


The Hidden Diversity of Temperate Mesophotic Ecosystems from Central Chile (Southeastern Pacific Ocean) Assessed through Towed Underwater Videos
 ,
,  ,
, 
Abstract
1. Introduction
2. Materials and Methods
2.1. Sampling Campaign
2.2. Reef Fish Assemblages
2.3. Benthic Community
2.4. Statistical Analyses
3. Results
3.1. Reef Fish Assemblages
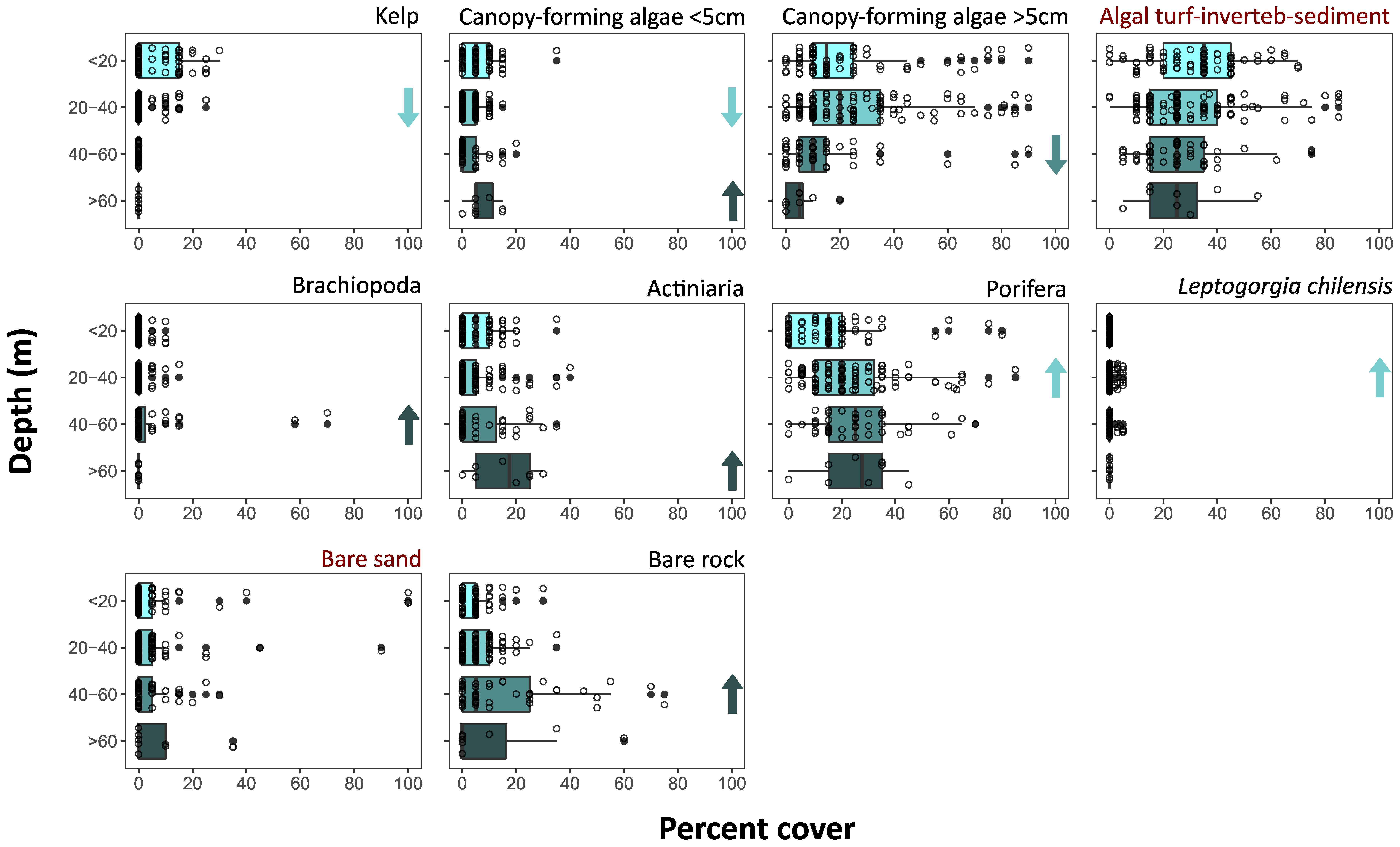
3.2. Benthic Community
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cerrano, C.; Bastari, A.; Calcinai, B.; Di Camillo, C.; Pica, D.; Puce, S.; Valisano, L.; Torsani, F. Temperate mesophotic ecosystems: Gaps and perspectives of an amerging conservation challenge for the Mediterranean Sea. Eur. Zool. J. 2019, 86, 370–388. [Google Scholar] [CrossRef]
- Pyle, R.L.; Copus, J.M. Mesophotic coral ecosystems: Introduction and overview. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer: Cham, Switzerland, 2019; pp. 3–27. [Google Scholar]
- Turner, J.A.; Andradi-Brown, D.A.; Gori, A.; Bongaerts, P.; Burdett, H.L.; Ferrier-Pagès, C.; Voolstra, C.R.; Weinstein, D.K.; Bridge, T.C.L.; Costantini, F.; et al. Key questions for research and conservation of mesophotic coral ecosystems and temperate mesophotic ecosystems. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer: Cham, Switzerland, 2019; pp. 989–1003. [Google Scholar]
- Eyal, G.; Pinheiro, H.T. Mesophotic ecosystems: The link between shallow and deep-sea habitats. Diversity 2020, 12, 411. [Google Scholar] [CrossRef]
- Puglise, K.A.; Hinderstein, L.M.; Marr, J.C.A.; Dowgiallo, M.J.; Martinez, F.A. Mesophotic Coral Ecosystems Research Strategy. In Proceedings of the International Workshop to Prioritize Research and Management Needs for Mesophotic Coral Ecosystems, Jupiter, FL, USA, 12–15 July 2008; NOAA/National Centers for Coastal Ocean Science: Silver Spring, MD, USA; p. 24. [Google Scholar]
- Hinderstein, L.; Marr, J.C.A.; Martinez, F.A.; Dowgiallo, M.J.; Puglise, K.A.; Pyle, R.L.; Zawada, D.G.; Appeldoorn, R. Theme section on “mesophotic coral ecosystems: Characterization, ecology, and management”. Coral Reefs 2010, 29, 247–251. [Google Scholar] [CrossRef]
- Rocha, L.A.; Pinheiro, H.T.; Shepherd, B.; Papastamatiou, Y.P.; Luiz, O.J.; Pyle, R.L.; Bongaerts, P. Mesophotic coral ecosystems are threatened and ecologically distinct from shallow water reefs. Science 2018, 361, 281–284. [Google Scholar] [CrossRef]
- Cerrano, C.; Danovaro, R.; Gambi, C.; Pusceddu, A.; Riva, A.; Schiaparelli, S. Gold coral (Savalia savaglia) and gorgonian forests enhance benthic biodiversity and ecosystem functioning in the mesophotic zone. Biodivers. Conserv. 2010, 19, 153–167. [Google Scholar] [CrossRef]
- Bell, J.J.; Micaroni, V.; Harris, B.; Strano, F.; Broadribb, M.; Rogers, A. Global status, impacts, and management of rocky temperate mesophotic ecosystems. Conserv. Biol. 2022, e13945. [Google Scholar] [CrossRef]
- Micaroni, V.; McAllen, R.; Turner, J.; Strano, F.; Morrow, C.; Picton, B.; Harman, L.; Bell, J.J. Vulnerability of Temperate Mesophotic Ecosystems (TMEs) to environmental impacts: Rapid ecosystem changes at Lough Hyne Marine Nature Reserve, Ireland. Sci. Total Environ. 2021, 789, 147708. [Google Scholar] [CrossRef]
- Shepherd, B.; Phelps, T.; Pinheiro, H.T.; Pérez-Matus, A.; Rocha, L.A. Plectranthias ahiahiata, a new species of perchlet from a mesophotic ecosystem at Rapa Nui (Easter Island) (Teleostei, Serranidae, Anthiadinae). ZooKeys 2018, 762, 105–116. [Google Scholar] [CrossRef]
- Sheperd, B.; Pinheiro, H.T.; Phelps, T.; Pérez-Matus, A.; Rocha, L.A. Luzonichthys kiomeamea (Teleostei: Serranidae: Anthiadinae), a new species from a mesophotic coral ecosystem of Rapa Nui (Easter Island). J. Ocean Sci. Found. 2019, 33, 17–27. [Google Scholar]
- Easton, E.E.; Gorny, M.; Mecho, A.; Sellanes, J.; Gaymer, C.F.; Spalding, H.L.; Aburto, J. Chile and the Salas y Gómez Ridge. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer: Cham, Switzerland, 2019; pp. 477–490. [Google Scholar]
- Hoeksema, B.W.; Sellanes, J.; Easton, E.E. A high-latitude, mesophotic Cycloseris field at 85 m depth off Rapa Nui (Easter Island). Bull. Mar. Sci. 2019, 95, 101–102. [Google Scholar] [CrossRef]
- Shepherd, B.; Pinheiro, H.T.; Phelps, T.A.Y.; Easton, E.E.; Pérez-Matus, A.; Rocha, L.A. A New Species of Chromis (Teleostei: Pomacentridae) from Mesophotic Coral Ecosystems of Rapa Nui (Easter Island) and Salas y Gómez, Chile. Copeia 2020, 108, 326–332. [Google Scholar] [CrossRef]
- Sellanes, J.; Gorny, M.; Zapata-Hernández, G.; Álvarez, G.; Muñoz, P.; Tala, F. A new threat to local marine biodiversity: Filamentous mats proliferating at mesophotic depths off Rapa Nui. PeerJ 2021, 9, e12052. [Google Scholar] [CrossRef]
- Mecho, A.; Dewitte, B.; Sellanes, J.; van Gennip, S.; Easton, E.E.; Gusmao, J.B. Environmental drivers of mesophotic echinoderm assemblages of the southeastern Pacific Ocean. Front. Mar. Sci. 2021, 8, 574780. [Google Scholar] [CrossRef]
- Försterra, G.; Häussermann, V.; Laudien, J. Animal Forests in the Chilean fjords: Discoveries, perspectives and threats in shallow and deep waters. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas, C., Eds.; Springer: Cham, Switzerland, 2016. [Google Scholar]
- Gorny, M.; Easton, E.E.; Sellanes, J. First record of black corals (Antipatharia) in shallow coastal waters of northern Chile by means of underwater video. Lat. Am. J. Aquat. Res. 2018, 46, 457–460. [Google Scholar] [CrossRef]
- Holstein, D.M.; Fletcher, P.; Groves, S.H.; Smith, T.B. Ecosystem services of mesophotic coral ecosystems and a call for better accounting. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer: Cham, Switzerland, 2019; pp. 943–956. [Google Scholar]
- Gómez, F.A.; Spitz, Y.H.; Batchelder, H.P.; Correa-Ramírez, M.A. Intraseasonal patterns in coastal plankton biomass off central Chile derived from satellite observations and a biochemical model. J. Mar. Syst. 2017, 174, 106–118. [Google Scholar] [CrossRef]
- Thiel, M.; Macaya, E.C.; Acuña, E. The Humboldt Current system of northern and central Chile. Oceanogr. Mar. Biol. 2007, 45, 195–344. [Google Scholar]
- Kahng, S.E.; Garcia-Sais, J.R.; Spalding, H.L.; Brokovich, E.; Wagner, D.; Weil, E.; Hinderstein, L.; Toonen, R.J. Community ecology of mesophotic coral reef ecosystems. Coral Reefs 2010, 29, 255–275. [Google Scholar] [CrossRef]
- Glynn, P.W. Coral reef bleaching: Facts, hypotheses and implications. Glob. Chang. Biol. 1996, 2, 495–509. [Google Scholar] [CrossRef]
- Asher, J.; Williams, I.D.; Harvey, E.S. Mesophotic depth gradients impact reef fish assemblage composition and functional group partitioning in the main Hawaiian Islands. Front. Mar. Sci. 2017, 4, 98. [Google Scholar] [CrossRef]
- Voerman, S.E. Ecosystem engineer morphological traits and taxon identity shape biodiversity across the euphotic-mesophotic transition. Proc. R. Soc. B. 2022, 289, 20211834. [Google Scholar] [CrossRef]
- Lesser, M.P.; Slattery, M.; Leichter, J.J. Ecology of mesophotic coral reefs. J. Exp. Mar. Biol. Ecol. 2009, 375, 1–8. [Google Scholar] [CrossRef]
- Bongaerts, P.; Ridgway, T.; Sampayo, E.M.; Hoegh-Guldberg, O. Assessing the “deep-reef refugia” hypothesis: Focus on Caribbean reefs. Coral Reefs 2010, 29, 309–327. [Google Scholar] [CrossRef]
- Loya, Y.; Eyal, G.; Treibitz, T.; Lesser, M.P.; Appeldoorn, R. Theme section on mesophotic coral ecosystems: Advances in knowledge and future perspectives. Coral Reefs 2016, 35, 1–9. [Google Scholar] [CrossRef]
- Sih, T.L.; Cappo, M.; Kingsford, M.J. Deep-reef fish assemblages of the Great Barrier Reef shelf-break (Australia). Sci. Rep. 2017, 7, 10886. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, H.T.; Goodbody-Grinley, G.; Jessup, M.E.; Shepherd, B.; Chequer, A.D.; Rocha, L.A. Upper and lower mesophotic coral reef fish communities evaluated by underwater visual censuses in two Caribbean locations. Coral Reefs 2016, 35, 139–151. [Google Scholar] [CrossRef]
- Bridge, T.C.L.; Hughes, T.P.; Guinotte, J.M.; Bongaerts, P. Call to protect all coral reefs. Nat. Clim. Chang. 2013, 3, 528–530. [Google Scholar] [CrossRef]
- Smith, T.B.; Glynn, P.W.; Maté, J.L.; Toth, L.T.; Gyory, J. A depth refugium from catastrophic coral bleaching prevents regional extinction. Ecology 2014, 95, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Matus, A.; Neubauer, P.; Shima, J.S.; Rivadeneira, M.M. Reef fish diversity across the Temperate South Pacific Ocean. Front. Ecol. Evol. 2022, 10, 768707. [Google Scholar] [CrossRef]
- Carbines, G.; Cole, R.C. Using a remote drift underwater video (DUV) to examine dredge impacts on demersal fishes and benthic habitat complexity in Foveaux Strait, Southern New Zealand. Fish. Res. 2009, 96, 230–237. [Google Scholar] [CrossRef]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef]
- Foster, M.S.; Harrold, C.; Hardin, D.D. Point vs. photo quadrat estimates of the cover of sessile marine organisms. J. Exp. Mar. Biol. Ecol. 1991, 146, 193–203. [Google Scholar] [CrossRef]
- Kohler, K.E.; Gill, S.M. Coral Point Count with Excel extensions (CPCe): A visual basic program for the determination of coral and substrate coverage using random point count methodology. Comput. Geosci. 2006, 32, 1259–1269. [Google Scholar] [CrossRef]
- Gotelli, N.J.; Colwell, R.K. Estimating Species Richness. In Biological Diversity: Frontiers in Measurement and Assessment; Magurran, A.E., McGill, B.J., Eds.; Oxford University Press: New York, NY, USA, 2011; Chapter 4; pp. 39–54. [Google Scholar]
- Chao, A.; Shen-Ming, L. Estimating the number of classes via sample coverage. JASA 1992, 87, 210–217. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar]
- Kruskal, J.B.; Wish, M. Multidimensional Scaling; Sage Publications: Thousand Oaks, CA, USA, 1978; p. 93. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. 2019. Available online: https://www.r-project.org (accessed on 25 January 2023).
- Delignette-Muller, M.L.; Dutang, C. Fitdistrplus: An R package for fitting distributions. J. Stat. Softw. 2015, 64, 1–34. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package ‘Corrplot’: Visualization of a Correlation Matrix (Version 0.92). 2021. Available online: https://github.com/taiyun/corrplot (accessed on 25 January 2023).
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting linear mixed-effects models using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Barton, K. MuMIn: Multi-Model Inference. R package Version 1.46.0. 2022. Available online: https://CRAN.R-project.org/package=MuMIn (accessed on 25 January 2023).
- Korner-Nievergelt, F.; Roth, T.; von Felten, S.; Guelat, J.; Almasi, B.; Korner-Nievergelt, P. Bayesian Data Analysis in Ecology Using Linear Models with R, BUGS and Stan; Elsevier: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Halekoh, U.; Højsgaard, S. A Kenward-Roger approximation and parametric bootstrap methods for tests in linear mixed models—The R package pbkrtest. J. Stat. Softw. 2014, 59, 1–30. [Google Scholar] [CrossRef]
- Haritg, F. DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. R Package Version 0.4.6. 2022. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 25 January 2023).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016. [Google Scholar]
- Lüdecke, D. Ggeffects: Tidy Data Frames of Marginal Effects from Regression Models. J. Open Source Softw. 2018, 3, 772. [Google Scholar] [CrossRef]
- Vavrek, M.J. Fossil: Palaeoecological and palaeogeographical analysis tools (version 0.4.0). Palaeontol. Electron. 2011, 14, 1. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; O’hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.; Wagner, H. Vegan: Community Ecology Package. R Package Version 2.6-2. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 25 January 2023).
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-81-2. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 25 January 2023).
- Camps-Castellà, J.; Breedy, O.; Vera-Escalona, I.; Vargas, S.; Silva, F.; Hinojosa, I.A.; Prado, P.; Brante, A. Distribution and divergence of shallow and upper mesophotic cold-water gorgonians (Octocorallia: Gorgoniidae) in Chilean coasts, with the description of a new species. In Proceedings of the VIII International Symposium on Marine Sciences, Las Palmas de Gran Canaria, Spain, 6–8 July 2022; p. 198. [Google Scholar]
- Montecino, V.; Pizarro, G. Phytoplankton acclimation and spectral penetration of UV irradiance off the central Chilean coast. Mar. Ecol. Prog. Ser. 1995, 121, 261–269. [Google Scholar] [CrossRef]
- Pyle, R.L.; Kosaki, R.K.; Pinheiro, H.T.; Rocha, L.A.; Whitton, R.K.; Copus, J.M. Fishes: Biodiversity. In Mesophotic Coral Ecosystems; Loya, Y., Puglise, K.A., Bridge, T.C.L., Eds.; Springer: New York, NY, USA, 2019; pp. 749–777. [Google Scholar]
- Thresher, R.E.; Colin, P.L. Trophic structure, diversity and abundance of fishes of the deep reef (30–300 m) at Enewetak, Marshall Islands. Bull. Mar. Sci. 1986, 38, 253–272. [Google Scholar]
- Connell, S.D.; Jones, G.P. The influence of habitat complexity on postrecruitment processes in a temperate reef fish population. J. Exp. Mar. Biol. Ecol. 1991, 151, 271–294. [Google Scholar] [CrossRef]
- Friedlander, A.M.; Brown, E.K.; Jokiel, P.L.; Smith, W.R.; Rodgers, K.S. Effects of habitat, wave exposure, and marine protected area status on coral reef fish assemblages in the Hawaiian archipelago. Coral Reefs 2003, 22, 291–305. [Google Scholar] [CrossRef]
- Bejarano, I.; Appeldoorn, R.S.; Nemeth, M. Fishes associated with mesophotic coral ecosystems in La Parguera, Puerto Rico. Coral Reefs 2014, 33, 313–328. [Google Scholar] [CrossRef]
- Stefanoudis, P.V.; Rivers, M.; Smith, S.R.; Schneider, C.W.; Wagner, D.; Ford, H.; Rogers, A.D.; Woodall, L.C. Low connectivity between shallow, mesophotic and rariphotic zone benthos. R. Soc. Open Sci. 2019, 6, 190958. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Matus, A.; Ferry-Graham, L.; Cea, A.; Vásquez, J. Community structure of temperate reef fishes in kelp-dominated subtidal habitats of northern Chile. Mar. Fresh. Res. 2007, 58, 1069–1085. [Google Scholar] [CrossRef]
- Navarrete-Fernández, T.; Landaeta, M.F.; Bustos, C.A.; Pérez-Matus, A. Nest building and description of parental care behavior in a temperate reef fish, Chromis crusma (Pisces: Pomacentridae). Rev. Chil. Hist. Nat. 2014, 87, 30. [Google Scholar] [CrossRef]
- Lee, M.R.; Hermosilla, C.; Castilla, J.C.; Fernández, M.; Clarke, M.; González, C.; Prado, L.; Rozbaczylo, N.; Valdovinos, C. Free-living benthic marine invertebrates in Chile. Rev. Chil. Hist. Nat. 2008, 81, 51–67. [Google Scholar] [CrossRef]
- Coni, E.O.C.; Ferreira, C.M.; de Moura, R.L.; Meirelles, P.M.; Kaufman, L.; Francini-Filho, R.B. An evaluation of the use of branching fire-corals (Millepora spp.) as refuge by reef fish in the Abrolhos Bank, eastern Brazil. Environ. Biol. Fishes 2013, 96, 45–55. [Google Scholar] [CrossRef]
- Ponti, M.; Turicchia, E.; Ferro, F.; Cerrano, C.; Abbiati, M. The understorey of gorgonian forests in mesophotic temperate reefs. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 1153–1166. [Google Scholar] [CrossRef]
- Rodríguez, S.R. Consumption of drift kelp by intertidal populations of the sea urchin Tetrapygus niger on the central Chilean coast: Possible consequences at different ecological levels. Mar. Ecol. Prog. Ser. 2003, 251, 141–151. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Area | Transects | Reef Fish Species | Benthic Community | ||
|---|---|---|---|---|---|---|
| N | Depth | N | Depth | |||
| Los Vilos | 8557.20 | 6 | 48 | 7.0–88.9 | 84 | 6.4–91 |
| Algarrobo | 5229.28 | 3 | 23 | 21.8–68.1 | 50 | 18–50.5 |
| El Quisco | 2631.07 | 7 | 22 | 8.4–63.9 | 48 | 8–50.7 |
| Las Cruces | 6927.13 | 2 | 27 | 11.9–60.9 | 51 | 12.3–55.8 |
| Total | 23,344.68 | 18 | 120 | 7.0–88.9 | 233 | 6.4–91 |
| Fish Abundance Negative Binomial Distribution (thetha = 1.23) | ||||||
| Fixed effects | Estimate | Std. Error | 2.5% | 97.5% | z-value | p-value |
| (Intercept) Depth Topographic complexity | −3.30 −0.57 0.97 | 0.22 0.15 0.13 | −6.39 −0.05 0.69 | −4.21 −0.01 1.19 | −14.73 −3.73 7.45 | <0.001 <0.001 <0.001 |
| Random effects | Variance | Std. Dev. | ||||
| Transect: site Site | 0.22 0.09 | 0.47 0.31 | ||||
| R2m 0.519, R2c 0.640 Overdispersion 1.02 | ||||||
| Fish richness Poisson Distribution | ||||||
| Fixed effects | Estimate | Std. Error | 2.5% | 97.5% | z-value | p-value |
| (Intercept) Sampled area Depth Topographic complexity | 0.64 0.03 −0.35 0.36 | 0.08 0.08 0.09 0.09 | 0.47 −0.12 −0.52 0.21 | 0.83 0.19 −0.17 0.53 | 8.07 0.43 −3.98 4.19 | <0.001 0.668 <0.001 <0.001 |
| Random effects | Variance | Std. Dev. | ||||
| Transect: site Site | 0.003 0.001 | 0.054 0.031 | ||||
| R2m 0.356, R2c 0.362 Overdispersion 0.85 | ||||||
| Df | Sum Sq | R2 | F | p | |
|---|---|---|---|---|---|
| Depth | 1 | 4.02 | 0.13 | 18.37 | 0.001 |
| Site | 3 | 1.87 | 0.06 | 2.84 | 0.003 |
| Topographic complexity | 1 | 2.71 | 0.09 | 12.38 | 0.001 |
| depth:site | 3 | 1.51 | 0.05 | 2.30 | 0.010 |
| Topographic complexity:site | 3 | 0.62 | 0.02 | 0.94 | 0.531 |
| Residuals | 91 | 19.90 | 0.65 | ||
| Total | 102 | 30.61 | 1.00 |
| Df | Sum Sq | R2 | F | p | |
|---|---|---|---|---|---|
| Depth | 1 | 1.73 | 0.05 | 14.88 | 0.001 |
| Site | 3 | 2.85 | 0.08 | 8.16 | 0.001 |
| depth:site | 3 | 2.85 | 0.08 | 8.17 | 0.001 |
| Residuals | 229 | 26.65 | 0.78 | ||
| Total | 236 | 34.08 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Campoy, A.N.; Pérez-Matus, A.; Wieters, E.A.; Alarcón-Ireland, R.; Garmendia, V.; Beldade, R.; Navarrete, S.A.; Fernández, M. The Hidden Diversity of Temperate Mesophotic Ecosystems from Central Chile (Southeastern Pacific Ocean) Assessed through Towed Underwater Videos. Diversity 2023, 15, 360. https://doi.org/10.3390/d15030360
Campoy AN, Pérez-Matus A, Wieters EA, Alarcón-Ireland R, Garmendia V, Beldade R, Navarrete SA, Fernández M. The Hidden Diversity of Temperate Mesophotic Ecosystems from Central Chile (Southeastern Pacific Ocean) Assessed through Towed Underwater Videos. Diversity. 2023; 15(3):360. https://doi.org/10.3390/d15030360
Chicago/Turabian StyleCampoy, Ana Navarro, Alejandro Pérez-Matus, Evie A. Wieters, Rodrigo Alarcón-Ireland, Vladimir Garmendia, Ricardo Beldade, Sergio A. Navarrete, and Miriam Fernández. 2023. "The Hidden Diversity of Temperate Mesophotic Ecosystems from Central Chile (Southeastern Pacific Ocean) Assessed through Towed Underwater Videos" Diversity 15, no. 3: 360. https://doi.org/10.3390/d15030360
APA StyleCampoy, A. N., Pérez-Matus, A., Wieters, E. A., Alarcón-Ireland, R., Garmendia, V., Beldade, R., Navarrete, S. A., & Fernández, M. (2023). The Hidden Diversity of Temperate Mesophotic Ecosystems from Central Chile (Southeastern Pacific Ocean) Assessed through Towed Underwater Videos. Diversity, 15(3), 360. https://doi.org/10.3390/d15030360