
Phenotypic and Molecular Characterization of Yeast Diversity Associated to Postharvest Fermentation Process of Coffee Fruits in Southern Ecuador

 ,
,  ,
,  , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
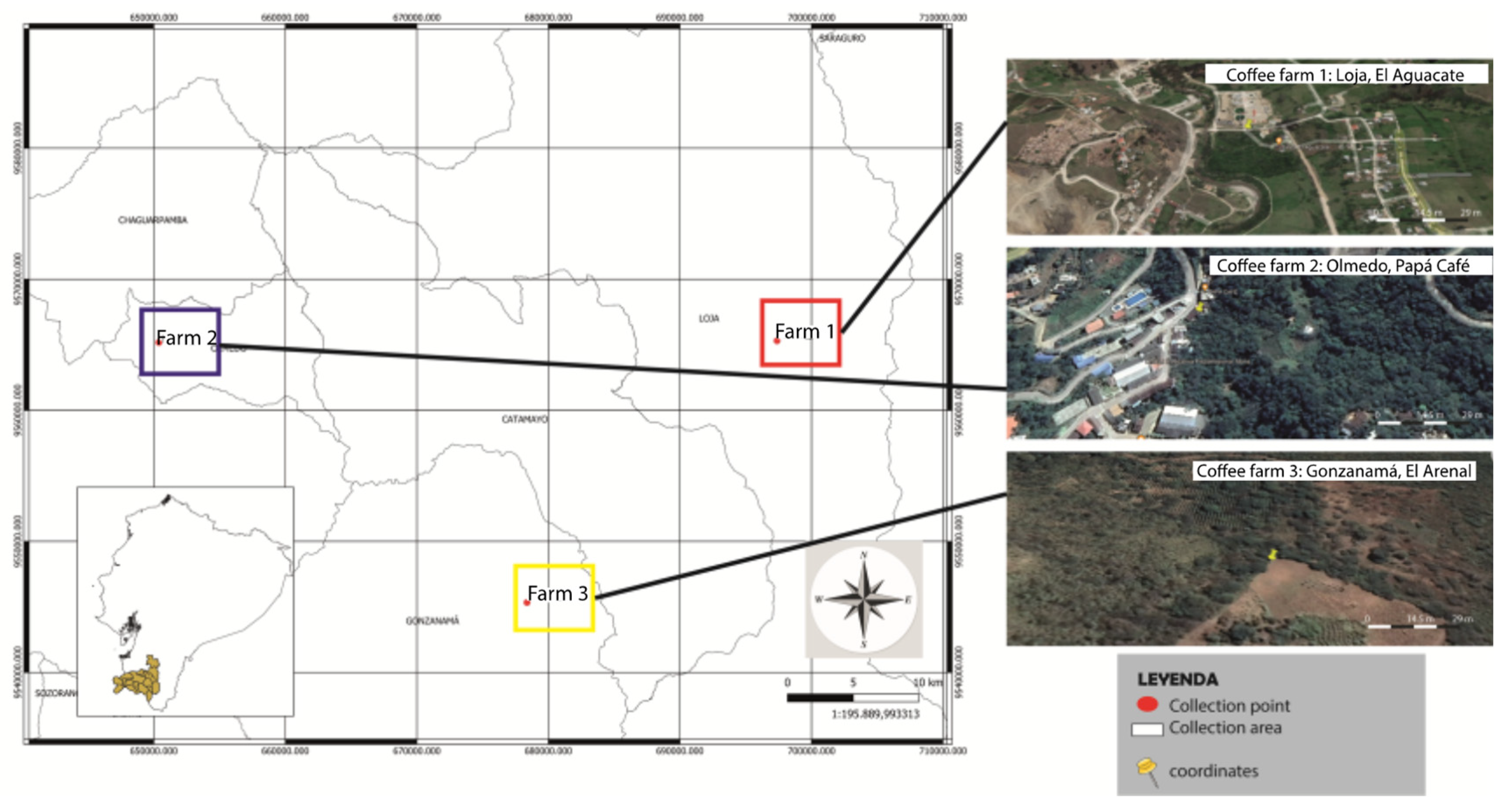
2.1. Sampling
2.2. Postharvest Fermentation Process
2.3. Yeast Isolation
2.4. Phenotypic Characterization of Pure Strain Sampling
2.5. Statistical Analysis of Ordination and Analysis of Isolates by Farms
2.6. DNA Extraction, Amplification and Sequencing
2.7. Phylogenetic Analysis
2.8. Strain Preservation
3. Results
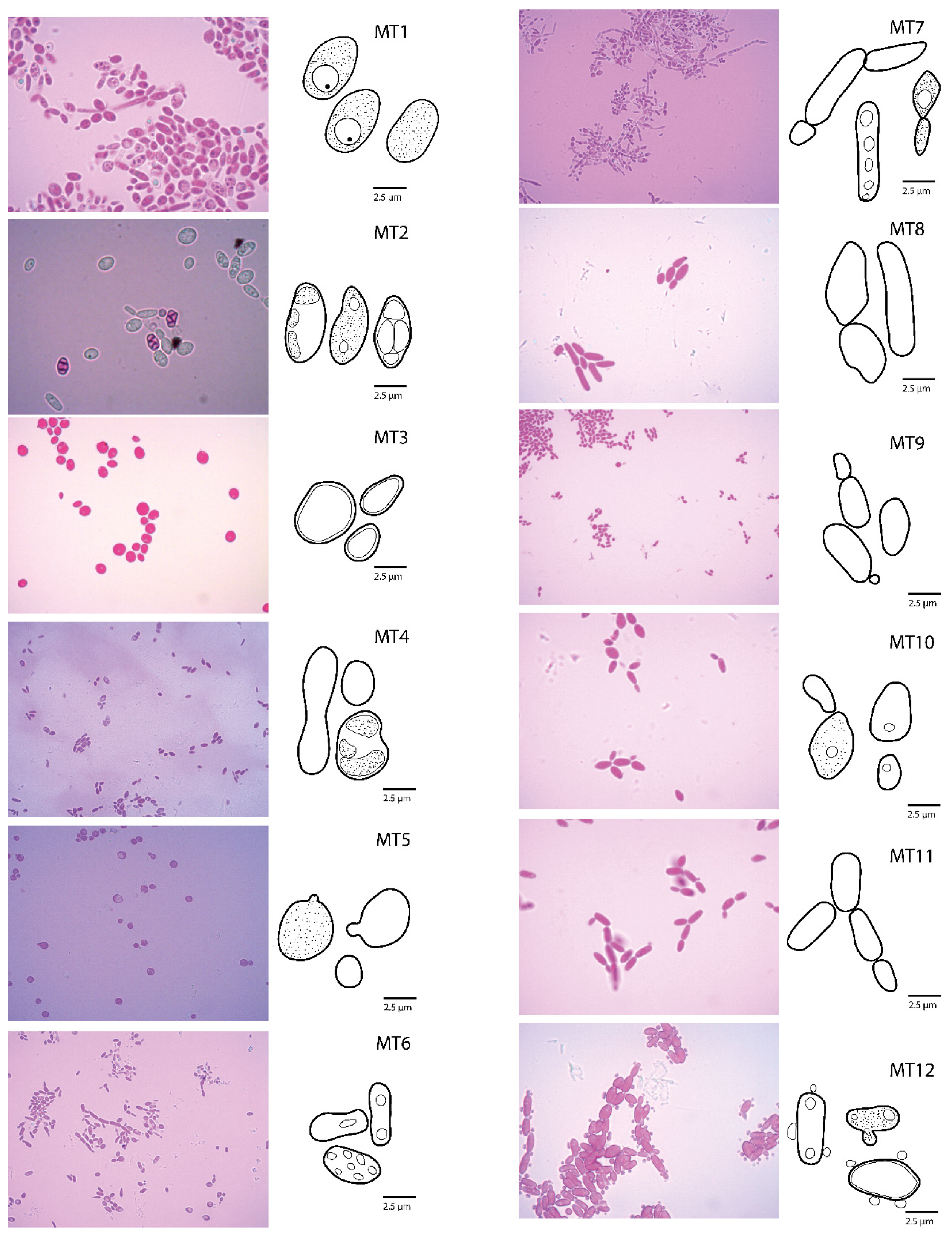
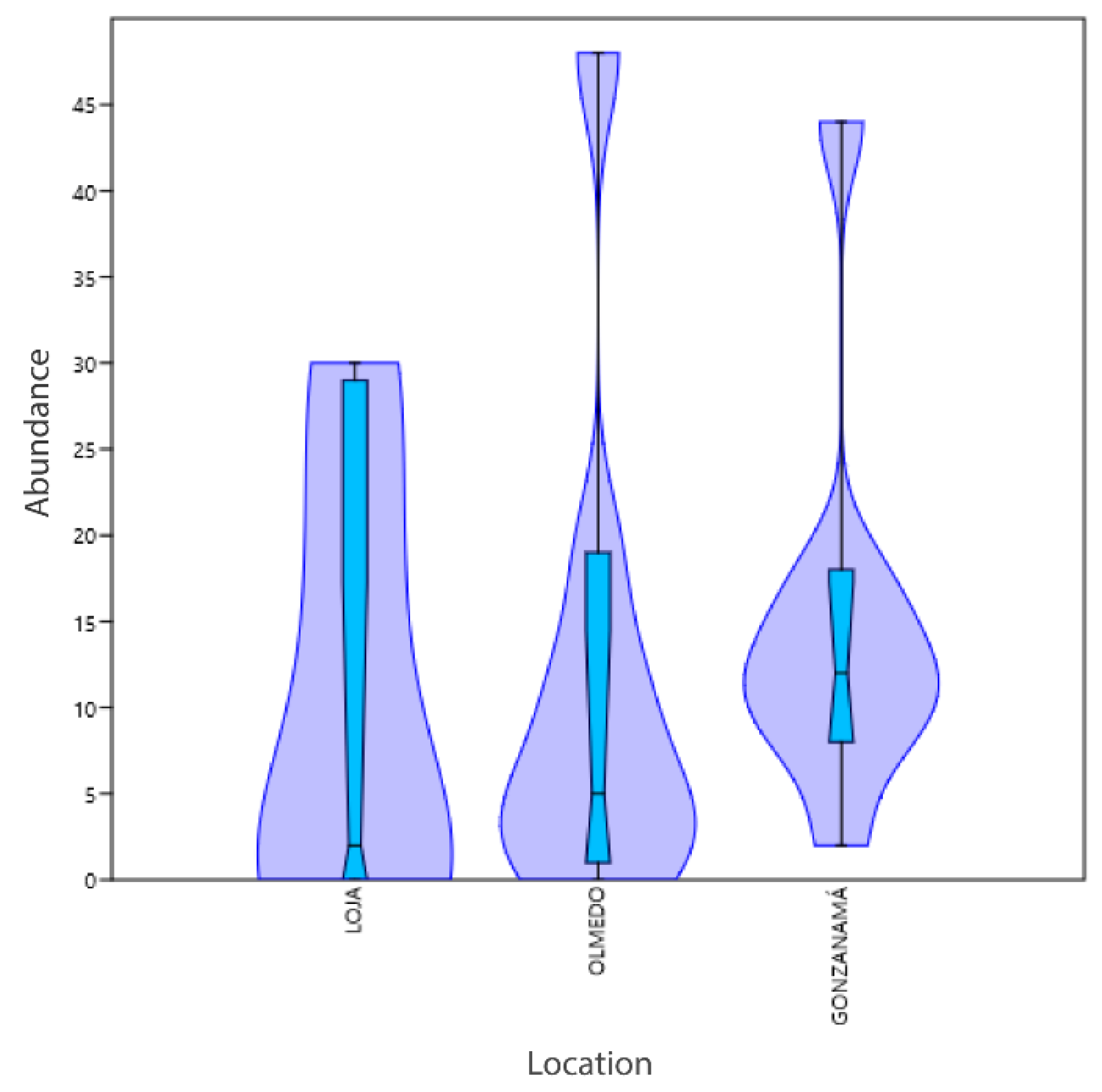
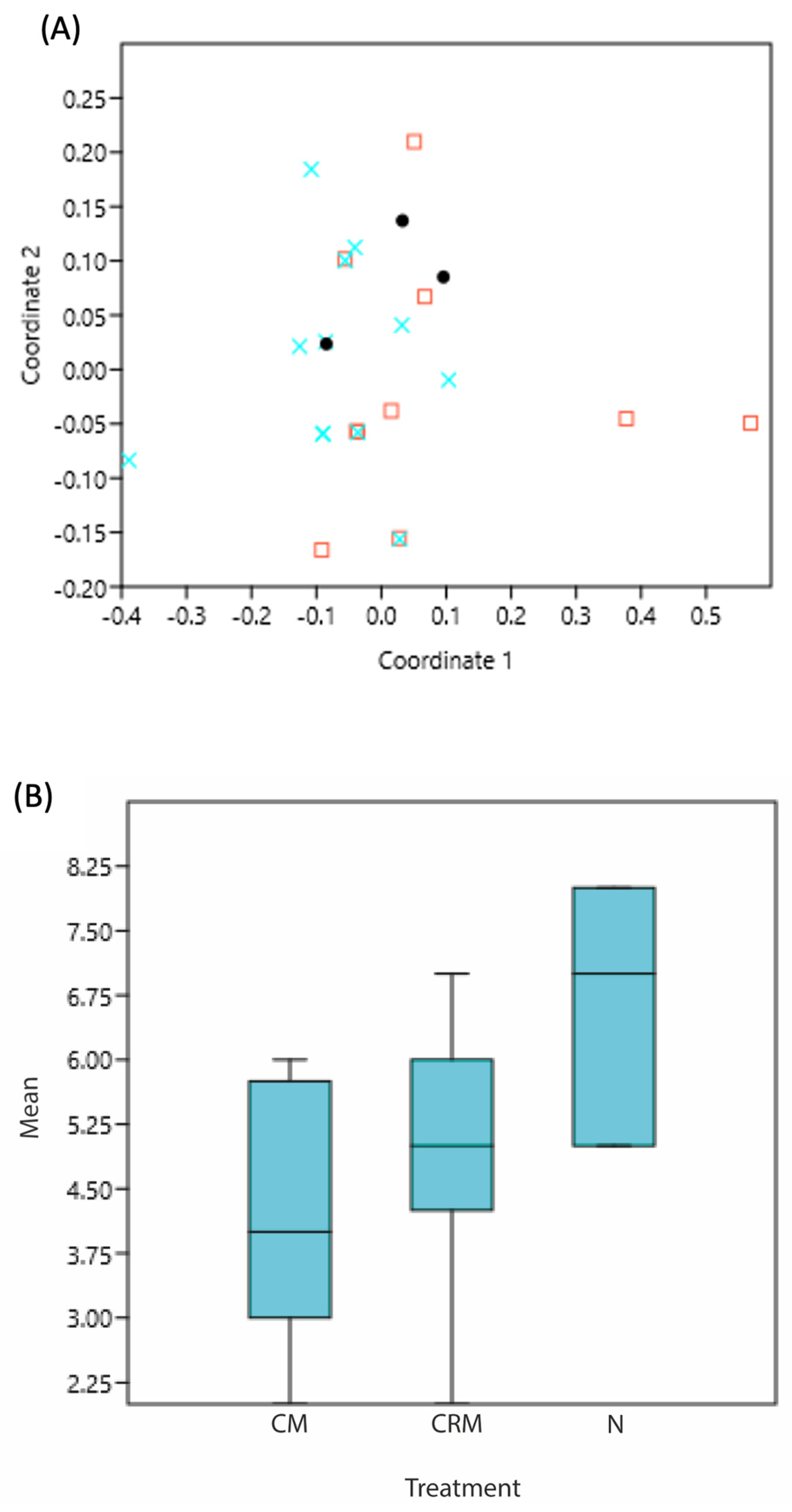
3.1. Phenotyping and Diversity
3.2. Molecular Species Definition
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barrezueta-Unda, S.; Moreira Blacio, W.; Quezada Abad, C. Análisis del cacao y café ecuatoriano desde su cadena de valor en el periodo 2010–2015. Rev. Científica Agroecosistemas 2018, 6, 6–17. [Google Scholar]
- Mercedes, M.S.; Otiniano, A.J.; Borjas Ventura, R. Sustentabilidad de fincas productoras de café en Jipijapa (Manabí, Ecuador). Saber Hacer-Rev. Fac. Ing. USIL 2016, 3, 23–25. [Google Scholar]
- Duicela, G.L.A.; Andrade, M.J.; Farfán, T.D.S.; Velásquez, C.S.D. Calidad organoléptica, métodos de beneficio y cultivares de café robusta (Coffea canephora Pierre ex Froehner) en la amazonía del Ecuador. Rev. Iberoam. Tecnol. Postcosecha 2018, 19, 2. [Google Scholar]
- Elhalis, H.; Cox, J.; Frank, D.; Zhao, J. The crucial role of yeasts in the wet fermentation of coffee beans and quality. Int. J. Food Microbiol. 2020, 333, 108796. [Google Scholar] [CrossRef] [PubMed]
- Guzzon, R.; Malacarne, M.; Larcher, R.; Franciosi, E.; Toffanin, A. The impact of grape processing and carbonic maceration on the microbiota of early stages of winemaking. J. Appl. Microbiol. 2020, 128, 209–224. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Sun, J.; Lassabliere, B.; Yu, B.; Liu, S.Q. Coffee flavour modification through controlled fermentations of green coffee beans by Saccharomyces cerevisiae and Pichia kluyveri: Part I. Effects from individual yeasts. Food Res. Int. 2020, 136, 109588. [Google Scholar] [CrossRef]
- Arastehfar, A.; Fang, W.; Pan, W.; Lackner, M.; Liao, W.; Badiee, P.; Boekhout, T. YEAST PANEL multiplex PCR for identification of clinically important yeast species: Stepwise diagnostic strategy, useful for developing countries. Diagn. Microbiol. Infect. Dis. 2019, 93, 112–119. [Google Scholar] [CrossRef]
- Yurkov, A.M.; Röhl, O.; Carvalho, C.; Maldonado, C.; Sampaio, J.P. Local climatic conditions constrain soil yeast diversity patterns in mediterranean forests, woodlands and scrubbiome. FEMS Yeast Res. 2015, 16, fov103. [Google Scholar] [CrossRef]
- Toro, M.; Oro, N.; Vega, A.; Maturano, Y.; Nally, M.; Fernández, E.; Pucheta, E.; Vázquez, F. Diversidad de levaduras en canopias y suelos asociados con Bulnesia retama y Larrea divaricata. Rev. Argent. De Microbiol. 2005, 37, 209–213. [Google Scholar]
- Kurtzman, C.; Robnett, C. Phylogenetic relationships among yeasts of the ‘Saccharomyces complex’ determined from multigene sequence analyses. FEMS Yeast Res. 2003, 3, 417–432. [Google Scholar] [CrossRef]
- Vásquez, C.J.A.; Ramirez, C.M.; Monsalve, F.Z.I. Actualización en caracterización molecular de Levaduras de Interés Industrial. Rev. Colomb. Biotecnol. 2016, 18, 129. [Google Scholar] [CrossRef]
- Nerva, L.; Turina, M.; Zanzotto, A.; Gardiman, M.; Gaiotti, F.; Gambino, G.; Chitarra, W. Isolation, molecular characterization and virome analysis of culturable wood fungal endophytes in esca symptomatic and asymptomatic grapevine plants. Environ. Microbiol. 2019, 21, 2886–2904. [Google Scholar] [CrossRef] [PubMed]
- Schoch, C.L.; Seifert, K.A.; Huhndorf, S.; Robert, V.; Spouge, J.L.; Levesque, C.A. Nuclear ribosomal internal transcribed spacer (ITS) region as a universal DNA barcode marker for fungi. Proc. Natl. Acad. Sci. USA 2012, 109, 6241–6246. [Google Scholar] [CrossRef] [PubMed]
- Segura, G.L.E.; Kirchmayr, M.R.; Flores, B.E.P.; Gschaedler, M.A.C. PCR-RFLP de las regiones ITS-5.8S como herramienta de identificación de levaduras: Ventajas y desventajas. e-Gnosis 2010, 8, 1–12. [Google Scholar]
- Ooi, T.S.; Ting, A.S.Y.; Siow, L.F. Influence of selected native yeast starter cultures on the antioxidant activities, fermentation index and total soluble solids of Malaysia cocoa beans: A simulation study. LWT 2020, 122, 108977. [Google Scholar] [CrossRef]
- Wei, J.; Zhang, Y.; Qiu, Y.; Guo, H.; Ju, H.; Wang, Y.; Yuan, Y.; Yue, T. Chemical composition, sensorial properties, and aroma-active compounds of ciders fermented with Hanseniaspora osmophila and Torulaspora quercuum in co- and sequential fermentations. Food Chem. 2020, 306, 12–56. [Google Scholar] [CrossRef]
- Evangelista, S.R.; Silva, C.F.; da Cruz Miguel, M.G.P.; de Souza, C.C.; Pinheiro, A.C.M.; Duarte, W.F.; Schwan, R.F. Improvement of coffee beverage quality by using selected yeasts strains during the fermentation in dry process. Food Res. Int. 2020, 61, 183–195. [Google Scholar] [CrossRef]
- Santos, D.K.F.; Resende, A.H.M.; de Almeida, D.G.; Soares da Silva, R.C.F.; Rufino, R.D.; Luna, J.M.; Banat, I.M.; Sarubbo, L.A. Candida lipolytica UCP0988 Biosurfactant: Potential as a Bioremediation Agent and in Formulating a Commercial Related Product. Front. Microbiol. 2017, 8, 767. [Google Scholar] [CrossRef]
- Sepúlveda, W.S.; Ureta, I.; Sepulveda-Sepúlveda, A. Profile and preference of Ecuadorian consumers for quality attributes in coffee production. Coffee Sci. 2016, 11, 298–307. [Google Scholar]
- Aguirre, L.A.; Rodríguez, Z.; Boucourt, R.; Saca, V.; Salazar, R.; Jiménez, M. Effect of whey on solid state fermentation of coffee (Coffea arabica L.) pulp for feeding ruminants. Rev. Cuba. De Cienc. Agrícola 2018, 52, 303–312. [Google Scholar]
- Jumbo, N.; Martínez, M. Efecto del Uso de Saccharomyces Cerevisiae Bajo Condiciones Fermentativas en la Calidad de Taza del Café (Coffea arabica L.) en el Cantón Loja; Trabajo de titulación, Universidad Nacional de Loja: Loja, Ecuador, 2021. [Google Scholar]
- Arcila Pulgarín, J. Crecimiento y Desarrollo de la Planta de Café. Sistemas de Producción de Café en Colombia; CENICAFE: Chinchiná, Colombia, 2007; pp. 21–60. [Google Scholar]
- Delgado-Ospina, J.; Triboletti, S.; Alessandria, V.; Serio, A.; Sergi, M.; Paparella, A.; Rantsiou, K.; Chaves-López, C. Functional Biodiversity of Yeasts Isolated from Colombian Fermented and Dry Cocoa Beans. Microorganisms 2020, 8, 1086. [Google Scholar] [CrossRef]
- Versalovic, J. (Ed.) Manual of Clinical Microbiology; American Society for Microbiology Press: Washington, DC, USA, 2011; Volume 1. [Google Scholar]
- Rodrigues, C.B.; Barabasz, R.F.; Silva, R.H.D.; Sustakowski, M.C.; Kuhn, O.J.; Carvalho, J.C.; Stangarlin, J.R. Yeast Potential for the Biological Control of Colletotrichum musae. J. Agric. Sci. 2020, 12, 301. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Gardes, M.; Bruns, T.D. ITS primers with enhanced specificity for basidiomycetes-application to the identification of mycorrhizae and rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Leaw, S.N.; Chang, H.C.; Sun, H.F.; Barton, R.; Bouchara, J.P.; Chang, T.C. Identification of medically important yeast species by sequence analysis of the internal transcribed spacer regions. J. Clin. Microbiol. 2006, 44, 693–699. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics, 1st ed.; Oxford University Press: New York, NY, USA, 2000; pp. 154–196. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. Mega X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Ocares, P.; Yocelyn; Castro, F.; Jean, F. Preservación de microorganismos por congelación. Inst. Investig. Agropecu. 2020, 428, 119–134. [Google Scholar]
- Wang, F.; Cui, M.; Liu, H.; Li, X.; Yu, J.; Huang, Y.; Liu, Y. Characterization and identification of a fraction from silver carp (Hypophthalmichthys molitrix) muscle hydrolysates with cryoprotective effects on yeast. LWT 2021, 137, 110388. [Google Scholar] [CrossRef]
- Xiao, M.; Wang, H.; Lu, J.; Chen, S.C.; Kong, F.; Ma, X.J.; Xu, Y.C. Three clustered cases of candidemia caused by Candida quercitrusa and mycological characteristics of this novel species. J. Clin. Microbiol. 2014, 52, 3044–3048. [Google Scholar] [CrossRef]
- Eddouzi, J.; Lohberger, A.; Vogne, C.; Manai, M.; Sanglard, D. Identification and antifungal susceptibility of a large collection of yeast strains isolated in Tunisian hospitals. Med. Mycol. 2013, 51, 737–746. [Google Scholar] [CrossRef][Green Version]
- Ben Taheur, F.; Mansour, C.; Ben Jeddou, K.; Machreki, Y.; Kouidhi, B.; Abdulhakim, J.A.; Chaieb, K. Aflatoxin B1 degradation by microorganisms isolated from kombucha culture. Toxicon 2020, 179, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.S.; Arruda, L.M.; Xavier, P.L.; Ramírez, M.X.; da Silveira, F.A.; Santana, W.C.; da Silva, P.H.; Fietto, L.G.; Eller, M.R. Selection of yeasts from bee products for alcoholic beverage production. Braz. J. Microbiol. 2020, 51, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Bourret, T.B.; Grove, G.G.; Vandemark, G.J.; Henick-Kling, T.; Glawe, D.A. Diversity and molecular determination of wild yeasts in a central Washington State vineyard. N. Am. Fungi 2013, 8, 1–32. [Google Scholar] [CrossRef]
- Zabukovec, P.; Čadež, N.; Čuš, F. Isolation and Identification of Indigenous Wine Yeasts and Their Use in Alcoholic Fermentation. Food Technol. Biotechnol. 2020, 58, 337–347. [Google Scholar] [CrossRef]
- Stavrou, A.A.; Lackner, M.; Lass-Flörl, C.; Boekhout, T. The changing spectrum of saccharomycotina yeasts causing candidemia: Phylogeny mirrors antifungal susceptibility patterns for azole drugs and amphothericin B. FEMS Yeast Res. 2019, 19, foz037. [Google Scholar] [CrossRef]
- Li, D.; Han, T.; Liao, J.; Hu, X.; Xu, S.; Tian, K.; Gu, X.; Cheng, K.; Li, Z.; Hua, H.; et al. Oridonin, a promising ent-kaurane diterpenoid lead compound. Int. J. Mol. Sci. 2016, 17, 1395. [Google Scholar] [CrossRef] [PubMed]
- Arteau, M.; Labrie, S.; Roy, D. Terminal-restriction fragment length polymorphism and automated ribosomal intergenic spacer analysis profiling of fungal communities in Camembert cheese. Int. Dairy J. 2010, 20, 545–554. [Google Scholar] [CrossRef]
- Vasques, D.T.; Ebihara, A.; Hirai, R.Y.; Prado, J.; Motomi, I. Phylogeny of Hymenophyllum subg. Mecodium (Hymenophyllaceae), with special focus on the diversity of the Hymenophyllum polyanthos species complex. Plant Syst. Evol. 2019, 305, 811–825. [Google Scholar] [CrossRef]
- Irinyi, L.; Serena, C.; Garcia-Hermoso, D.; Arabatzis, M.; Desnos-Ollivier, M.; Vu, D.; Meyer, W. International Society of Human and Animal Mycology (ISHAM)-ITS reference DNA barcoding database—The quality controlled standard tool for routine identification of human and animal pathogenic fungi. Med. Mycol. 2015, 53, 313–337. [Google Scholar] [CrossRef]
- Wu, Z.W.; Robert, V.; Bai, F.Y. Genetic diversity of the Pichia membranifaciens strains revealed from rRNA gene sequencing and electrophoretic karyotyping, and the proposal of Candida californica comb. nov. FEMS Yeast Res. 2006, 6, 305–311. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Golić, N.; Čadež, N.; Terzić-Vidojević, A.; Šuranská, H.; Beganović, J.; Lozo, J.; Topisirović, L. Evaluation of lactic acid bacteria and yeast diversity in traditional white pickled and fresh soft cheeses from the mountain regions of Serbia and lowland regions of Croatia. Int. J. Food Microbiol. 2013, 166, 294–300. [Google Scholar] [CrossRef] [PubMed]
- Vu, D.; Groenewald, M.; Szöke, S.; Cardinali, G.; Eberhardt, U.; Stielow, B.; Robert, V. DNA barcoding analysis of more than 9 000 yeast isolates contributes to quantitative thresholds for yeast species and genera delimitation. Stud. Mycol. 2016, 85, 91–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.M.; Xu, J.; Wang, H.; Li, J.; Bai, F.Y. Torulaspora quercuum sp. nov. and Candida pseudohumilis sp. nov., novel yeasts from human and forest habitats. FEMS Yeast Res. 2009, 9, 1322–1326. [Google Scholar] [CrossRef] [PubMed]
- Zha, Y.; Hossain, A.H.; Tobola, F.; Sedee, N.; Havekes, M.; Punt, P.J. Pichia anomala 29X: A resistant strain for lignocellulosic biomass hydrolysate fermentation. FEMS Yeast Res. 2013, 13, 609–617. [Google Scholar] [CrossRef]
- Tanahashi, M.; Hawes, C. The presence of a mycangium in European Sinodendron cylindricum (Coleoptera: Lucanidae) and the associated yeast symbionts. J. Insect Sci. 2016, 16, 76. [Google Scholar] [CrossRef] [PubMed]
- Glushakova, A.M.; Maximova, I.A.; Kachalkin, A.V.; Yurkov, A.M. Ogataea cecidiorum sp. nov., a methanol-assimilating yeast isolated from galls on willow leaves. Antonie Van Leeuwenhoek 2010, 98, 93–101. [Google Scholar] [CrossRef]
- Knop, M. Yeast cell morphology and sexual reproduction–A short overview and some considerations. Comptes Rendus Biol. 2011, 334, 599–606. [Google Scholar] [CrossRef]
- De Melo Pereira, G.V.; Soccol, V.T.; Pandey, A.; Medeiros, A.B.P.; Lara, J.M.R.A.; Gollo, A.L.; Soccol, C.R. Isolation, selection and evaluation of yeasts for use in fermentation of coffee beans by the wet process. Int. J. Food Microbiol. 2014, 188, 60–66. [Google Scholar] [CrossRef]
- Lopes, M.R.; Santos, A.R.O.; Moreira, J.D.; Santa-Brígida, R.; Martins, M.B.; Pinto, F.O.; Valente, P.; Morais, P.B.; Jacques, N.; Grondin, C.; et al. Kurtzmaniella hittingeri f.a., sp. nov., isolated from rotting wood and fruits, and transfer of three Candida species to the genus Kurtzmaniella as new combinations. Int. J. Syst. Evol. Microbiol. 2019, 69, 1504–1508. [Google Scholar] [CrossRef]
- De Carvalho Neto, D.P.; De Melo Pereira, G.V.; Tanobe, V.O.; Thomaz, S.V.G.; Da Silva, B.J.; Rodrigues, C.; Soccol, C.R. Yeast diversity and physicochemical characteristics associated with coffee bean fermentation from the Brazilian Cerrado Mineiro region. Fermentation 2017, 3, 11. [Google Scholar] [CrossRef]
- Kurtzman, C.; Fell, J.W.; Boekhout, T. The Yeasts a Taxonomic Study, 5th ed.; Elsevier Science: Madrid, Spain, 2011; pp. 543–562. [Google Scholar]
- Rodrigues, J.D.O.; Höfling, J.F.; Tavares, F.C.A.; Duarte, K.M.R.; Gonçalves, R.B.; Azevedo, R.A.D. Evaluation of biochemical and serological methods to identify and clustering yeast cells of oral Candida species by CHROMagar test, SDS-PAGE and ELISA. Braz. J. Biol. 2004, 64, 317–326. [Google Scholar] [CrossRef]
- Martins, P.M.M.; Ribeiro, L.S.; Miguel, M.G.D.C.P.; Evangelista, S.R.; Schwan, R.F. Production of coffee (Coffea arabica) inoculated with yeasts: Impact on quality. J. Sci. Food Agric. 2019, 99, 5638–5645. [Google Scholar] [CrossRef] [PubMed]
- Op De Beeck, M.; Lievens, B.; Busschaert, P.; Declerck, S.; Vangronsveld, J.; Colpaert, J.V. Comparison and validation of some ITS primer pairs useful for fungal metabarcoding studies. PLoS ONE 2014, 9, 6–29. [Google Scholar] [CrossRef]
- Armijos, C.; Valarezo, E.; Cartuche, L.; Zaragoza, T.; Finzi, P.V.; Mellerio, G.G.; Vidari, G. Chemical composition and antimicrobial activity of Myrcianthes fragrans essential oil, a natural aromatizer of the traditional Ecuadorian beverage colada morada. J. Ethnopharmacol. 2018, 225, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Shapaval, V.; Brandenburg, J.; Blomqvist, J.; Tafintseva, V.; Passoth, V.; Sandgren, M.; Kohler. Biochemical profiling, prediction of total lipid content and fatty acid profile in oleaginous yeasts by FTIR spectroscopy. Biotechnol. Biofuels 2019, 12, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Zhou, Y.; Zeng, K. Effect of Pichia membranaefaciens on ROS metabolism and postharvest disease control in citrus fruit. Crop Prot. 2013, 53, 96–102. [Google Scholar] [CrossRef]
- Vicente, J.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S. High potential of Pichia kluyveri and other Pichia species in wine technology. Int. J. Mol. Sci. 2021, 22, 1196. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Canton | Farms by Sector | Latitude and Longitude | Altitude (m.a.s.l.) | No Sample Code | Postharvest Fermentation Process |
|---|---|---|---|---|---|
| Loja | El Aguacate | 3°55.848′ S 79°13.340′ W | 2010 | A | Natural |
| AC | Carbonic maceration with CO2 atmosphere at room temperature | ||||
| AF | Carbonic refrigerated maceration with CO2 atmosphere up to 10 °C | ||||
| Olmedo | Papá Café | 3°55.960′ S 79°38.729′ W | 1230 | B | Natural |
| BMF | Carbonic maceration with CO2 atmosphere at room temperature | ||||
| BMC | Carbonic refrigerated maceration with CO2 atmosphere up to 10 °C | ||||
| Gonzanamá | El Arenal | 4°06′02.0″ S 79°23′40.4″ W | 1500 m | M | Natural |
| MC | Carbonic maceration with CO2 atmosphere at room temperature | ||||
| MF | Carbonic refrigerated maceration with CO2 atmosphere up to 10 °C |
| Sum of Squares | Df | Mean Square F | F | p Value | ||
|---|---|---|---|---|---|---|
| Richness according to the type of fermentation | Between groups | 15.7407 | 2 | 7.87037 | 4.077 | 0.02991 |
| Within groups | 46.3333 | 24 | 1.93056 | |||
| Total | 62.0741 | 26 | 0.3307 | |||
| Richness by location | Between groups | 11.6296 | 2 | 5.81481 | 2.767 | 0.08295 |
| Within groups | 50.4444 | 24 | 2.10185 | |||
| Total | 62.0741 | 26 | 0.08731 | |||
| Species | Collection Number | Morphotype Referential | GenBank Accession Number | Country | Literature Reference |
|---|---|---|---|---|---|
| Kurztmaniella quercitrusa | CM1L1 | MT7 | OR145791 | Ecuador | This study |
| Candida quercitrusa | HA 1669 | - | AM160627 | Austria | Xiao [34] |
| Kurztmaniella quercitrusa | BMF2L3 | MT7 | OR145790 | Ecuador | This study |
| Hannseniaspora opuntiae | MF3L2 | MT8 | OR145783 | Ecuador | This study |
| Hannseniaspora opuntiae | JEY258 | - | KC111445 | Tunisia | Eddouzi et al. [35] |
| Hannseniaspora opuntiae | KOT1 | - | KY849378 | Tunisia | Ben Taheur [36] |
| Hannseniaspora opuntiae | SM10UFAM | - | MN268780 | Brazil | Silva et al. [37] |
| Hanseniaspora uvarum | AC3L1 | MT9 | OR145784 | Ecuador | This study |
| Hanseniaspora uvarum | P43C012 | - | JX188164 | USA | Bourret et al. [38] |
| Hanseniaspora uvarum | Mer383 | - | MT734682 | Slovenia | Zabukovec et al. [39] |
| Krutzmaniella quercitrusa | CBS 4412 | - | MK394107 | Netherlands | Stavrou et al. [40] |
| Krutzmaniella quercitrusa | LHCT1 | - | KX961193. | China | Li et al. [41] |
| Nuerospora crassa | AR14R | - | OK148118 | India | Direct submission |
| Nuerospora crassa | I1-3 | - | MH507014 | China | Direct submission |
| Nuerospora crassa | I1-3 | - | MK064500 | China | Direct submission |
| Pichia fermentans | AF2L7 | MT4 | OR145756 | Ecuador | This study |
| Pichia fermentans | CM1L6 | MT4 | OR145758 | Ecuador | This study |
| Pichia fermentans | AC3L10 | MT4 | OR145755 | Ecuador | This study |
| Pichia fermentans | AC4L2 | MT4 | OR145757 | Ecuador | This study |
| Pichia fermentans | HR-1 DS1308 | - | KM029994 | USA | Direct submission |
| Pichia fermentans | HA-1 | - | KU820951 | Republic of Korea | Direct submission |
| Pichia fermentans | ATCC 10651 | - | GQ458040 | Canadá | Arteau et al. [42] |
| Pichia fermentans | CBS 187 | - | MK394169 | Netherlands | Stavrou et al. [40] |
| Pichia kluyveri | BMC4L9 | MT1 | OR145759 | Ecuador | This study |
| Pichia kluyveri | AF2L3 | MT10 | OR145761 | Ecuador | This study |
| Pichia kluyveri | BMF4L1 | MT10 | OR145762 | Ecuador | This study |
| Pichia kluyveri | AF2L1 | MT10 | OR145760 | Ecuador | This study |
| Pichia kluyveri | P25B004 | - | JX188197 | USA | Bourret et al. [38] |
| Pichia kluyveri | H12 | - | MN266784 | Japón | Vasques et al. [43] |
| Pichia kluyveri | KBP:Y-4954 | - | MG367291 | Moscow | Direct Submission |
| Pichia kluyveri | PMM10-1033666L | - | KP132502 | Australia | Irinyi et al. [44] |
| Pichia kluyveri | CBS 188 | - | MK394165 | Netherlands | Stavrou et al. [40] |
| Pichia membranifaciens | M2L7 | MT6 | OR145770 | Ecuador | This study |
| Pichia membranifaciens | MF4L8 | MT6 | OR145772 | Ecuador | This study |
| Pichia membranifaciens | BMF1L6 | MT6 | OR145774 | Ecuador | This study |
| Pichia membranifaciens | AC1L5 | MT10 | OR145767 | Ecuador | This study |
| Pichia membranifaciens | BMF1L2 | MT6 | OR145769 | Ecuador | This study |
| Pichia membranifaciens | BMF1L5 | MT6 | OR145775 | Ecuador | This study |
| Pichia membranifaciens | BMF4L9 | MT12 | OR145773 | Ecuador | This study |
| Pichia membranifaciens | AC3L3 | MT6 | OR145766 | Ecuador | This study |
| Pichia membranifaciens | MF1L3 | MT6 | OR145776 | Ecuador | This study |
| Pichia membranifaciens | MF1L7 | MT6 | OR145777 | Ecuador | This study |
| Pichia membranifaciens | AC2L2 | MT11 | OR145764 | Ecuador | This study |
| Pichia membranifaciens | AF2L6 | MT6 | OR145765 | Ecuador | This study |
| Pichia membranifaciens | P43C010 | - | JX188208 | USA | Bourret et al. [38] |
| Pichia membranifaciens | CBS 82 | - | DQ198951 | China | Wu et al. [45] |
| Pichia membranifaciens | CBS 107 | - | MK394163 | Netherlands | Stavrou et al. [40] |
| Pichia membranifaciens | MC1L9 | MT6 | OR145771 | Ecuador | This study |
| Pichia membranifaciens | BMF1L9 | MT6 | OR145768 | Ecuador | This study |
| Pichia membranifaciens | MF4L5 | MT12 | OR145763 | Ecuador | This study |
| Torulaspora delbrueckii | MC4L2 | MT2 | OR145788 | Ecuador | This study |
| Torulaspora delbrueckii | MC1L1 | MT2 | OR145787 | Ecuador | This study |
| Torulaspora delbrueckii | MC2L1 | MT2 | OR145789 | Ecuador | This study |
| Torulaspora delbrueckii | B2/III/17 | - | HE799671 | Slovenia | Golic et al. [46] |
| Torulaspora delbrueckii | CBS 1146 | - | MK394138 | Netherlands | Stavrou et al. [40] |
| Torulaspora quercuum | BMF4L7 | MT3 | OR145786 | Ecuador | This study |
| Torulaspora quercuum | AF2L8 | MT3 | OR145785 | Ecuador | This study |
| Torulaspora quercuum | CBS:11403 | - | KY105673 | Netherlands | Vu et al. [47] |
| Torulaspora quercuum | CGMCC AS 2 | - | NR137029 | China | Wang et al. [48] |
| Torulaspora quercuum | XZ-46A | - | FJ888525 | China | Wang et al. [48] |
| Torulaspora quercuum | XZ19100-5 | - | MW710147 | China | Direct Submission |
| Wickerhamomyces anomalus | BMC2L7 | MT5 | OR145778 | Ecuador | This study |
| Wickerhamomyces anomalus | AC4L9 | MT5 | OR145782 | Ecuador | This study |
| Wickerhamomyces anomalus | BMF2L7 | MT5 | OR145779 | Ecuador | This study |
| Wickerhamomyces anomalus | MF1L9 | MT5 | OR145780 | Ecuador | This study |
| Wickerhamomyces anomalus | MC3L1 | MT5 | OR145781 | Ecuador | This study |
| Wickerhamomyces anomalus | CBS 5759 | - | MK394130 | Netherlands | Stavrou et al. [40] |
| Wickerhamomyces anomalus | I 29 | - | HF952836 | Netherlands | Zha et al. [49] |
| Wickerhamomyces anomalus | VIT-ASN01 | - | KX253664 | India | Direct Submission |
| Wickerhamomyces anomalus | DBL01s1 | - | LC120363 | Japan | Tanahashi and Hawes [50] |
| Wickerhamomyces anomalus | iL-51-2 | - | FN868149 | Germany | Glushakova et al. [51] |
| Wickerhamomyces anomalus | CBS 5759 | - | MH545921 | Netherlands | Stavrou et al. [40] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Apolo, D.; Fernández, J.M.; Benítez, Á.; Figueroa, J.G.; Estrada, K.; Cruz, D. Phenotypic and Molecular Characterization of Yeast Diversity Associated to Postharvest Fermentation Process of Coffee Fruits in Southern Ecuador. Diversity 2023, 15, 984. https://doi.org/10.3390/d15090984
Apolo D, Fernández JM, Benítez Á, Figueroa JG, Estrada K, Cruz D. Phenotypic and Molecular Characterization of Yeast Diversity Associated to Postharvest Fermentation Process of Coffee Fruits in Southern Ecuador. Diversity. 2023; 15(9):984. https://doi.org/10.3390/d15090984
Chicago/Turabian StyleApolo, David, José Miguel Fernández, Ángel Benítez, Jorge G. Figueroa, Karla Estrada, and Darío Cruz. 2023. "Phenotypic and Molecular Characterization of Yeast Diversity Associated to Postharvest Fermentation Process of Coffee Fruits in Southern Ecuador" Diversity 15, no. 9: 984. https://doi.org/10.3390/d15090984
APA StyleApolo, D., Fernández, J. M., Benítez, Á., Figueroa, J. G., Estrada, K., & Cruz, D. (2023). Phenotypic and Molecular Characterization of Yeast Diversity Associated to Postharvest Fermentation Process of Coffee Fruits in Southern Ecuador. Diversity, 15(9), 984. https://doi.org/10.3390/d15090984