



Use of Formalin-Preserved Collections to Infer Trophic Indicators of Marine Zooplankton from Stable Isotopes



Abstract
1. Introduction
2. Materials and Methods
2.1. Zooplankton Sampling
2.2. Stable Isotope Analysis
2.3. Statistical Analysis
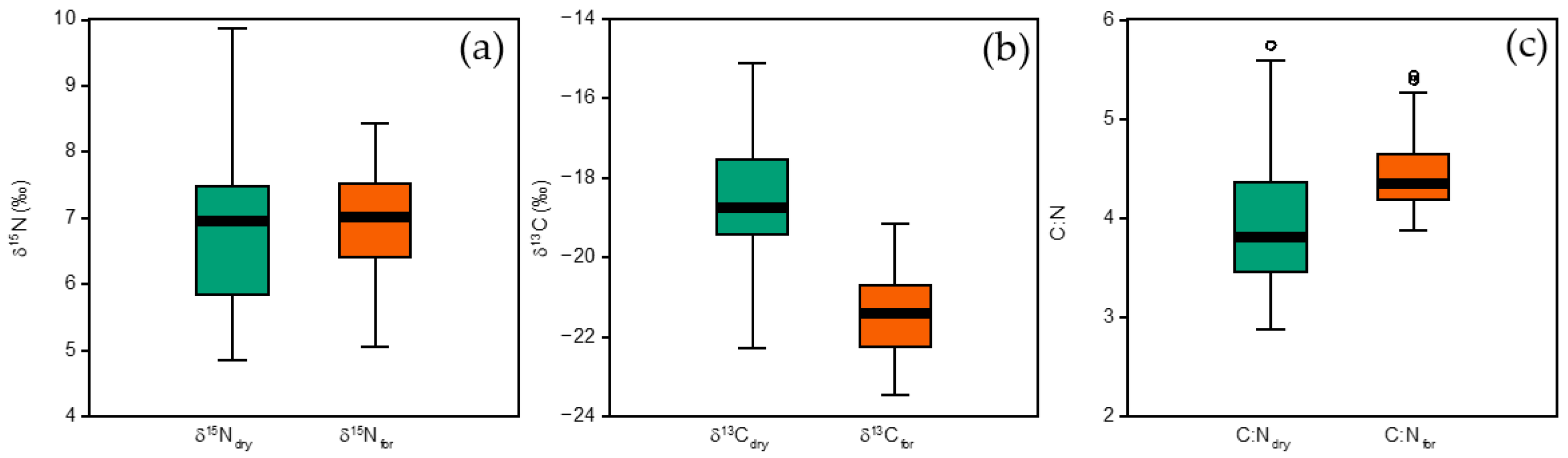
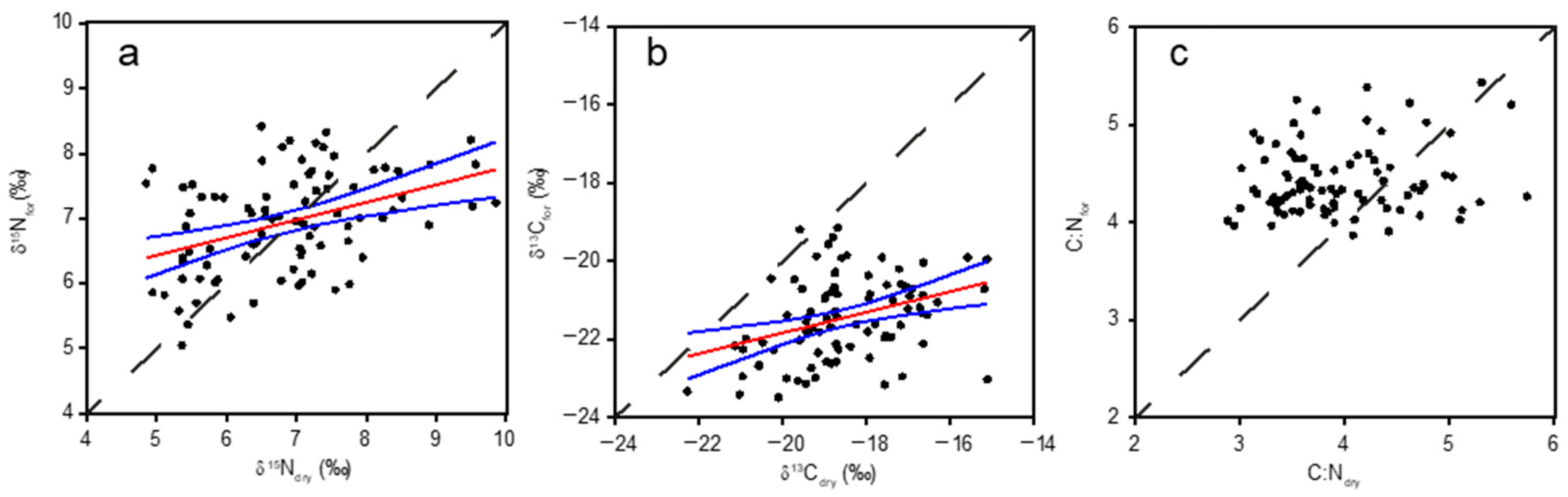
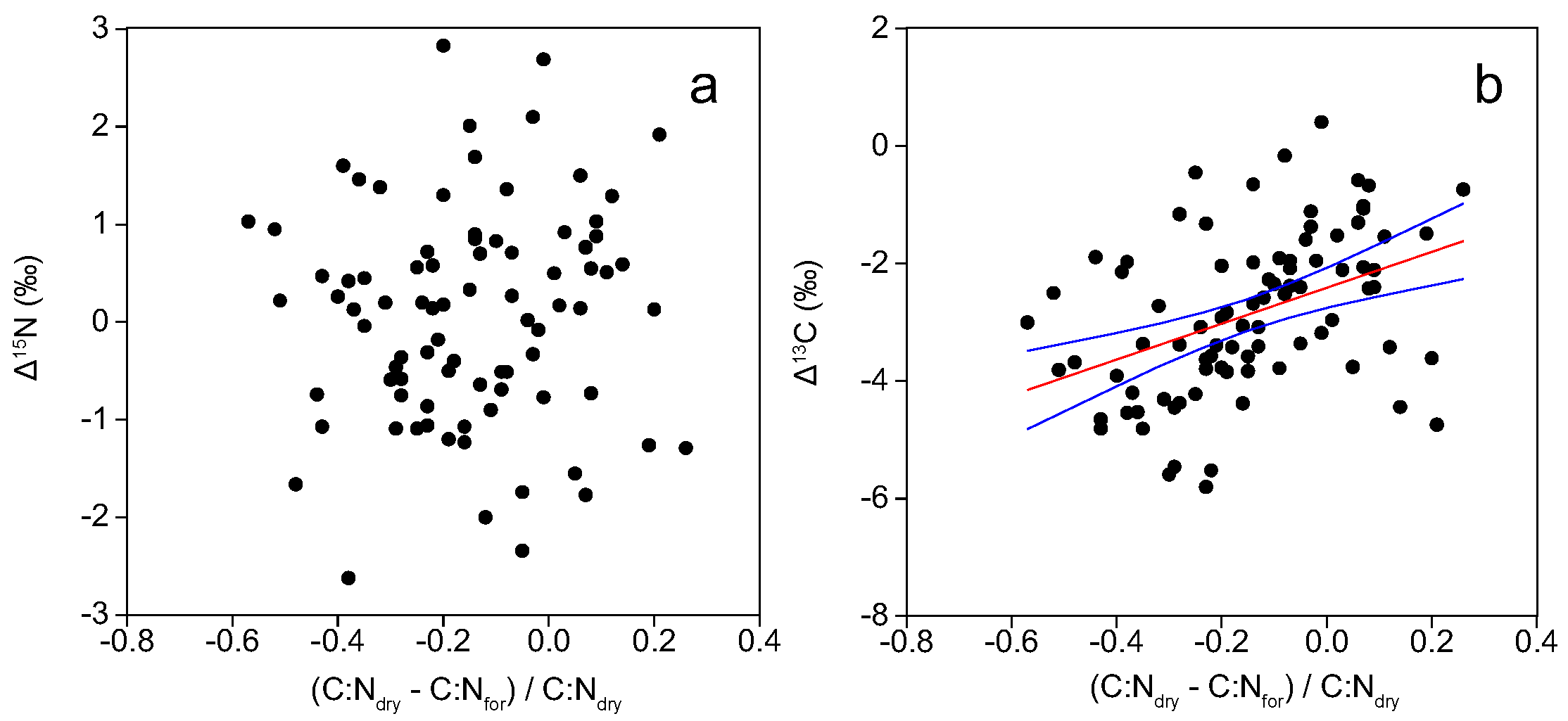
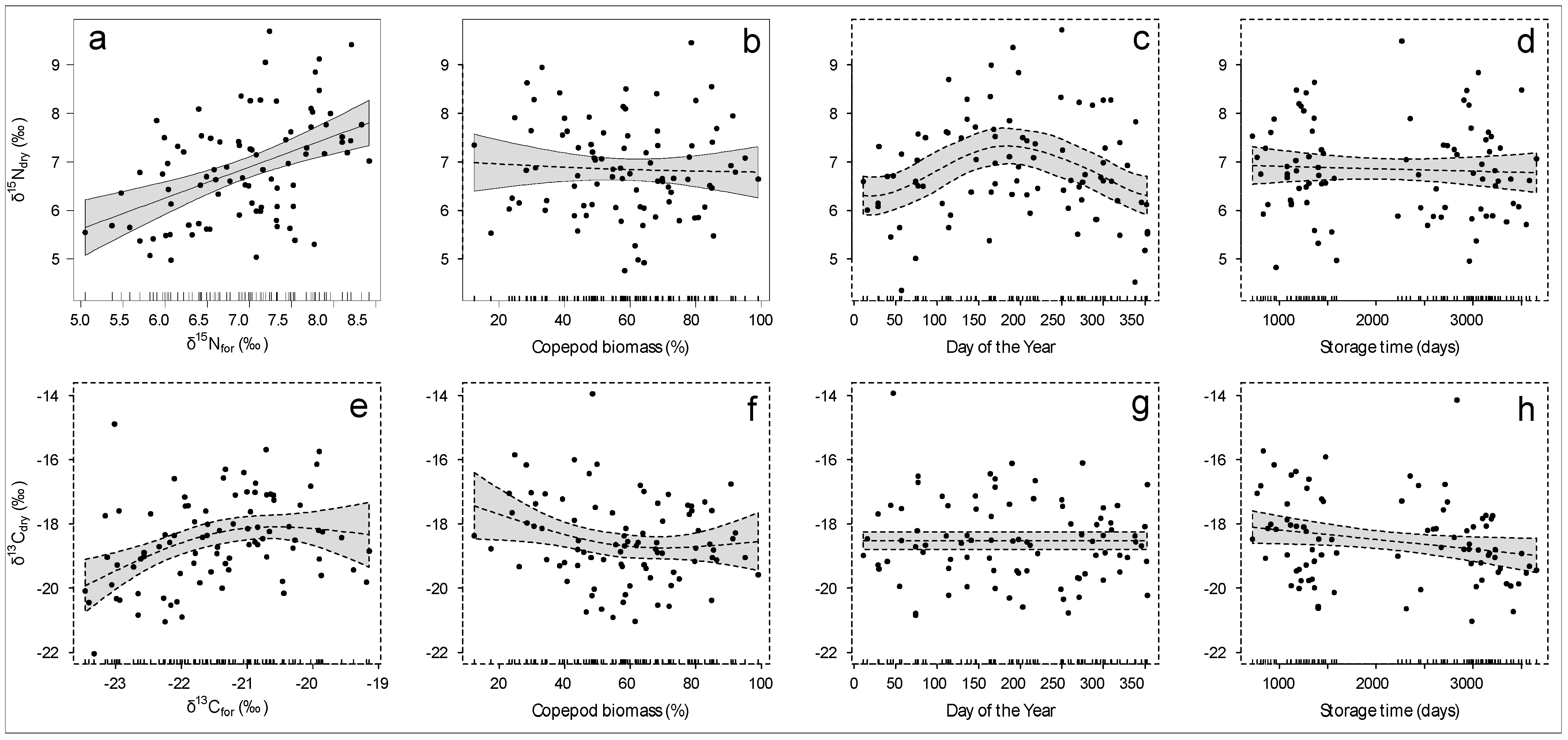
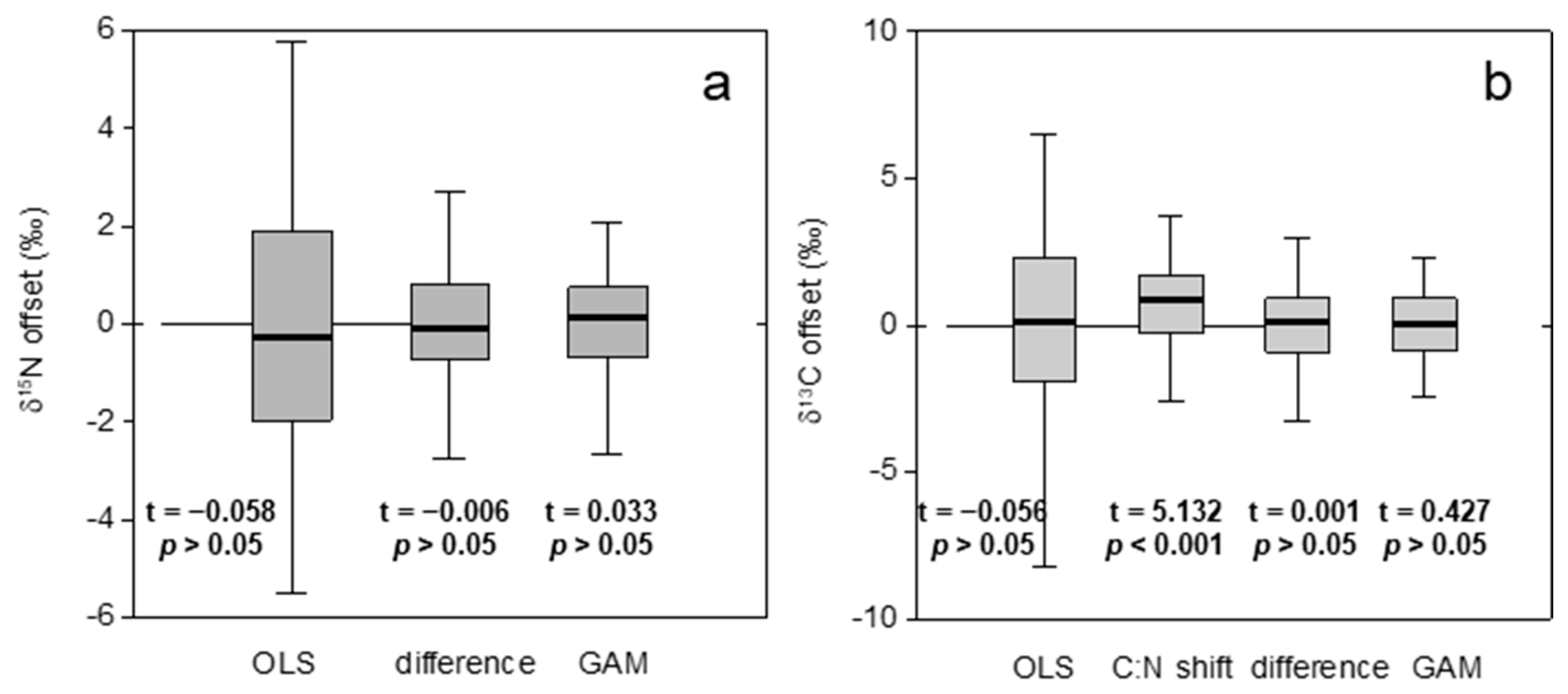
3. Results
3.1. Comparison of Formalin vs. Dry Sample Measurements
3.2. Models for Correction of Measurements in Formalin
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; et al. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Cooley, R.I.; Gerrodette, T.; Fiedler, P.C.; Chivers, S.J.; Danil, K.; Ballance, L.T. Temporal variation in pelagic food chain length in response to environmental change. Sci. Adv. 2017, 3, e1701140. [Google Scholar] [CrossRef] [PubMed]
- Barton, A.D.; Finkel, Z.V.; Ward, B.A.; Johns, D.G.; Follows, M.J. On the roles of cell size and trophic strategy in North Atlantic diatom and dinoflagellate communities. Limnol. Oceanogr. 2013, 58, 254–266. [Google Scholar] [CrossRef]
- Malone, T.C.; Azzaro, M.; Bode, A.; Brown, E.; Duce, R.; Kamykowski, D.; Kang, S.H.; Kedong, Y.; Thorndyke, M.; Wang, J.; et al. Primary production, cycling of nutrients, surface layer and plankton. In The First Global Integrated Marine Assessment: World Ocean; Cambridge University Press: Cambridge, UK, 2017; pp. 119–148. [Google Scholar]
- Walther, G.-R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Reum, J.C.P.; Jennings, S.; Hunsicker, M.E. Implications of scaled δ15N fractionation for community predator-prey body mass ratio estimates in size-structured food webs. J. Anim. Ecol. 2015, 84, 1618–1627. [Google Scholar] [CrossRef]
- Ruiz-Cooley, R.I.; Koch, P.L.; Fiedler, P.C.; McCarthy, M.D. Carbon and Nitrogen Isotopes from Top Predator Amino Acids Reveal Rapidly Shifting Ocean Biochemistry in the Outer California Current. PLoS ONE 2014, 9, e110355. [Google Scholar] [CrossRef]
- Rau, G.H.; Ohman, M.D.; Pierrot-Bults, A. Linking nitrogen dynamics to climate variability off central California: A 51 year based on 15N/14N in CalCOFI zooplankton. Deep Sea Res. II 2003, 50, 2431–2447. [Google Scholar] [CrossRef]
- El-Sabaawi, R.W.; Trudel, M.; Mackas, D.L.; Dower, J.F.; Mazumder, A. Interannual variability in bottom-up processes in the upstream range of the California Current system: An isotopic approach. Prog. Oceanogr. 2012, 106, 16–27. [Google Scholar] [CrossRef]
- Chiba, S.; Sugisaki, H.; Kuwata, A.; Tadokoro, K.; Kobari, T.; Yamaguchi, A.; Mackas, D.L. Pan-NorthPacific comparison of long-term variation in Neocalanus copepods based on stable isotope analysis. Prog. Oceanogr. 2012, 97, 63–75. [Google Scholar] [CrossRef]
- Bicknell, A.W.J.; Campbell, M.; Knight, M.E.; Bilton, D.T.; Newton, J.; Votier, S.C. Effects of formalin preservation on stable carbon and nitrogen isotope signatures in Calanoid copepods: Implications for the use of Continuous Plankton Recorder Survey samples in stable isotope analyses. Rapid Commun. Mass Spectrom. 2011, 25, 1794–1800. [Google Scholar] [CrossRef]
- Mullin, M.M.; Rau, G.H.; Eppley, R.W. Stable nitrogen isotopes in zooplankton: Some geographic and temporal variations in the North Pacific. Limnol. Oceanogr. 1984, 29, 1267–1273. [Google Scholar] [CrossRef]
- Hannides, C.C.S.; Popp, B.N.; Landry, M.R.; Graham, B.S. Quantification of zooplankton trophic position in the North Pacific Subtropical Gyre using stable nitrogen isotopes. Limnol. Oceanogr. 2009, 54, 50–61. [Google Scholar] [CrossRef]
- De Lecea, A.M.; Cooper, R.; Omarjee, A.; Smit, A.J. The effects of preservation methods, dyes and acidification on the isotopic values (δ15N and δ13C) of two zooplankton species from the KwaZulu-Natal Bight, South Africa. Rapid Commun. Mass Spectrom. 2011, 25, 1853–1861. [Google Scholar] [CrossRef]
- Hetherington, E.D.; Kurle, C.M.; Ohman, M.D.; Popp, B.N. Effects of chemical preservation on bulk and amino acid isotope ratios of zooplankton, fish, and squid tissues. Rapid Commun. Mass Spectrom. 2019, 33, 935–945. [Google Scholar] [CrossRef]
- Ventura, M.; Jeppesen, E. Effects of fixation on freshwater invertebrate carbon and nitrogen isotope composition and its arithmetic correction. Hydrobiologia 2009, 632, 297–308. [Google Scholar] [CrossRef]
- Feuchtmayr, H.; Grey, J. Effect of preparation and preservation procedures on carbon and nitrogen stable isotope determinations from zooplankton. Rapid Commun. Mass Spectrom. 2003, 17, 2605–2610. [Google Scholar] [CrossRef]
- Beaugrand, G.; Reid, P.C.; Ibañez, F.; Lindley, J.A.; Edwards, M. Reorganization of North Atlantic Marine Copepod Biodiversity and Climate. Science 2002, 296, 1692–1694. [Google Scholar] [CrossRef]
- Reid, P.C.; Hari, R.E.; Beaugrand, G.; Livingstone, D.M.; Marty, C.; Straile, D.; Barichivich, J.; Goberville, E.; Adrian, R.; Aono, Y.; et al. Global impacts of the 1980s regime shift. Glob. Change Biol. 2016, 22, 682–703. [Google Scholar] [CrossRef]
- Bode, A.; Álvarez, M.; García García, L.M.; Louro, M.A.; Nieto-Cid, M.; Ruíz-Villarreal, M.; Varela, M.M. Climate and local hydrography underlie recent regime shifts in plankton communities off Galicia (NW Spain). Oceans 2020, 1, 181–197. [Google Scholar] [CrossRef]
- Valdés, L.; Bode, A.; Latasa, M.; Nogueira, E.; Somavilla, R.; Varela, M.M.; González-Pola, C.; Casas, G. Three decades of continuous ocean observations in North Atlantic Spanish waters: The RADIALES time series project, context, achievements and challenges. Prog. Oceanog. 2021, 198, 102671. [Google Scholar] [CrossRef]
- Bode, A.; Alvarez-Ossorio, M.T.; Miranda, A.; López-Urrutia, A.; Valdés, L. Comparing copepod time-series in the north of Spain: Spatial autocorrelation of community composition. Prog. Oceanogr. 2012, 97–100, 108–119. [Google Scholar] [CrossRef]
- Razouls, C.; Desreumaux, N.; Kouwenberg, J.; de Bovée, F. Biodiversity of Marine Planktonic Copepods (Morphology, Geographical Distribution and Biological Data). 2005–2023. Available online: http://copepodes.obs-banyuls.fr/en (accessed on 30 June 2022).
- Acuña, J.L.; Lopez-Urrutia, A.; Colin, S. Data compilation of marine pelagic organism oxygen respiration rates and biomasses. Pangaea 2014. [Google Scholar] [CrossRef]
- Wiebe, P. Functional regression equations for zooplankton displacement volume, wet weight, dry weight, and carbon: A correction. Fish. Bull. 1988, 86, 833–835. Available online: https://spo.nmfs.noaa.gov/node/14372 (accessed on 30 June 2022).
- Bode, A.; Mompeán, C.; Lamas, Á.F. Stable C and N isotope data for mesozooplankton samples unpreserved and preserved in formalin. Pangaea 2021. [Google Scholar] [CrossRef]
- Sokal, R.R.; Rohlf, F.J. Biometry, 2nd ed.; W.H. Freeman: San Francisco, CA, USA, 1981; p. 858. [Google Scholar]
- Smyntek, P.M.; Teece, M.A.; Schulz, K.L.; Thackeray, S.J. A standard protocol for stable isotope analysis of zooplankton in aquatic food web research using mass balance correction models. Limnol. Oceanogr. 2007, 52, 2135–2146. [Google Scholar] [CrossRef]
- Wood, S. Generalized Additive Models: An Introduction with R; Chapman & Hall/CRC Press: Boca Raton, FL, USA, 2006; p. 384. [Google Scholar]
- R Core Team. R: A language and environment for statistical computing. In R Foundation for Statistical Computing; The R Foundation: Vienna, Austria, 2022; ISBN 3-900051-07-0. Available online: http://R-project.org (accessed on 30 June 2022).
- Sano, M.; Makabe, R.; Kurosawa, N.; Moteki, M.; Odate, T. Effects of Lugol’s iodine on long-term preservation of marine plankton samples for molecular and stable carbon and nitrogen isotope analyses. Limnol. Oceanogr. Methods 2020, 18, 635–643. [Google Scholar] [CrossRef]
- Hobson, K.A.; Gibbs, H.L.; Gloutney, M.L. Preservation of blood and tissue samples for stable-carbon and stable-nitrogen isotope analysis. Can. J. Zool.-Rev. Can. Zool. 1997, 75, 1720–1723. [Google Scholar] [CrossRef]
- Bosley, K.L.; Wainright, S.C. Effects of preservatives and acidification on the stable isotope ratios (15N:14N, 13C:12C) of two species of marine animals. Can. J. Fish. Aquat. Sci. 1999, 56, 2181–2185. [Google Scholar] [CrossRef]
- Sarakinos, H.C.; Johnson, M.L.; Vander Zanden, M.J. A synthesis of tissue-preservation effects on carbon and nitrogen stable isotope signatures. Can. J. Zool. 2002, 80, 381–387. [Google Scholar] [CrossRef]
- Valdés, L.; López-Urrutia, A.; Beaugrand, G.; Harris, R.P.; Irigoien, X. Seasonality and interannual variability of copepods in the Western English Channel, Celtic Sea, Bay of Biscay, and Cantabrian Sea with a special emphasis to Calanus helgolandicus and Acartia clausi. ICES J. Mar. Sci. 2022, 79, 727–740. [Google Scholar] [CrossRef]
- Le Bourg, B.; Lepoint, G.; Michel, L.C.N. Effects of preservation methodology on stable isotope compositions of sea stars. Rapid Commun. Mass Spectrom. 2020, 34, e8589. [Google Scholar] [CrossRef]
- Durante, L.M.; Sabadel, A.J.M.; Frew, R.D.; Ingram, T.; Wing, S.R. Effects of fixatives on stable isotopes of fish muscle tissue: Implications for trophic studies on preserved specimens. Ecol. Appl. 2020, 30, e02080. [Google Scholar] [CrossRef]
- Willert, M.S.; France, C.A.M.; Brooks, B.L.; Baldwin, C.C.; Hay, M.E. Effects of formalin preservation on carbon and nitrogen stable isotopes of seaweeds: A foundation for looking back in time. Limnol. Oceanogr. Methods 2020, 18, 717–724. [Google Scholar] [CrossRef]
- Welicky, R.L.; Rolfe, T.; Leazer, K.; Maslenikov, K.P.; Tornabene, L.; Holtgrieve, G.W.; Wood, C.L. Fluid-preserved fishes are one solution for assessing historical change in fish trophic level. Ecol. Evol. 2021, 11, 415–426. [Google Scholar] [CrossRef]
- Silberberger, M.J.; Koziorowska-Makuch, K.; Kulinski, K.; Kedra, M. Stable isotope mixing models are biased by the choice of sample preservation and pre-treatment: Implications for studies of aquatic food webs. Front. Mar. Sci. 2021, 7, 621978. [Google Scholar] [CrossRef]
- Carretero, O.; Gago, J.; Filgueiras, A.V.; Viñas, L. The seasonal cycle of micro and meso-plastics in surface waters in a coastal environment (Ría de Vigo, NW Spain). Sci. Total Environ. 2022, 803, 150021. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| x | y | a | sea | b | seb | r | r2 | p |
|---|---|---|---|---|---|---|---|---|
| δ15Ndry | δ15Nfor | 5.075 | 0.463 | 0.272 | 0.067 | 0.404 | 0.164 | 0.000 |
| δ13Cdry | δ13Cfor | −16.575 | 1.396 | 0.263 | 0.075 | 0.355 | 0.126 | 0.001 |
| C:Ndry | C:Nfor | ---- | ---- | ---- | ---- | 0.168 | 0.028 | 0.121 |
| Δ15N | (C:Ndry − C:Nfor)/C:Ndry | ---- | ---- | ---- | ---- | 0.069 | 0.005 | 0.528 |
| Δ13C | (C:Ndry − C:Nfor)/C:Ndry | −2.443 | 0.184 | 3.216 | 0.780 | 0.408 | 0.167 | 0.000 |
| Isotope | Parameter | Estimate (±SE) | EDF | t-Value | F-Value | p |
|---|---|---|---|---|---|---|
| δ15Ndry | Intercept | 6.85 (±0.11) | 64.91 | <0.001 | ||
| N = 87 | δ15Nfor | 1.00 | 20.76 | <0.001 | ||
| r2adj = 0.27 | Copepod biomass | 1.08 | 0.10 | 0.779 | ||
| Day of the Year | 2.10 | 3.40 | 0.001 | |||
| Storage time | 1.00 | 0.20 | 0.659 | |||
| δ13Cdry | Intercept | −18.52 (±0.14) | −133.90 | <0.001 | ||
| N = 87 | δ13Cfor | 1.81 | 7.81 | 0.003 | ||
| r2adj = 0.20 | Copepod biomass | 1.69 | 1.90 | 0.108 | ||
| Day of the Year | 0.00 | 0.00 | 0.746 | |||
| Storage time | 1.00 | 4.04 | 0.048 |
| Variable | Type of Correction | Interquartile Range | 95% Interval |
|---|---|---|---|
| δ15N | OLS | 3.87 | 1.12 |
| difference | 1.52 | 0.47 | |
| GAM | 1.43 | 0.41 | |
| δ13C | OLS | 4.21 | 1.32 |
| C:N shift | 1.99 | 0.59 | |
| difference | 1.85 | 0.58 | |
| GAM | 1.80 | 0.50 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bode, A.; Otero, J.; Lamas, Á.F.; Mompeán, C. Use of Formalin-Preserved Collections to Infer Trophic Indicators of Marine Zooplankton from Stable Isotopes. Diversity 2023, 15, 459. https://doi.org/10.3390/d15030459
Bode A, Otero J, Lamas ÁF, Mompeán C. Use of Formalin-Preserved Collections to Infer Trophic Indicators of Marine Zooplankton from Stable Isotopes. Diversity. 2023; 15(3):459. https://doi.org/10.3390/d15030459
Chicago/Turabian StyleBode, Antonio, Jaime Otero, Ángel F. Lamas, and Carmen Mompeán. 2023. "Use of Formalin-Preserved Collections to Infer Trophic Indicators of Marine Zooplankton from Stable Isotopes" Diversity 15, no. 3: 459. https://doi.org/10.3390/d15030459
APA StyleBode, A., Otero, J., Lamas, Á. F., & Mompeán, C. (2023). Use of Formalin-Preserved Collections to Infer Trophic Indicators of Marine Zooplankton from Stable Isotopes. Diversity, 15(3), 459. https://doi.org/10.3390/d15030459