

Ontogeny of Different Tetrad Types in the Single Microsporangium of Mitrephora tomentosa (Annonaceae)



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Material and Methods
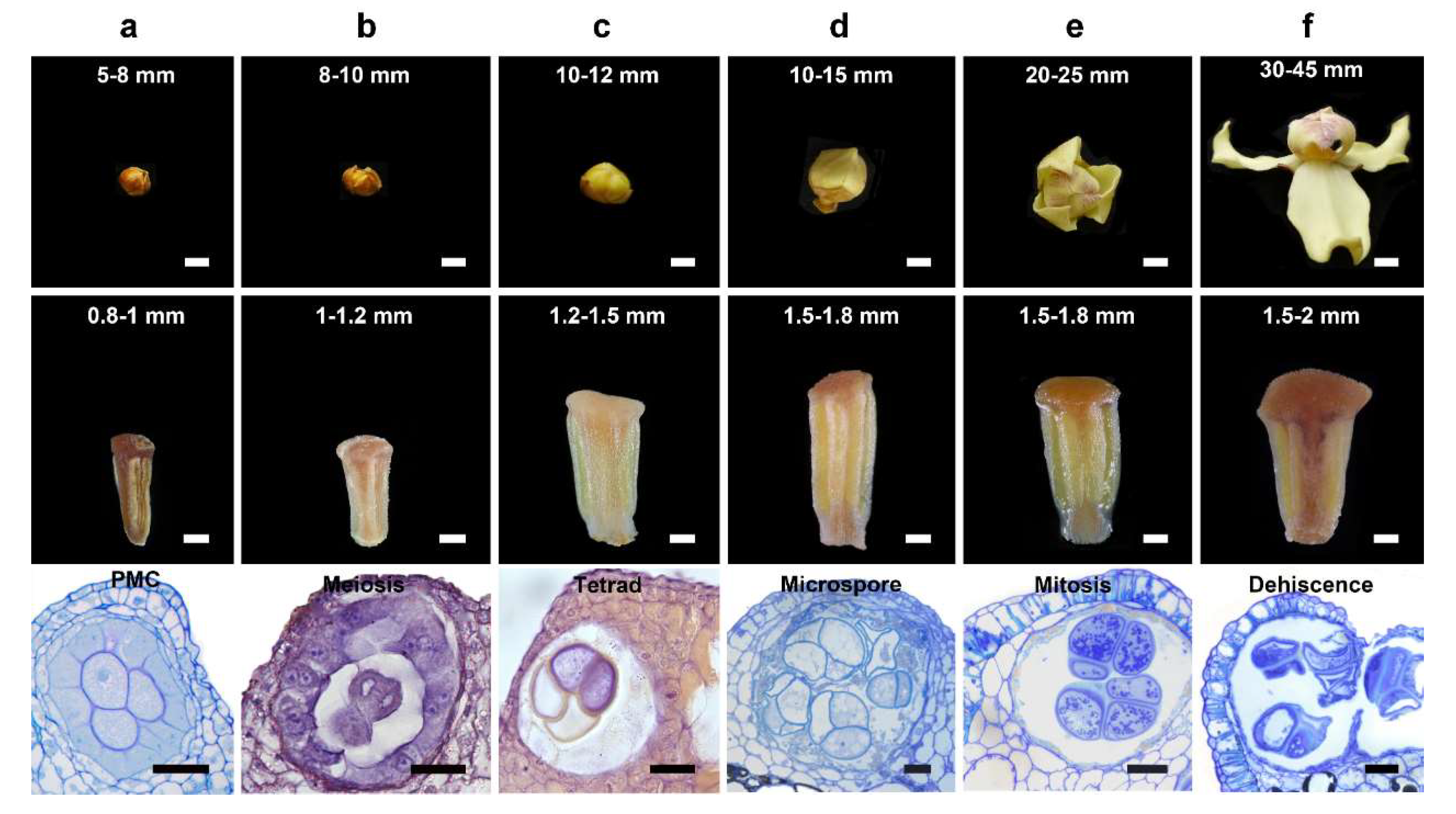
2.1. Plant Materials
2.2. Scanning Electron Microscopy (SEM)
2.3. Light Microscopy (LM)
2.4. Transmission Electron Microscopy (TEM)
2.5. Confocal Laser Scanning Microscopy (CLSM)
3. Results
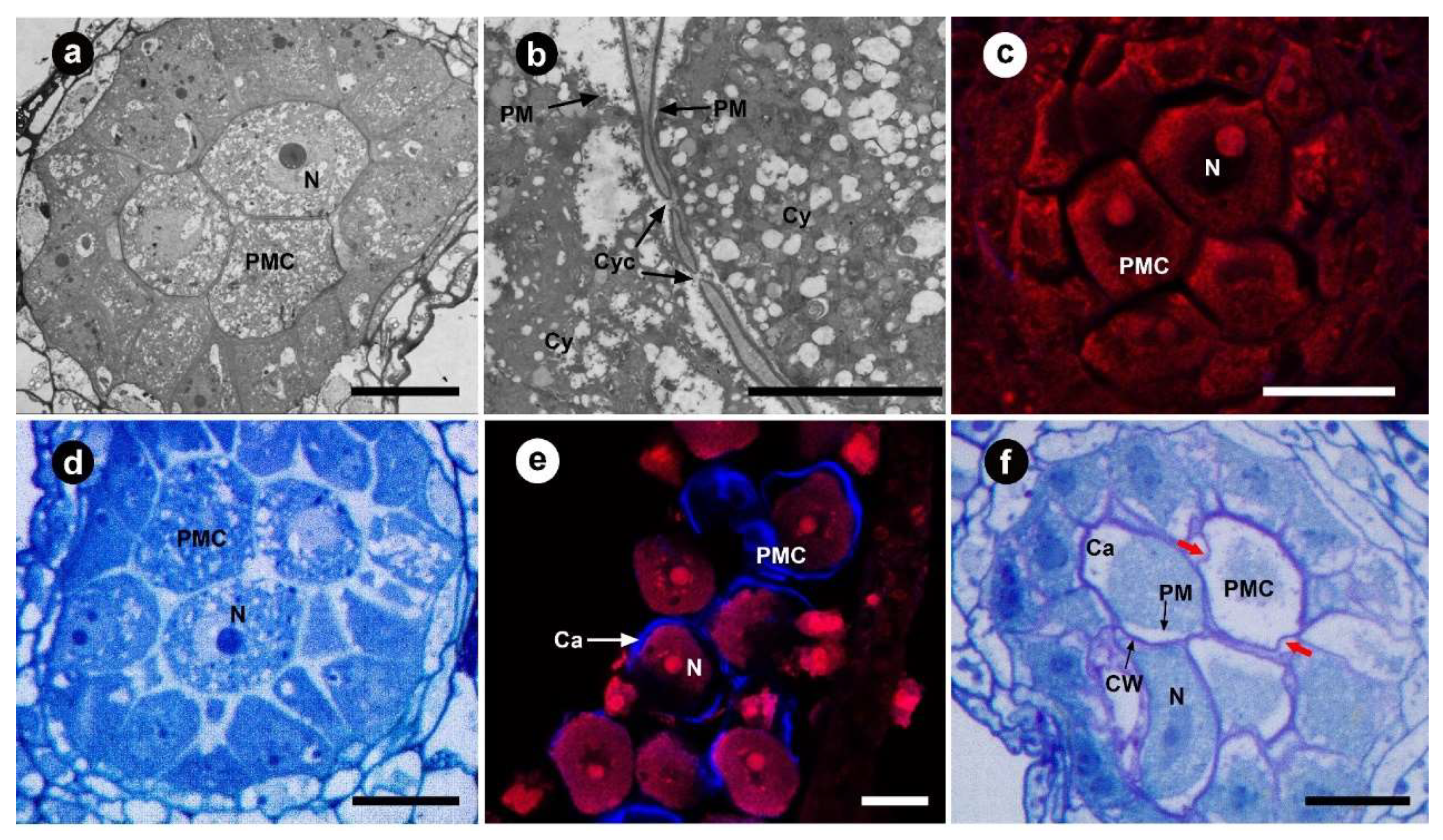
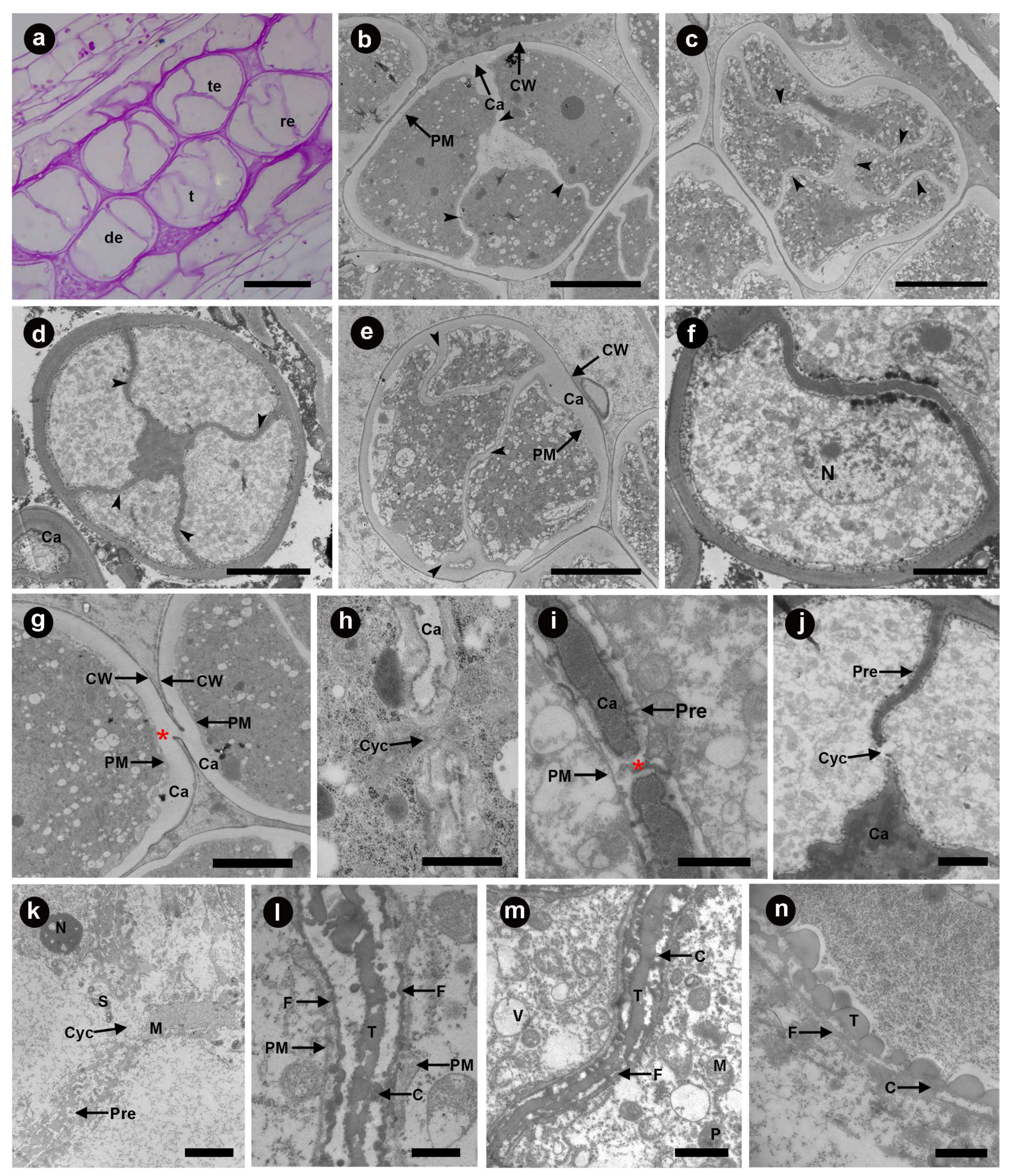
3.1. Pollen Mother Cell Stage
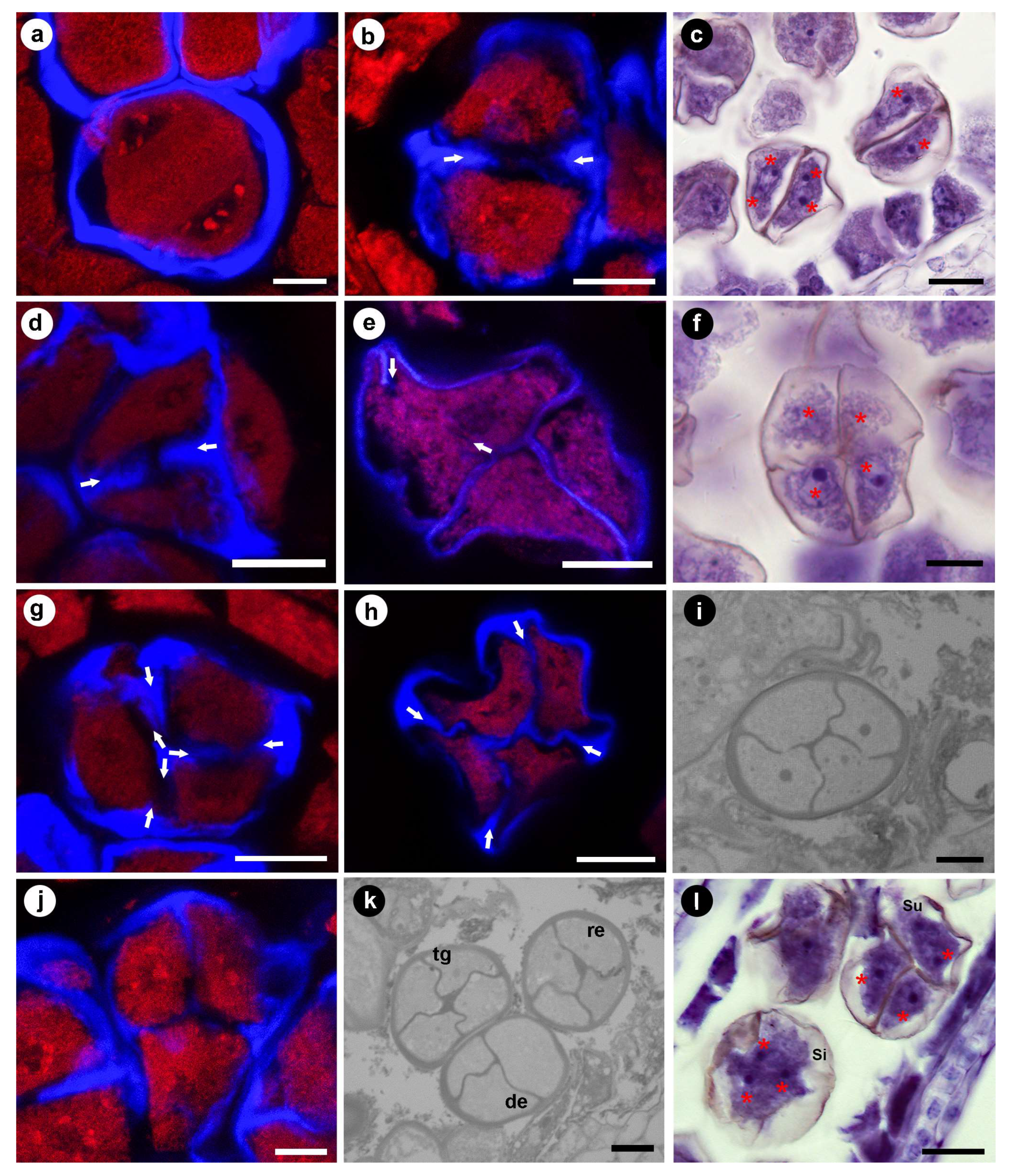
3.2. Meiosis Stage
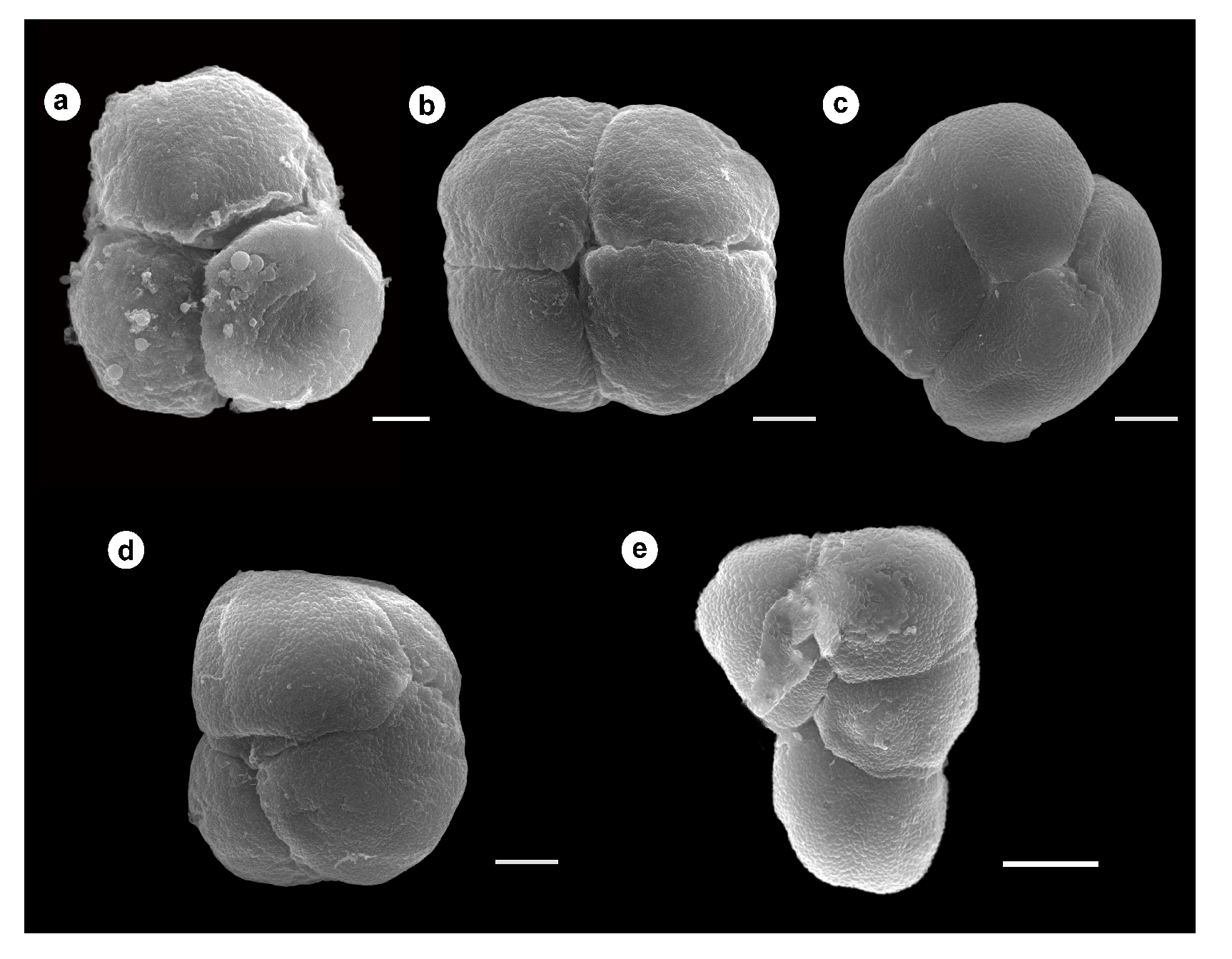
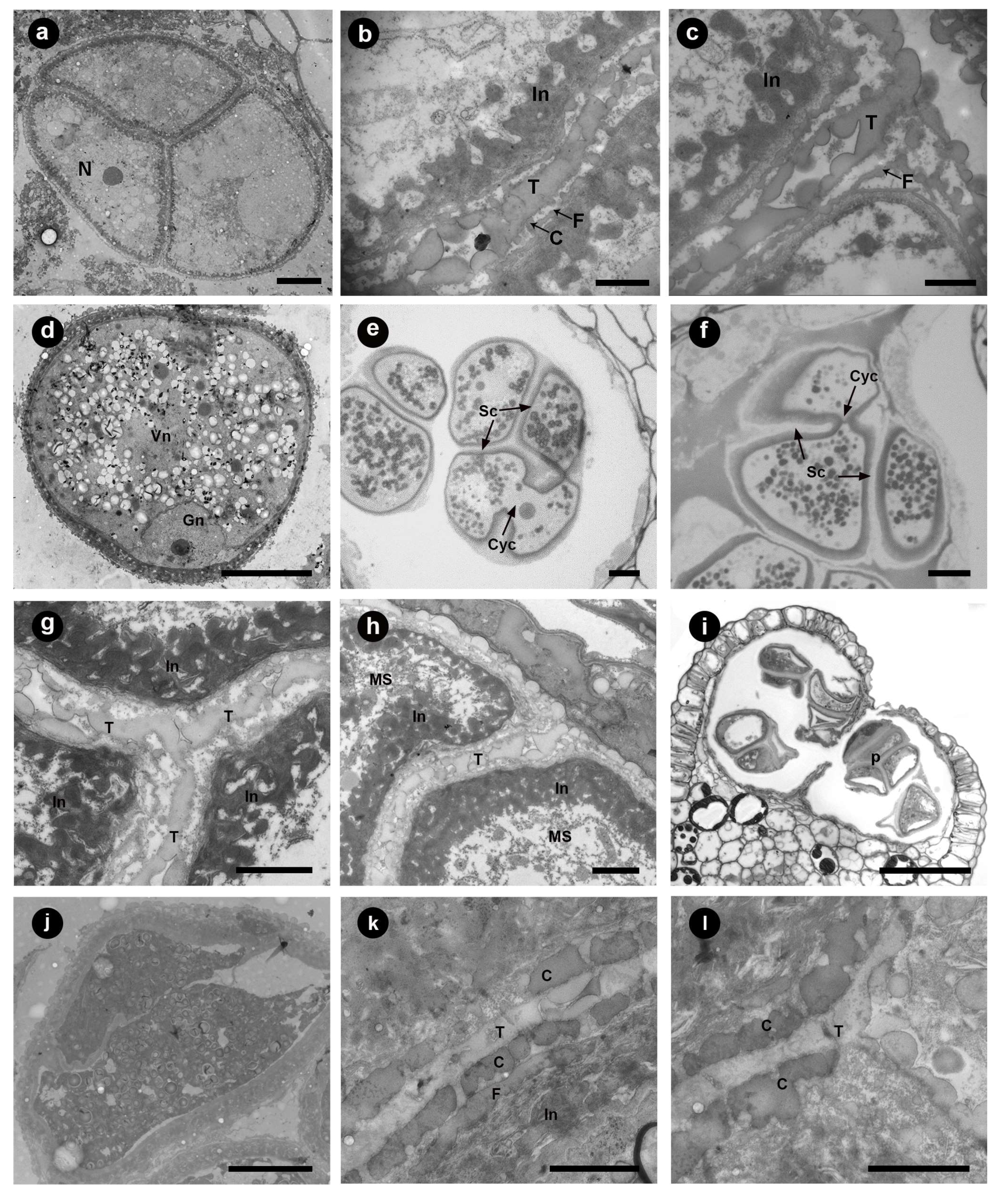
3.3. Tetrad Stage
3.4. Unicellular, Bicellular Microspore and Mature Pollen Stage
4. Discussion
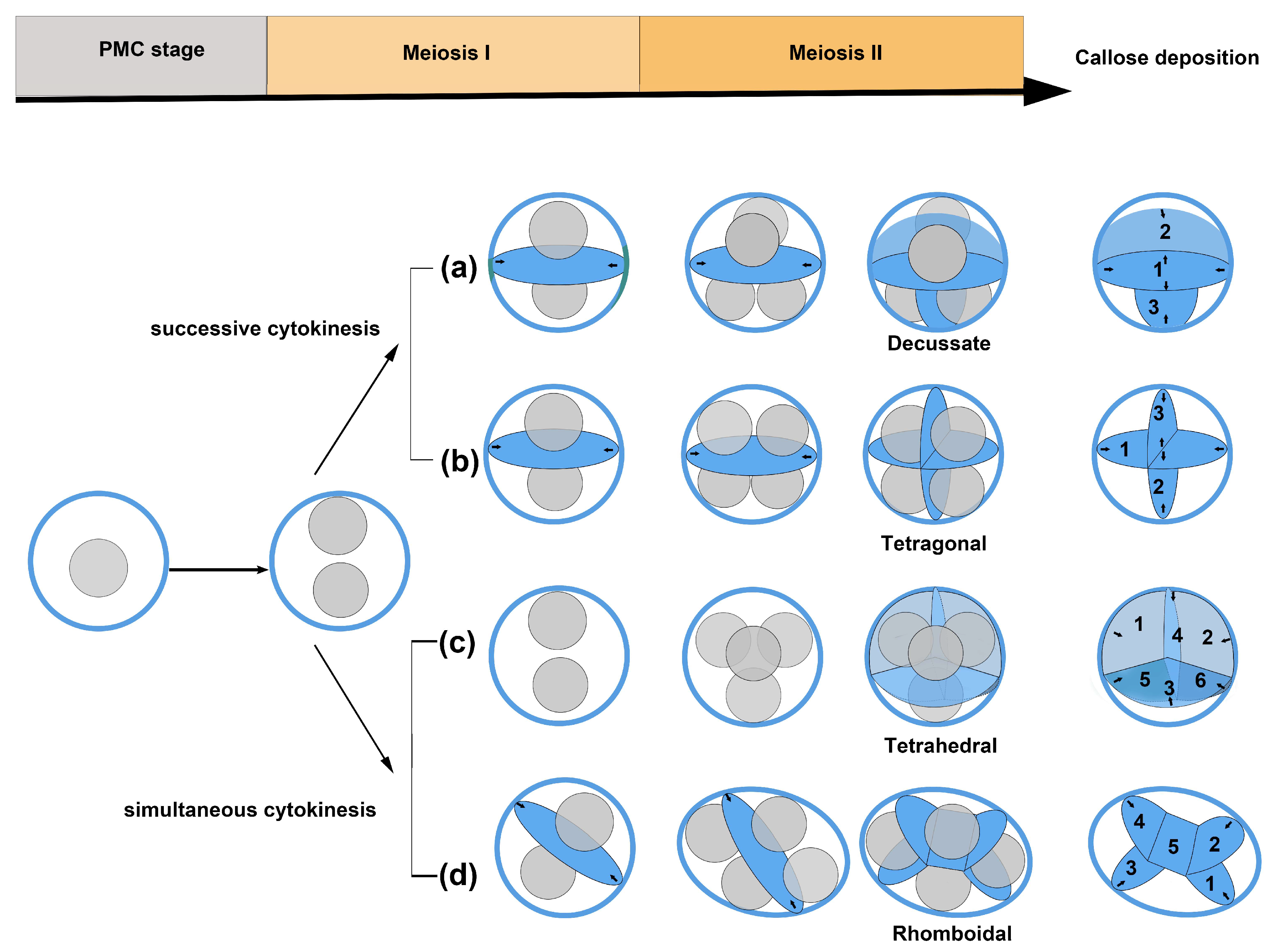
4.1. The Diversity of Microsporogenesis
4.2. Cohesion Mechanism of the Permanent Tetrad Pollen
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erdtman, G. Pollen Morphology and Plant Taxonomy. Angiosperms. An Introduction to Palynology I; Hafner Publishing: New York, NY, USA, 1966. [Google Scholar]
- Copenhaver, G.P. A compendium of plant species producing pollen tetrads. J. N. Carol. Acad. Sci. 2005, 121, 17–35. [Google Scholar]
- Walker, J.W. Pollen morphology, phytogeography, and phylogeny of the Annonaceae. Contrib. Gray Herb. Harv. Univ. 1971, 202, 1–130. [Google Scholar] [CrossRef]
- Walker, J.W. Contributions to the pollen morphology and phylogeny of the Annonaceae I. Grana 1971, 11, 45–54. [Google Scholar] [CrossRef]
- Zhang, M.Y.; Lu, L.; Wortley, A.H.; Wang, H.; Li, D.Z.; Blackmore, S. Evolution of angiosperm pollen. 4. The basal angiosperms. Ann. Mo. Bot. Gard. 2017, 102, 141–182. [Google Scholar] [CrossRef]
- Furness, C.A.; Rudall, P.J.; Sampson, F.B. Evolution of Microsporogenesis in Angiosperms. Int. J. Plant Sci. 2002, 163, 235–260. [Google Scholar] [CrossRef]
- Erdtman, G. Handbook of Palynology. Morphology, Taxonomy, Ecology. An Introduction to the Study of Pollen Grains and Spores; Hafner: New York, NY, USA, 1969. [Google Scholar]
- Albert, B.; Nadot, S.; Dreyer, L.; Ressayre, A. The influence of tetrad shape and intersporal callose wall formation on pollen aperture pattern ontogeny in two eudicot species. Ann. Bot. 2010, 106, 557–564. [Google Scholar] [CrossRef]
- Matamoro-Vidal, A.; Furness, C.A.; Gouyon, P.H.; Wurdack, K.J.; Albert, B. Evolutionary stasis in Euphorbiaceae pollen: Selection and constraints. J. Evol. Biol. 2012, 25, 1077–1096. [Google Scholar] [CrossRef]
- Penet, L. Premeiotic microsporocyte cell shape influences shape of tetrads during microsporogenesis. Int. J. Plant Sci. 2012, 173, 375–381. [Google Scholar] [CrossRef]
- Albert, B.; Toghranegar, Z.; Nadot, S. Diversity and evolution of microsporogenesis in Bromeliaceae. Bot. J. Linn. Soc. 2014, 176, 36–45. [Google Scholar] [CrossRef]
- Nadot, S.; Furness, C.A.; Sannier, J.; Penet, L.; Triki-Teurtroy, S.; Albert, B.; Ressayre, A. Phylogenetic comparative analysis of microsporogenesis in angiosperms with a focus on monocots. Am. J. Bot. 2008, 95, 1426–1436. [Google Scholar] [CrossRef]
- Ressayre, A.; Godelle, B.; Raquin, C.; Gouyon, P.H. Aperture pattern ontogeny in angiosperms. J. Exp. Zool. 2002, 294, 122–135. [Google Scholar] [CrossRef] [PubMed]
- Penet, L.; Nadot, S.; Ressayre, A.; Forchioni, A.; Dreyer, L.; Gouyon, P.H. Multiple developmental pathways leading to a single morph: Monosulcate pollen (examples from the Asparagales). Ann. Bot. 2005, 95, 331–343. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nadot, S.; Forchioni, A.; Penet, L.; Sannier, J.; Ressayre, A. Links between early pollen development and aperture pattern in monocots. Protoplasma 2006, 228, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Ressayre, A. Equatorial aperture pattern in monocots: Same definition rules as in eudicots? The example of two species of Pontederiaceae. Int. J. Plant Sci. 2001, 162, 1219–1224. [Google Scholar] [CrossRef]
- Ouyang, H.B.; Li, Y.; Zhang, S.Z.; Li, N.; Wu, H. Microsporogenesis of Cycas elongata and its systematic implication. Acta Bot. Sin. 2004, 42, 500–512. (In Chinese) [Google Scholar]
- Yan, X.; Bai, M.; Ning, X.; Ouyang, H.; Zhang, S.; Yang, M.; Wu, H. Spatiotemporal features of microsporogenesis in the cycad species Macrozamia communis. Am. J. Bot. 2015, 102, 1061–1072. [Google Scholar] [CrossRef]
- Zhang, H.F.; Ouyang, H.B.; Du, J.Y.; Zhang, S.Z.; Li, Y.; Wu, H. Microsporogenesis of Cycas and its systematic implications. J. Syst. Evol. 2012, 50, 125–134. [Google Scholar] [CrossRef]
- Galati, B.; Zarlavsky, G.; Rosenfeldt, S.; Gotelli, M. Pollen ontogeny in Magnolia liliflora Desr. Plant Syst. Evol. 2011, 298, 527–534. [Google Scholar] [CrossRef]
- Li, B.X.; Xu, F.X. Formation pattern in five types of pollen tetrad in Pseuduvaria trimera (Annonaceae). Protoplasma 2019, 256, 53–68. [Google Scholar] [CrossRef]
- Xu, F.X.; Chen, D.Q.; Specht, C. Comparative microsporogenesis and anther development of selected species from Magnoliaceae. Nord. J. Bot. 2013, 31, 291–300. [Google Scholar] [CrossRef]
- Sampson, F.B. Studies on the Monimiaceae. Aust. J. Bot. 1969, 17, 403–424. [Google Scholar] [CrossRef]
- Rudall, P.J.; Furness, C.A.; Chase, M.W.; Fay, M.F. Microsporogenesis and pollen sulcus type in Asparagales (Lilianae). Canadian J. Bot. 1997, 75, 408–430. [Google Scholar] [CrossRef]
- Chatrou, L.W.; Pirie, M.D.; Erkens, R.H.J.; Couvreur, T.L.P.; Neubig, K.M.; Abbott, J.R.; Mols, J.B.; Maas, J.W.; Saunders, R.M.K.; Chase, M.W. A new subfamilial and tribal classification of the pantropical flowering plant family Annonaceae informed by molecular phylogenetics. Bot. J. Linn. Soc. 2012, 169, 5–40. [Google Scholar] [CrossRef]
- Guo, X.; Tang, C.C.; Thomas, D.C.; Couvreur, T.L.P.; Saunders, R.M.K. A mega-phylogeny of the Annonaceae: Taxonomic placement of five enigmatic genera and support for a new tribe, Phoenicantheae. Sci. Rep. 2017, 7, 7323. [Google Scholar] [CrossRef]
- APG IV. An update of the Angiosperm phylogeny group classification for the orders and families of flowering plants: APG IV. Bot. J. Linn. Soc. 2016, 181, 1–20. [Google Scholar] [CrossRef]
- Xue, B.; Guo, X.; Landis, J.B.; Sun, M.; Tang, C.C.; Soltis, P.S.; Soltis, D.E.; Saunders, R.M.K. Accelerated diversification correlated with functional traits shapes extant diversity of the early divergent angiosperm family Annonaceae. Mol. Phylogenet. Evol. 2020, 142, 106659. [Google Scholar] [CrossRef] [PubMed]
- Tsou, C.H.; Fu, Y.L. Tetrad pollen formation in Annona (Annonaceae): Proexine formation and binding mechanism. Am. J. Bot. 2002, 89, 734–747. [Google Scholar] [CrossRef]
- Tsou, C.H.; Fu, Y.L. Octad pollen formation in Cymbopetalum (Annonaceae): The binding mechanism. Plant Syst. Evol. 2007, 263, 13–23. [Google Scholar] [CrossRef]
- Su, Y.C.F.; Saunders, R.M.K. Pollen structure, tetrad cohesion and pollen-connecting in Pseuduvaria (Annonaceae). Bot. J. Linn. Soc. 2003, 143, 69–78. [Google Scholar] [CrossRef]
- Lora, J.; Testillano, P.S.; Risueno, M.C.; Hormaza, J.I.; Herrero, M. Pollen development in Annona cherimola Mill. (Annonaceae). Implications for the evolution of aggregated pollen. Bmc Plant Biol. 2009, 9, 129. [Google Scholar] [CrossRef] [PubMed]
- Lora, J.; Herrero, M.; Hormaza, J.I. Microspore development in Annona (Annonaceae): Differences between monad and tetrad pollen. Am. J. Bot. 2014, 101, 1508–1518. [Google Scholar] [CrossRef]
- Halbritter, H.; Silvia, U.; Grímsson, F.; Weber, M.; Frosch-Radivo, A. Illustrated Pollen Terminology, 2nd ed.; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Tidy, A.C.; Ferjentsikova, I.; Vizcay-Barrena, G.; Liu, B.; Yin, W.Z.; Higgins, J.D.; Xu, J.; Zhang, D.B.; Geelen, D.; Wilson, Z.A. Sporophytic control of pollen meiotic progression is mediated by tapetum expression of Aborted microspores. J. Exp. Bot. 2022, 73, 5543–555840. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.F.; Wu, Y.; Zhang, G.F. Formation pattern and regulatory mechanisms of pollen wall in Arabidopsis. J. Plant Physiol. 2021, 260, 153388. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.J.; Kang, J.H.; Zhao, M.A.; Kwon, J.K.; Choi, H.S.; Bae, J.H.; Lee, H.A.; Joung, Y.H.; Choi, D.; Kang, B.C. Tomato Male sterile 1035 is essential for pollen development and meiosis in anthers. J. Exp. Bot. 2014, 65, 6693–6709. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Bai, M.; Yang, M.; Wu, H. Cell polarity, asynchronous nuclear divisions, and bidirectional cytokinesis in male meiosis in Magnolia denudata. Protoplasma. 2021, 258, 621–632. [Google Scholar] [CrossRef]
- Wu, H.; Hu, M.L.; Yang, M. Nymphaea colorata, another basal angiosperm species with bidirectional cytokinesis in microsporogenesis. Plant Signal. Behav. 2021, 16, 1559–2316. [Google Scholar] [CrossRef]
- Sannier, J.; Nadot, S.; Forchioni, A.; Harley, M.; Albert, B.; Harley, M. Variation in the microsporogenesis of monosulcate palm pollen. Bot. J. Linn. Soc. 2006, 151, 93–102. [Google Scholar] [CrossRef][Green Version]
- Blakmore, S.; Wortley, A.H.; Skvarla, J.J.; Rowley, J.R. Pollen wall development in flowering plants. New Phytol. 2007, 174, 483–498. [Google Scholar] [CrossRef]
- Matamoro-Vidal, A.; Prieu, C.; Furness, C.A.; Albert, B.; Gouyon, P.H. Evolutionary stasis in pollen morphogenesis due to natural selection. New Phytol. 2016, 209, 376–394. [Google Scholar] [CrossRef]
- Albert, B.; Raquin, C.; Prigent, M.; Nadot, S.; Brisset, F.; Yang, M.; Ressayre, A. Successive microsporogenesis affects pollen aperture pattern in the tam mutant of Arabidopsis thaliana. Ann. Bot. 2011, 107, 1421–1426. [Google Scholar] [CrossRef]
- Walker, J.W.; Doyle, J.A. Bases of angiosperm phylogeny-palynology. Ann. Mo. Bot. Gard. 1975, 62, 664–723. [Google Scholar] [CrossRef]
- Ressayre, A.; Dreyer, L.; Triki-Teurtroy, S.; Forchioni, A.; Nadot, S. Post-meiotic cytokinesis and pollen aperture pattern ontogeny: Comparison of development in four species differing in aperture pattern. Am. J. Bot. 2005, 92, 576–583. [Google Scholar] [CrossRef]
- Albert, B.; Matamoro-Vidal, A.; Prieu, C.; Nadot, S.; Till-Bottraud, I.; Ressayre, A.; Gouyon, P.H. A review of the developmental processes and selective pressures shaping aperture pattern in Angiosperms. Plants 2022, 11, 357. [Google Scholar] [CrossRef] [PubMed]
- Furness, C.A.; Rudall, P.J. Pollen and anther characters in monocot systematics. Grana 2001, 40, 17–25. [Google Scholar] [CrossRef]
- Toghranegar, Z.; Nadot, S.; Albert, B. Variation of microsporogenesis in monocots producing monosulcate pollen grains. Ann. Bot. 2013, 112, 135–139. [Google Scholar] [CrossRef]
- Periasamy, K.; Thangavel, S. Anther development in Xylopia nigricans. Proc. Indian Acad. Sci. 1988, 98, 251–255. [Google Scholar] [CrossRef]
- Locke, J.F. Microsporogenesis and cytokinesis in Asimina triloba. Bot Gaz. 1936, 98, 159–168. [Google Scholar] [CrossRef]
- Gabarayeva, N.I. Sporoderm development in Asimina triloba (Annonaceae). Grana 1992, 31, 213–222. [Google Scholar] [CrossRef][Green Version]
- Gabarayeva, N.I. Sporoderm Development in Asimina triloba (Annonaceae). II. The developmental events after callose dissolution. Grana 1993, 32, 210–220. [Google Scholar] [CrossRef]
- Taylor, M.L.; Hudson, P.J.; Rigg, J.M.; Strandquist, J.N.; Schwartz, G.J.; Thiemann, T.C.; Osborn, J.M. Pollen ontogeny in Victoria (Nymphaeales). Int. J. Plant Sci. 2013, 174, 1259–1276. [Google Scholar] [CrossRef]
- Sarwar, A. Ontogenetic development of pollen tetrads in Vaccinium smallii A. Gray (Ericaceae). Jpn. J. Palynol. 2012, 58, 61–71. [Google Scholar]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, B.; Liu, X.; Cheng, M.; Xu, F.; Liao, H. Ontogeny of Different Tetrad Types in the Single Microsporangium of Mitrephora tomentosa (Annonaceae). Diversity 2023, 15, 898. https://doi.org/10.3390/d15080898
Li B, Liu X, Cheng M, Xu F, Liao H. Ontogeny of Different Tetrad Types in the Single Microsporangium of Mitrephora tomentosa (Annonaceae). Diversity. 2023; 15(8):898. https://doi.org/10.3390/d15080898
Chicago/Turabian StyleLi, Bingxin, Xiu Liu, Mei Cheng, Fengxia Xu, and Haimin Liao. 2023. "Ontogeny of Different Tetrad Types in the Single Microsporangium of Mitrephora tomentosa (Annonaceae)" Diversity 15, no. 8: 898. https://doi.org/10.3390/d15080898
APA StyleLi, B., Liu, X., Cheng, M., Xu, F., & Liao, H. (2023). Ontogeny of Different Tetrad Types in the Single Microsporangium of Mitrephora tomentosa (Annonaceae). Diversity, 15(8), 898. https://doi.org/10.3390/d15080898