. Fish Biol. 2000, 56, 1062–1073. [Google Scholar] [CrossRef]
- Hoffmann, S.L.; Porter, M.E. Body and pectoral fin kinematics during routine yaw turning in bonnethead sharks (Sphyrna tiburo). Integr. Org. Biol. 2019, 1, obz014. [Google Scholar] [CrossRef] [PubMed]
- Motani, R. Phylogeny of the Ichthyopterygia. J. Vertebr. Paleontol. 1999, 19, 473–496. [Google Scholar] [CrossRef]
- Wade, M. Platypterygius australis, an Australian Cretaceous Ichthyosaur. Lethaia 1984, 17, 99–113. [Google Scholar] [CrossRef]
- Fernández, M.S. Nuevo Material de Caypullisaurus bonapartei Fernández (Reptilia: Ichthyosauridae) Del Jurásico Superior de La Cuenca Neuquina, Argentina. Ameghiniana 1998, 35, 21–24. [Google Scholar]
- Lee, S.; Kim, J.; Park, H.; Jabłoński, P.G.; Choi, H. The Function of the Alula in Avian Flight. Sci. Rep. 2015, 5, 9914. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, E.E.; Kear, B.P. Postcranial Anatomy of Platypterygius americanus (Reptilia: Ichthyosauria) from the Cretaceous of Wyoming. J. Vertebr. Paleontol. 2010, 30, 1059–1068. [Google Scholar] [CrossRef]
- Pardo-Pérez, J.; Frey, E.; Stinnesbeck, W.; Fernández, M.S.; Rivas, L.; Salazar, C.; Leppe, M. An Ichthyosaurian Forefin from the Lower Cretaceous Zapata Formation of Southern Chile: Implications for Morphological Variability within Platypterygius. Palaeobiodivers. Palaeoenviron. 2012, 92, 287–294. [Google Scholar] [CrossRef]
- Stinnesbeck, W.; Frey, E.; Rivas, L.; Perez, J.P.; Cartes, M.L.; Soto, C.S.; Lobos, P.Z. A Lower Cretaceous Ichthyosaur Graveyard in Deep Marine Slope Channel Deposits at Torres Del Paine National Park, Southern Chile. Geol. Soc. Am. Bull. 2014, 126, 1317–1339. [Google Scholar] [CrossRef]
- Vermeij, G.J. Historical Contingency and the Purported Uniqueness of Evolutionary Innovations. Proc. Natl. Acad. Sci. USA 2006, 103, 1804–1809. [Google Scholar] [CrossRef]
- Jablonski, D. Scale and Hierarchy in Macroevolution. Palaeontology 2007, 50, 87–109. [Google Scholar] [CrossRef]
- Motani, R.; Jiang, D.-Y.; Tintori, A.; Rieppel, O.; Chen, G.-B.; You, H. First Evidence of Centralia in Ichthyopterygia Reiterating Bias from Paedomorphic Characters on Marine Reptile Phylogenetic Reconstruction. J. Vertebr. Paleontol. 2015, 35, e948547. [Google Scholar] [CrossRef]
- Fischer, V.; Weis, R.; Thuy, B. Refining the Marine Reptile Turnover at the Early–Middle Jurassic Transition. PeerJ 2021, 9, e10647. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.S. A New Ichthyosaur from the Los Molles Formation (Early Bajocian), Neuquen Basin, Argentina. J. Paleontol. 1999, 73, 677–681. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | Nodes | Edges | Density | Clustering | Path Length | Heterogeneity | Parcellation | Group |
|---|---|---|---|---|---|---|---|---|
| Aegirosaurus leptospondylus | 109 | 199 | 0.034 | 0.141 | 8.079 | 0.253 | 0.854 | ICHTHYOSAURS |
| Brachypterygius extremus | 70 | 139 | 0.058 | 0.262 | 5.761 | 0.337 | 0.825 | |
| Chacaicosaurus cayi | 48 | 84 | 0.074 | 0.227 | 5.213 | 0.381 | 0.795 | |
| Cryopterygius kristiansenae | 77 | 155 | 0.053 | 0.307 | 6.504 | 0.322 | 0.817 | |
| Hauffiopteryx typicus | 59 | 108 | 0.063 | 0.224 | 5.745 | 0.350 | 0.798 | |
| Macgowania janiceps | 65 | 138 | 0.066 | 0.331 | 5.726 | 0.322 | 0.825 | |
| Myobradypterygius hauthali | 198 | 453 | 0.023 | 0.332 | 11.364 | 0.255 | 0.865 | |
| Ophthalmosaurus icenicus | 61 | 113 | 0.062 | 0.207 | 5.198 | 0.346 | 0.816 | |
| Platypterygius hercynicus | 193 | 436 | 0.024 | 0.303 | 9.803 | 0.242 | 0.869 | |
| Stenopterygius quadriscissus | 89 | 163 | 0.042 | 0.167 | 7.484 | 0.279 | 0.823 | |
| Temnodontosaurus trigonodon | 50 | 114 | 0.093 | 0.494 | 5.442 | 0.263 | 0.799 | |
| Undurosaurus nessovi | 97 | 187 | 0.040 | 0.259 | 8.419 | 0.406 | 0.840 | |
| Ichthyosaurus somersetensis | 113 | 252 | 0.040 | 0.420 | 6.965 | 0.334 | 0.871 | |
| Caypullisaurus bonapartei | 103 | 245 | 0.047 | 0.433 | 6.685 | 0.280 | 0.844 | |
| Mixosaurus cornalianus | 78 | 171 | 0.057 | 0.425 | 5.861 | 0.362 | 0.804 | |
| Hupehsuchus | 37 | 49 | 0.074 | 0.230 | 5.041 | 0.446 | 0.873 | REPTILE SECAD |
| Nanchangosaurus | 56 | 76 | 0.049 | 0.160 | 6.097 | 0.495 | 0.884 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernández, M.S.; Campos, L.; Manzo, A.; Vlachos, E. Bone Connectivity and the Evolution of Ichthyosaur Fins. Diversity 2024, 16, 349. https://doi.org/10.3390/d16060349
Fernández MS, Campos L, Manzo A, Vlachos E. Bone Connectivity and the Evolution of Ichthyosaur Fins. Diversity. 2024; 16(6):349. https://doi.org/10.3390/d16060349
Chicago/Turabian StyleFernández, Marta S., Lisandro Campos, Agustina Manzo, and Evangelos Vlachos. 2024. "Bone Connectivity and the Evolution of Ichthyosaur Fins" Diversity 16, no. 6: 349. https://doi.org/10.3390/d16060349
APA StyleFernández, M. S., Campos, L., Manzo, A., & Vlachos, E. (2024). Bone Connectivity and the Evolution of Ichthyosaur Fins. Diversity, 16(6), 349. https://doi.org/10.3390/d16060349