
Temporal Trends in Skull Morphology of the European Bison from the 1950s to the Present Day



Abstract
1. Introduction
2. Materials and Methods
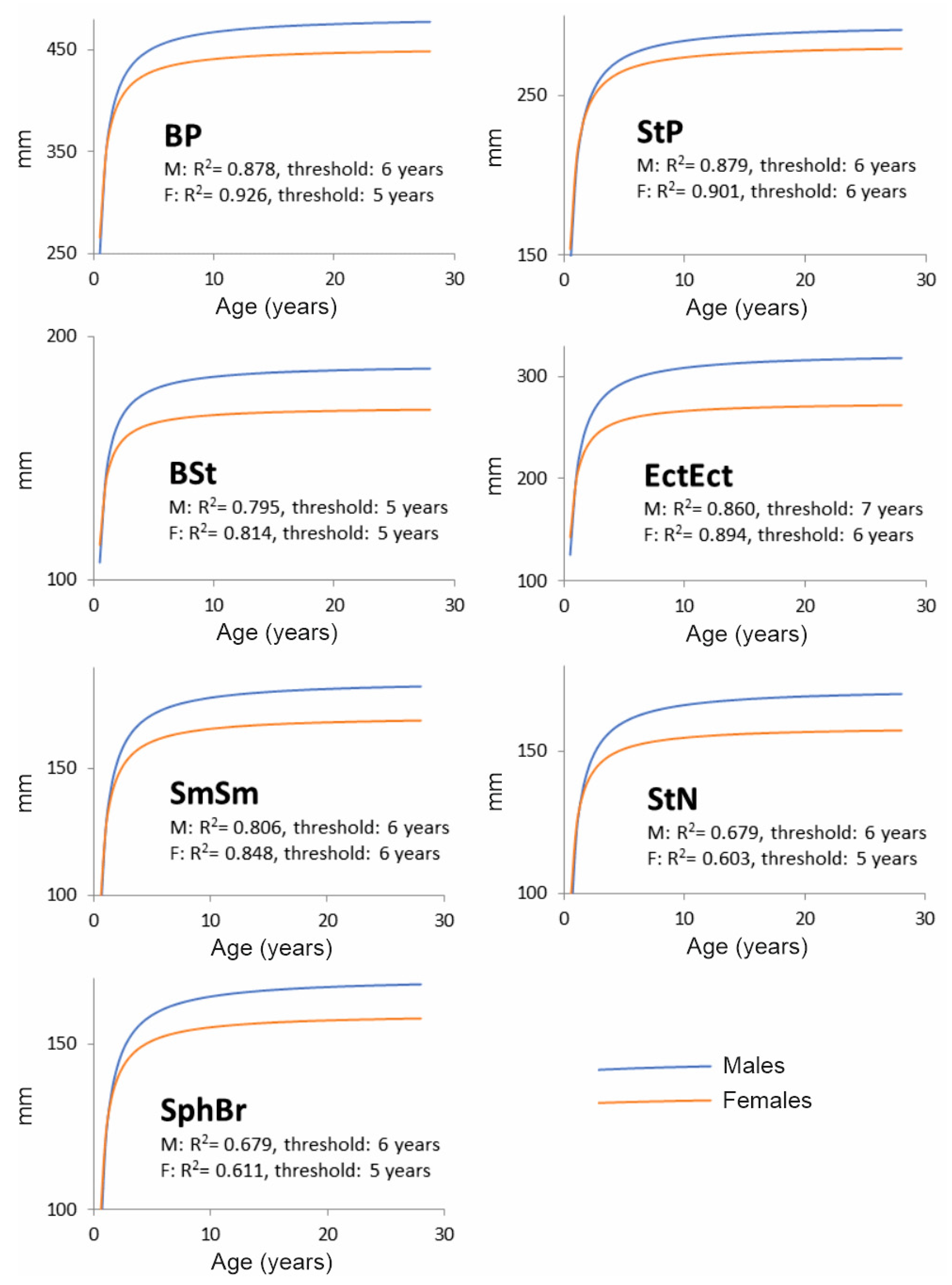
- Basal length: Basion–Prosthion (BP)
- Length of the splanchnocranium: Staphylon–Prosthion (StP)
- Length of the neurocranium: Basion–Staphylon (BSt)
- Orbital breadth: Ectorbitale–Ectorbitale (EctEct)
- Breadth of the splanchnocranium: Supramolare–Supramolare (SmSm)
- Height of the splanchnocranium: Staphylion–Nasion (StN)
- Height of the neurocranium: Sphenobasion–Brema (SphBr)
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hanken, J.; Thorogood, P. Evolution and development of the vertebrate skull: The role of pattern formation. Trends Ecol. Evol. 1993, 8, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Zelditch, M.L.; Lundrigan, B.L.; Garland, T., Jr. Developmental regulation of skull morphology. I. Ontogenetic dynamics of variance. Evol. Dev. 2004, 6, 194–206. [Google Scholar] [CrossRef] [PubMed]
- Logan, C.J.; Clutton-Brock, T.H. Validating methods for estimating endocranial volume in individual red deer (Cervus elaphus). Behav. Process. 2013, 92, 143–146. [Google Scholar] [CrossRef]
- Finarelli, J.A. Estimation of endocranial volume through the use of external skull measures in the Carnivora (Mammalia). J. Mammal. 2006, 87, 1027–1036. [Google Scholar] [CrossRef]
- Brudnicki, W. Comparison of craniometric features and cranial cavity volume in domestic pig (Sus scrofa forma domestica) and wild boar (Sus scrofa) in view of development. Folia Biol. 2005, 53, 25–30. [Google Scholar] [CrossRef]
- Van Cakenberghe, V.; Herrel, A.; Aguirre, L.F. Evolutionary relationships between cranial shape and diet in bats (Mammalia: Chiroptera). In Topics in Functional and Ecological Vertebrate Morphology; Shaker Publishing BV: Maastricht, The Netherlands, 2002; pp. 205–236. ISBN 90-423-0204-6. [Google Scholar]
- Lundrigan, B. Morphology of horns and fighting behavior in the family Bovidae. J. Mammal. 1996, 77, 462–475. [Google Scholar] [CrossRef]
- Kitchener, A. An analysis of the forces of fighting of the blackbuck (Antilope cervicapra) and the bighorn sheep (Ovis canadensis) and the mechanical design of the horn of bovids. J. Zool. 1988, 214, 1–20. [Google Scholar] [CrossRef]
- Kobryńczuk, F.; Krasińska, M.; Szara, T. Sexual dimorphism in skulls of the lowland European bison, Bison bonasus bonasus. Ann. Zool. Fenn. 2008, 45, 335–340. [Google Scholar] [CrossRef]
- Kobryńczuk, F.; Krasińska, M.; Szara, T. Polarization of skull shapes in adult Lowland European bison, Bison bonasus bonasus. Ann. Zool. Fenn. 2008, 45, 341–346. [Google Scholar] [CrossRef]
- Krasińska, M.; Szuma, E.; Kobryńczuk, F.; Szara, T. Morphometric variation of the skull during postnatal development in the Lowland European bison Bison bonasus bonasus. Acta Theriol. 2008, 53, 193–216. [Google Scholar] [CrossRef]
- Krasinska, M. Variability of the skull shape in hybrids between European bison and domestic cattle. Acta Theriol 1988, 33, 147–186. [Google Scholar] [CrossRef]
- Ścibor, J. Hodowla żubrów w Polsce. In Proceedings of the Conferences “European Bison Breeding in Białowieska Forest”; Ministry of Forestry and Wood Industry: Warsaw, Poland, 1965; pp. 19–30. [Google Scholar]
- Pucek, Z. Stan i potrzeby badań nad żubrem. In Proceedings of the Conference “European bison breeding in Białowieska Forest”; PWN: Warsaw, Poland, 1969; pp. 123–135. [Google Scholar]
- Von den Driesch, A. A Guide to the Measurement of Animal Bones from Archaeological Sites: As Developed by the Institut für Palaeoanatomie; Domestikationsforschung und Geschichte der Tiermedizin of the University of Munich, Peabody Museum Press: Münich, Germany, 1976; Volume 1. [Google Scholar]
- Krasińska, M.; Krasiński, Z.A. Body mass and measurements of the European bison during postnatal development. Acta Theriol. 2002, 47, 85–106. [Google Scholar] [CrossRef]
- Kobryńczuk, F. The influence of inbreeding on the shape and size of the skeleton of the European bison. Acta Theriol. 1985, 30, 379–422. [Google Scholar] [CrossRef]
- Empel, W. Morphologie des Schädels von Bison bonasus (Linnaeus 1758). Acta Theriol. 1962, 6, 53–112. [Google Scholar] [CrossRef]
- Preston, B.T.; Stevenson, I.R.; Pemberton, J.M.; Coltman, D.W.; Wilson, K. Overt and covert competition in a promiscuous mammal: The importance of weaponry and testes size to male reproductive success. Proceedings of the Royal Society of London. Ser. B Biol. Sci. 2003, 270, 633–640. [Google Scholar] [CrossRef]
- Tidière, M.; Lemaître, J.F.; Pélabon, C.; Gimenez, O.; Gaillard, J.M. Evolutionary allometry reveals a shift in selection pressure on male horn size. J. Evol. Biol. 2017, 30, 1826–1835. [Google Scholar] [CrossRef] [PubMed]
- Martin, J.M.; Mead, J.I.; Barboza, P.S. Bison body size and climate change. Ecol. Evol. 2018, 8, 4564–4574. [Google Scholar] [CrossRef]
- Palmer, A.R. Waltzing with asymmetry. BioScience 1996, 46, 518–532. [Google Scholar] [CrossRef]
- Siegal, M.L.; Bergman, A. Waddington’s canalization revisited: Developmental stability and evolution. Proc. Natl. Acad. Sci. USA 2002, 99, 10528–10532. [Google Scholar] [CrossRef]
- Debat, V.; David, P. Mapping phenotypes: Canalization, plasticity and developmental stability. Trends Ecol. Evol. 2001, 16, 555–561. [Google Scholar] [CrossRef]
- Breno, M.; Leirs, H.; Van Dongen, S. No relationship between canalization and developmental stability of the skull in a natural population of Mastomys natalensis (Rodentia: Muridae). Biol. J. Linn. Soc. 2011, 104, 207–216. [Google Scholar] [CrossRef]
- Baranov, A.S.; Pucek, Z.; Kiseleva, E.G.; Zakharov, V.M. Developmental stability of skull morphology in European bison Bison bonasus. Acta Theriol. 1997, 42, 79–85. [Google Scholar] [CrossRef]
- Baranov, A.S.; Zakharov, V.M. Developmental stability in hybrids of European bison Bison bonasus and domestic cattle. Acta Theriol. 1997, 42, 87–90. [Google Scholar] [CrossRef]
- Cardini, A.; Polly, D.; Dawson, R.; Milne, N. Why the long face? Kangaroos and wallabies follow the same ‘rule’ of cranial evolutionary allometry (CREA) as placentals. Evol. Biol. 2015, 42, 169–176. [Google Scholar] [CrossRef]
- Jędrzejewski, W.; Jędrzejewska, B.; Okarma, H.; Ruprecht, A.L. Wolf predation and snow cover as mortality factors in the ungulate community of the Bialowieża National Park, Poland. Oecologia 1992, 90, 27–36. [Google Scholar] [CrossRef]
- Benítez-López, A.; Santini, L.; Gallego-Zamorano, J.; Milá, B.; Walkden, P.; Huijbregts, M.A.; Tobias, J.A. The island rule explains consistent patterns of body size evolution in terrestrial vertebrates. Nat. Ecol. Evol. 2021, 5, 768–786. [Google Scholar] [CrossRef] [PubMed]
- Silva, M.; Downing, J.A. The allometric scaling of density and body mass: A nonlinear relationship for terrestrial mammals. Am. Nat. 1995, 145, 704–727. [Google Scholar] [CrossRef]
- Krasiński, Z. Free living European bisons. Acta Theriol. 1967, 12, 391–405. [Google Scholar] [CrossRef]
- Owczarek, M.; Filipiak, J. Contemporary changes of thermal conditions in Poland, 1951–2015. Bull. Geogr. Phys. Geogr. Ser. 2016, 10, 31–50. [Google Scholar] [CrossRef]
- Sheridan, J.A.; Bickford, D. Shrinking body size as an ecological response to climate change. Nat. Clim. Chang. 2011, 1, 401–406. [Google Scholar] [CrossRef]
- Owen-Smith, N.; Cromsigt, J.P.; Le Roux, E. Smaller ungulates are first to incur imminent extirpation from an African protected area. Biol. Conserv. 2017, 216, 108–114. [Google Scholar] [CrossRef]
- Damuth, J. Interspecific allometry of population density in mammals and other animals: The independence of body mass and population energy-use. Biol. J. Linn. Soc. 1987, 31, 193–246. [Google Scholar] [CrossRef]
- Dirzo, R.; Young, H.S.; Galetti, M.; Ceballos, G.; Isaac, N.J.; Collen, B. Defaunation in the Anthropocene. Science 2014, 345, 401–406. [Google Scholar] [CrossRef] [PubMed]
- Hilbers, J.P.; Schipper, A.M.; Hendriks, A.J.; Verones, F.; Pereira, H.M.; Huijbregts, M.A.J. An allometric approach to quantify the extinction vulnerability of birds and mammals. Ecology 2016, 97, 615–626. [Google Scholar] [CrossRef]
- Cardillo, M.; Mace, G.M.; Jones, K.E.; Bielby, J.; Bininda-Emonds, O.R.P.; Sechrest, W.; Orme, C.D.L.; Purvis, A. Multiple causes of high extinction risk in large mammal species. Science 2005, 309, 1239–1241. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
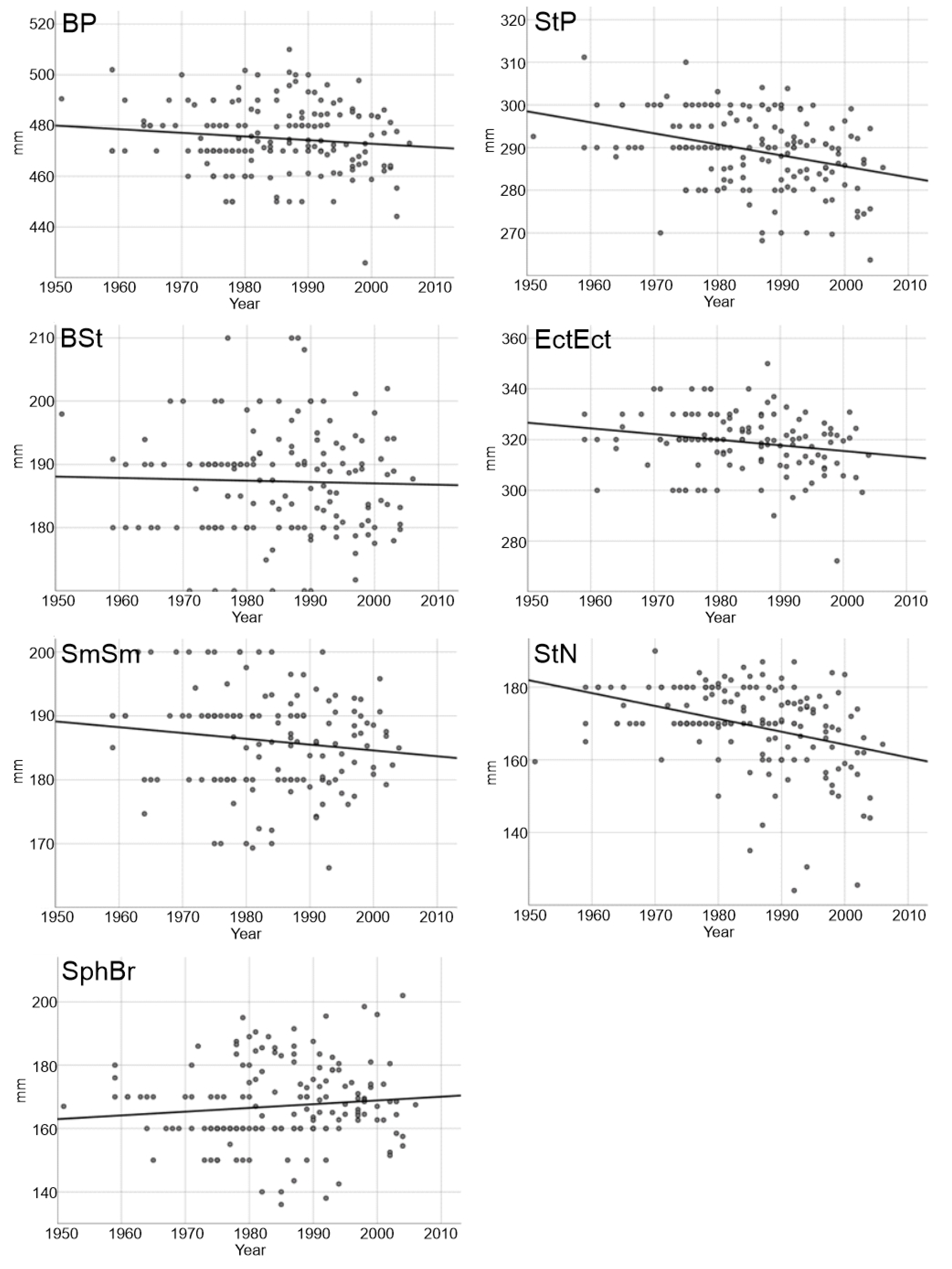
| Model Outputs | BP | StP | BSt | EctEct | SmSm | StN | SphBr |
|---|---|---|---|---|---|---|---|
| F | 2.43 | 19.38 | 0.14 | 5.31 | 2.40 | 23.24 | 1.87 |
| P | 0.121 | 0.000 | 0.710 | 0.023 | 0.123 | 0.000 | 0.173 |
| Intercept [B0 (±SE)] | 760.18 (183.02) | 801.71 (116.37) | 230.24 (115.11) | 761.61 (192.05) | 366.46 (116.41) | 874.08 (146.12) | −67.39 (171.32) |
| Year [B1 (±SE)] | −0.14 (0.09) | −0.26 (0.06) | −0.02 (0.06) | −0.22 (0.10) | −0.09 (0.06) | −0.35 (0.07) | 0.12 (0.09) |
| R2 | 0.015 | 0.107 | 0.001 | 0.040 | 0.017 | 0.124 | 0.011 |
| N | 165 | 163 | 160 | 128 | 143 | 166 | 167 |
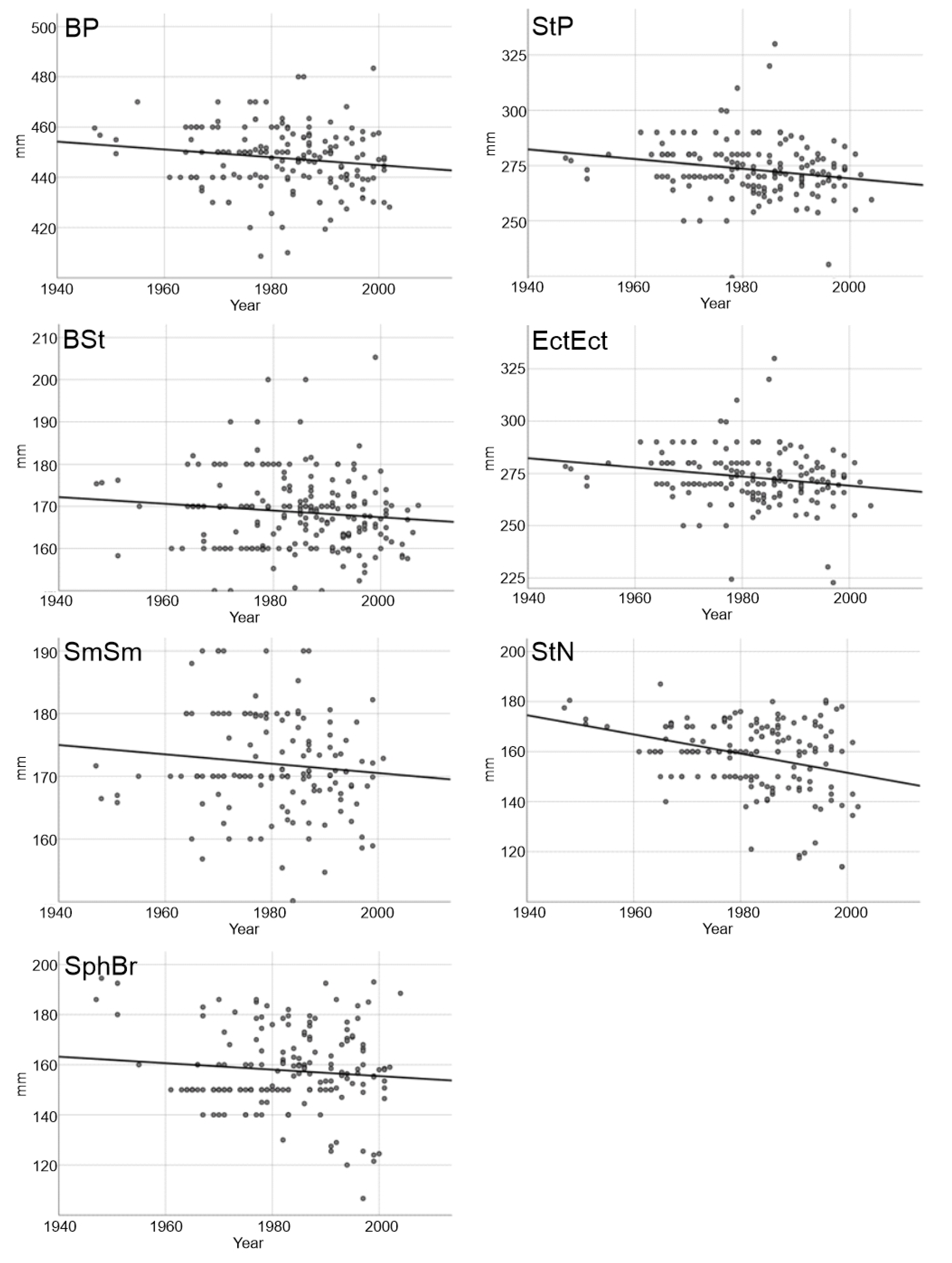
| Model Outputs | BP | StP | BSt | EctEct | SmSm | StN | SphBr |
|---|---|---|---|---|---|---|---|
| F | 3.90 | 7.57 | 2.53 | 5.89 | 1.63 | 17.78 | 1.62 |
| P | 0.050 | 0.007 | 0.110 | 0.016 | 0.203 | 0.000 | 0.204 |
| Intercept [B0 (±SE)] | 755.97 (156.12) | 584.66 (110.77) | 327.08 (99.53) | 707.08 (178.77) | 319.53 (115.44) | 918.03 (180.13) | 413.36 (200.71) |
| Year [B1 (±SE)] | −0.16 (0.08) | −0.15 (0.06) | −0.08 (0.05) | −0.22 (0.09) | −0.07 (0.06) | −0.38 (0.09) | −0.13 (0.10) |
| R2 | 0.022 | 0.046 | 0.012 | 0.036 | 0.011 | 0.101 | 0.009 |
| N | 175 | 162 | 213 | 163 | 146 | 164 | 178 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szara, T.; Klich, D.; Wójcik, A.M.; Olech, W. Temporal Trends in Skull Morphology of the European Bison from the 1950s to the Present Day. Diversity 2023, 15, 377. https://doi.org/10.3390/d15030377
Szara T, Klich D, Wójcik AM, Olech W. Temporal Trends in Skull Morphology of the European Bison from the 1950s to the Present Day. Diversity. 2023; 15(3):377. https://doi.org/10.3390/d15030377
Chicago/Turabian StyleSzara, Tomasz, Daniel Klich, Anna M. Wójcik, and Wanda Olech. 2023. "Temporal Trends in Skull Morphology of the European Bison from the 1950s to the Present Day" Diversity 15, no. 3: 377. https://doi.org/10.3390/d15030377
APA StyleSzara, T., Klich, D., Wójcik, A. M., & Olech, W. (2023). Temporal Trends in Skull Morphology of the European Bison from the 1950s to the Present Day. Diversity, 15(3), 377. https://doi.org/10.3390/d15030377