


The Complete Mitochondrial Genomes of Aelia sibirica and A. fieberi (Hemiptera, Pentatomidae), and Phylogenetic Implications

Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling, DNA Extraction, and Sequencing
2.2. Genome Annotations and Sequence Analyses
2.3. Phylogenetic Analysis
3. Results
3.1. Genomic Features
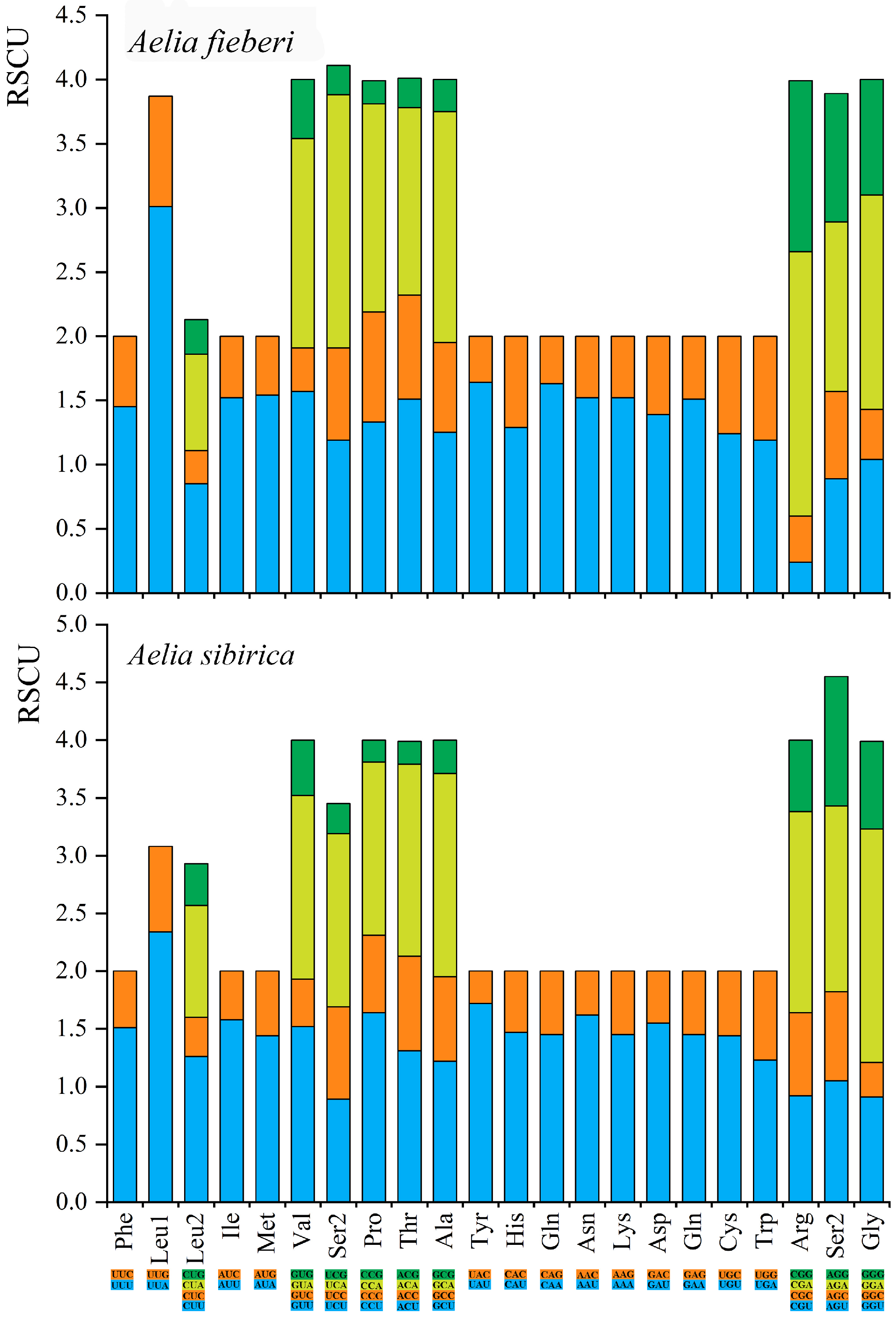
3.2. Nucleotide Composition and Codon Usage
3.3. PCGs
3.4. Ribosomal and Transfer RNA Genes
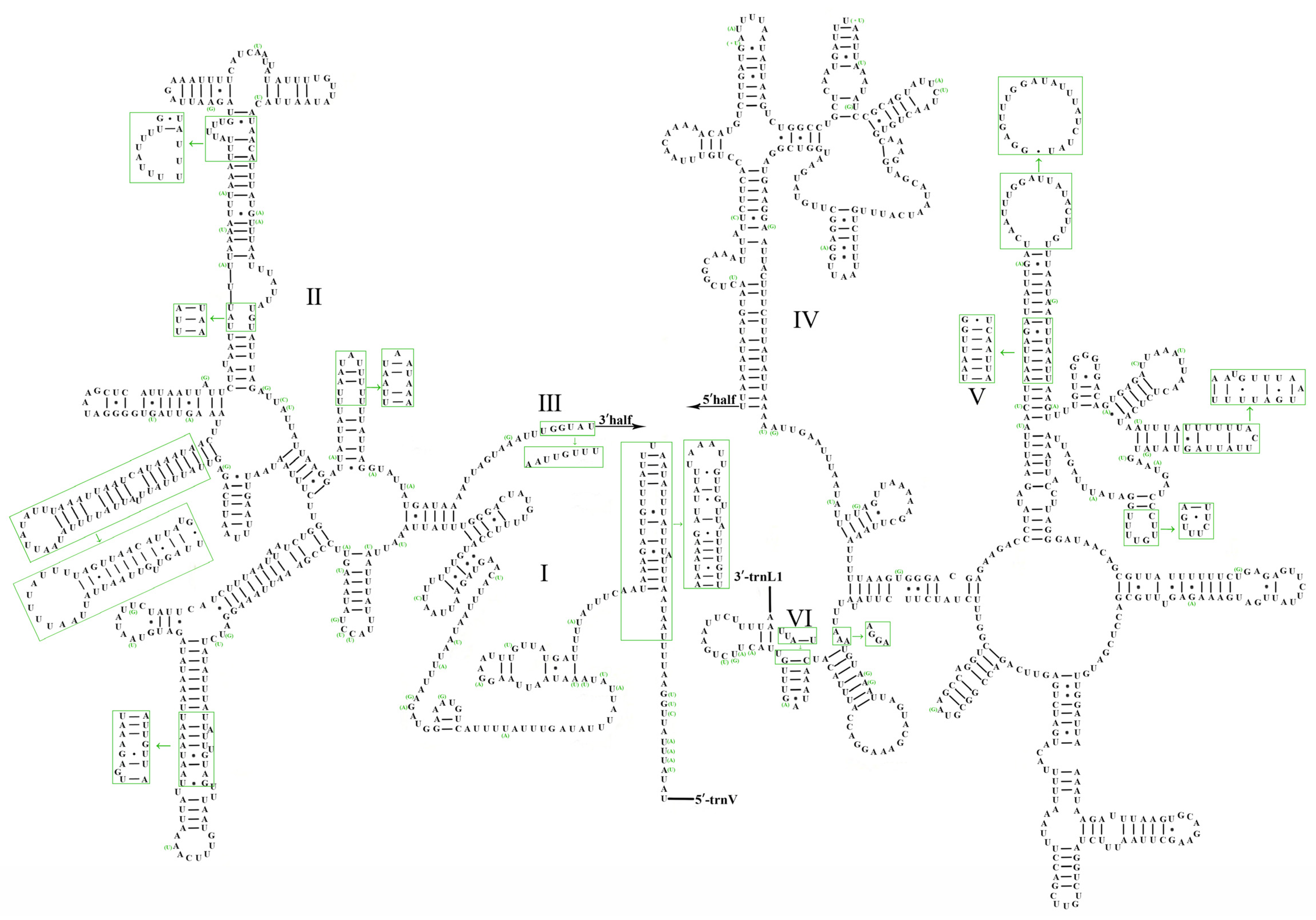
3.5. Control Region
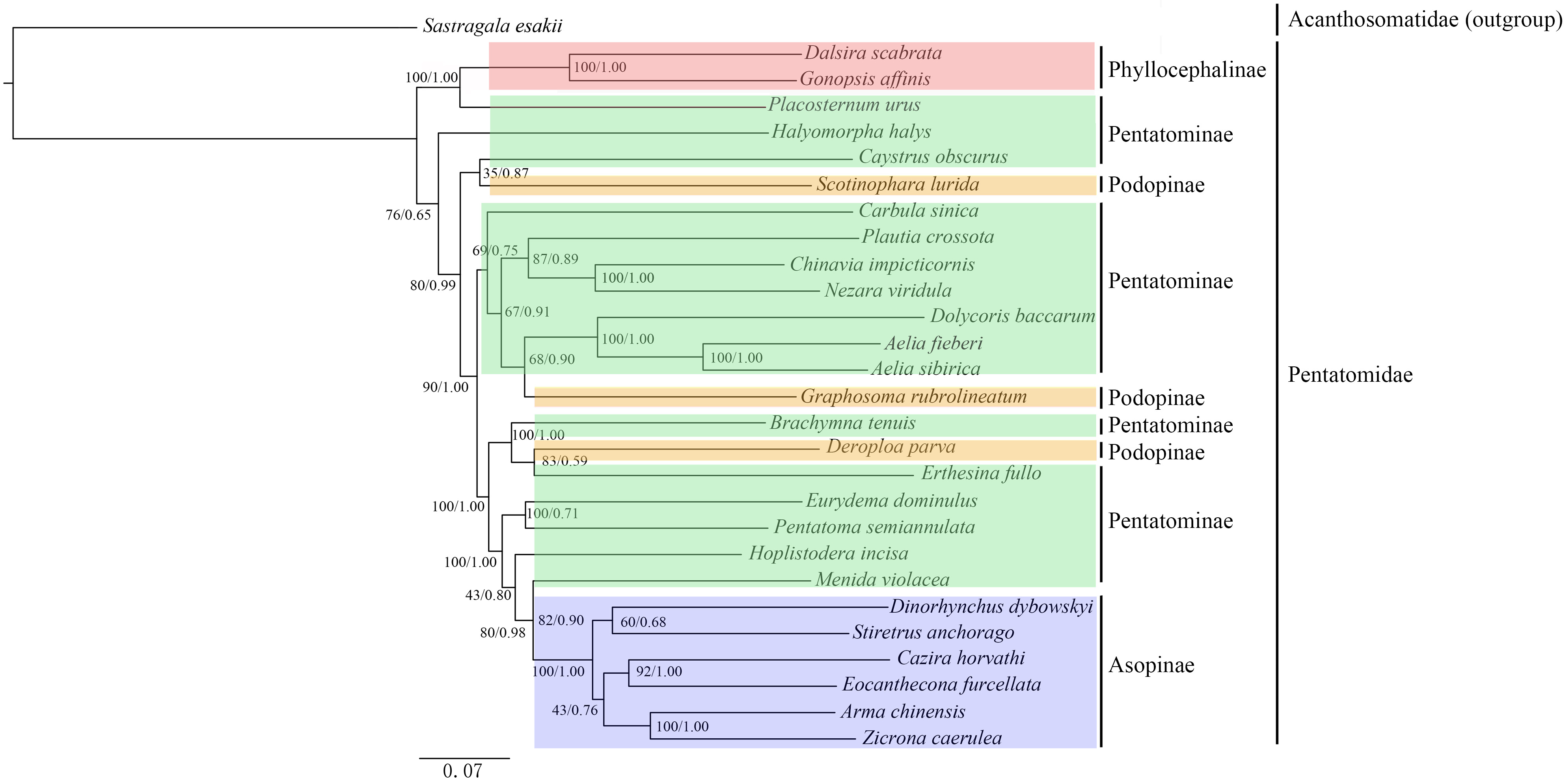
3.6. Phylogenetic Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wolstenholme, D.R. Animal Mitochondrial DNA: Structure and Evolution. In International Review of Cytology; Wolstenholme, D.R., Jeon, K.W., Eds.; Academic Press: Cambridge, MA, USA, 1992; Volume 141, pp. 173–216. [Google Scholar]
- Boore, J.L. Animal Mitochondrial Genomes. Nucleic Acids Res. 1999, 27, 1767–1780. [Google Scholar] [CrossRef]
- Simon, C.; Buckley, T.R.; Frati, F.; Stewart, J.B.; Beckenbach, A.T. Incorporating Molecular Evolution into Phylogenetic Analysis, and a New Compilation of Conserved Polymerase Chain Reaction Primers for Animal Mitochondrial DNA. Annu. Rev. Ecol. Evol. Syst. 2006, 37, 545–579. [Google Scholar] [CrossRef]
- Liu, Y.; Li, H.; Song, F.; Zhao, Y.; Wilson, J.-J.; Cai, W. Higher-Level Phylogeny and Evolutionary History of Pentatomomorpha (Hemiptera: Heteroptera) Inferred from Mitochondrial Genome Sequences. Syst. Entomol. 2019, 44, 810–819. [Google Scholar] [CrossRef]
- Jeong, J.S.; Park, J.S.; Sohn, J.-C.; Kim, M.J.; Kim, I. Complete Mitochondrial Genome of the Hawthorn Moth Scythropia Crataegella Linnaeus, 1767 (Lepidoptera: Scythropiidae). J. Asia-Pac. Entomol. 2023, 26, 102103. [Google Scholar] [CrossRef]
- Song, N.; Wu, Q.; Lin, X.; Zhang, Y.; Zhao, T. The Complete Mitochondrial Genome of Nephrotoma Scalaris Parvinotata (Brunetti 1918) (Diptera: Tipulidae) and Insights into the Phylogeny of Tipulomorpha. J. Asia-Pac. Entomol. 2023, 26, 102133. [Google Scholar] [CrossRef]
- Song, N.; Zhang, H.; Zhao, T. Insights into the Phylogeny of Hemiptera from Increased Mitogenomic Taxon Sampling. Mol. Phylogenet. Evol. 2019, 137, 236–249. [Google Scholar] [CrossRef]
- Mu, Y.-L.; Zhang, C.-H.; Zhang, Y.-J.; Yang, L.; Chen, X.-S. Characterizing the Complete Mitochondrial Genome of Arma Custos and Picromerus Lewisi (Hemiptera: Pentatomidae: Asopinae) and Conducting Phylogenetic Analysis. J. Insect Sci. 2022, 22, 6. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, D.; Jia, Q.; Wu, X.; Zhang, H. Characterization of the Complete Mitochondrial Genome of Myrmus Lateralis (Heteroptera, Rhopalidae) and Its Implication for Phylogenetic Analyses. ZooKeys 2021, 1070, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Dowton, M.; Castro, L.R.; Austin, A.D. Mitochondrial Gene Rearrangements as Phylogenetic Characters in the Invertebrates: The Examination of Genome “Morphology”. Invert. Syst. 2002, 16, 345–356. [Google Scholar] [CrossRef]
- Françoso, E.; Gomes, F.; Arias, M.C. A Protocol for Isolating Insect Mitochondrial Genomes: A Case Study of NUMT in Melipona Flavolineata (Hymenoptera: Apidae). Mitochondrial DNA Part A 2016, 27, 2401–2404. [Google Scholar] [CrossRef] [PubMed]
- Djoumad, A.; Nisole, A.; Zahiri, R.; Freschi, L.; Picq, S.; Gundersen-Rindal, D.E.; Sparks, M.E.; Dewar, K.; Stewart, D.; Maaroufi, H.; et al. Comparative Analysis of Mitochondrial Genomes of Geographic Variants of the Gypsy Moth, Lymantria Dispar, Reveals a Previously Undescribed Genotypic Entity. Sci. Rep. 2017, 7, 14245. [Google Scholar] [CrossRef]
- Talavera, G.; Vila, R. What Is the Phylogenetic Signal Limit from Mitogenomes? The Reconciliation between Mitochondrial and Nuclear Data in the Insecta Class Phylogeny. BMC Evol. Biol. 2011, 11, 315. [Google Scholar] [CrossRef]
- Fedorov, V.B.; Trucchi, E.; Goropashnaya, A.V.; Stenseth, N.C. Conflicting Nuclear and Mitogenome Phylogenies Reveal Ancient Mitochondrial Replacement between Two North American Species of Collared Lemmings (Dicrostonyx Groenlandicus, D. Hudsonius). Mol. Phylogenet. Evol. 2022, 168, 107399. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, D.; Gao, Z.; Liu, J.; Yi, W.; Zhang, H. Complete mitochondrial genome sequence and phylogenetic implications of Chorosoma macilentum (Heteroptera: Rhopalidae). Mitochondrial DNA Part B 2019, 4, 3882–3883. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.Y.; Gui, H. Insect Classification; Nanjing Normal University Press: Nanjing, China, 1999. [Google Scholar]
- McPherson, J.E.; Esquivel, J.F.; Musolin, D.L.; Jones, W.A.; Hoelmer, K.A. Invasive Stink Bugs and Related Species (Pentatomoidea): Biology, Higher Systematics, Semiochemistry, and Management; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Rider, D.A.; Schwertner, C.F.; Vilimová, J.; Redei, D.; Kment, P.; Thomas, D.B. Higher Systematics of the Pentatomoidea. In Invasive Stink Bugs and Related Species (Pentatomoidea); CRC Press: Boca Raton, CA, USA, 2018. [Google Scholar]
- Genevcius, B.C.; Greve, C.; Koehler, S.; Simmons, R.B.; Rider, D.A.; Grazia, J.; Schwertner, C.F. Phylogeny of the Stink Bug Tribe Chlorocorini (Heteroptera, Pentatomidae) Based on DNA and Morphological Data: The Evolution of Key Phenotypic Traits. Syst. Entomol. 2021, 46, 327–338. [Google Scholar] [CrossRef]
- Roca-Cusachs, M.; Schwertner, C.F.; Kim, J.; Eger, J.; Grazia, J.; Jung, S. Opening Pandora’s Box: Molecular Phylogeny of the Stink Bugs (Hemiptera: Heteroptera: Pentatomidae) Reveals Great Incongruences in the Current Classification. Syst. Entomol. 2022, 47, 36–51. [Google Scholar] [CrossRef]
- Xu, Z.Q. Phylogeny and Cladistic Analysis of the Pentatomoidea (Hemiptera: Heteroptera). Acta Agric. Univ. Pekin. 1993, 19, 49–53. [Google Scholar]
- Weirauch, C.; Schuh, R.T. Systematics and Evolution of Heteroptera: 25 Years of Progress. Annu. Rev. Entomol. 2011, 56, 487–510. [Google Scholar] [CrossRef] [PubMed]
- Hua, J.; Li, M.; Dong, P.; Cui, Y.; Xie, Q.; Bu, W. Comparative and Phylogenomic Studies on the Mitochondrial Genomes of Pentatomomorpha (Insecta: Hemiptera: Heteroptera). BMC Genom. 2008, 9, 610. [Google Scholar] [CrossRef] [PubMed]
- Grazia, J.; Schuh, R.T.; Wheeler, W.C. Phylogenetic Relationships of Family Groups in Pentatomoidea Based on Morphology and DNA Sequences (Insecta: Heteroptera). Cladistics 2008, 24, 932–976. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.Z.; Rédei, D.; Eger, J., Jr.; Wang, Y.H.; Wu, H.Y.; Carapezza, A.; Kment, P.; Cai, B.; Sun, X.-Y.; Guo, P.-L.; et al. Phylogeny and the Colourful History of Jewel Bugs (Insecta: Hemiptera: Scutelleridae). Cladistics 2018, 34, 502–516. [Google Scholar] [CrossRef]
- Yuan, M.-L.; Zhang, Q.-L.; Guo, Z.-L.; Wang, J.; Shen, Y.-Y. Comparative Mitogenomic Analysis of the Superfamily Pentatomoidea (Insecta: Hemiptera: Heteroptera) and Phylogenetic Implications. BMC Genom. 2015, 16, 460. [Google Scholar] [CrossRef]
- Zhao, W.; Zhao, Q.; Li, M.; Wei, J.; Zhang, X.; Zhang, H. DNA Barcoding of Chinese Species of the Genus Eurydema Laporte, 1833 (Hemiptera: Pentatomidae). Zootaxa 2017, 4286, 151–175. [Google Scholar] [CrossRef]
- Ding, X.; Chen, C.; Wei, J.; Gao, X.; Zhang, H.; Zhao, Q. Comparative Mitogenomics and Phylogenetic Analyses of the Genus Menida (Hemiptera, Heteroptera, Pentatomidae). ZooKeys 2023, 1138, 29–48. [Google Scholar] [CrossRef]
- Derzhansky, V.; Péricart, J.; Pluot-Sigwalt, D. Notes on Synonymy and Distribution of Some Species of the Genus Aelia (Heteroptera: Pentatomidae). Zoosyst. Ross. 2007, 15, 298. [Google Scholar] [CrossRef]
- Gibicsár, S.; Keszthelyi, S. Topographical Based Significance of Sap-Sucking Heteropteran in European Wheat Cultivations: A Systematic Review. Diversity 2023, 15, 109. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-Miseq: An Updated Pipeline to Assemble Microbial Genomes from Illumina MiSeq Data. Bioinformatics 2014, 31, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de Novo Metazoan Mitochondrial Genome Annotation. Mol. Phylogenet. Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef]
- Grant, J.R.; Stothard, P. The CGView Server: A Comparative Genomics Tool for Circular Genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Perna, N.T.; Kocher, T.D. Patterns of Nucleotide Composition at Fourfold Degenerate Sites of Animal Mitochondrial Genomes. J. Mol. Evol. 1995, 41, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Benson, G. Tandem Repeats Finder: A Program to Analyze DNA Sequences. Nucleic Acids Res. 1999, 27, 573–580. [Google Scholar] [CrossRef] [PubMed]
- Vaidya, G.; Lohman, D.J.; Meier, R. SequenceMatrix: Concatenation Software for the Fast Assembly of Multi-Gene Datasets with Character Set and Codon Information. Cladistics 2011, 27, 171–180. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Stamatakis, A. Using RAxML to Infer Phylogenies. Curr. Protoc. Bioinform. 2015, 51, 6–14. [Google Scholar] [CrossRef]
- Yan, L.; Xu, W.; Zhang, D.; Li, J. Comparative Analysis of the Mitochondrial Genomes of Flesh Flies and Their Evolutionary Implication. Int. J. Biol. Macromol. 2021, 174, 385–391. [Google Scholar] [CrossRef]
- Yuan, L.; Ge, X.; Xie, G.; Liu, H.; Yang, Y. First Complete Mitochondrial Genome of Melyridae (Coleoptera, Cleroidea): Genome Description and Phylogenetic Implications. Insects 2021, 12, 87. [Google Scholar] [CrossRef]
- Lee, W.; Kang, J.; Jung, C.; Hoelmer, K.; Lee, S.H.; Lee, S. Complete Mitochondrial Genome of Brown Marmorated Stink Bug Halyomorpha Halys (Hemiptera: Pentatomidae), and Phylogenetic Relationships of Hemipteran Suborders. Mol. Cells 2009, 28, 155–165. [Google Scholar] [CrossRef]
- Mi, Q.; Zhang, J.; Gould, E.; Chen, J.; Sun, Z.; Zhang, F. Biology, Ecology, and Management of Erthesina Fullo (Hemiptera: Pentatomidae): A Review. Insects 2020, 11, 346. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Ji, Y.; Li, H.; Song, F.; Zhang, L.; Wang, M. Characterization of the Complete Mitochondrial Genome of Pentatoma Semiannulata (Hemiptera: Pentatomidae). Mitochondrial DNA Part B 2021, 6, 750–752. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Cassis, G.; Zhao, L.; He, Y.; Zhang, H.; Wei, J. The Complete Mitochondrial Genome of Zicrona caerulea (Linnaeus) (Hemiptera: Pentatomidae: Asopinae) and Its Phylogenetic Implications. Zootaxa 2020, 4747, 547–561. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wei, J.; Zhao, W.; Chen, C.; Gao, X.; Zhao, Q. The Complete Mitochondrial Genome of Pentatoma Rufipes (Hemiptera, Pentatomidae) and Its Phylogenetic Implications. ZooKeys 2021, 1042, 51–72. [Google Scholar] [CrossRef]
- Hassanin, A.; Leger, N.; Deutsch, J. Evidence for Multiple Reversals of Asymmetric Mutational Constraints during the Evolution of the Mitochondrial Genome of Metazoa, and Consequences for Phylogenetic Inferences. Syst. Biol. 2005, 54, 277–298. [Google Scholar] [CrossRef]
- Jiang, P. Studies on the Comparative Mitochondrial Genomics and Phylogeny of Heteroptera (Insecta: Hemiptera). Ph.D. Thesis, China Agricultural University, Beijing, China, 2017. [Google Scholar]
- Sun, C.-H.; Zhang, Y.-N.; Zeng, X.-S.; Liu, D.-W.; Huang, Q.; Zhang, X.-L.; Zhang, Q. Mitogenome of Knodus Borki (Cypriniformes: Characidae): Genomic Characterization and Phylogenetic Analysis. Mol. Biol. Rep. 2022, 49, 1741–1748. [Google Scholar] [CrossRef]
- He, H.; Yan, B.; Yu, X.; Yang, M. Characterization and Phylogenetic Implications of Newly Sequenced Mitogenomes of Five Mileewa and Processina Species from China (Hemiptera: Cicadellidae: Mileewinae). Sci. Rep. 2022, 12, 20852. [Google Scholar] [CrossRef]
- Xu, S.; Wu, Y.; Liu, Y.; Zhao, P.; Chen, Z.; Song, F.; Li, H.; Cai, W. Comparative Mitogenomics and Phylogenetic Analyses of Pentatomoidea (Hemiptera: Heteroptera). Genes 2021, 12, 1306. [Google Scholar] [CrossRef] [PubMed]
- Lian, D.; Wei, J.; Ding, X.; Liu, Y.; Zhao, Q. Comparison and application of tRNA and rRNA genes in the mitochondrial genome of Pentatomidae (Hemiptera: Pentatomoidea). J. Fujian Agric. For. Univ. (Nat. Sci. Ed.) 2022, 51, 782–791. [Google Scholar]
- Lian, D.; Wei, J.; Chen, C.; Niu, M.; Zhang, H.; Zhao, Q. Comparative analysis and phylogeny of mitochondrial genomes of Pentatomidae (Hemiptera: Pentatomoidea). Front. Genet. 2022, 13, 1045193. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.I. Hemiptera, Pentatomidae. In Economic Insect Fauna of China; Academia Sinica, Science Press: Beijing, China, 1962; Volume 2, pp. 67–128. [Google Scholar]
- Zhao, W.; Zhao, Q.; Li, M.; Wei, J.; Zhang, X.; Zhang, H. Comparative mitogenomic analysis of the Eurydema genus in the context of representative Pentatomidae (Hemiptera: Heteroptera) taxa. J. Insect Sci. 2019, 19, 20. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Li, M.; Yan, J.; Bai, M.; Zhang, H. Five mitochondrial genomes of the genus Eysarcoris Hahn, 1834 with phylogenetic implications for the Pentatominae (Hemiptera: Pentatomidae). Insects 2021, 12, 597. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placement | Species | GenBank Accession Number | ||
|---|---|---|---|---|
| Acanthosomatidae | Sastragala esakii | NC_058975 | ||
| Pentatomidae | Phyllocephalinae | Gonopsis affinis | NC_036745 | |
| Dalsira scabrata | KX505855 | |||
| Asopinae | Arma chinensis | MW355500 | ||
| Cazira horvathi | MF497718 | |||
| Dinorhynchus dybowskyi | NC_037724 | |||
| Eocanthecona furcellata | MZ440302 | |||
| Stiretrus anchorago | BK059217 | |||
| Zicrona caerulea | MW847250 | |||
| Podopinae | Deroploa parva | MW679032 | ||
| Graphosoma rubrolineatum | KX267740 | |||
| Scotinophara lurida | MF497733 | |||
| Pentatominae | Aeliini | Aelia fieberi | ON059969 | |
| Aelia sibirica | NC_067883 | |||
| Antestiini | Plautia crossota | MK757497 | ||
| Sephelini | Brachymna tenuis | MF497711 | ||
| Eysarcorini | Carbula sinica | NC_037741 | ||
| Caystrini | Caystrus obscurus | MF497717 | ||
| Carpocorini | Dolycoris baccarum | JQ743672 | ||
| Halyini | Erthesina fullo | MK374364 | ||
| Strachiini | Eurydema dominulus | MG584833 | ||
| Cappaeini | Halyomorpha halys | LC579925 | ||
| Hoplistoderini | Hoplistodera incisa | MF620037 | ||
| Menidini | Menida violacea | MF497728 | ||
| Nezarini | Nezara viridula | NC_011755 | ||
| Chinavia impicticornis | MG253262 | |||
| Pentatomini | Pentatoma semiannulata | MT985377 | ||
| Placosternum urus | MF497730 | |||
| Gene | Size (bp) | Position | IGS (bp) | Anti- Codon | Start Codon | Stop Codon | |
|---|---|---|---|---|---|---|---|
| Start | Stop | ||||||
| trnI | 66/66 | 1/1 | 66/66 | 0/8 | GAU | — | — |
| trnQ | 69/69 | 64/75 | 132/143 | −3/−3 | UUG | — | — |
| trnM | 66/66 | 152/150 | 217/215 | 19/6 | CAU | — | — |
| nad2 | 987/987 | 218/216 | 1204/1202 | 0/0 | — | ATC | TAA |
| trnW | 66/66 | 1211/1221 | 1276/1286 | 6/18 | UCA | — | — |
| trnC | 62/66 | 1269/1279 | 1330/1344 | −8/−8 | GCA | — | — |
| trnY | 65/65 | 1334/1348 | 1398/1412 | 3/3 | GUA | — | — |
| cox1 | 1542/1542 | 1408/1422 | 2949/2963 | 9/9 | — | TTG | TAA |
| trnL2UUR | 65/66 | 2945/2959 | 3009/3024 | −5/−5 | UAA | — | — |
| cox2 | 679/679 | 3010/3025 | 3688/3703 | −5/0 | — | ATA | T |
| trnK | 71/70 | 3689/3704 | 3759/3773 | 0/0 | CUU | — | — |
| trnD | 66/66 | 3774/3770 | 3839/3835 | 14/−4 | GUC | — | — |
| atp8 | 159/159 | 3840/3836 | 3998/3994 | 0/0 | — | TTG/GTG | TAA |
| atp6 | 675/675 | 3992/3988 | 4666/4662 | −7/3 | — | ATG | TAA |
| cox3 | 789/789 | 4669/4665 | 5457/5453 | 2/2 | — | ATG | TAA |
| trnG | 63/63 | 5464/5458 | 5526/5520 | 6/4 | UCC | — | — |
| nad3 | 352/377 | 5527/5521 | 5878/5897 | 0/0 | — | ATA | TAG/T |
| trnA | 64/66 | 5879/5898 | 5942/5963 | 0/0 | UGC | — | — |
| trnR | 64/61 | 5947/5973 | 6010/6033 | 4/9 | UCG | — | — |
| trnN | 68/67 | 6016/6037 | 6083/6103 | 5/3 | GUU | — | — |
| trnS1AGN | 70/69 | 6083/6103 | 6152/6171 | −1/−1 | GCU | — | — |
| trnE | 68/69 | 6155/6173 | 6222/6241 | 2/2 | UUC | — | — |
| trnF | 66/66 | 6221/6240 | 6286/6305 | −2/−2 | GAA | — | — |
| nad5 | 1707/1706 | 6286/6307 | 7992/8011 | −1/1 | — | ATT | TAA/T |
| trnH | 64/64 | 7994/8013 | 8057/8076 | 1/1 | GUG | — | — |
| nad4 | 1329/1329 | 8061/8080 | 9389/9408 | 3/3 | — | ATT/ATG | TAA |
| nad4L | 288/288 | 9383/9402 | 9670/9689 | −7/−7 | — | ATG/ATT | TAA |
| trnT | 67/65 | 9673/9692 | 9739/9756 | 2/2 | UGU | — | — |
| trnP | 66/66 | 9740/9757 | 9805/9822 | 0/0 | UGG | — | — |
| nad6 | 465/480 | 9816/9833 | 10,280/10,312 | 10/10 | — | ATG/ATA | TAA |
| cytb | 1137/1147 | 10,288/10,317 | 11,424/11,456 | 7/4 | — | ATG | TAA |
| trnS2UCN | 69/69 | 11,426/11,465 | 11,494/11,533 | 1/8 | UGA | — | — |
| nad1 | 924/924 | 11,522/11,558 | 12,445/12,481 | 27/24 | — | TTG | TAG |
| trnL1CUN | 65/65 | 12,446/12,482 | 12,510/12,546 | 0/0 | UAG | — | — |
| 16S rRNA | 1282/1272 | 12,510/12,546 | 13,791/13,817 | −1/−1 | — | — | — |
| trnV | 68/68 | 13,792/13,818 | 13,859/13,885 | 0/0 | UAC | — | — |
| 12S rRNA | 796/795 | 13,860/13,886 | 14,655/14,680 | 0/0 | — | — | — |
| Control region | 717/768 | 14,656/14,681 | 15,372/15,448 | 0 | — | — | — |
| Gene | A/% | T/% | G/% | C/% | (A + T)/% | AT-Skew | GC-Skew |
|---|---|---|---|---|---|---|---|
| Whole genome | 41.8/41.8 | 31.7/31.7 | 11.0/11.0 | 15.5/15.5 | 73.5/73.5 | 0.138/0.138 | −0.167/−0.167 |
| 13PCGs | 33.1/32.6 | 40.4/40.3 | 13.1/13.3 | 13.5/13.8 | 73.5/72.9 | −0.100/−0.105 | −0.015/−0.017 |
| 22tRNAs | 38.4/37.6 | 37.1/37.2 | 13.4/13.9 | 11.1/11.3 | 75.5/74.8 | 0.017/0.005 | 0.094/0.106 |
| 2rRNAs | 33.1/33.4 | 44.1/43.6 | 14.1/14.7 | 8.6/8.2 | 77.2/77.1 | −0.142/−0.132 | 0.242/0.283 |
| Control region | 35.8/34.0 | 34.7/35.2 | 12.3/12.3 | 17.2/18.5 | 70.5/69.2 | 0.016/−0.017 | −0.166/−0.203 |
| nad1 | 24.5/24.7 | 50.0/50.2 | 16.8/16.6 | 8.8/8.5 | 74.5/74.9 | −0.343/−0.341 | 0.313/0.319 |
| nad2 | 44.2/44.3 | 35.6/34.5 | 8.9/9.2 | 11.3/12.0 | 79.8/78.8 | 0.108/0.123 | −0.120/−0.129 |
| nad3 | 38.4/39.6 | 35.8/34.2 | 10.8/11.2 | 15.1/15.0 | 74.2/73.8 | 0.034/0.074 | −0.165/−0.146 |
| nad4 | 26.4/25.8 | 49.3/50.6 | 14.6/13.9 | 9.6/10.3 | 75.7/75.8 | −0.318/−0.335 | 0.137/0.149 |
| nad4L | 26.0/24.7 | 51.4/49.7 | 12.8/14.2 | 9.7/11.5 | 77.4/74.3 | −0.327/−0.336 | 0.323/0.108 |
| nad5 | 26.5/24.6 | 49.3/49.1 | 14.6/15.4 | 9.6/11.0 | 75.8/73.6 | −0.301/−0.333 | 0.205/0.169 |
| nad6 | 39.8/39.4 | 38.9/40.8 | 8.2/7.9 | 13.1/11.9 | 78.7/80.2 | 0.011/−0.0188 | −0.232/−0.200 |
| cox1 | 34.8/34.1 | 32.9/32.9 | 14.7/15.2 | 17.5/17.8 | 67.7/67.0 | 0.027/0.018 | −0.091/−0.018 |
| cox2 | 39.8/39.8 | 29.6/30.2 | 13.0/13.3 | 17.7/16.8 | 69.4/70.0 | 0.146/0.137 | −0.154/−0.118 |
| cox3 | 34.1/34.3 | 34.0/33.8 | 15.0/14.6 | 17.0/17.2 | 68.1/68.2 | 0.002/0.007 | −0.063/−0.084 |
| cytb | 34.5/34.8 | 35.1/35.4 | 12.8/12.1 | 17.7/17.7 | 69.6/70.2 | −0.009/−0.009 | −0.162/−0.187 |
| atp6 | 38.1/37.2 | 35.1/35.3 | 10.7/10.7 | 16.1/16.9 | 73.2/72.4 | 0.041/0.027 | −0.204/−0.226 |
| atp8 | 40.3/42.8 | 32.1/37.1 | 9.4/8.2 | 18.2/11.9 | 72.4/79.9 | 0.113/0.071 | −0.318/−0.188 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, D.; Zhang, H.; Fu, S.; Wang, Y.; Zhao, W.; Zhao, Q. The Complete Mitochondrial Genomes of Aelia sibirica and A. fieberi (Hemiptera, Pentatomidae), and Phylogenetic Implications. Diversity 2024, 16, 111. https://doi.org/10.3390/d16020111
Liu D, Zhang H, Fu S, Wang Y, Zhao W, Zhao Q. The Complete Mitochondrial Genomes of Aelia sibirica and A. fieberi (Hemiptera, Pentatomidae), and Phylogenetic Implications. Diversity. 2024; 16(2):111. https://doi.org/10.3390/d16020111
Chicago/Turabian StyleLiu, Dajun, Hufang Zhang, Shuhui Fu, Yating Wang, Wanqing Zhao, and Qing Zhao. 2024. "The Complete Mitochondrial Genomes of Aelia sibirica and A. fieberi (Hemiptera, Pentatomidae), and Phylogenetic Implications" Diversity 16, no. 2: 111. https://doi.org/10.3390/d16020111
APA StyleLiu, D., Zhang, H., Fu, S., Wang, Y., Zhao, W., & Zhao, Q. (2024). The Complete Mitochondrial Genomes of Aelia sibirica and A. fieberi (Hemiptera, Pentatomidae), and Phylogenetic Implications. Diversity, 16(2), 111. https://doi.org/10.3390/d16020111