


Genome-Based Species Diversity Assessment in the Pseudomonas chlororaphis Phylogenetic Subgroup and Proposal of Pseudomonas danubii sp. nov. Isolated from Freshwaters, Soil, and Rhizosphere

 , , , , , , and
, , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Phylogenetic Analyses of the 16S rRNA, gyrB, rpoB, and rpoD Gene Sequences and Multilocus Sequence Analysis (MLSA)
2.3. Genome Sequencing
2.4. Phylogenomic Analyses
2.5. Genome Insights
2.6. Morphological, Biochemical, and Physiological Tests
2.7. Chemotaxonomic Analysis
3. Results
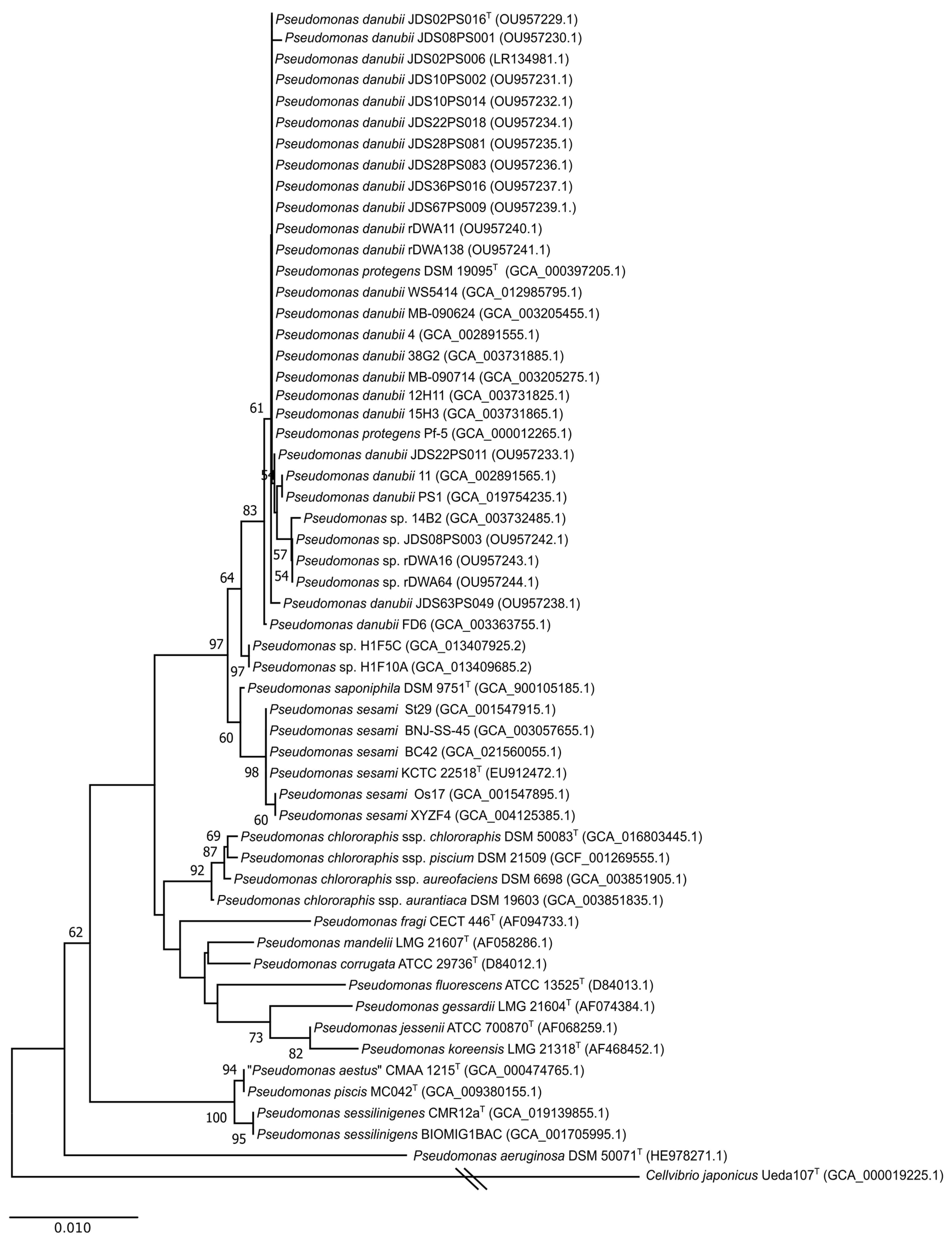
3.1. 16S rRNA and rpoD Gene Phylogenies
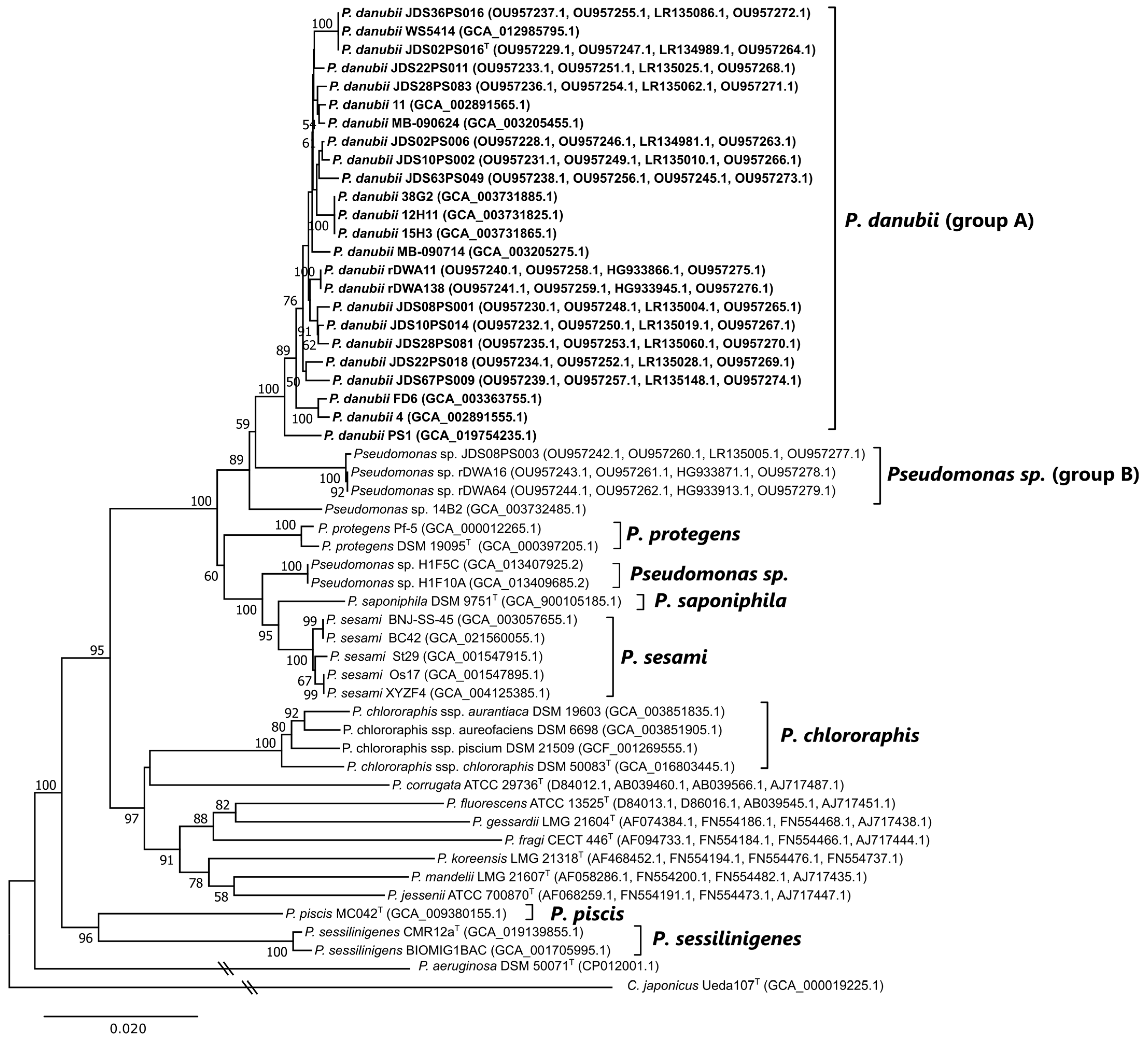
3.2. Multilocus Sequence Analysis (MLSA)
3.3. Genome Characteristics
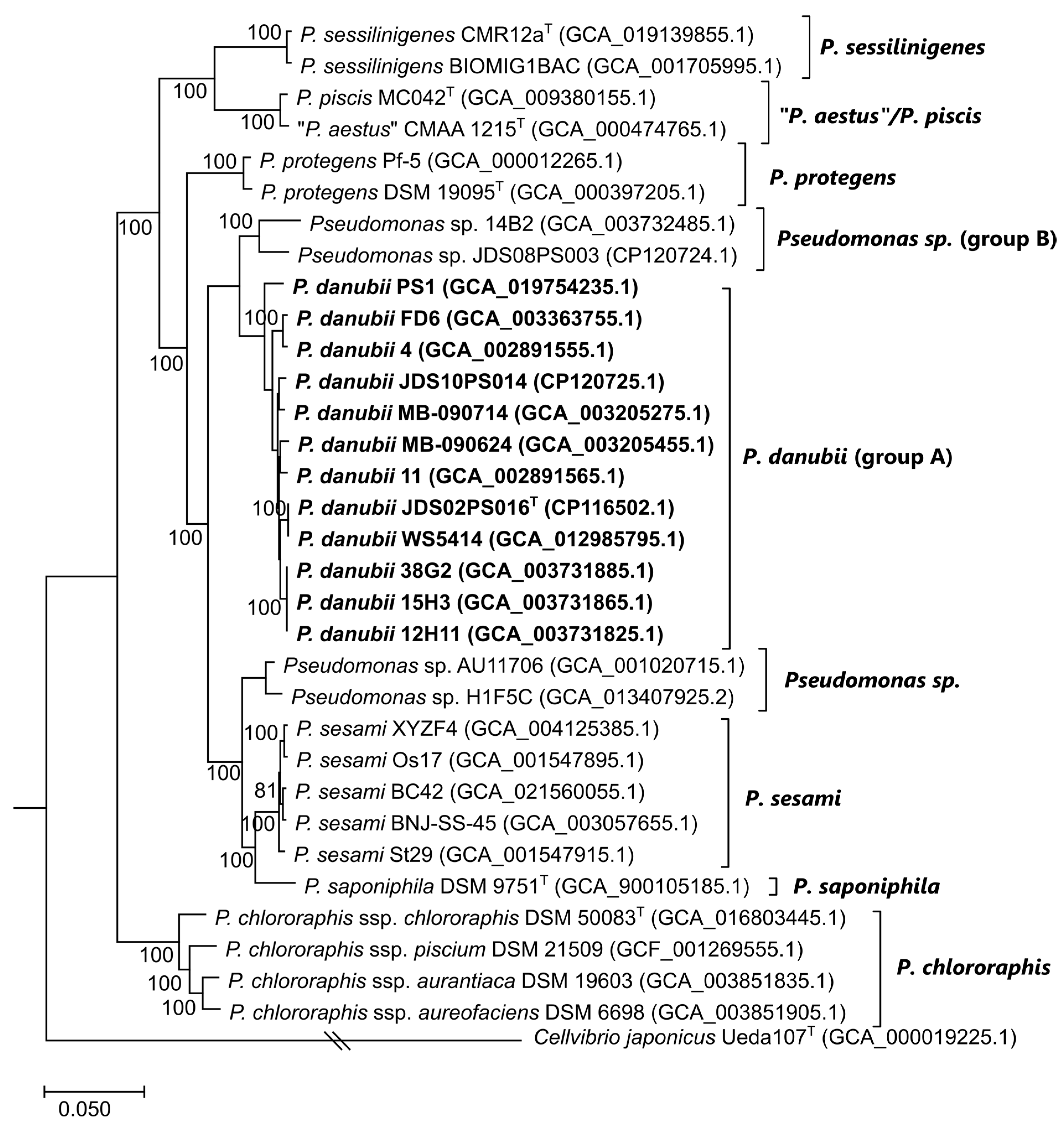
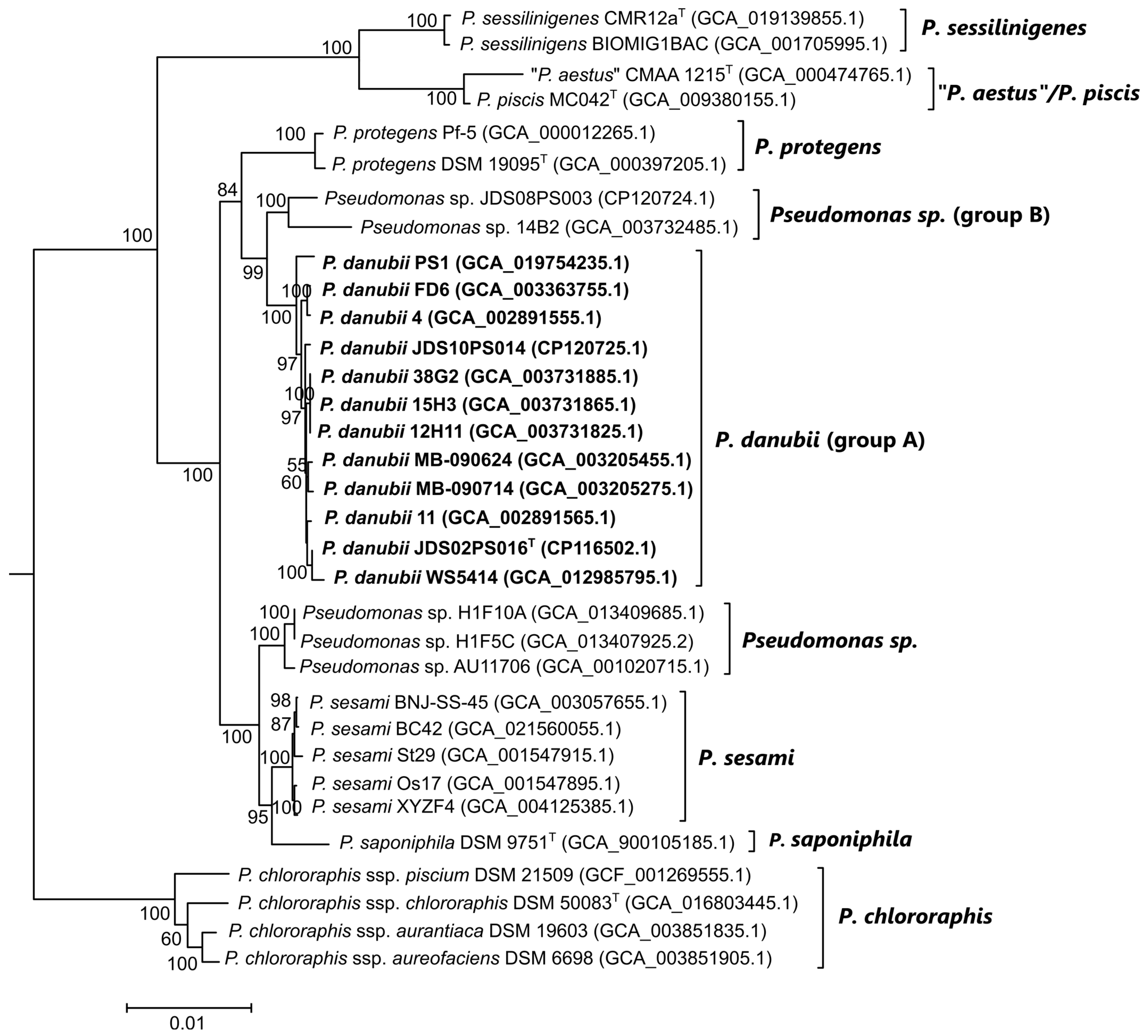
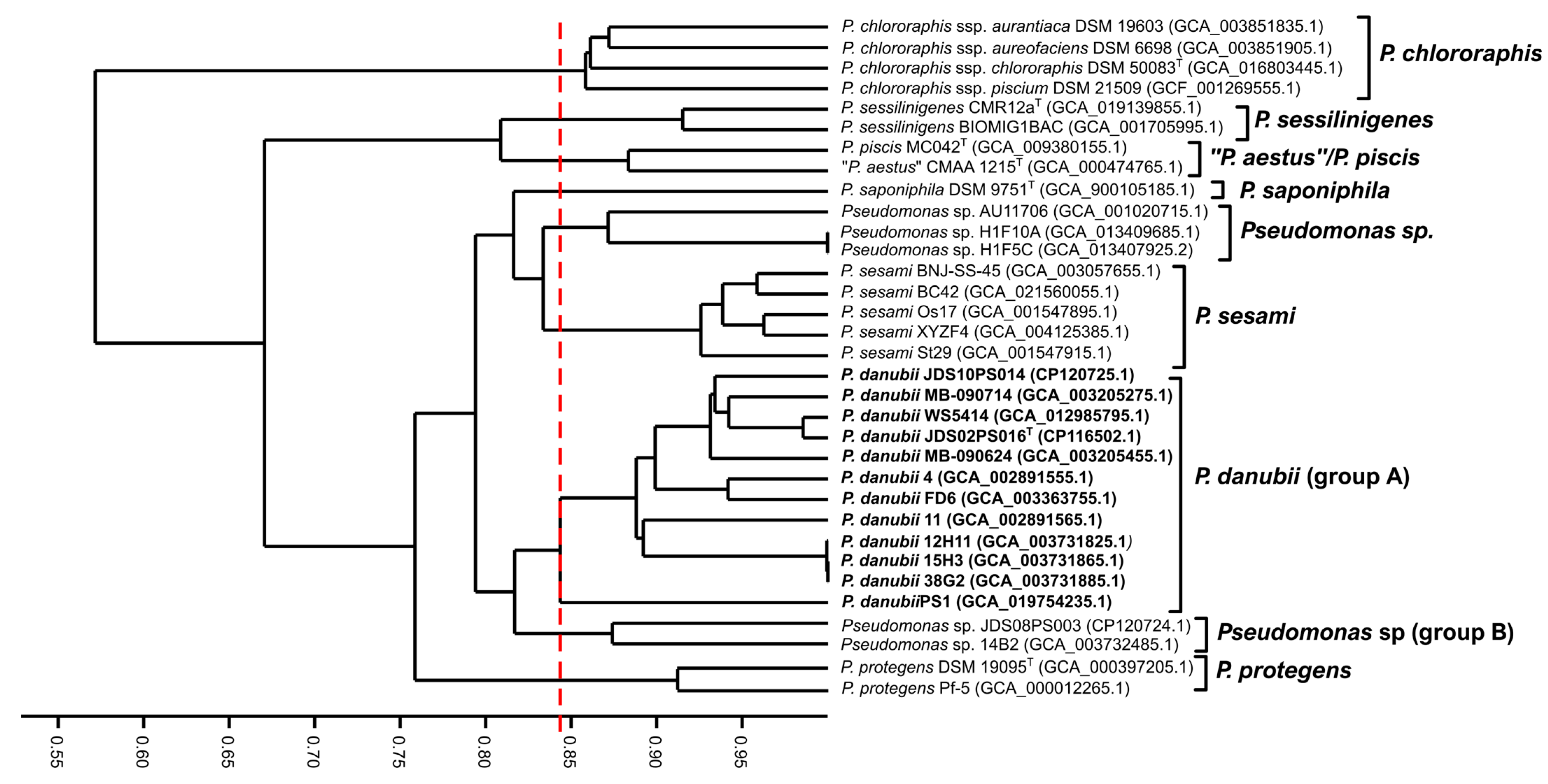
3.4. Genome-Based Phylogeny and Species Circumscription
3.5. Gene Content Comparisons
3.6. Genome Insights
3.7. Phenotypic Traits: Morphology, Physiology, Biochemical Traits, and Chemotaxonomy
4. Discussion
5. Protologue
Description of Pseudomonas danubii sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANI | genome-aggregated average nucleotide identity |
| autoMLST | automated multilocus species tree |
| FAME | fatty acid methyl ester analysis |
| GGDC | genome to genome distance calculator |
| MALDI-TOF MS | matrix-assisted laser desorption/ionization (MALDI) time-of-flight (TOF) mass spectrometry |
| MLSA | multilocus sequence analysis |
References
- Palleroni, N.J.; Genus, I. Pseudomonas. In Bergey’s Manual of Systematic Bacteriology. Part B. the Gammaproteobacteria; Brenner, D.J., Krieg, N.R., Staley, J.T., Eds.; Springer: New York, NY, USA, 2005; pp. 323–379. [Google Scholar]
- Parte, A.C.; Sardà Carbasse, J.; Meier-Kolthoff, J.P.; Reimer, L.C.; Göker, M. List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. Int. J. Syst. Evol. Microbiol. 2020, 70, 5607–5612. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Mulet, M.; Gomila, M.; García-Valdés, E. Genomics in Bacterial Taxonomy: Impact on the Genus Pseudomonas. Genes 2020, 11, 139. [Google Scholar] [CrossRef]
- Rudra, B.; Gupta, R.S. Phylogenomic and comparative genomic analyses of species of the family Pseudomonadaceae: Proposals for the genera Halopseudomonas gen. nov. and Atopomonas gen. nov., merger of the genus Oblitimonas with the genus Thiopseudomonas, and transfer of some misclassified species of the genus Pseudomonas into other genera. Int. J. Syst. Evol. Microbiol. 2021, 71, 5011. [Google Scholar] [CrossRef]
- Saati-Santamaría, Z.; Peral-Aranega, E.; Velázquez, E.; Rivas, R.; García-Fraile, P. Phylogenomic Analyses of the Genus Pseudomonas Lead to the Rearrangement of Several Species and the Definition of New Genera. Biology 2021, 10, 782. [Google Scholar] [CrossRef]
- Mulet, M.; Lalucat, J.; García-Valdés, E. DNA sequence-based analysis of the Pseudomonas species. Environ. Microbiol. 2010, 12, 1513–1530. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J.; Gomila, M.; Mulet, M.; Zaruma, A.; García-Valdés, E. Past, present and future of the boundaries of the Pseudomonas genus: Proposal of Stutzerimonas gen. Nov. Syst. Appl. Microbiol. 2022, 45, 126289. [Google Scholar] [CrossRef]
- Nikolaidis, M.; Mossialos, D.; Oliver, S.G.; Amoutzias, G.D. Comparative Analysis of the Core Proteomes among the Pseudomonas Major Evolutionary Groups Reveals Species-Specific Adaptations for Pseudomonas aeruginosa and Pseudomonas chlororaphis. Diversity 2020, 12, 289. [Google Scholar] [CrossRef]
- Girard, L.; Lood, C.; Höfte, M.; Vandamme, P.; Rokni-Zadeh, H.; van Noort, V.; Lavigne, R.; De Mot, R. The Ever-Expanding Pseudomonas Genus: Description of 43 New Species and Partition of the Pseudomonas putida Group. Microorganisms 2021, 9, 1766. [Google Scholar] [CrossRef]
- Scales, B.S.; Dickson, R.P.; LiPuma, J.J.; Huffnagle, G.B. Microbiology, Genomics, and Clinical Significance of the Pseudomonas fluorescens Species Complex, an Unappreciated Colonizer of Humans. Clin. Microbiol. Rev. 2014, 27, 927–948. [Google Scholar] [CrossRef]
- Scales, B.S.; Erb-Downward, J.R.; LiPuma, J.J.; Huffnagle, G.B. Draft Genome Sequences of Five Pseudomonas fluorescens Subclade I and II Strains, Isolated from Human Respiratory Samples. Genome Announc. 2015, 3, e00837-15. [Google Scholar] [CrossRef]
- Kittinger, C.; Lipp, M.; Baumert, R.; Folli, B.; Koraimann, G.; Toplitsch, D.; Liebmann, A.; Grisold, A.J.; Farnleitner, A.H.; Kirschner, A.; et al. Antibiotic Resistance Patterns of Pseudomonas spp. Isolated from the River Danube. Front. Microbiol. 2016, 7, 586. [Google Scholar] [CrossRef] [PubMed]
- Mulet, M.; Montaner, M.; Román, D.; Gomila, M.; Kittinger, C.; Zarfel, G.; Lalucat, J.; García-Valdés, E. Pseudomonas Species Diversity along the Danube River Assessed by rpoD Gene Sequence and MALDI-TOF MS Analyses of Cultivated Strains. Front. Microbiol. 2020, 11, 2114. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, D.; Matthijs, S.; Gomila, M.; Tricot, C.; Mulet, M.; García-Valdés, E.; Lalucat, J. rpoD Gene Pyrosequencing for the Assessment of Pseudomonas Diversity in a Water Sample from the Woluwe River. Appl. Environ. Microbiol. 2014, 80, 4738–4744. [Google Scholar] [CrossRef] [PubMed]
- Vasconcellos, R.L.F.; Santos, S.N.; Zucchi, T.D.; Silva, F.S.P.; Souza, D.T.; Melo, I.S. Pseudomonas aestus sp. nov., a plant growth-promoting bacterium isolated from mangrove sediments. Arch. Microbiol. 2017, 199, 1223–1229. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Rao, Q.; Blom, J.; Lin, Q.; Luo, T. Pseudomonas piscis sp. nov., isolated from the profound head ulcers of farmed Murray cod (Maccullochella peelii peelii). Int. J. Syst. Evol. Microbiol. 2020, 70, 2732–2739. [Google Scholar] [CrossRef]
- Peix, A.; Valverde, A.; Rivas, R.; Igual, J.M.; Bahena, M.H.R.; Mateos, P.; Santa-Regina, I.; Rodríguez-Barrueco, C.; Martínez-Molina, E.; Velázquez, E. Reclassification of Pseudomonas aurantiaca as a synonym of Pseudomonas chlororaphis and proposal of three subspecies, P. chlororaphis subsp. chlororaphis subsp. nov., P. chlororaphis subsp. aureofaciens subsp. nov., comb. nov. and P. chlororaphis subsp. aurantiaca subsp. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 2007, 57, 1286–1290. [Google Scholar] [CrossRef]
- Burr, S.; Gobeli, S.; Kuhnert, P.; Goldschmidt-Clermont, E.; Frey, J. Pseudomonas chlororaphis subsp. piscium subsp. nov., isolated from freshwater fish. Int. J. Syst. Evol. Microbiol. 2010, 60, 2753–2757. [Google Scholar] [CrossRef]
- Ramette, A.; Frapolli, M.; Saux, M.F.-L.; Gruffaz, C.; Meyer, J.-M.; Défago, G.; Sutra, L.; Moënne-Loccoz, Y. Pseudomonas protegens sp. nov., widespread plant-protecting bacteria producing the biocontrol compounds 2,4-diacetylphloroglucinol and pyoluteorin. Syst. Appl. Microbiol. 2011, 34, 180–188. [Google Scholar] [CrossRef]
- Lang, E.; Burghartz, M.; Spring, S.; Swiderski, J.; Spröer, C. Pseudomonas benzenivorans sp. nov. and Pseudomonas saponiphila sp. nov., Represented by Xenobiotics Degrading Type Strains. Curr. Microbiol. 2010, 60, 85–91. [Google Scholar] [CrossRef]
- Madhaiyan, M.; Poonguzhali, S.; Saravanan, V.S.; Selvapravin, K.; Duraipandiyan, V.; Al-Dhabi, N.A. Pseudomonas sesami sp. nov., a plant growth-promoting Gammaproteobacteria isolated from the rhizosphere of Sesamum indicum L. Antonie Van Leeuwenhoek 2017, 110, 843–852. [Google Scholar] [CrossRef]
- Paulsen, I.T.; Press, C.; Ravel, J.; Kobayashi, D.Y.; Myers, G.; Mavrodi, D.; DeBoy, R.T.; Seshadri, R.; Ren, Q.; Madupu, R.; et al. Complete genome sequence of the plant commensal Pseudomonas fluorescens Pf-5. Nat. Biotechnol. 2005, 23, 873–878, Erratum in Nat. Biotechnol. 2006, 24, 466. [Google Scholar] [CrossRef] [PubMed]
- Mulet, M.; Gomila, M.; Ramírez, A.; Cardew, S.; Moore, E.R.B.; Lalucat, J.; García-Valdés, E. Uncommonly isolated clinical Pseudomonas: Identification and phylogenetic assignation. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Jukes, T.; Cantor, C. Evolution of protein molecules. In Mammalian Protein Metabolism; Munro, H.N., Ed.; Academic Press: New York, NY, USA, 1969; pp. 21–132. [Google Scholar]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Alanjary, M.; Steinke, K.; Ziemert, N. AutoMLST: An automated web server for generating multi-locus species trees highlighting natural product potential. Nucleic Acids Res. 2019, 47, W276–W282. [Google Scholar] [CrossRef]
- Wagner, G.E.; Dabernig-Heinz, J.; Lipp, M.; Cabal, A.; Simantzik, J.; Kohl, M.; Scheiber, M.; Lichtenegger, S.; Ehricht, R.; Leitner, E.; et al. Real-Time Nanopore Q20+ Sequencing Enables Extremely Fast and Accurate Core Genome MLST Typing and Democratizes Access to High-Resolution Bacterial Pathogen Surveillance. J. Clin. Microbiol. 2023, 61, e0163122. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Bickhart, D.M.; Behsaz, B.; Gurevich, A.; Rayko, M.; Shin, S.B.; Kuhn, K.; Yuan, J.; Polevikov, E.; Smith, T.P.L.; et al. metaFlye: Scalable long-read metagenome assembly using repeat graphs. Nat. Methods 2020, 17, 1103–1110. [Google Scholar] [CrossRef]
- Aziz, R.K.; Bartels, D.; Best, A.A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST server: Rapid annotations using subsystems technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef]
- Richter, M.; Rosselló-Móra, R.; Oliver Glöckner, F.O.; Peplies, J. JSpeciesWS: A web server for prokaryotic species circumscription based on pairwise genome comparison. Bioinformatics 2016, 32, 929–931. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Göker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef] [PubMed]
- Avram, O.; Rapoport, D.; Portugez, S.; Pupko, T. M1CR0B1AL1Z3R—A user-friendly web server for the analysis of large-scale microbial genomics data. Nucleic Acids Res. 2019, 47, W88–W92. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. Available online: http://palaeo-electronica.org/2001_1/past/issue1_01.htm (accessed on 1 February 2023).
- Huson, D.H.; Bryant, D. Application of Phylogenetic Networks in Evolutionary Studies. Mol. Biol. Evol. 2006, 23, 254–267. [Google Scholar] [CrossRef]
- Oliveros, J.C. Venny. An Interactive Tool for Comparing Lists with Venn Diagrams. Available online: http://bioinfogp.cnb.csic.es/tools/venny/index.html (accessed on 1 February 2023).
- Alcock, B.P.; Huynh, W.; Chalil, R.; Smith, K.W.; Raphenya, A.R.; A Wlodarski, M.; Edalatmand, A.; Petkau, A.; A Syed, S.; Tsang, K.K.; et al. CARD 2023: Expanded curation, support for machine learning, and resistome prediction at the comprehensive antibiotic resistance database. Nucleic Acids Res. 2023, 51, D690–D699. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishardt, D.S. PHASTER: A better faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Moriya, Y.; Itoh, M.; Okuda, S.; Yoshizawa, A.C.; Kanehisa, M. KAAS: An automatic genome annotation and pathway reconstruction server. Nucleic Acids Res. 2007, 35 (Suppl. S2), W182–W185. [Google Scholar] [CrossRef] [PubMed]
- Lalucat, J. Analysis of Refractile (R) Bodies. In Methods in Microbiology; Academic Press: Cambridge, MA, USA, 1988; Volume 20, pp. 79–90. [Google Scholar]
- Sánchez, D.; Mulet, M.; Rodríguez, A.C.; David, Z.; Lalucat, J.; García-Valdés, E. Pseudomonas aestusnigri sp. nov., isolated from crude oil-contaminated intertidal sand samples after the Prestige oil spill. Syst. Appl. Microbiol. 2014, 37, 89–94. [Google Scholar] [CrossRef]
- Sasser, M. Identification of Bacteria by Gas Chromatography of Cellular Fatty Acids; MIDI Technical Note 101; MIDI Inc.: Newark, NJ, USA, 1990. [Google Scholar]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.-A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Mulet, M.; Bennasar, A.; Lalucat, J.; García-Valdés, E. An rpoD-based PCR procedure for the identification of Pseudomonas species and for their detection in environmental samples. Mol. Cell. Probes 2009, 23, 140–147. [Google Scholar] [CrossRef]
- Ghyselinck, J.; Coorevits, A.; Van Landschoot, A.; Samyn, E.; Heylen, K.; De Vos, P. An rpoD gene sequence based evaluation of cultured Pseudomonas diversity on different growth media. Microbiology 2013, 159, 2097–2108. [Google Scholar] [CrossRef]
- Girard, L.; Lood, C.; Rokni-Zadeh, H.; Van Noort, V.; Lavigne, R.; De Mot, R. Reliable Identification of Environmental Pseudomonas Isolates Using the rpoD Gene. Microorganisms 2020, 8, 1166. [Google Scholar] [CrossRef] [PubMed]
- Fortier, L.-C.; Sekulovic, O. Importance of prophages to evolution and virulence of bacterial pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef] [PubMed]
- Tao, X.; Zhang, H.; Gao, M.; Li, M.; Zhao, T.; Guan, X. Pseudomonas species isolated via high-throughput screening significantly protect cotton plants against verticillium wilt. AMB Express 2020, 10, 193. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Genome | Previous Identification in the GTDB Taxonomy or Corresponding Reference | NCBI Organism Name in the Assembly Information | NCBI Taxonomy: Species Check in the Assembly Information | Identification in This Study | Reference |
|---|---|---|---|---|---|---|
| JDS02PS006 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS02PS016T | CP116502 | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS08PS001 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS10PS002 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS10PS014 | CP120725 | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS22PS011 | -- | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS22PS018 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS28PS081 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS28PS083 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS36PS016 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS63PS049 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| JDS67PS009 | - | Pseudomonas sp. PSNew6 | - | - | P. danubii | [12,13] |
| rDWA11 | - | Pseudomonas sp. | - | - | P. danubii | [14] |
| rDWA138 | - | Pseudomonas sp. | - | - | P. danubii | [14] |
| JDS08PS003 | CP120724 | Pseudomonas sp. PSNew6 | - | - | Pseudomonas sp. | [12,13] |
| rDWA16 | - | Pseudomonas sp. | - | - | Pseudomonas sp. | [14] |
| rDWA64 | - | Pseudomonas sp. | - | - | Pseudomonas sp. | [14] |
| Closely related type strains in the Pseudomonas chlororaphis phylogenetic subgroup | ||||||
| CMAA 1215 T | GCA_000474765.1 | “P. aestus” | P. aestus | inconclusive | P. piscis | [15] |
| DSM 19603 T | GCA_003851835.1 | P. chlororaphis ssp. aurantiaca | P. chlororaphis ssp. aurantiaca | OK; type material | P. chlororaphis ssp. aurantiaca | [17] |
| DSM 6698 T | GCA_003851905.1 | P. chlororaphis ssp. aureofaciens | P. chlororaphis ssp. aureofaciens | OK; type material | P. chlororaphis ssp. aureofaciens | [17] |
| DSM 50083 T | GCA_016803445.1 | P. chlororaphis ssp. chlororaphis | P. chlororaphis ssp. chlororaphis | OK; type material | P. chlororaphis ssp. chlororaphis | [17] |
| DSM 21509 T | GCF_001269555.1 | P. chlororaphis ssp. piscium | P. chlororaphis ssp. piscium | OK; type material | P. chlororaphis ssp. piscium | [18] |
| DSM 19095 T | GCA_000397205.1 | P. protegens | P. protegens | OK; type material | P. protegens | [19] |
| DSM 9751 T | GCA_900105185.1 | P. saponiphila | P. saponiphila | OK; type material | P. saponiphila | [20] |
| KCTC 22518T | not available | P. sesami | P. sesami | P. sesami | [21] | |
| CMR12a T | GCA_019139855.1 | P. sessilinigenes | P. sessilinigenes | OK; type material | P. sessilinigenes | [9] |
| MC042 T | GCA_009380155.1 | P. piscis | P. piscis | inconclusive | P. piscis | [16] |
| Closely related strains in public databases of genome sequences | ||||||
| BIOMIG1BAC | GCA_001705995.1 | Pseudomonas_E sp001705835 | Pseudomonas sp. | OK | P. sessilinigens | NCBI web page |
| FD6 | GCA_003363755.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| 11 | GCA_002891565.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| WS5414 | GCA_012985795.1 | Pseudomonas_E protegens_A | Pseudomonas sp. | inconclusive | P. danubii | NCBI web page |
| MB-090624 | GCA_003205455.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| 4 | GCA_002891555.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| 38G2 | GCA_003731885.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| MB-090714 | GCA_003205275.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| 12H11 | GCA_003731825.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| 15H3 | GCA_003731865.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| Go58 | GCA_017347385.1 | Pseudomonas_E protegens_A | P. protegens | inconclusive | P. danubii | NCBI web page |
| B2-1059 | GCF_026016285.1 | not found | Pseudomonas sp. | inconclusive | P. danubii | NCBI web page |
| PS1 | GCA_019754235.1 | not found | P. protegens | inconclusive | P. danubii | NCBI web page |
| 14B2 | GCA_003732485.1 | Pseudomonas_E protegens_B | P. protegens | inconclusive | Pseudomonas sp. | NCBI web page |
| AU11706 | GCA_001020715.1 | Pseudomonas_E fluorescens_AP | P. fluorescens | inconclusive | Pseudomonas sp. | [11] |
| H1F5C | GCA_013407925.2 | Pseudomonas_E fluorescens_AP | P. protegens | inconclusive | Pseudomonas sp. | NCBI web page |
| H1F10A | GCA_013409685.2 | Pseudomonas_E fluorescens_AP | P. protegens | inconclusive | Pseudomonas sp. | NCBI web page |
| Pf-5 | GCA_000012265.1 | Pseudomonas_E protegens | P. protegens | OK | P. protegens | [22] |
| PF-1 | GCA_005887595.1 | Pseudomonas_E protegens | P. protegens | OK | P. protegens | NCBI web page |
| St29 | GCA_001547915.1 | Pseudomonas_E sp001547895 | Pseudomonas sp. | inconclusive | P. sesami | NCBI web page |
| Os17 | GCA_001547895.1 | Pseudomonas_E sp001547895 | Pseudomonas sp. | inconclusive | P. sesami | NCBI web page |
| XYZF4 | GCA_004125385.1 | Pseudomonas_E sp001547895 | P. protegens | inconclusive | P. sesami | NCBI web page |
| BNJ-SS-45 | GCA_003057655.1 | Pseudomonas_E sp001547895 | P. protegens | inconclusive | P. sesami | NCBI web page |
| BC42 | GCA_021560055.1 | not found | Pseudomonas sp. | inconclusive | P. sesami | NCBI web page |
| NFPP19 | GCA_900110785.1 | Pseudomonas_E protegens | Pseudomonas sp. | OK | P. protegens | NCBI web page |
| NFPP12 | GCA_900103205.1 | Pseudomonas_E protegens | Pseudomonas sp. | OK | P. protegens | NCBI web page |
| NFPP10 | GCA_900109535.1 | Pseudomonas_E protegens | Pseudomonas sp. | OK | P. protegens | NCBI web page |
| NFPP05 | GCA_900114815.1 | Pseudomonas_E protegens | Pseudomonas sp. | OK | P. protegens | NCBI web page |
| NFPP08 | GCA_900113795.1 | Pseudomonas_E protegens | Pseudomonas sp. | OK | P. protegens | NCBI web page |
| NFPP09 | GCA_900119575.1 | Pseudomonas_E protegens | Pseudomonas sp. | OK | P. protegens | NCBI web page |
| R26 | GCF_002112545.1 | Pseudomonas_E piscis | Pseudomonas sp. | OK | P. piscis | NCBI web page |
| B6(2017) | GCF_002112765.1 | Pseudomonas_E piscis | Pseudomonas sp. | OK | P. piscis | NCBI web page |
| FW50712TSA | GCF_017350535.1 | Pseudomonas_E piscis | Pseudomonas sp. | OK | P. piscis | NCBI web page |
| FW50714TSA | GCF_017350515.1 | Pseudomonas_E piscis | Pseudomonas sp. | OK | P. piscis | NCBI web page |
| P. danubii JDS02PS016T | P. danubii JDS10PS014 | Pseudomonas sp. JDS08PS003 | |
|---|---|---|---|
| GeneBank ID | CP116502 | CP120725 | CP120724 |
| BioProject | PRJNA922378 | PRJNA922378 | PRJNA922378 |
| BioSample | SAMN32652947 | SAMN33777683 | SAMN33777684 |
| Genome size (bp) | 6,722,326 | 6,746,028 | 7,107,113 |
| GC-content (%) | 62.3 | 62.3 | 62.1 |
| Total genes | 6021 | 6055 | 6357 |
| Protein-coding genes (CDS) | 5931 | 5965 | 6267 |
| RNA genes (clusters) | 90 | 90 | 90 |
| tRNAs | 70 | 70 | 70 |
| Pseudogenes | 58 | 56 | 65 |
| Strain | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| JDS02PS016T | JDS10PS014 | JDS08PS003 | |||||||
| ARO Term | AMR Gene Family | Drug Class | Resistance Mechanism | % Identity of Matching Region | % Length of Reference Sequence | % Identity of Matching Region | % Length of Reference Sequence | % Identity of Matching Region | % Length of Reference Sequence |
| adeF | resistance-nodulation-cell division (RND) antibiotic efflux pump | fluoroquinolone antibiotic, tetracycline antibiotic | antibiotic efflux | 41.48 | 98.87 | 41.48 | 98.87 | 41.38 | 98.87 |
| adeF | 44.02 | 97.45 | 44.02 | 97.45 | 44.59 | 97.45 | |||
| adeF | 67.36 | 100.00 | 67.36 | 100.00 | 67.36 | 100.00 | |||
| adeF | 67.79 | 100.76 | |||||||
| FosA8 | fosfomycin thiol transferase | phosphonic acid antibiotic | antibiotic inactivation | 61.76 | 97.87 | ||||
| FosA | 73.13 | 102.22 | |||||||
| vanG | glycopeptide resistance gene cluster, Van ligase | glycopeptide antibiotic | antibiotic target alteration | 36.91 | 104.30 | 36.91 | 104.30 | 36.91 | 104.30 |
| vanW gene in vanG cluster | glycopeptide antibiotic | antibiotic target alteration | 30.57 | 100.71 | 30.57 | 100.71 | 28.83 | 102.91 | |
| YajC | resistance-nodulation-cell division (RND) antibiotic efflux pump | fluoroquinolone antibiotic, cephalosporin, glycylcycline, penam, tetracycline antibiotic, oxazolidinone antibiotic, glycopeptide antibiotic, rifamycin antibiotic, phenicol antibiotic, disinfecting agents, and antiseptics | antibiotic efflux | 90.18 | 100.00 | 90.18 | 100.00 | 89.29 | 100.00 |
| Characteristics | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NaCl (%) (/v) | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–6 | 0–5 | 0–5 | 0–6 | 0–6 | 0–6 | 0–6 | 0–5 | 0–6 | 0–6 | 0–8 |
| pH | 5–10 | 5–10 | 5–10 | 5–9 | 5–9 | 5–10 | 5–10 | 5–9 | 5–10 | 5–9 | 5–9 | 5–10 | 5–9 | 5–9 | 5–9 | 5–9 | 5–9 | 5–10 | 5–10 | 5–9 | 5–9 | 5–9 |
| API 20 NE test: | ||||||||||||||||||||||
| Reduction of nitrate to nitrite | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| Reduction of nitrite to N2 | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| Hydrolysis of gelatin | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | − |
| Assimilation of: | ||||||||||||||||||||||
| Arabinose | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + |
| Adipate | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | − | − | − | − |
| Phenylacetate | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | − | − |
| BIOLOG GENIII test: | ||||||||||||||||||||||
| Carbon source utilization assays | ||||||||||||||||||||||
| D-Sorbitol | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + |
| p-Hydroxy-Phenylacetic Acid | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | − |
| Tween 40 | w | w | w | w | − | w | + | − | − | w | w | w | w | + | + | − | − | + | + | + | + | + |
| glycyl-L-proline | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + | + | + |
| D-Galacturonic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + |
| Methyl Pyruvate | w | w | w | w | − | w | w | − | − | − | − | − | − | − | w | − | − | w | w | w | + | w |
| D-Arabitol | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + | + | + |
| alfa-Hydroxy Butyric Acid | w | − | − | − | − | w | w | − | − | w | − | w | − | w | − | − | w | w | w | w | w | + |
| D-Trehalose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + |
| D-Galactose | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | + |
| myo-Inositol | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + |
| D-Glucuronic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + |
| alfa-Keto-Butyric Acid | w | − | − | − | − | w | w | − | − | w | − | w | − | + | − | − | − | − | w | + | w | + |
| D-Fucose | w | w | − | − | − | w | w | − | − | w | − | − | − | − | w | − | w | w | w | w | + | + |
| D-glucose-6-PO4 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | − | − |
| Glucuronamide | + | + | + | + | w | w | w | w | w | w | w | − | w | − | + | w | w | + | w | + | + | + |
| Sucrose | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + | + | − | + |
| D-fructose-6-PO4 | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | w | − | w |
| Mucic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | + |
| D-Malic Acid | − | − | − | − | − | − | − | − | − | − | − | w | − | + | − | − | + | − | − | + | + | + |
| L-Rhamnose | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + |
| D-Aspartic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | − | − |
| D-Serine | − | w | − | − | − | − | + | − | − | − | − | − | − | − | − | − | − | + | w | + | + | + |
| D-Saccharic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | + |
| Bromo-Succinic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | w | w | + | + |
| Formic Acid | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | + | + | + | + | + |
| Chemical sensitivity assays | ||||||||||||||||||||||
| Sodium Butyrate | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | + | − | + | w | + |
| 8% NaCl | w | w | w | + | w | w | w | + | − | w | + | + | w | − | + | + | + | + | − | − | − | + |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mulet, M.; Martínez, M.J.; Gomila, M.; Dabernig-Heinz, J.; Wagner, G.E.; Kittinger, C.; Zarfel, G.; Lalucat, J.; García-Valdés, E. Genome-Based Species Diversity Assessment in the Pseudomonas chlororaphis Phylogenetic Subgroup and Proposal of Pseudomonas danubii sp. nov. Isolated from Freshwaters, Soil, and Rhizosphere. Diversity 2023, 15, 617. https://doi.org/10.3390/d15050617
Mulet M, Martínez MJ, Gomila M, Dabernig-Heinz J, Wagner GE, Kittinger C, Zarfel G, Lalucat J, García-Valdés E. Genome-Based Species Diversity Assessment in the Pseudomonas chlororaphis Phylogenetic Subgroup and Proposal of Pseudomonas danubii sp. nov. Isolated from Freshwaters, Soil, and Rhizosphere. Diversity. 2023; 15(5):617. https://doi.org/10.3390/d15050617
Chicago/Turabian StyleMulet, Magdalena, María José Martínez, Margarita Gomila, Johanna Dabernig-Heinz, Gabriel E. Wagner, Clemens Kittinger, Gernot Zarfel, Jorge Lalucat, and Elena García-Valdés. 2023. "Genome-Based Species Diversity Assessment in the Pseudomonas chlororaphis Phylogenetic Subgroup and Proposal of Pseudomonas danubii sp. nov. Isolated from Freshwaters, Soil, and Rhizosphere" Diversity 15, no. 5: 617. https://doi.org/10.3390/d15050617
APA StyleMulet, M., Martínez, M. J., Gomila, M., Dabernig-Heinz, J., Wagner, G. E., Kittinger, C., Zarfel, G., Lalucat, J., & García-Valdés, E. (2023). Genome-Based Species Diversity Assessment in the Pseudomonas chlororaphis Phylogenetic Subgroup and Proposal of Pseudomonas danubii sp. nov. Isolated from Freshwaters, Soil, and Rhizosphere. Diversity, 15(5), 617. https://doi.org/10.3390/d15050617