

The Relationship between Inbreeding and Fitness Is Different between Two Genetic Lines of European Bison


Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
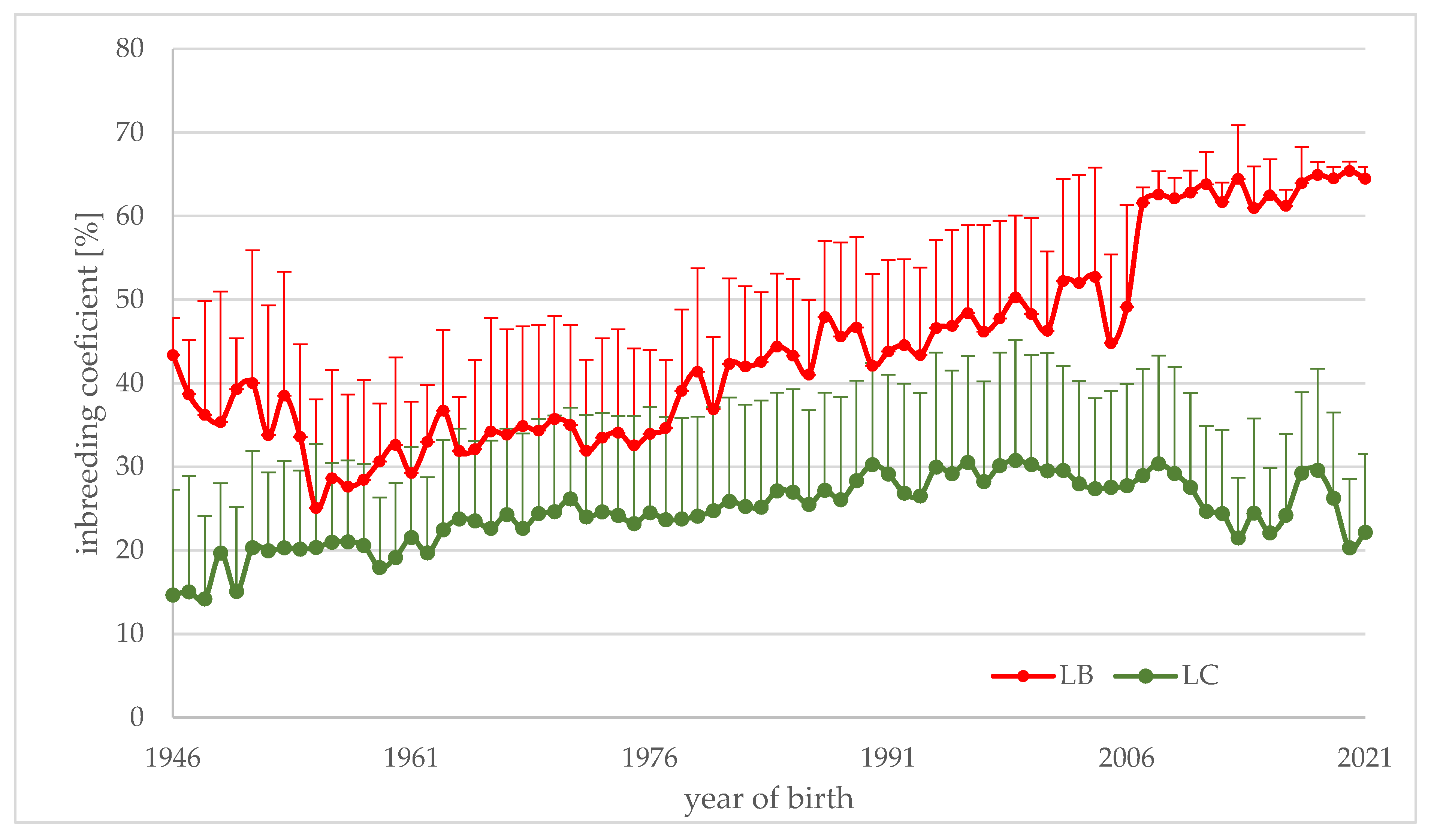
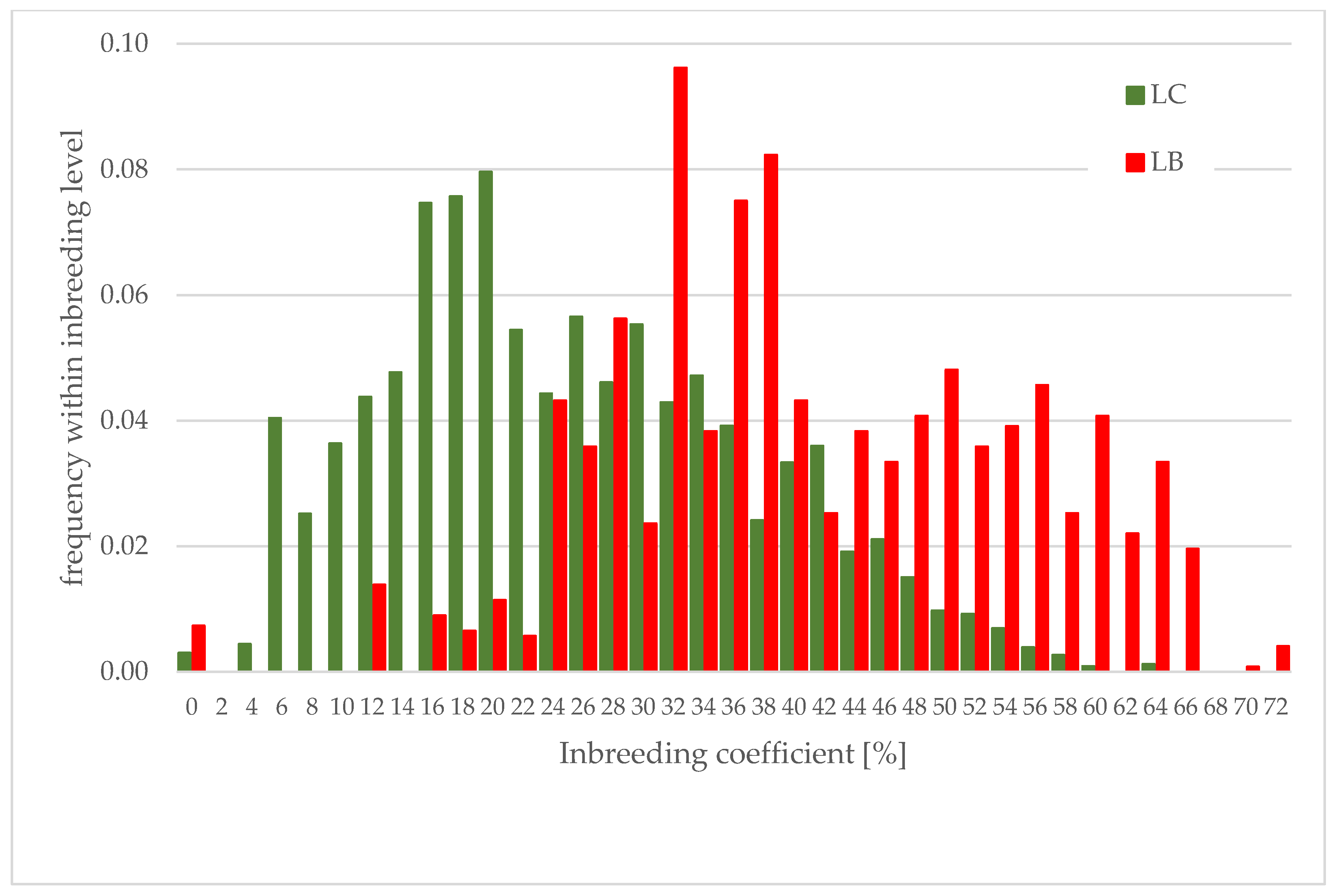
- The European bison, in comparison with other species, is very highly inbred. For animals with a fully known pedigree born in years 1946–2021, the average value of the inbreeding coefficient varies between lines: in the LB line, it is equal to 0.419, and in the LC line, it is 0.253, which is the effect of the different numbers of founders.
- Survival up to one month is negatively influenced by inbreeding only for animals belonging to the LC line. In the LB line, the relation is the opposite, probably as a result of purging in the period of the beginning of the restitution.
- The study of inbreeding and its relation with fitness should be continued.
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Raczyński, J. European Bison; PWRiL: Warszawa, Poland, 1978; pp. 1–246. (In Polish) [Google Scholar]
- Pucek, Z. History of the European bison and problems of its protection and management. In Global Trends in Wildlife Management; Bobek, B., Perzanowski, K., Regelin, W., Eds.; Swiat Press: Krakow-Warszawa, Poland, 1991; pp. 19–39. [Google Scholar]
- von der Groeben, G. Das Zuchtbuch. In Berichte Internationale, Geselschaft zur Erhaltung des Wisents; Kommission bei Dr W. Stichel: Leipzig, Germany, 1932; pp. 5–50. [Google Scholar]
- Raczyński, J.; Bołbot, M. European Bison Pedigree Book 2021; Białowieski National Park: Białowieża, Poland, 2022; pp. 1–80. [Google Scholar]
- Slatis, H.M. An analysis of inbreeding in the European bison. Genetics 1960, 45, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Olech, W. The number of ancestors and their contribution to European bison (Bison bonasus L.) population. Ann. Wars. Agric. Univ. Anim. Sci. 1999, 35, 111–117. [Google Scholar]
- Olech, W. The changes of founders’ numbers and their contribution to the European bison population during 80 years of species’ restitution. Eur. Bison Conserv. Newsl. 2009, 2, 54–60. [Google Scholar]
- Olech, W. The participation of ancestral genes in the existing population of European bison. Acta Theriol. 1989, 34, 397–407. [Google Scholar] [CrossRef]
- Olech, W. The analysis of European bison genetic diversity using pedigree data. In Health Threats for the European Bison Particularly in Free-Roaming Populations in Poland; Kita, J., Anusz, K., Eds.; The SGGW Publishers: Warszawa, Poland, 2006; pp. 205–236. [Google Scholar]
- Parusel, J.B. European bison from Pszczyna and their role in species restitution. Eur. Bison Conserv. Newsl. 2009, 2, 129–136. (In Polish) [Google Scholar]
- Olech, W. Analysis of inbreeding in European bison. Acta Theriol. 1987, 32, 373–387. [Google Scholar] [CrossRef]
- Ballou, J.D. Ancestral inbreeding only minimally affects inbreeding depression in mammalian populations. J. Hered. 1997, 88, 169–178. [Google Scholar] [CrossRef]
- Pucek, Z.; Belousova, I.P.; Krasińska, M.; Krasiński, Z.A.; Olech, W. European Bison Status Survey and Conservation Action Plan; IUCN The World Conservation Union: Gland, Switzerland, 2004; pp. 1–49. [Google Scholar]
- Matuszewska, M.; Olech, W.; Bielecki, W.; Osińska, B. The influence of inbreeding into pathological changes occurrence in European bison males reproduction tract. Park. Nar. Rezerw. Przyr. Polsce 2004, 23, 679–685. (In Polish) [Google Scholar]
- Sobieraj, A.; Olech, W. Twenty years of the European bison Lowland line Bison bonasus bonasus conservation in captivity. Ann. WULS—Anim. Sci. 2018, 57, 171–182. [Google Scholar] [CrossRef]
- Johnson, H.E.; Mills, L.S.; Wehausen, J.D.; Stephenson, T.R.; Luikart, G. Translating effects of inbreeding depression on component vital rates to overall population growth in endangered bighorn sheep. Conserv. Biol. 2011, 25, 1240–1249. [Google Scholar] [CrossRef]
- Reed, T.E.; Grotan, V.; Jenouvrier, S.; Saether, B.-E.; Visser, M.E. Population growth in a wild bird is buffered against phenological mismatch. Science 2013, 340, 488–491. [Google Scholar] [CrossRef] [PubMed]
- Belousova, I.P.; Smirnov, K.A.; Kasmin, V.D.; Kudryavtsev, I.V. Genetic characteristics and prognosis for existence of the European bison free-living population created in the Orlovskoe Poles’e National Park. Russ. J. Genet. 2004, 40, 295–299. [Google Scholar] [CrossRef]
- Olech, W. The Influence of Individual and Maternal Inbreeding on the Survival of European Bison Calves; Warsaw University of Life Sciences: Warsaw, Poland, 2003; pp. 1–78. (In Polish) [Google Scholar]
- Olech, W. The influence of inbreeding on European bison sex ratio. In Animals, Zoos and Conservation; Zgrabczyńska, E., Ćwiertnia, P., Ziomek, J., Eds.; Life Sicience University: Poznań, Poland, 2006; pp. 29–33. [Google Scholar]
- Kobryńczuk, F. The influence of inbreeding on the shape and size of the skeleton of the European bison. Acta Theriol. 1985, 30, 379–422. [Google Scholar] [CrossRef]
- Gill, J. The Physiology of European Bison; Severus: Warszawa, Poland, 1999; pp. 1–176. (In Polish) [Google Scholar]
- Ralls, K.; Ballou, J.D.; Templeton, A. Estimates of lethal equivalents and the cost of inbreeding in mammals. Conserv. Biol. 1988, 2, 185–193. [Google Scholar] [CrossRef]
- Ralls, K.; Brugger, K.; Ballou, J. Inbreeding and juvenile mortality in small populations of ungulates. Science 1979, 206, 1101–1103. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to Conservation Genetics; Cambridge University Press: Cambridge, UK, 2002; pp. 1–140. [Google Scholar]
- Ralls, K.; Ballou, J. Effects of inbreeding on juvenile mortality in some small mammal species. Lab. Anim. 1982, 16, 159–166. [Google Scholar] [CrossRef]
- Frankham, R.; Ralls, K. Inbreeding leads to extinction. Nature 1998, 392, 441–442. [Google Scholar] [CrossRef]
- Saccheri, I.; Kuussaari, M.; Kankare, M.; Vikman, P.; Fortellus, W.; Hanski, I. Inbreeding and extinction in a butterfly metapopulation. Nature 1998, 392, 491–494. [Google Scholar] [CrossRef]
- Laikre, L. Hereditary effects and conservation genetic management of captive population. Zoo Biol. 1999, 18, 81–99. [Google Scholar] [CrossRef]
- Bouwmeester, J.; Mulder, J.L.; van Bree, P.H.J. High incidence of malocclusion in an isolated population of the red fox (Vulpes vulpes) in the Netherland. J. Zool. 1989, 219, 123–136. [Google Scholar] [CrossRef]
- Pemberton, J. Measuring inbreeding depression in the wild: The old ways are the best. Trends Ecol. Evol. 2004, 19, 613–615. [Google Scholar] [CrossRef]
- Slate, J.; Kruuk, L.E.B.; Marshall, T.C.; Pemberton, J.M.; Clutton-Brock, T.H. Inbreeding depression influences lifetime breeding success in a wild population of red deer (Cervus elaphus). Proc. R. Soc. Lond. B 2000, 267, 1657–1662. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P.H.; Garcia-Dorado, A. Understanding inbreeding depression, purging and genetic rescue. Trends Ecol. Evol. 2016, 31, 940–952. [Google Scholar] [CrossRef] [PubMed]
- Ballou, J.D.; Lacy, R.C. Identifying genetically important individuals for management of genetic diversity in pedigreed populations. In Population Management for Survival and Recovery; Ballou, J.D., Foose, T.J., Gilpin, M.E., Eds.; Columbia University Press: New York, NY, USA, 1995; pp. 76–111. [Google Scholar]
- Byers, D.L.; Waller, D.M. Do plant populations purge their genetic load? Effects of population size and mating history on inbreeding depression. Annu. Rev. Ecol. Syst. 1999, 30, 479–513. [Google Scholar] [CrossRef]
- Crnokrak, P.; Barrett, S.C.D. Perspective: Purging the genetic load: A review of the experimental evidence. Evolution 2002, 56, 2347–2358. [Google Scholar] [PubMed]
- Boakes, E.H.; Wang, J.; Amos, W. An investigation of inbreeding depression and purging in captive pedigreed populations. Heredity 2007, 98, 172–182. [Google Scholar] [CrossRef]
- Kalinowski, S.T.; Hedrick, P.W.; Miller, P.S. No inbreeding depression observed in Mexican and red wolf captive breeding programs. Conserv. Biol. 1999, 13, 1371–1377. [Google Scholar] [CrossRef]
- Bouzat, J.l. Conservation genetics of population bottlenecks: The role of chance, selection, and history. Conserv. Genet. 2010, 11, 463–478. [Google Scholar] [CrossRef]
- López-Cortegano, E.; Moreno, E.; García-Dorado, A. Genetic purging in captive endangered ungulates with extremely low effective population sizes. Heredity 2021, 127, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Carr, D.E.; Dudash, M. Recent approaches into the genetic basis of inbreeding depression in plants. Philos. Trans. R. Soc. Lond. B 2003, 358, 1071–1084. [Google Scholar] [CrossRef]
- Wang, J.; Hill, W.G.; Charlesworth, D.; Charlesworth, B. Dynamics of inbreeding depression due to deleterious mutations in small populations: I. Mutation parameters and inbreeding rate. Genet. Res. 1999, 74, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Krasińska, M.; Krasiński, Z.A.; Perzanowski, K.; Olech, W. European bison Bison bonasus (Linnaeus, 1758). In Ecology, Evolution and Behaviour of Wild Cattle: Implications for Conservation; Melleti, M., Burton, J., Eds.; Cambridge University Press: Cambridge, UK, 2014; pp. 115–173. [Google Scholar]
- Krasiński, Z. The restitution of European bison in Białowieża in years 1929–1952. Park. Nar. Rezerw. Przyr. 1994, 4, 3–23. (In Polish) [Google Scholar]
- Kaczmarek-Okrój, M.; Olech, W. Reproduction parameters of wisent in ex situ condition. Eur. Bison Conserv. Newsl. 2022, 14, 29–42. [Google Scholar]
- Krasiński, Z.; Raczyński, J. The reproduction biology of European bison living in reserves and in freedom. Acta Theriol. 1967, 29, 407–444. [Google Scholar] [CrossRef]
- EBPB—European Bison Pedigree Book; URL. 1133. Available online: https://bpn.com.pl/index.php?option=com_content&task=view&id=1133&Itemid=213 (accessed on 20 December 2022).
- Olech, W.; Michalska, E. Comparison of inbreeding in two closed herd. Zwierz. Lab. 1999, 27, 3–8. [Google Scholar]
- Quaas, R.L. Computing the diagonal elements and inverse of a large numerator relationship matrix. Biometrics 1976, 32, 949–953. [Google Scholar] [CrossRef]
- Olech, W.; Perzanowski, K. (Eds.) European Bison (Bison bonasus) Strategic Species Status Review 2020; IUCN SSC Bison Specialist Group and European Bison Conservation Center, European Bison Friends Society: Warsaw, Poland, 2022; pp. 1–138. [Google Scholar]
- Weiser, E.L.; Grueber, C.E.; Kennedy, E.S.; Jamieson, I.G. Unexpected positive and negative effects of continuing inbreeding in one of the world”s most inbred wild animals. Evolution 2015, 70, 154–166. [Google Scholar] [CrossRef] [PubMed]
- Luis, C.; Cothran, E.G.; do Mar Oomm, M. Inbreeding and Genetic Structure in the Endangered Sorraia Horse Breed: Implications for its Conservation and Management. J. Hered. 2007, 98, 232–237. [Google Scholar] [CrossRef]
- Haanes, H.; Markussen, S.S.; Herfindal, I.; Røed, K.H.; Solberg, E.J.; Heim, M.; Midthjell, L.; Sæther, B.E. Effects of inbreeding on fitness-related traits in a small isolated moose population. Ecol. Evol. 2013, 3, 4230–4242. [Google Scholar] [CrossRef]
- Walling, C.A.; Nussey, D.H.; Morris, A.; Clutton-Brock, T.H.; Kruuk, L.E.B.; Pemberton, J.M. Inbreeding depression in red deer calves. Evol. Biol. 2011, 11, 318. [Google Scholar] [CrossRef] [PubMed]
- Farquharson, K.A.; Hogg, C.J.; Grueber, C.E. Offspring survival changes over generations of captive breeding. Nat. Commun. 2021, 12, 3045. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, C.; Zhang, L.; Zhong, Z.; Jiang, Z. No correlation between neonatal fitness and heterozygosity in a reintroduced population of Père David’s deer. Curr. Zool. 2013, 59, 249–256. [Google Scholar] [CrossRef]
- Skotarczak, E.; Ćwiertnia, P.; Szwaczkowski, T. Pedigree structure of American bison (Bison bison) population. Czech J. Anim. Sci. 2018, 63, 507–517. [Google Scholar] [CrossRef]
- Sausman, K.A. Survival of captive born Ovis canadensis in North American zoos. Zoo Biol. 1984, 3, 111–121. [Google Scholar] [CrossRef]
- Zhang, S.; Li, C.; Li, Y.; Chen, Q.; Hu, D.; Cheng, Z.; Wang, X.; Shan, Y.; Bai, J.; Liu, G. Genetic differentiation of reintroduced Père David’s deer (Elaphurus davidianus) based on population genomics analysis. Front Genet. 2021, 12, 705337. [Google Scholar] [CrossRef]
- Zchokke, S.; Baur, B. Inbreeding, outbreeding, infant growth and size dimorphism in captive Indian rhinoceros. Can. J. Zool. 2002, 80, 2014–2023. [Google Scholar] [CrossRef]
- Parusel, J.B. If it had not been for the European bison of Pszczyna…. Eur. Bison Conserv. Newsl. 2019, 12, 69–78. (In Polish) [Google Scholar]
- Templeton, R.A.; Read, B. Elimination of inbreeding depression from a captive populations of Speke’s gazelle. Validity of original statistical analysis and confirmation by permutation testing. Zoo Biol. 1998, 17, 77–94. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Group | Number | Survival to First Month | Males | ||
|---|---|---|---|---|---|
| Number | % | Number | % | ||
| Global population: all registered born until 2021 | 13,750 | 12,404 | 90.2 | 6716 | 48.8 |
| Line LC * | 9801 | 8778 | 89.6 | 4874 | 49.7 |
| Line LB * | 3949 | 3627 | 91.8 | 1842 | 46.6 |
| Analyzed: (with full known pedigree and born 1946–2021) | 6868 | 6051 | 88.1 | 3435 | 50.0 |
| Line LC | 5641 | 4943 | 87.6 | 2849 | 50.5 |
| Line LB | 1227 | 1108 | 90.3 | 586 | 47.8 |
| Line LC | Line LB | Both Lines Together | ||||||
|---|---|---|---|---|---|---|---|---|
| Number | Average | s.d. | Number | Average | s.d. | Average | s.d. | |
| All analyzed animals | 5641 | 0.253 | 0.121 | 1227 | 0.410 | 0.136 | 0.281 | 0.137 |
| F | 2792 | 0.257 | 0.122 | 641 | 0.402 | 0.136 | 0.284 | 0.137 |
| M | 2849 | 0.250 | 0.119 | 586 | 0.419 | 0.135 | 0.279 | 0.137 |
| not surviving until 1st month | 698 | 0.276 | 0.127 | 119 | 0.379 | 0.124 | 0.291 | 0.132 |
| surviving to 1st month | 4943 | 0.250 | 0.119 | 1108 | 0.413 | 0.136 | 0.280 | 0.138 |
| born in Spring | 3546 | 0.248 | 0.119 | 857 | 0.410 | 0.136 | 0.280 | 0.139 |
| born in Autumn | 1776 | 0.260 | 0.121 | 335 | 0.408 | 0.136 | 0.283 | 0.134 |
| born in Winter | 244 | 0.287 | 0.132 | 32 | 0.415 | 0.145 | 0.302 | 0.139 |
| Source | B * | Standard Error | Wald χ2 * | p Value | Exp (B) * |
|---|---|---|---|---|---|
| Intercept | 1.211 | 0.1643 | 54.289 | 0.000 | 3.356 |
| Sex—Female | 0.021 | 0.0749 | 0.075 | 0.784 | 1.021 |
| Season—Spring | 0.992 | 0.1459 | 46.264 | 0.000 | 2.698 |
| Season—Autumn | 0.938 | 0.1534 | 37.365 | 0.000 | 2.555 |
| Inbreeding coefficient | −0.527 | 0.2696 | 3.820 | 0.051 | 0.590 |
| Source | B * | Standard Error | Wald χ2 | p Value | Exp (B) * |
|---|---|---|---|---|---|
| Intercept | 1.477 | 0.1796 | 67.638 | 0.000 | 4.380 |
| Sex—Female | 0.046 | 0.0816 | 0.320 | 0.572 | 1.047 |
| Season—Spring | 0.923 | 0.1549 | 35.490 | 0.000 | 2.516 |
| Season—Autumn | 0.961 | 0.1633 | 34.645 | 0.000 | 2.615 |
| Inbreeding coefficient | −1.658 | 0.3299 | 25.268 | 0.000 | 0.190 |
| Source | B * | Standard Error | Wald χ2 | p Value | Exp (B) * |
|---|---|---|---|---|---|
| Intercept | 0.733 | 0.546 | 1.803 | 0.179 | 2.081 |
| Sex—Female | −0.054 | 0.195 | 0.078 | 0.780 | 0.947 |
| Season—Spring | 0.940 | 0.4722 | 3.964 | 0.046 | 2.560 |
| Season—Autumn | 0.522 | 0.4853 | 1.157 | 0.282 | 1.685 |
| Inbreeding coefficient | 1.882 | 0.7222 | 6.793 | 0.009 | 6.569 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olech, W. The Relationship between Inbreeding and Fitness Is Different between Two Genetic Lines of European Bison. Diversity 2023, 15, 368. https://doi.org/10.3390/d15030368
Olech W. The Relationship between Inbreeding and Fitness Is Different between Two Genetic Lines of European Bison. Diversity. 2023; 15(3):368. https://doi.org/10.3390/d15030368
Chicago/Turabian StyleOlech, Wanda. 2023. "The Relationship between Inbreeding and Fitness Is Different between Two Genetic Lines of European Bison" Diversity 15, no. 3: 368. https://doi.org/10.3390/d15030368
APA StyleOlech, W. (2023). The Relationship between Inbreeding and Fitness Is Different between Two Genetic Lines of European Bison. Diversity, 15(3), 368. https://doi.org/10.3390/d15030368