
Callovian Marine Reptiles of European Russia


 and
and
Abstract
1. Introduction
2. Historical Background
3. Geological and Paleogeographic Settings
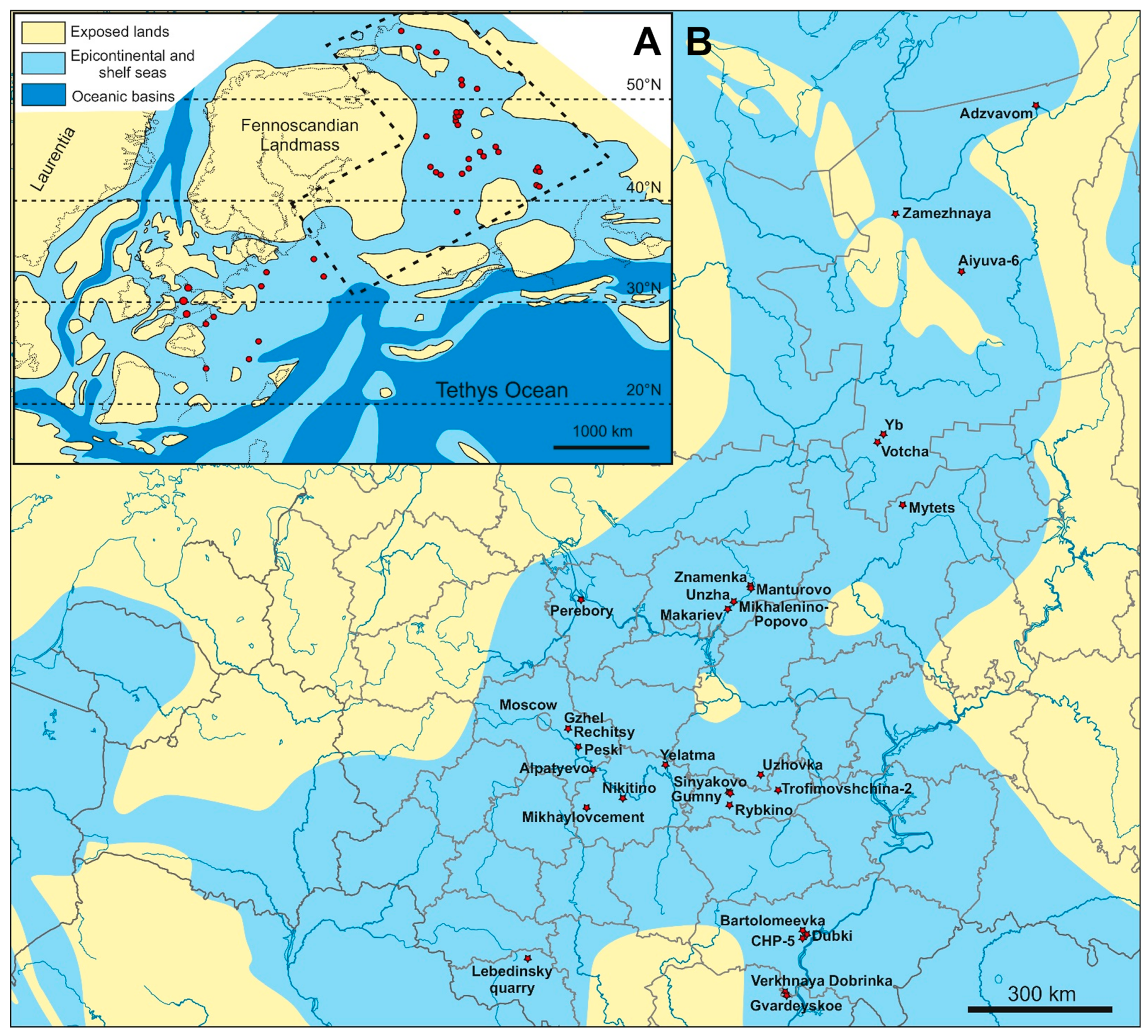
3.1. Paleogeographic Position of the Region
3.2. Localities with Marine Reptiles
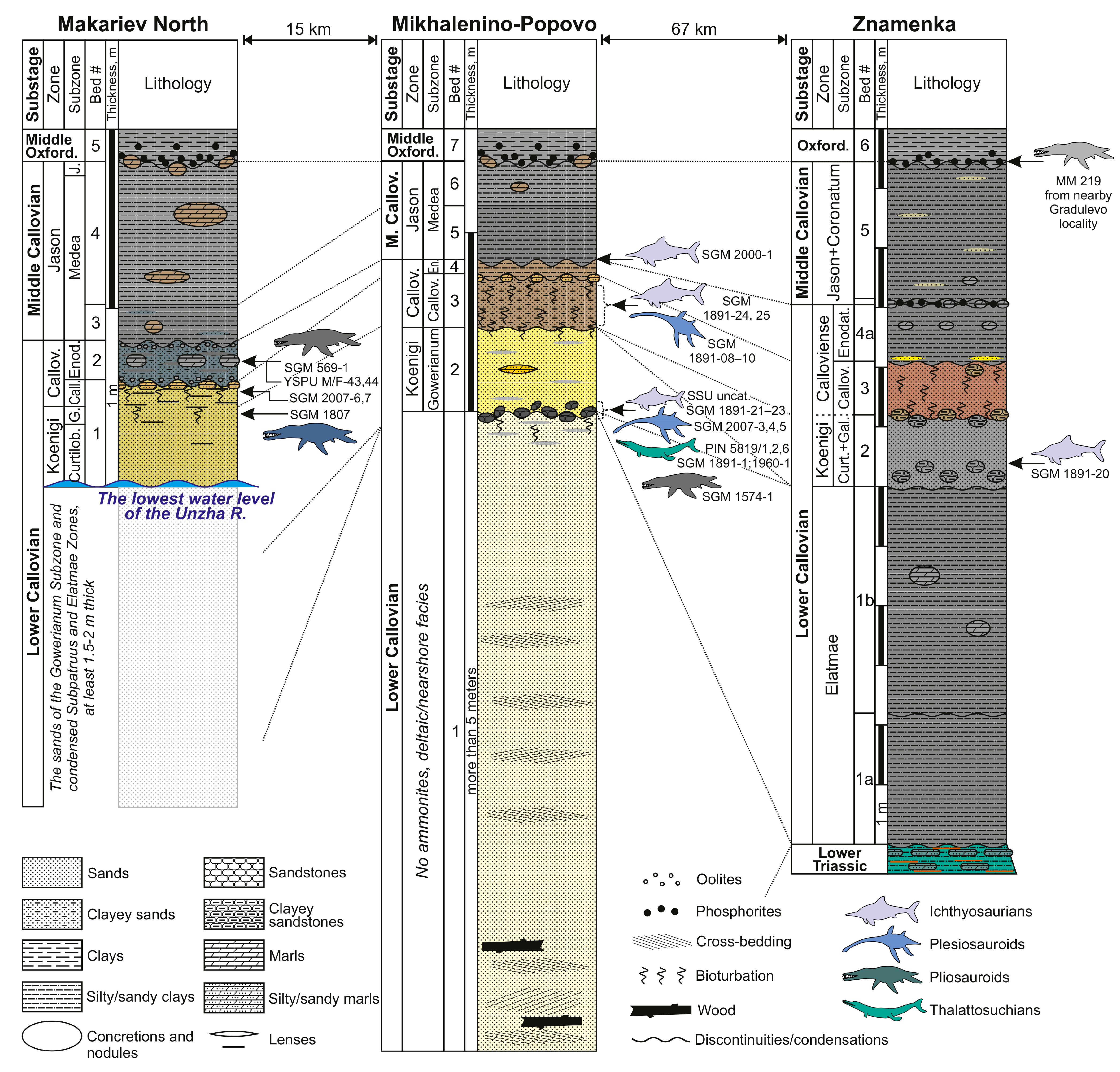
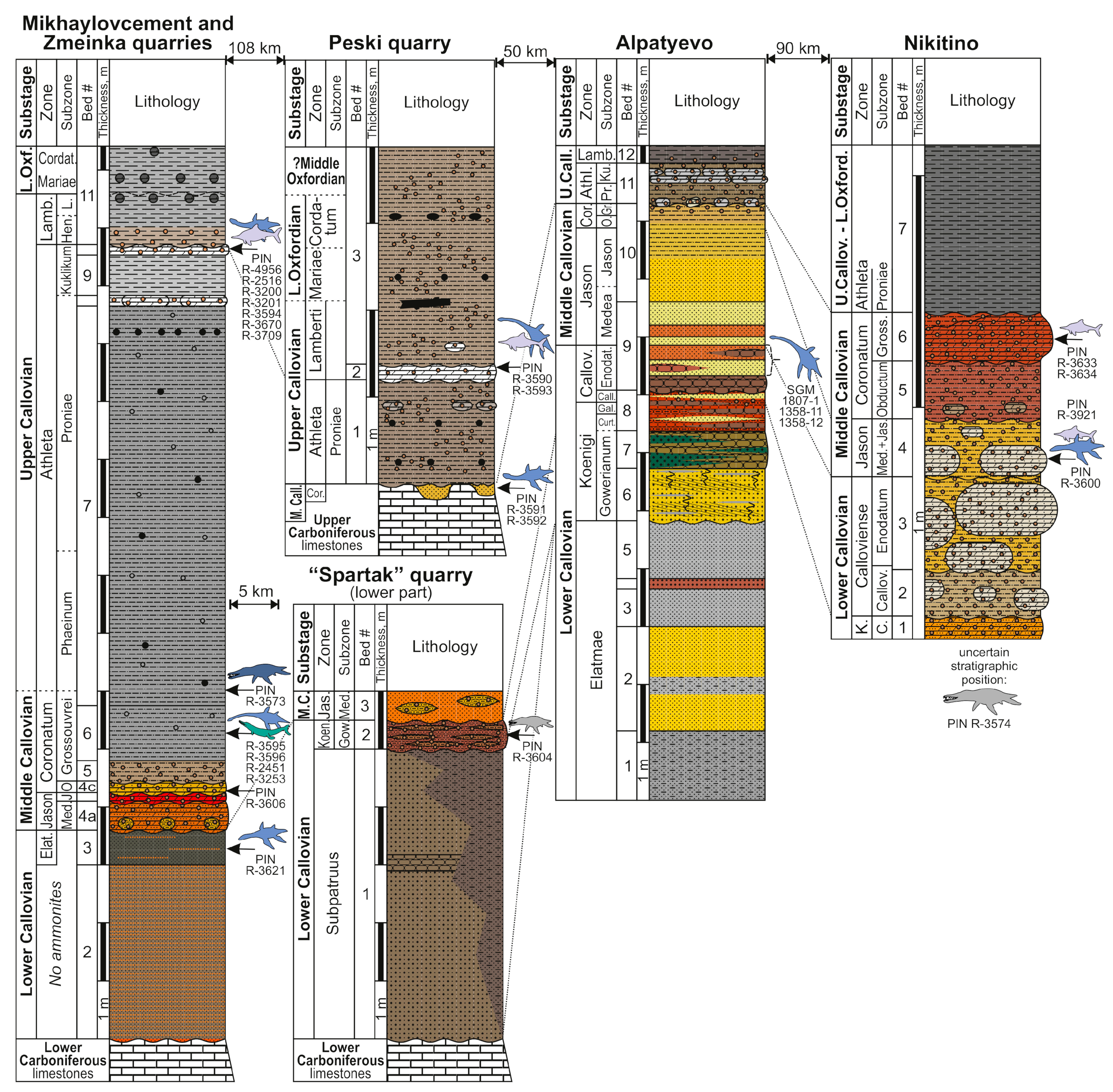
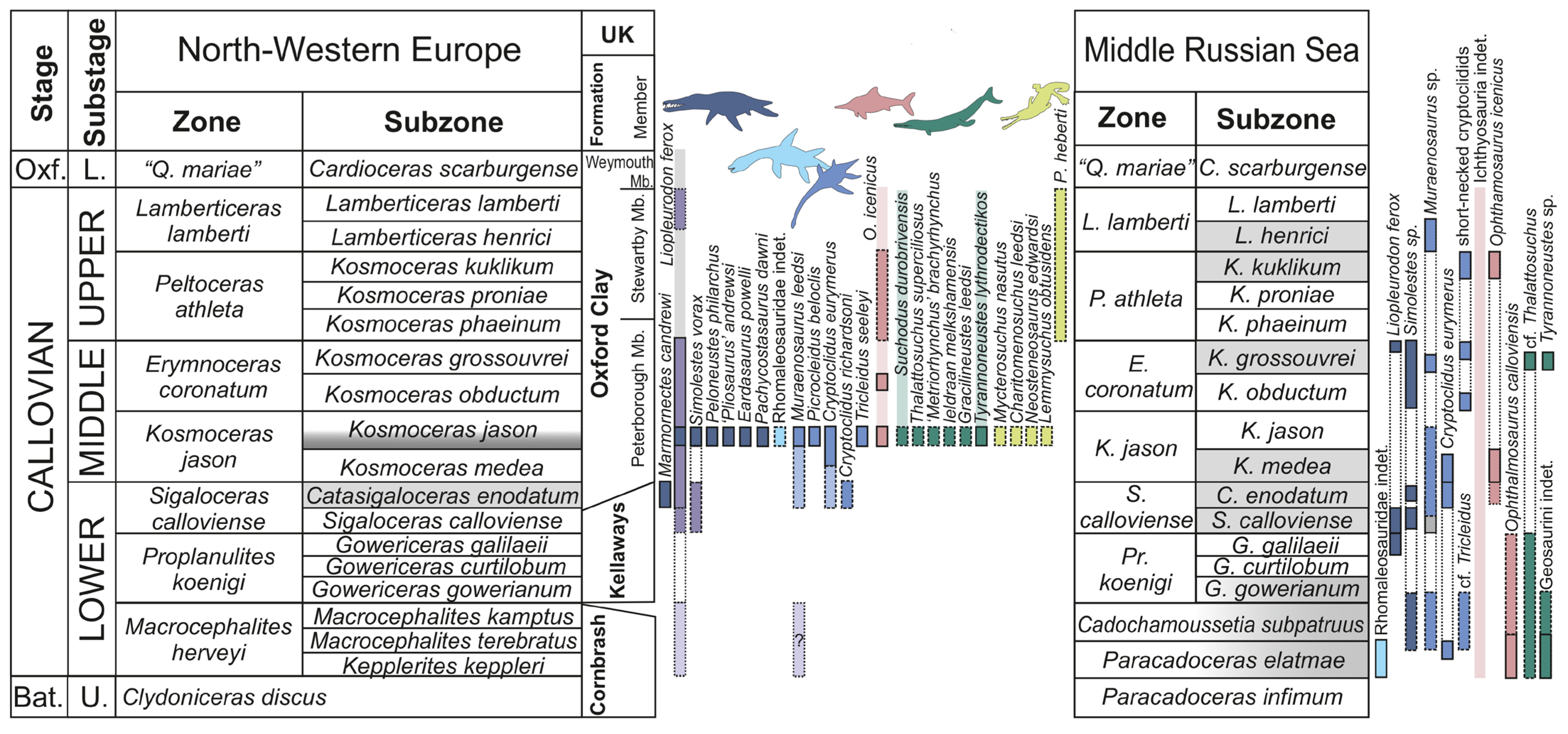
3.3. Geological Features of the Callovian of European Russia and Patterns of Stratigraphical Distributions of Marine Reptile Remains
4. Materials and Methods

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Repository and Catalogue No, Status | Identification (Historical in Parentheses) | Material | Locality | Stratigraphic Position | Reference |
|---|---|---|---|---|---|
| ZIN PH 1/215 | Ichthyosauria indet. | posterior dorsal centrum | Zamezhnaya, “Outcrop 14” (label info; but “outcrop 16” in [109]), Komi Republic | middle/upper Callovian boundary strata, beds with Longaeviceras | collected by V.S. Kravets |
| Specimen location unknown | Ichthyosauria indet. | centrum | Adzvavom, Inta District, Komi Republic, “outcrop 14”in [110] | Upper Callovian, Longaeviceras nikitini Zone (Boreal standard) | [110] |
| UMLH MPZ KP 1982/1 | Ichthyosauria indet. | anterior presacral centrum | Aiyuva-6, Sosnogorsk District, Komi Republic | middle Callovian? | [111] |
| IG 722/6 | Ichthyosauria indet. | posterior caudal centrum | Aiyuva-6 | middle Callovian, Rondiceras milaschevici Zone (Boreal standard) | [111] |
| IG 93/13 | Ichthyosauria indet. | anterior presacral centrum | Votcha, Sysola District, Komi Republic | ex situ, likely from the middle Callovian, K. jason–E. coronatum zones | [112] |
| SGM 1358-53 | Plesiosauria indet. (“Cryptoclidus (?) sp.” in [33]) | caudal vertebra | Kargort (Yb) at Sysola River, Komi Republic | middle Callovian | [33] (p. 230) |
| B.A. Mal’kov’s private collection | Plesiosauria indet., Pliosauridae indet., Ichthyosauria indet., Muraenosaurus sp. | cervical and dorsal vertebrae, plesiosaurian distal propodial | Kargort (Yb) | middle Callovian? | [112,113] |
| SGM uncatalogued | Ichthyosauria indet. | cast of posterior dorsal centrum | Mytets River, Sychugovy Village (abandoned), Nagorsk District, Kirov Region | Callovian? | [114] collected by Khabakov, 1924 |
| NNGASU 147/2080 | Pliosauridae indet. | proximal portion of a large propodial bone | Vyatka–Kama phosphorite field, Verkhnekamsky District, Kirov Region | middle Jurassic, Callovian? | collected by Yu. S. Tamoykin |
| MM 219 | Pliosauridae indet. | partial postcranial skeleton | Gradulevo, Manturovo District, Kostroma Region | middle Callovian or middle Oxfordian | [41] |
| SGM 1891-02, 03 and 20 | cf. Ophthalmosaurus | anterior dorsal vertebra, indet. bone fragments, and partial mandible with teeth. | Znamenka, Manturovo District, Kostroma Region | lower Callovian, Pr. koenigi Zone, G. galilaeii Subzone | collected by V.V. Mitta, 2015 |
| SGM 1807 | Liopleurodon ferox | isolated posterior tooth and partial postcranial skeleton; small plesiosaurian vertebra found in association | Makariev North locality, Makariev District, Kostroma Region | lower Callovian, upper part of Pr. koenigi Zone, G. galilaeii Subzone | collected by A.V. Stupachenko |
| SGM 2007-6,7 | Liopleurodon sp. | three associated teeth | Makariev, Makariev District, Kostroma Region | lower Callovian, S. calloviense Zone | collected by A.V. Stupachenko, 2016 |
| SGM 569-1 | Simolestes sp. | propodial, likely femur | Makariev | lower Callovian, S. calloviense Zone, C. enodatum Subzone | collected by A.V. Stupachenko and V.V. Mitta |
| YSPU M/F-45 | Pliosauridae indet. | articular surface of a large vertebral centrum | Makariev North | lower Callovian, S. calloviense Zone | collected by D.N. Kiselev |
| YSPU M/F-44 | cf. Cryptoclidus | nearly complete dorsal vertebra | Makariev North | same as above | collected by D.N. Kiselev |
| SGM 1574-1 | Simolestes sp. | left maxilla | Mikhalenino, Makariev District, Kostroma Region | lower Callovian, horizon of reworked fauna of P. elatmae and C. subpatruus zones at the base of G. gowerianum Subzone | [37] collected by A.V. Stupachenko |
| SGM 1891-10 | cf. Muraenosaurus | posterior cervical vertebra | Mikhalenino | lower Callovian, S. calloviense Zone | collected by A.V. Stupachenko, 2001 |
| SGM 1891-09 | cf. Muraenosaurus | posterior cervical vertebra | Mikhalenino | same as above | collected by A.V. Stupachenko |
| SGM 1891-08 | Muraenosaurus sp. | anterior cervical vertebra | Mikhalenino | same as above | collected by A.V. Stupachenko, |
| SGM 2007-3(1–5) | Muraenosaurus sp. | five isolated teeth | Mikhalenino | lower Callovian, horizon of reworked fauna of P. elatmae and C. subpatruus zones at the base of G. gowerianum Subzone | collected by A.V. Stupachenko, 1999 |
| PIN 5819/3 | Muraenosaurus sp. | partial tooth | Mikhalenino | same as above | collected by A.V. Stupachenko, 1999 |
| SGM 2007-4 | cf. Tricleidus | tooth crown | Mikhalenino | same as above | collected by A.V. Stupachenko, 1999 |
| SGM 2007-5 | Pliosauridae indet. | fragmant of a large tooth | Mikhalenino | same as above | collected by A.V. Stupachenko, 2016 |
| SGM 2007-8 | cf. Ophthalmosaurus | tooth | Mikhalenino | same as above | collected by A.V. Stupachenko, |
| PIN 5819/4 | cf. Ophthalmosaurus | tooth crown | Mikhalenino | same as above | collected by A.V. Stupachenko, 1999 |
| SGM 1891-22 | cf. Ophthalmosaurus | left jugal | Mikhalenino | same as above | collected by A.V. Stupachenko, 1999 |
| SSU uncatalogued | cf. Ophthalmosaurus | left quadrate | Mikhalenino | same as above | collected by A.V. Stupachenko, 2001 |
| SSU uncatalogued | Ophthalmosauria indet. | right nasal, anterodorsal centrum, rib | Mikhalenino | same as above | collected by A.V. Stupachenko, 1998 |
| SGM 1891-21 | Ophthalmosauria indet. | a series of nine tail fluke centra with neural arches | Mikhalenino | same as above | collected by A.V. Stupachenko, 1999 |
| SGM 1891-23 | Ichthyosauria indet. | long dorsal rib (50 cm proximoditally) | Mikhalenino | same as above | collected by A.V. Stupachenko, 1999 |
| SGM 1960-01 | Geosaurini indet. | tooth crown | Mikhalenino | same as above | [40] (as “SGM BX-12”) collected by A.V. Stupachenko |
| SGM 1891-01 | Thalattosuchia indet. | autopodial element | Mikhalenino | same as above | Collected by A.V. Stupachenko |
| PIN 5819/1, 2, 6 | Metriorhynchidae indet. | three miscellaneous teeth | Mikhalenino | same as above | [40] |
| SGM 1891-25 | Ichthyosauria indet. | posterior presacral centrum | Mikhalenino | lower Callovian, S. calloviense Zone | collected by A.V. Stupachenko, |
| SGM 1891-24 | Ichthyosauria indet. | caudal centrum | Mikhalenino | same as above | collected by A.V. Stupachenko, |
| SGM 2000-1 | Ophthalmosaurus icenicus | left premaxilla | Mikhalenino | clays at the boundary of the lower and middle Callovian | collected by A.V. Stupachenko, 1999 |
| SGM 1961 | Ophthalmosaurus icenicus | skull fragments including quadrate, angular and tooth; atlas-axis and vertebrae from all regions of the column; fragments of pectoral girdle and partial forelimb | Perebory, Rybinsk District, Yaroslavl Region | middle Callovian; K. jason Zone, K. medea Subzone | collected by K.K. Kotov and N.Z., 2021. |
| specimen is lost, holotype | Simolestes sp. (Thaumatosaurus calloviensis) | isolated tooth crown | Rechitsy near Gzhel village, Ramenskoe District, Moscow Region | middle Callovian | [33] (p. 200, pl. II, figs. 1 and 6) |
| PIN R-3589 | cf. Simolestes | partial crown | Gzhel, Ramenskoe District, Moscow Region | middle Callovian | collected by M.S. Boiko, 1990 |
| PIN 5818/8 | Pliosauridae indet. | partial crown | Rechitsy | middle Callovian | collected by P.A. Gerasimov, 1928 |
| PIN 5818/9 | Tyrannoneustes sp. | tooth crown | Rechitsy | middle Callovian, likely E. coronatum Zone | [40] collected by P.A. Gerasimov, 1928 |
| PIN 5819/7 | Metriorhynchidae indet. | tooth crown | Rechitsy | middle to lower upper Callovian | [40] |
| PIN R-3590 | cf. Muraenosaurus | posterior cervical centrum | Peski Quarry, Kolomensk District, Moscow Region | upper Callovian, L. lamberti Zone | [39] (fig. 5b) collected by A. Kuzmenko, 2015 |
| PIN R-3591, PIN R-3592 | Cryptoclididae indet. | cervical of juvenile | Peski Quarry | middle Callovian, E. coronatum Zone | [39] (fig. 5d); collected by N. Denisova and M. Sushko, 2015 |
| PIN R-3593 | Ichthyosauria indet. | four posterior caudal centra | Peski Quarry | upper Callovian, L. lamberti Zone | collected by I.V. Ilyasov, 1990s |
| SGM 1358-11 | Plesiosauria indet. | two articulated pectoral or sacral centra | Gorky (nowadays Fruktovaya Station), Lukhovitsy District, Moscow Region | lower Callovian, likely S. calloviense Zone, C. enodatum Subzone | [33] (p. 149, pl. I) |
| SGM 1358-12 | Plesiosauria indet. | proximal fragment of propodial | Alpatyevo, Lukhovitsy District, Moscow Region | lower Callovian? | [33] (p. 151) |
| SGM 1358-37 | Muraenosaurus cf. leedsi | cervical vertebra | Alpatyevo | lower Callovian, likely S. calloviense Zone, C. enodatum Subzone | [33] (p. 235, pl. III, figs. 1 and 2) |
| PIN R-3600 | Cryptoclidus eurymerus | partial skeleton, including several cervical, pectoral, dorsal and caudal vertebrae, dorsal ribs and gastralia, partial pubis, humerus, radii, femora | Nikitino, Spasskiy District, Ryazan Region | middle Callovian, K. jason Zone | collected by the Club of Junior Paleontologists of the Paleontological Museum, lead by A.S., in 2014–2015 |
| PIN R-3574 | Pliosauridae indet. | partial tooth crown | Nikitino | middle Callovian | collected by D.N. Kasantsev, 2011 |
| PIN R-3633, 3634 | Ichthyosauria indet. | caudal centrum and jawbone fragments | Nikitino | middle Callovian, E. coronatum Zone, K. grossouvrei Subzone | collected by the Club of Junior Paleontologists of the Paleontological Museum, lead by A.S., in 2014-2015 |
| PIN R-3573 | Liopleurodon ferox | large caniniform tooth crown | Mikhaylovcement Quarry, Mikhaylov District, Ryazan Region | same as above | [39] collected by A. Kuraev and A.S., 2012 |
| V. Bakhtin private collection | Simolestes sp. | partial tooth crown | Zmeinka Quarry, Mikhaylov District, Ryazan Region | upper Callovian | [39] (fig. 1C) |
| PIN R-3604 | Pliosauridae indet. | tooth crown fragment | Spartak Quarry, Mikhaylov District, Ryazan Region | lower Callovian, Pr. koenigi Zone | collected by A.S. |
| PIN R-3595 | cf. Muraenosaurus | posterior cervical vertebra | Mikhaylovcement Quarry | middle Callovian, E. coronatum Zone, K. grossouvrei Subzone | [39] (fig. 5c) collected by A. Churkin |
| PIN R-3621 | Cryptoclidus sp. | tooth crown | Mikhaylovcement Quarry | lower Callovian, upper part of P. elatmae Zone | collected by D.G., 2017 |
| PIN R-3670 | Cryptoclididae indet. | cervical vertebra | Mikhaylovcement Quarry | upper Callovian, P. athleta Zone, K. kuklikum Subzone | collected by S.V. Grishin, 2017 |
| PIN R-3709 | Cryptoclididae indet. | pectoral vertebra | Mikhaylovcement Quarry | same as above | collected by Ershova O.G. |
| PIN R-3606 | Cryptoclididae indet. | vertebra | Mikhaylovcement Quarry | middle Callovian, E. coronatum Zone, K. obductum Subzone | collected by Ershova O.G. |
| PIN R-3201, PIN R-3202 | Plesiosauria indet. | caudal centrum and dorsal neural arch fragment | Mikhaylovcement Quarry | upper Callovian, P. athleta Zone, K. kuklikum Subzone | collected by K. Nazarov, 2011 |
| PIN R-3596 | Plesiosauia indet. | posterior caudal centrum | Mikhaylovcement Quarry | middle Callovian, E. coronatum Zone, K. grossouvrei Subzone | collected by S. Rossiysky, 2016 |
| PIN R-4956 | Ophthalmosaurus icenicus | humerus, radius, intermedium, radiale | Zmeinka Quarry | upper Callovian, P. athleta Zone K. kuklikum Subzone | collected by L.V. Kulagina, 2023 |
| PIN R-2516 | cf. Ophthalmosaurus | anterior accessory epipodial element | Mikhaylovcement Quarry | same as above | [39] (fig. 12D) collected by A.S., 2010 |
| PIN R-3200 | Ichthyosauria indet. | posterodorsal centrum | Mikhaylovcement Quarry | same as above | collected by N. Ushakov, 2011 |
| PIN R-3594 | Ichthyosauria indet. | dorsal centrum | Zmeinka Quarry | upper Callovian, L. lamberti Zone | collected by I.A. Dadykin, 2016 |
| PIN R-2451 | Tyrannoneustes sp. | tooth crown | Mikhaylovcement Quarry | middle Callovian, E. coronatum Zone, K. grossouvrei Subzone | [39,40] collected by A.S., 2010 |
| PIN R-3253 | cf. Thalattosuchus | tooth crown | Mikhaylovcement Quarry | same as above | [39,40] collected by A.S. Shmakov, 2012 |
| SGM w/o number | Geosaurinae indet. | cervical vertebra | Mikhaylovcement Quarry | middle to upper Callovian; collected ex situ | collected by K. Volkov |
| TsNIGR 157a/649 | Pliosauridae indet. | sacral vertebra | Yelatma, Kasimov District, Ryazan Region | indet. Callovian | collected by N.A Bogoslovsky |
| TsNIGR 144/1712 | cf. Muraenosaurus | dorsal vertebra | Yelatma | indet. Callovian | collected by E.M. Lutkevich, 1925 |
| SGM 1891-13, 14, 16, 17, 19 | Ichthyosauria indet. | weathered centrum (17), anterior to middle caudal centrum (19); posterior caudal centrum (14); coracoid lateral fragment (13); small rib fragment (16) | Uzhovka Quarry, Pochinki District, Nizhny Novgorod Region | lower Callovian, lower part of Pr. koenigi Zone | collected by D.G., 1995 |
| SGM 1891-18 | Ichthyosauria indet. | small caudal vertebra | Uzhovka Quarry | middle Callovian, K. jason Zone, K. obductum Subzone | collected by D.G., 1995 |
| SGM 1891-15 | Ichthyosauria indet. | posterior dorsal centrum | Uzhovka Quarry | lower Callovian, P. elatmae or C. subpatruus zones | collected by D.G., 1995 |
| SGM 1891-05 | Plesiosauria indet. | dorsal centrum of juvenile | Trofimovshchina-2, Romodanovo District, Republic of Mordovia | lower Callovian, P. elatmae Zone | collected by N.Z., 2018 |
| SGM 1891-06 | Ophthalmosaurus cf. calloviensis | radius | Trofimovshchina-2 | same as above | collected by N.Z., 2018 |
| SGM 1891-04 | Ichthyosauria indet. | middle dorsal centrum | Trofimovshchina-2 | lower Callovian, P. elatmae Zone, upper clayey part | collected by A.P. Ippolitov, 2016 |
| SGM 1891-07 | Ichthyosauria indet. | rib | Trofimovshchina-2 | same as above | collected by A. I., 2016 |
| SGM 1891-11 | Ichthyosauria indet. | weathered vertebral centrum and proximal rib fragments | Trofimovshchina-2 | same as above | collected by A. I. and N.Z., 2018 |
| PSM 3999-4004 | Ophthalmosaurus cf. calloviensis | jaw fragments with teeth; sclerotic ring; dorsal and caudal centra; neural arches; proximal fragment of humerus; radius, ulna and phalanges | Moksha River near Rybkino Village, Kovylkino District, Republic of Mordovia | lower Callovian, P. elatmae to Pr. koenigi zones | collected by A.A. Stuckenberg, 1925 |
| Collection of V.M. Efimov | Ophthalmosaurus sp. | anterior part of the skeleton including skull, ribs and limb elements | Sinyakovo Village, Krasnoslobodsk District, Republic of Mordovia | lower Callovian, P. elatmae Zone | [38] |
| MRUM 1315/1 | cf. Thalattosuchus | tooth crown | Gumny, Krasnoslobodsk District, Republic of Mordovia | lower Callovian, P. elatmae to Pr. koenigi Zones | [40] |
| SGM 1891-27 | Plesiosauria indet. | scapular fragment | Lebedinsky Quarry; Belgorod Region | lower Callovian, P. elatmae Zone | collected by N.Y. Bragin, 2000 |
| SSU 137/211 | cf. Muraenosaurus | cervical vertebra | CHP-5 (TETs-5), Saratov, Saratov Region | lower Callovian, C. subpatruus–S. calloviense zones | collected 1985 |
| SSU 137/200 | Ichthyosauria indet. | caudal centrum | sovkhoz Leninskiy Put’ near CHP 5 power station (TETs-5), Saratov, Saratov Region | lower Callovian, P. elatmae Zone | collected by M.A., 1984 |
| SSU 14/31 (historical No 104a/29) | Geosaurini indet. | tooth crown | sovkhoz Leninskiy Put’ near CHP 5 | lower Callovian, P. elatmae Zone | [35,40] collected by M.A., 1984 |
| SSU 104a/27, holotype | Ophthalmosaurus calloviensis (Khudiakovia calloviensis) | partial forelimb | construction pit near Dubki Village, Saratov District | upper Callovian? (or lower Callovian) | [35] collected 1977 by the Children’s Club of Local History of Saratov |
| PIN R-3575 | Pliosauridae indet. | Crown fragment | Dubki, Saratov District, Saratov Region | upper Callovian, L. lamberti Zone | collected by R. Yu. Stredinin, 2011 |
| SGM 1445-97−120 | Rhomaleosauridae indet. | Ilium, partial ischium and pubis; dorsal and caudal centra; rib, metatarsals and phalanges | Verkhnaya Dobrinka, Zhirnovsk district, Volgograd Region | lower Callovian, P. elatmae Zone | [34,36] collected by E.V. Milanovsky, 1920 |
| SSTU MEZ 3/4 | cf. Ophthalmosaurus | partial skeleton | Gvardeyskoe Village, Krasnoarmeysk District, Saratov Region | lower Callovian, P. elatmae Zone | donated to SSTU by S.A. Bratashova in early 2000s |
| Whereabouts of specimen unknown | Ichthyosauria indet. | Partial skeleton | 0.5 km south of Bartolomeevka, Saratov District, Saratov Region | lower Callovian, P. elatmae Zone | [115] collected by V. Grizbovskiy, 1998 |
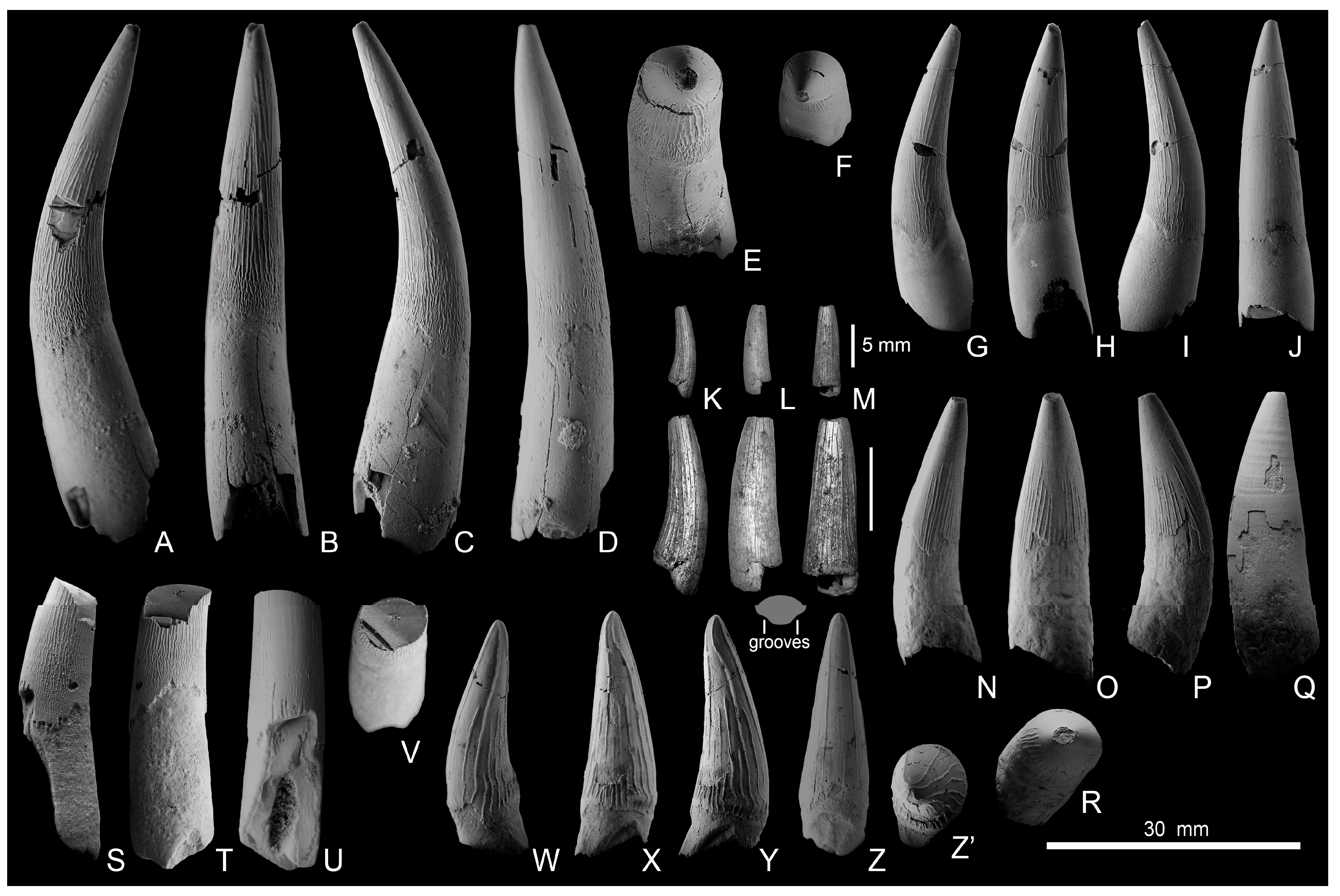
5. Results of the Study of the Marine Reptile Material
5.1. Pliosaurids
5.1.1. Liopleurodon ferox
5.1.2. Simolestes sp.
5.1.3. Pliosauridae gen. et sp. indet.
5.2. Plesiosauroids
5.2.1. Cryptoclidus eurymerus
5.2.2. Isolated Plesiosauroid Teeth
5.2.3. Isolated Plesiosauroid Vertebrae
5.3. Ichthyosaurians
5.3.1. Ophthalmosaurian Ichthyosaurs
5.3.2. Indeterminate Ichthyosaurians
5.4. Thalattosuchian Crocodylomorphs
6. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benson, R.B.J.; Butler, R.J.; Lindgren, J.; Smith, A.S. Mesozoic marine tetrapod diversity: Mass extinctions and temporal heterogeneity in geological megabiases affecting the vertebrates. Proc. R. Soc. Lond. B, Biol. Sci. 2010, 277, 829–834. [Google Scholar] [CrossRef] [PubMed]
- Benson, R.B.J.; Butler, R.J. Uncovering the diversification history of marine tetrapods: Ecology influences the effect of geological sampling biases. Geol. Soc. Spec. Publ. 2011, 358, 191–208. [Google Scholar] [CrossRef]
- Raja, N.B.; Dunne, E.M.; Matiwane, A.; Khan, T.M.; Nätscher, P.S.; Ghilardi, A.M.; Chattopadhyay, D. Colonial history and global economics distort our understanding of deep-time biodiversity. Nat. Ecol. Evol. 2022, 6, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Andrews, C.W. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. Part 1; British Museum (Natural History): London, UK, 1910. [Google Scholar]
- Andrews, C.W. A Descriptive Catalogue of the Marine Reptiles of the Oxford Clay. Part 2; British Museum (Natural History): London, UK, 1913. [Google Scholar]
- Tarlo, L.B. A review of the Upper Jurassic pliosaurs. Bull. Br. Mus. Geol. 1960, 4, 147–189. [Google Scholar]
- Brown, D.S. The English Upper Jurassic Plesiosauroidea (Reptilia) and a review of the phylogeny and classification of the Plesiosauria. Bull. of the Brit. Mus. Geol. 1981, 35, 253–347. [Google Scholar]
- Martill, D.M. The preservation of marine vertebrates in the Lower Oxford Clay (Jurassic) of central England. Philos. Trans. R. Soc. Lond. B 1985, 311, 155–165. [Google Scholar]
- Ketchum, H.F.; Benson, R.B.J. A new pliosaurid (Sauropterygia, Plesiosauria) from the Oxford Clay Formation (Middle Jurassic, Callovian) of England: Evidence for a gracile, longirostrine grade of Early–Middle Jurassic pliosaurids. Spec. Pap. Palaeontol. 2011, 86, 109–129. [Google Scholar]
- Ketchum, H.F.; Benson, R.B.J. The cranial anatomy and taxonomy of Peloneustes philarchus (Sauropterygia, Pliosauridae) from the Peterborough Member (Callovian, Middle Jurassic) of the United Kingdom. Palaeontology 2011, 54, 639–665. [Google Scholar] [CrossRef]
- Ketchum, H.F.; Benson, R.B.J. A new pliosaurid from the Oxford Clay Formation of Oxfordshire, UK. Acta Palaeontol. Pol. 2022, 67, 297–315. [Google Scholar] [CrossRef]
- Sauvage, H.-E. Notes sur les reptiles fossiles. Bull. Soc. Géol. Fr. 1873, 3, 365–386. [Google Scholar]
- Sauvage, H.-E. Synopsis des poisons et des reptiles des terrains jurassiques de Boulogne-sur-Mer. Bull. Soc. Géol. Fr. 1880, 8, 524–547. [Google Scholar]
- Sauvage, H.-E. Catalogue des Reptiles Jurassiques du Boulonnais. Bull Sc Acad de l’Arrondissement de Boulogne-sur-Mer 1914, 10, 253–264. [Google Scholar]
- Bigot, A. Sauroptérygiens du Jurassique du Calvados. Bull. Soc. Géol. Fr. 1938, 5, 631–636. [Google Scholar]
- Brunet, M. Note préliminaire sur une faune de vertébrés du Callovien des environs de Poitiers. C. R. Hebd. Séanc. Acad. Sci. Série D Sci. Nat. 1969, 268, 2667–2670. [Google Scholar]
- Bardet, N.; Pennetier, G.; Pennetier, E. Muraenosaurus leedsii Seeley (Plesiosauria, Elasmosauridae) dans le Callovien du Calvados (France). Neues Jahrb. Geol. Paläontol. Monatshefte 1991, 7, 402–408. [Google Scholar] [CrossRef]
- Bardet, N. Pliosaurs and plesiosaurs from the Middle Jurassic (Callovian) of Normandy. Rev. Paléobiol. 1993, 7, 1–7. [Google Scholar]
- Bardet, N.; Pennetier, G.; Pennetier, E.; Queromain, J. Présence du pliosaure Liopleurodon ferox Sauvage dans le Callovien de Villers-sur-Mer (Normandie). Bull. Trim. Soc. Géol. Normandie Amis Mus. Havre 1993, 80, 11–14. [Google Scholar]
- Bardet, N. Les plésiosaures du Callovien de Normandie. L’Écho Des Falaises 1996, 2, 11–20. [Google Scholar]
- Bardet, N. Les ichthyosaures et les plésiosaures du Jurassique et du Crétacé des falaises des Vaches-Noires (Normandie, France). Fossiles 2013, Hors-Série 4, 98–104. [Google Scholar]
- Hermann, R. Pliosaurus sp. aus dem Ornatenton des fränkischen Jura. Cbl Mineral Geol. Palaontol. 1907, 1907, 667–669. [Google Scholar]
- Von Huene, F. Ein großer Pliosaurus aus dem schwäbischen Ornatenton. Jahresh. Ges. Naturkunde Wurtt. 1934, 90, 31–46. [Google Scholar]
- Michelis, I.; Sander, P.M.; Metzdorf, R.; Breitkreutz, H. Die Vertebratenfauna des Calloviums (Mittlerer Jura) aus dem Steinbruch Störmer (Wallücke, Wiehengebirge). Geol. Paläontol. Westfal. 1996, 44, 1–66. [Google Scholar]
- Sachs, S.; Nyhuis, C.J. Plesiosaurier-Funde aus dem Mittleren Jura von Hildesheim. Naturhistorica 2018, 160, 115–128. [Google Scholar]
- Waskow, K.; Grzegorczyk, D.; Sander, P.M. The first record of Tyrannoneustes (Thalattosuchia: Metriorhynchidae): A complete skull from the Callovian (late Middle Jurassic) of Germany. PalZ 2018, 92, 457–480. [Google Scholar] [CrossRef]
- Gasparini, Z.; Chong-Díaz, G. Metriorhynchus casamiquelai n. sp. (Crocodilia, Thalattosuchia) a marine crocodile from the Jurassic (Callovian) of Chile, South America. Neues Jahrb. Geol. Paläontol. Abh. 1977, 153, 341–360. [Google Scholar]
- Gasparini, Z.B. Un nuevo cocodrilo marino (Crocodylia, Metriorhynchidae) del Caloviano del norte de Chile. Ameghiniana 1980, 17, 97–103. [Google Scholar]
- Gasparini, Z. Marine reptiles from the circum-Pacific region. In The Jurassic of the Circum Pacific; Westermann, G.E.G., Ed.; Cambridge University Press: Cambridge, UK, 1992; pp. 361–364. [Google Scholar]
- Gasparini, Z.; Spaletti, L. First Callovian plesiosaurs from the Neuquén Basin, Argentina. Ameghiniana 1993, 30, 245–254. [Google Scholar]
- Russell, D.A. Jurassic marine reptiles from Cape Grassy, Melville Island, Arctic Canada. Bull. Geol. Surv. Can. 1994, 450, 195–201. [Google Scholar]
- Sato, T.; Wu, X.-C. A new Jurassic pliosaur from Melville Island, Canadian Arctic Archipelago. Can. J. Earth Sci. 2008, 45, 303–320. [Google Scholar] [CrossRef]
- Bogolubov, N.N. On the History of Plesiosaurs in Russia; Tipografia Imperatorskogo Moskovskogo Universiteta: Moscow, Russia, 1911. (In Russian) [Google Scholar]
- Milanovsky, E.V. A find of plesiosaur in the Jurassic deposits of Saratov province. Geol. Vestn. 1921, 4, 118–120. (In Russian) [Google Scholar]
- Arkhangelsky, M.S. On an ichthyosaur from the Callovian stage of the Volga Region near Saratov. Paleontol. J. 1999, 33, 88–91. [Google Scholar]
- Benson, R.B.J.; Zverkov, N.G.; Arkhangelsky, M.S. Youngest occurrences of rhomaleosaurid plesiosaurs indicate survival of an archaic marine reptile clade at high palaeolatitudes. Acta Palaeontol. Pol. 2015, 60, 769–780. [Google Scholar] [CrossRef]
- Zverkov, N.G.; Arkhangelsky, M.S. On a find of a Callovian short-snouted pliosaur in Kostroma Region (Russia). In Jurassic System of Russia: Problems of Stratigraphy and Paleogeography, Proceedings of the Sixth All-Russian Meeting; Zakharov, V.A., Rogov, M.A., Ippolitov, A.P., Eds.; ALEF: Makhachkala, Russia, 2015; pp. 135–138. (In Russian) [Google Scholar]
- Efimov, V.M.; Meleshin, I.A. The first find of an ichthyosaur in the Jurassic deposits of Mordovia. In Proceedings of the all-Russian Scientific Conference “Treshnikovskie Chteniya-2017”, Ulyanovsk, Russia, 30–31 March 2017; pp. 22–23. (In Russian). [Google Scholar]
- Zverkov, N.G.; Shmakov, A.S.; Arkhangelsky, M.S. Jurassic Marine Reptiles of Moscow and Surroundings. In Jurassic Deposits of the Southern Part of the Moscow Syneclyse and Their Fauna; Rogov, M.A., Zakharov, V.A., Eds.; GEOS: Moscow, Russia, 2017; pp. 230–263. (In Russian) [Google Scholar]
- Young, M.T.; Zverkov, N.G.; Arkhangelsky, M.S.; Ippolitov, A.P.; Meleshin, I.A.; Mirantsev, G.V.; Shmakov, A.S.; Stenshin, I.M. Thalattosuchian crocodylomorphs from European Russia, and new insights into metriorhynchid tooth serration evolution and their palaeolatitudinal distribution. PeerJ 2023, 11, e15781. [Google Scholar] [CrossRef]
- Riabinin, A.N. Zwei Plesiosaurier aus den Jura- und Kreideablagerungen Russlands. Mém. Com. Géol. St. Petersburg 1909, 43, 1–49. (In Russian) [Google Scholar]
- Bogolubov, N.N. Sur quelques restes de deux reptiles (Cryptodidus simbirskensis n. sp. et Ichthyosaurus steleodon n. sp.), trouvés par Mr. le Profes. A.P. Pavlow sur les bords de la Volga dans les couches mésozoiques de Simbirsk. Ann. Géol. Minéral. Russ. 1909, 11, 42–64. (In Russian) [Google Scholar]
- Arkhangelsky, M.S.; Sennikov, A.G. Subclass Synaptosauria. In Fossil Vertebrates of Russia and Neighboring Countries. Fossil Reptiles and Birds. Part 1; Ivakhnenko, M.F., Kurochkin, E.N., Eds.; GEOS: Moscow, Russia, 2008; pp. 224–243. (In Russian) [Google Scholar]
- Storrs, G.W.; Arkhangel’sky, M.S.; Efimov, V.M. Mesozoic marine reptiles of Russia and other former Soviet republics. In The Age of Dinosaurs in Russia and Mongolia; Benton, M.J., Shishkin, M.A., Unwin, D.M., Kurochkin, E.N., Eds.; Cambridge University Press: Cambridge, UK, 2000; pp. 187–210. [Google Scholar]
- Knutsen, E.M.; Druckenmiller, P.S.; Hurum, J.H. A new plesiosauroid (Reptilia: Sauropterygia from the Agardhfjellet Formation (middle Volgian) of central Spitsbergen, Norway. Nor. J. Geol. 2012, 92, 213–234. [Google Scholar]
- Smith, A.S. The back-to-front plesiosaur Cryptoclidus (Apractocleidus) aldingeri from the Kimmeridgian of Milne Land, Greenland. Bull. Geol. Soc. Den. 2007, 55, 1–7. [Google Scholar] [CrossRef]
- Riabinin, A.N. Class Reptilia. In Atlas of the Guide Forms of the Fossil Faunas of USSR. Vol. IX. Upper Series of the Jurassic System; Krymgoltz, G.Y., Ed.; Gosgeolizdat: Moscow, Russia, 1949; pp. 290–298. (In Russian) [Google Scholar]
- Zverkov, N.G.; Fischer, V.; Madzia, D.; Benson, R.B.J. Increased pliosaurid dental disparity across the Jurassic-Cretaceous transition. Palaeontology 2018, 61, 825–846. [Google Scholar] [CrossRef]
- Nesov, L.A.; Ivanov, A.O.; Khozatskii, L.I. On the finds of the remains of ichthyosaurs in the USSR and the problem of faunal change in the middle Cretaceous. Vestn. Leningr. Universiteta. Seriya 7 Geol. Geogr. 1988, 1, 15–25. (In Russian) [Google Scholar]
- Maisch, M.W.; Matzke, A.T. The Ichthyosauria. Stutt Beitr. Naturkd B 2000, 298, 1–159. [Google Scholar]
- McGowan, C.; Motani, R. Handbook of Paleoherpetology. Part 8. Ichthyopterygia; Verlag Dr. Friedrich Pfeil: München, Germany, 2003. [Google Scholar]
- Moon, B.C.; Kirton, A.M. Ichthyosaurs of the British Middle and Upper Jurassic Part 1. Palaeontogr. Soc. Monogr. 2016, 170, 1–84. [Google Scholar] [CrossRef]
- Arkhangelsky, M.S. Subclass Ichthyopterygia. In Fossil Vertebrates of Russia and Neighboring Countries. Fossil Reptiles and birds. Part 1; Ivakhnenko, M.F., Kurochkin, E.N., Eds.; GEOS: Moscow, Russia, 2008; pp. 244–262. (In Russian) [Google Scholar]
- Mitta, V.V.; Seltzer, V.B. First finds of Arctocephalitinae (Ammonoidea) in Jurassic of the southeastern Russian Platform and correlation of the boreal Bathonian with the standard scale. Trans. Sci. Res. Geol. Inst. Saratov State Univ. 2002, 10, 12–39. (In Russian) [Google Scholar]
- Seltzer, V.B. Boreal event in the Early Bathonian basin of the southeastern part of the Russian Plate. In Proceedings of the LV session of the Paleontological Society of RAS “Paleontology and Development of the Stratigraphic Basis for Geologic Mapping”, St. Petersburg, Russia, 6–10 April 2009. (In Russian). [Google Scholar]
- Mitta, V.V.; Zakharov, V.A.; Barskov, I.S.; Sel’tser, V.B.; Ivanov, A.V. The Upper Bajocian–Lower Bathonian boundary section in the outskirts of Saratov: Molluscan characteristics and biostratigraphy. Stratigr. Geol. Correl. 2011, 19, 502–514. [Google Scholar] [CrossRef]
- Mitta, V.; Kostyleva, V.; Dzyuba, O.; Glinskikh, L.; Shurygin, B.; Seltzer, V.; Ivanov, A.; Urman, O. Biostratigraphy and sedimentary settings of the Upper Bajocian–Lower Bathonian in the vicinity of Saratov (Central Russia). Neues Jahrb. Geol. Paläontol. Abh. 2014, 271, 95–121. [Google Scholar] [CrossRef]
- Ippolitov, A.P. Belemnites and biostratigraphy of the Lower Bathonian of central and southern parts of East European platform. Part 2. Cylindroteuthididae and Belemnotheutididae. Stratigr. Geol. Correl. 2018, 26, 433–458. [Google Scholar] [CrossRef]
- Gulyaev, D.B. Ammonites and Infrazonal Stratigraphy of the Lower Bathonian Besnosovi Zone of the Russian Plate. Stratigr. Geol. Correl. 2019, 27, 95–117. [Google Scholar] [CrossRef]
- Ippolitov, A.P.; Gulyaev, D.B. Palaeogeography and marine connections of the Middle Russian Sea across the Middle Jurassic: Cephalopod-based model. In Proceedings of the 10th International Symposium “Cephalopods—Present and Past”, Fes, Morocco, 26 March–3 April 2018; pp. 53–54. [Google Scholar]
- Gulyaev, D.B. Infrazonal Ammonite Scale for the Upper Bathonian–Lower Callovian of Central Russia. Stratigr. Geol. Correl. 2001, 9, 65–92. [Google Scholar]
- Gulyaev, D.B. New data on ammonite biostratigraphy of the Upper Batonian and Lower Callovian deposits of Churkinskaya Stchelya reference stratigraphic section (Pizhma River, river basin of Pechora). In Jurassic System of Russia: Problems of Stratigraphy and Paleogeography. Second All-Russian Meeting: Scientific Materials; Zakharov, V.A., Dzyuba, O.S., Kiselev, D.N., Rogov, M.A., Eds.; Yaroslavl State Pedagogical University: Yaroslavl, Russia, 2007; pp. 49–58. (In Russian) [Google Scholar]
- Kiselev, D.N.; Rogov, M.A. Stratigraphy of the Bathonian–Callovian boundary deposits in the Prosek secion (Middle Volga Region), Article 1. Ammonites and Infrazonal biostratigraphy. Stratigr. Geol. Correl. 2007, 15, 485–515. [Google Scholar] [CrossRef]
- Gulyaev, D.B.; Rogov, M.A. Macrocephalitids (Sphaeroceratidae, Ammonoidea) in the Early Callovian of the East-European subboreal sea. In Contributions to current cephalopod research: Morphology, Systematics, Evolution, Ecology and Biostratigraphy, Issue 2; Leonova, T.B., Barskov, I.S., Mitta, V.V., Eds.; PIN RAS: Moscow, Russia, 2009; pp. 72–74. (In Russian) [Google Scholar]
- Gulyaev, D.B.; Ippolitov, A.P. Lower Callovian of Kanev dislocations, Cherkasy oblast, Ukraine: Ammonites and stratigraphy. Stratigr. Geol. Correl. 2021, 29, 767–847. [Google Scholar] [CrossRef]
- Mönnig, E. Der Macrocephalen Oolith von Hildesheim; Georg Olms Verlag: Hildesheim, Germany, 1995; pp. 1–77. [Google Scholar]
- Mönnig, E.; Dietl, G. The systematics of the ammonite genus Kepplerites (upper Bathonian and basal Callovian, Middle Jurassic) and the proposed basal boundary stratotype (GSSP) of the Callovian Stage. Neues Jahrb. Geol. Paläontol. Abh. 2017, 286, 235–287. [Google Scholar] [CrossRef]
- Mitta, V.V.; Dietl, G.; Callomon, J.H.; Schweigert, G.; Dietze, V. The ammonite genus Cadoceras (Cardioceratidae) in the Lower Callovian (Middle Jurassic) of the Swabian Alb and the Wutach area (Southern Germany), Neues Jahrb. Geol. Paläontol. Abh. 2015, 278, 303–321. [Google Scholar] [CrossRef]
- Sasonova, I.G.; Sasonov, N.T. Paleogeography of the Russian Platform during Jurassic and Early Cretaceous Time; Nedra: Leningrad, Russia, 1967. (In Russian) [Google Scholar]
- Ippolitov, A.P. The history of Middle Jurassic belemnites in the Middle Russian Sea, in the light of recent discoveries. In Contributions to Current Cephalopod Research: Morphology, Systematics, Evolution, Ecology and Biostratigraphy. Vol. 5, Proceeding of Conference, Moscow, 29–31 October 2018; Leonova, T.B., Barskov, I.S., Mitta, V.V., Eds.; PIN RAS: Moscow, Russia, 2018; pp. 49–52. (In Russian) [Google Scholar]
- Vinogradov, A.P. (Ed.) Atlas of the Lithological-Paleogeographic Maps of the Russian Platform and its Geosynclinal Margins. Volume II. Mesozoic and Cenozoic; Gosudarstvennoe Nauchno-Tehnicheskoe Izdatelstvo Literatury po Geologii i Ohrane nedr: Moscow–Leningrad, Russia, 1961. (In Russian) [Google Scholar]
- Wierzbowski, H.; Rogov, M. Reconstructing the palaeoenvironment of the Middle Russian Sea during the Middle–Late Jurassic transition using stable isotope ratios of cephalopod shells and variations in faunal assemblages. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 299, 250–264. [Google Scholar] [CrossRef]
- Benton, M.J.; Spencer, P.S. British Mid Jurassic fossil reptile sites. In Fossil Reptiles of Great Britain; Benton, M.J., Spencer, P.S., Eds.; Chapman and Hall: New York, NY, USA, 1995; pp. 124–164. [Google Scholar]
- Riou, B.; Bennourine, M.; Klee, N.; Lutz, M. Jurassic marine crocodiles in the Monts d’Ardèche UNESCO Global Geopark. Geoconservation Res. 2021, 4, 368–377. [Google Scholar]
- Parrilla-Bel, J.; Young, M.T.; Moreno-Azanza, M.; Canudo, J.I. The first metriorhynchid crocodylomorph from the Middle Jurassic of Spain, with implications for evolution of the subclade Rhacheosaurini. PLoS ONE 2013, 8, e54275. [Google Scholar] [CrossRef] [PubMed]
- Madzia, D.; Sachs, S.; Klug, C. Historical significance and taxonomic status of Ischyrodon meriani (Pliosauridae) from the Middle Jurassic of Switzerland. PeerJ 2022, 10, e13244. [Google Scholar] [CrossRef] [PubMed]
- Madzia, D.; Szczygielski, T.; Wolniewicz, A.S. The giant pliosaurid that wasn’t-revising the marine reptiles from the Kimmeridgian, Upper Jurassic, of Krzyżanowice, Poland. Acta Palaeontol. Pol. 2021, 66, 99–129. [Google Scholar] [CrossRef]
- Young, M.T.; Andrade, M.B.; Brusatte, S.L.; Sakamoto, M.; Liston, J. The oldest known metriorhynchid super-predator: A new genus and species from the Middle Jurassic of England, with implications for serration and mandibular evolution in predacious clades. J. Syst. Palaeontol. 2013, 4, 475–513. [Google Scholar] [CrossRef]
- Kiselev, D.N.; Rogov, M.A. Detailed biostratigraphy of the Middle Callovian–lowest Oxfordian in the Mikhaylov reference section (Ryazan region, European part of Russia) by ammonites. Vol. Jurass. 2018, 16, 73–186. [Google Scholar] [CrossRef]
- Tesakova, E.M. Callovian and Oxfordian Ostracodes from the Central Region of the Russian Plate. Paleont J. 2003, 37, S107–S227. [Google Scholar]
- Gulyaev, D.B. New data on stratigraphy of the classical Callovian section near the village Alpat’evo (Oka river, Moscow Region). In Jurassic System of Russia: Problems of Stratigraphy and Paleogeography, Proceedings of the Fifth all-Russian Meeting; Zakharov, V.A., Rogov, M.A., Schurygin, B.N., Eds.; IzdatNaukaServis LLC: Yekaterinburg, Russia, 2013; pp. 53–57. (In Russian) [Google Scholar]
- Sasonov, N.T. Jurassic Deposits of the Central Areas of the Russian Platform; Gosgeoltehizdat: Leningrad, Russia, 1957. (In Russian) [Google Scholar]
- Meledina, S.V. Ammonites and zonal stratigraphy of the Callovian of Subboreal regions of USSR. Trans. Inst. Geol. Geophys. Sib. Branch. USSR Acad. Sci. 1987, 691, 1–182. (In Russian) [Google Scholar]
- Kiselev, D.N. Zones, Subzones, and Biohorizons of the Middle Callovian of Central Russia; Pedagogical University of Yaroslavl: Yaroslavl, Russia, 2001; pp. 1–38. (In Russian) [Google Scholar]
- Gulyaev, D.B.; Ippolitov, A.P. Detailed biostratigraphy of the Lower Callovian of the Kanev dislocations area (Cherkassy region, Ukraine). In Jurassic System of Russia: Problems of Stratigraphy and Paleogeography, Proceedings of the Fifth all-Russian meeting; Zakharov, V.A., Rogov, S.B.N., Eds.; IzdatNaukaServis LLC: Yekaterinburg, Russia, 2013; pp. 65–72. (In Russian) [Google Scholar]
- Tesakova, E. Late Callovian and Early Oxfordian ostracods from the Dubki section (Saratov area, Russia): Implications for stratigraphy, paleoecology, eustatic cycles and palaeobiogeography. Neues Jahrb. Geol. Paläontol. Abh. 2008, 249, 25–45. [Google Scholar] [CrossRef]
- Kiselev, D.; Rogov, M.; Glinskikh, L.; Guzhikov, A.; Pimenov, M.; Mikhailov, A.; Dzyuba, O.; Matveev, A.; Tesakova, E. Integrated stratigraphy of the reference sections for the Callovian/Oxfordian boundary in European Russia. Vol. Jurass. 2013, 11, 59–96. [Google Scholar]
- Mitta, V.V.; Alekseev, A.S.; Shik, S.M. (Eds.) Unified Regional Stratigraphic Scheme of the Jurassic of East European Platform; PIN RAS—VNIGNI: Moscow, Russia, 2012. (In Russian) [Google Scholar]
- Gulyaev, D.B. Stratigraphy of the Bathonian-Callovian nearboundary deposits of European Russia. In Jurassic System of Russia: Problems of Stratigraphy and Paleogeography, Proceedings of the Sixth All-Russian Meeting; Zakharov, V.A., Rogov, M.A., Ippolitov, A.P., Eds.; ALEF: Makhachkala, Russia, 2015; pp. 94–101. (In Russian) [Google Scholar]
- Kiselev, D.N. Ammonites and biostratigraphy of the Callovian beds in the Votcha section on the Sysola river. News Paleontol. Strat. 2006, 9, 47–69. [Google Scholar]
- Kiselev, D.N. Ammonites and Infrazonal Stratigraphy of the Boreal and Subboreal Bathonian and Callovian; GEOS: Moscow, Russia, 2022. (In Russian) [Google Scholar]
- Gulyaev, D.B.; Ippolitov, A.P. On the Bathonian-Callovian boundary deposits in the reference sections of the north of European Russia (Komi Republic). In Golden Age of Russian Malacology. Collective Volume of the All-Russia Research Cconference Dedicated to the 100th Anniversary of the Birth of Professor Viktor Nikolaevich Shimansky; Barskov, I.S., Ivanov, A.V., Leonova, T.B., Nikolayeva, S.V., Yashkov, I.A., Eds.; Borissiak Paleontological Institute of RAS–Yuri Gagarin Saratov State Technical University: Moscow–Saratov, Russia, 2016; pp. 235–248. (In Russian) [Google Scholar]
- Kiselev, D.N.; Ippolitov, A.P.; Rogov, M.A. Jurassic section near the Votcha village. In Jurassic Deposits of the Sysola River Basin in the Vicinities of Syktyvkar (Komi Republic) Guidebook of the Field Geological Excursion of the IX All-Russian Meeting “Jurassic System of Russia: Problems of Stratigraphy and Paleogeography”; Kiselev, D.N., Beznosov, P.A., Rogov, M.A., Ippolitov, A.P., Zverkov, N.G., Lyyurov, S.V., Eds.; Institute of Geology, Komi SC UB RAS: Syktyvkar, Russia, 2023; pp. 5–20. (In Russian) [Google Scholar]
- Mitta, V.V. Ammonites and stratigraphy of the Lower Callovian of the Russian Platform. Bull. VNIGRI 2000, 3, 1–144. (In Russian) [Google Scholar]
- Gründel, J.; Mitta, V. Gastropoden aus dem Untercallovium des Unzha-Beckens (Zentralrussland). Freib. Forschungshefte 2013, C545, 107–139. [Google Scholar]
- Keupp, H.; Mitta, V. Cephalopod jaws from the Middle Jurassic of Central Russia. Neues Jahrb. Geol. Paläontol. Abh. 2013, 270, 23–54. [Google Scholar] [CrossRef]
- Mitta, V.V. The genus Cadochamoussetia in the phylogeny of the Jurassic Cardioceratidae (Ammonoidea). In Advancing Research on Living and Fossil Cephalopods. Development and Evolution Form, Construction, and Function Taphonomy, Palaeoecology, Palaeobiogeography, Biostratigraphy, and Basin Analysis, Proceedings of the IV International Symposium on Cephalopods: Present and Past, Granada, Spain, 14–18 July 1996; Olóriz, F., Rodríguez-Tovar, F.J., Eds.; Kluwer Acad./Plenum Publ.: New York, NY, USA, 1999; pp. 125–136. [Google Scholar]
- Mitta, V.V. Late Bathonian Cardioceratidae (Ammonoidea) from the middle reaches of the Volga River. Paleontol. J. 2005, 39, 629–644. [Google Scholar]
- Gulyaev, D.B. Macrocephalitinae and Gowericeratinae (Ammonoidea) from the Elatmae Zone and the stratigraphy of the Lower Callovian of central areas of Russian Platform. In Problems of the Mesozoic Stratigraphy and Palaeontology. Lecturing in Memory of M.S. Mesezhnikov; Kozlova, G.E., Prozorovskyi, V.A., Eds.; VNIGRI: St.-Petersburg, Russia, 1999; pp. 63–85. (In Russian) [Google Scholar]
- Kiselev, D.N. The zone and subzone ammonite assemblages from the Middle Callovian in Central Russia. In Problems of the Mesozoic Stratigraphy and Palaeontology. Lecturing in memory of M.S. Mesezhnikov; Kozlova, G.E., Prozorovskyi, V.A., Eds.; VNIGRI: St.-Petersburg, Russia, 1999; pp. 87–115. (In Russian) [Google Scholar]
- Gerasimov, P.A.; Mitta, V.V.; Kochanova, M.D.; Tesakova, E.M. Fossils of the Callovian Stage of Central Russia; VNIGNI: Moscow, Russia, 1996; pp. 1–127. (In Russian) [Google Scholar]
- Tesakova, E.M.; Strezh, A.S.; Gulyaev, D.B. New lower Callovian ostracodes from the Kursk Region. Paleontol. J. 2009, 43, 258–271. [Google Scholar] [CrossRef]
- Ippolitov, A.P.; Gulyaev, D.B. Belemnite stratigraphy of the Lower Callovian of south-west of the Western Europe platform: Preliminary results. In Jurassic System of Russia: Problems of Stratigraphy and Paleogeography, Proceedings of the Fifth All-Russian Meeting; Zakharov, V.A., Rogov, S.B.N., Eds.; IzdatNaukaServis LLC: Yekaterinburg, Russia, 2013; pp. 85–89. (In Russian) [Google Scholar]
- Benson, R.B.J.; Druckenmiller, P.S. Faunal turnover of marine tetrapods during the Jurassic–Cretaceous transition. Biol. Rev. 2014, 89, 1–23. [Google Scholar] [CrossRef]
- Zverkov, N.G. A Problem of naming of the families of Late Jurassic and Cretaceous ichthyosaurs. Paleontol. J. 2022, 56, 463–470. [Google Scholar] [CrossRef]
- Johnson, M.M.; Young, M.T.; Brusatte, S.L. The phylogenetics of Teleosauroidea (Crocodylomorpha, Thalattosuchia) and implications for their ecology and evolution. PeerJ 2020, 8, e9808. [Google Scholar] [CrossRef]
- Young, M.T.; Brignon, A.; Sachs, S.; Hornung, J.; Foffa, D.; Kitson, J.J.N.; Johnson, M.M.; Steel, L. Cutting the Gordian knot: A historical and taxonomic revision of the Jurassic crocodylomorph Metriorhynchus. Zool. J. Linn. Soc. 2021, 192, 510–553. [Google Scholar] [CrossRef]
- Young, M.T.; Wilberg, E.W.; Johnson, M.M.; Herrera, Y.; de Andrade, M.B.; Brignon, A.; Sachs, S.; Abel, P.; Foffa, D.; Fernández, M.S.; et al. The history, systematics, and nomenclature of Thalattosuchia (Archosauria: Crocodylomorpha). Zool. J. Linn. Soc. 2024, 200, 547–617. [Google Scholar] [CrossRef]
- Meledina, S.V. Boreal Middle Jurassic of Russia. Trans. Inst. Geol. Geophys. Sib. Branch. Russ. Acad. Sci. 1994, 819, 1–184. (In Russian) [Google Scholar]
- Repin, Y.S.; Zakharov, V.A.; Meledina, S.V.; Nalnyaeva, T.I. Atlas of the mollusks of the Pechora Jurassic. Bull. VNIGRI 2006, 3, 1–262. (In Russian) [Google Scholar]
- Beznosov, P.A.; Zverkov, N.G. Fossil vertebrates in the Jurassic of the Ukhta area. In Jurassic deposits of the Ukhta area (Komi Republic); Rogov, M.A., Ed.; Institute of Geology Komi SC UB RAS: Syktyvkar, Russia, 2023; pp. 88–93. [Google Scholar]
- Beznosov, P.A.; Zverkov, N.G. Vertebrate remains from the Jurassic deposits of the Sysola River basin. In Jurassic Deposits of the Sysola River Basin in the Vicinities of Syktyvkar (Komi Republic). Guidebook of the Field Geological Excursion of the IX All-Russian Meeting “Jurassic System of Russia: Problems of Stratigraphy and Paleogeography”; Institute of Geology Komi SC UB RAS: Syktyvkar, Russia, 2023; p. 29. (In Russian) [Google Scholar]
- Malkov, B.A.; Lysuk, A.Y.; Zhornyak, A.V. New finds of fossil bones of Jurassic plesiosaurs and ichthyosaurs in the Callovian clays of Sysola depression. Vestn. Inst. Geol. Komi SC UB RAS. 2004, 9, 28–30. (In Russian) [Google Scholar]
- Kassin, N. Carte Géologique Générale de la Partie Européenne De l’U.R.S.S. Feuille 107. Région Viatka—Slobodskoi—Omoutninsk—Kai; Comité Géologique: Leningrad, Russia, 1928. (In Russian) [Google Scholar]
- Arkhangelsky, M.S. On a find of incomplete ichthyosaur skeleton in the lower Callovian deposits of Saratov Region. In Materials of the First All-Russian Meeting “Jurassic System of Russia: Problems of Stratigraphy and Paleogeography”; Zakharov, V.A., Rogov, M.A., Dzyuba, O.S., Eds.; GIN RAS: Moscow, Russia, 2005; p. 4. (In Russian) [Google Scholar]
- Noè, L.F. A Taxonomic and Functional Study of the Callovian (Middle Jurassic) Pliosauroidea (Reptilia; Sauropterygia). Ph.D. Thesis, School of Environmental and Applied Sciences, University of Derby, Derby, UK, 2001. [Google Scholar]
- Linder, H. Beitrage zur Kenntnis der Plesiosaurier-Gattungen Peloneustes und Pliosaurus. Geol. Palæontol. Abh. 1913, 11, 339–401. [Google Scholar]
- Benson, R.; Evans, M.; Smith, A.S.; Sassoon, J.; Moore-Faye, S.; Ketchum, H.F.; Forrest, R. A giant pliosaurid skull from the Late Jurassic of England. PLoS ONE 2013, 8, e65989. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.S.; Benson, R.B.J. Osteology of Rhomaleosaurus thorntoni (Sauropterygia: Rhomaleosauridae) from the Lower Jurassic (Toarcian) of Northamptonshire, England. Monogr. Palaeontogr. Soc. 2014, 168, 1–40. [Google Scholar] [CrossRef]
- Newman, B.; Tarlo, L.B. A giant marine reptile from Bedfordshire. Animals 1967, 10, 61–63. [Google Scholar]
- O’Keefe, F.R.; Street, H.P.; Wilhelm, B.C.; Richards, C.D.; Zhu, H. A new skeleton of the cryptoclidid plesiosaur Tatenectes laramiensis reveals a novel body shape among plesiosaurs. J. Vertebr. Paleontol. 2011, 31, 330–339. [Google Scholar] [CrossRef]
- Brown, D.S.; Keen, J.A. An extensive marine vertebrate fauna from the Kellaways Sand (Callovian, Middle Jurassic) of Lincolnshire. Mercian Geol. 1991, 12, 87–96. [Google Scholar]
- O’Keefe, F.R.; Street, H.P. Osteology of the cryptocleidoid plesiosaur Tatenectes laramiensis, with comments on the taxonomic status of the Cimoliasauridae. J. Vertebr. Paleontol. 2009, 29, 48–57. [Google Scholar] [CrossRef]
- Zverkov, N.G.; Prilepskaya, N.E. A prevalence of Arthropterygius (Ichthyosauria: Ophthalmosauridae) in the Late Jurassic—Earliest Cretaceous of the Boreal Realm. PeerJ 2019, 7, e6799. [Google Scholar] [CrossRef] [PubMed]
- Zverkov, N.G.; Jacobs, M.L. Revision of Nannopterygius (Ichthyosauria: Ophthalmosauridae): Reappraisal of the “inaccessible” holotype resolves a taxonomic tangle and reveals an obscure ophthalmosaurid lineage with a wide distribution. Zool. J. Linn. Soc. 2021, 191, 228–275. [Google Scholar] [CrossRef]
- Fischer, V.; Weis, R.; Thuy, B. Refining the marine reptile turnover at the Early–Middle Jurassic transition. PeerJ 2021, 9, e10647. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, C.W. Osteology of Baptanodon. Mem. Carnegie Mus. 1905, 2, 77–129. [Google Scholar] [CrossRef]
- Arkhangelsky, M.S.; Zverkov, N.G.; Spasskaya, O.S.; Evgrafov, A.V. On the first reliable record of the ichthyosaur Ophthalmosaurus icenicus Seeley in the Oxfordian–Kimmeridgian beds of European Russia. Paleontol. J. 2018, 52, 49–57. [Google Scholar] [CrossRef]
- Bardet, N. Extinction events among Mesozoic marine reptiles. Hist. Biol. 1994, 7, 313–324. [Google Scholar] [CrossRef]
- Fernández, M. A new long-snouted ichthyosaur from the early Bajocian of Neuquén basin (Argentina). Ameghiniana 1994, 31, 291–297. [Google Scholar]
- Fernández, M. A new ichthyosaur from the Los Molles Formation (Early Bajocian), Neuquén basin, Argentina. J. Paleontol. 1999, 73, 677–681. [Google Scholar] [CrossRef]
- Fernández, M. Ophthalmosauria (Ichthyosauria) forefin from the Aalenian-Bajocian boundary of Mendozo Province, Argentina. J. Vertebr. Paleontol. 2003, 23, 691–694. [Google Scholar] [CrossRef] [PubMed]
- Cau, A.; Fanti, F. The oldest known metriorhynchid crocodylian from the Middle Jurassic of North-eastern Italy: Neptunidraco ammoniticus gen. et sp. nov. Gondwana Res. 2011, 19, 550–565. [Google Scholar] [CrossRef]
- Maxwell, E.E.; Fernández, M.S.; Schoch, R.R. First diagnostic marine reptile remains from the Aalenian (Middle Jurassic): A new ichthyosaur from southwestern Germany. PLoS ONE 2012, 7, e41692. [Google Scholar] [CrossRef] [PubMed]
- Bardet, N.; Falconnet, J.; Fischer, V.; Houssaye, A.; Jouve, S.; Pereda Suberbiola, X.; Pérez-García, A.; Rage, J.-C.; Vincent, P. Mesozoic marine reptile palaeobiogeography in response to drifting plates. Gondwana Res. 2014, 26, 869–887. [Google Scholar] [CrossRef]
- Sachs, S.; Hornung, J. Marine Tetrapoden aus dem Mittleren Jura (Aalenium und Bajocium) von Bielefeld (Nordwestdeutschland) Berichte Naturwiss. Ver. Für Bielef. Und Umgeg. 2015, 53, 52–73. [Google Scholar]
- Arkell, W.J. The Jurassic System in Great Britain; Clarendon Press: Oxford, UK, 1933. [Google Scholar]
- Araujo, R.; Smith, A.S.; Liston, J. The Alfred Leeds Fossil Vertebrate Collection of the National Museum of Ireland—Natural History. Ir. J. Earth Sci. 2008, 26, 17–32. [Google Scholar]
- Martill, D.M. The stratigraphic distribution of fossil vertebrates in the Oxford Clay of England. Mercian Geol. 1986, 10, 161–186. [Google Scholar]
- Martill, D.M.; Hudson, J.D. (Eds.) Fossils of the Oxford Clay, Palaeontological Association Field Guide to Fossils; Palaeontological Association: London, UK, 1991. [Google Scholar]
- Hudson, J.D.; Martill, D.M. The Peterborough Member (Callovian, Middle Jurassic) of the Oxford Clay Formation at Peterborough, UK. J. Geol. Soc. 1994, 151, 113–124. [Google Scholar] [CrossRef]
- Delair, J.B. The Mesozoic reptiles of Dorset, part two. Proc. Dorset Nat. Hist. Archaeol. Soc. 1960, 80, 52–75. [Google Scholar]
- Foffa, D.; Young, M.; Brusatte, S.L.; Graham, M.R.; Steel, L. A new metriorhynchid crocodylomorph from the Oxford Clay Formation (Middle Jurassic) of England, with implications for the origin and diversification of Geosaurini. J. Syst. Palaeontol. 2018, 16, 1123–1143. [Google Scholar] [CrossRef]
- Rogov, M.; Zakharov, V.; Kiselev, D. Molluscan immigrations via biogeographical ecotone of the Middle Russian Sea during the Jurassic. Vol. Jurass. 2008, 6, 143–152. [Google Scholar]
- Foffa, D.; Young, M.; Brusatte, S. Filling the Corallian gap: New information on Late Jurassic marine reptile faunas from England. Acta Palaeontol. Pol. 2018, 63, 287–313. [Google Scholar] [CrossRef]

















Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zverkov, N.; Arkhangelsky, M.; Gulyaev, D.; Ippolitov, A.; Shmakov, A. Callovian Marine Reptiles of European Russia. Diversity 2024, 16, 290. https://doi.org/10.3390/d16050290
Zverkov N, Arkhangelsky M, Gulyaev D, Ippolitov A, Shmakov A. Callovian Marine Reptiles of European Russia. Diversity. 2024; 16(5):290. https://doi.org/10.3390/d16050290
Chicago/Turabian StyleZverkov, Nikolay, Maxim Arkhangelsky, Denis Gulyaev, Alexey Ippolitov, and Alexey Shmakov. 2024. "Callovian Marine Reptiles of European Russia" Diversity 16, no. 5: 290. https://doi.org/10.3390/d16050290
APA StyleZverkov, N., Arkhangelsky, M., Gulyaev, D., Ippolitov, A., & Shmakov, A. (2024). Callovian Marine Reptiles of European Russia. Diversity, 16(5), 290. https://doi.org/10.3390/d16050290