



How Epiphytic Are Filmy Ferns? A Semi-Quantitative Approach



Abstract
1. Introduction
2. Materials and Methods
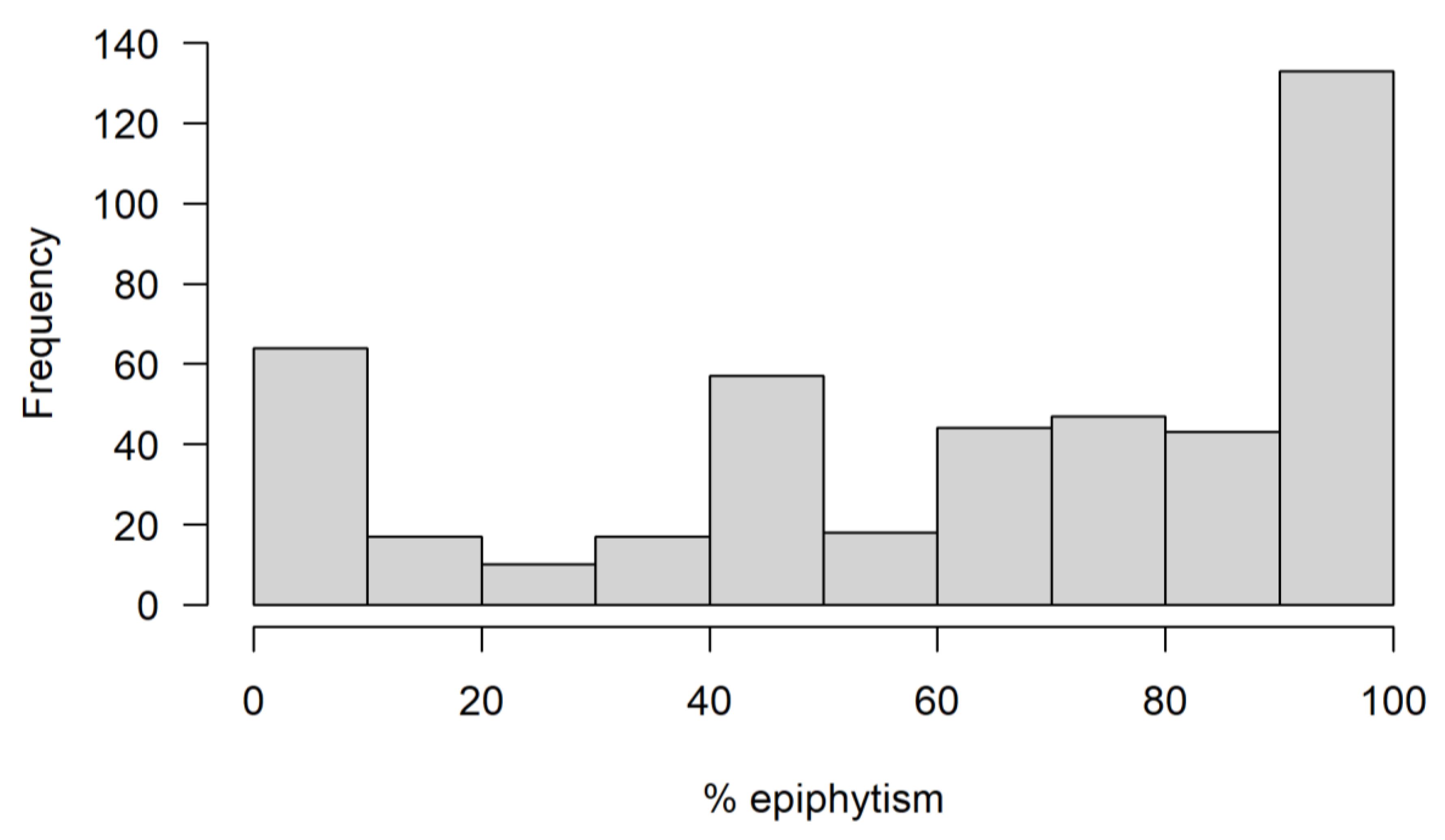
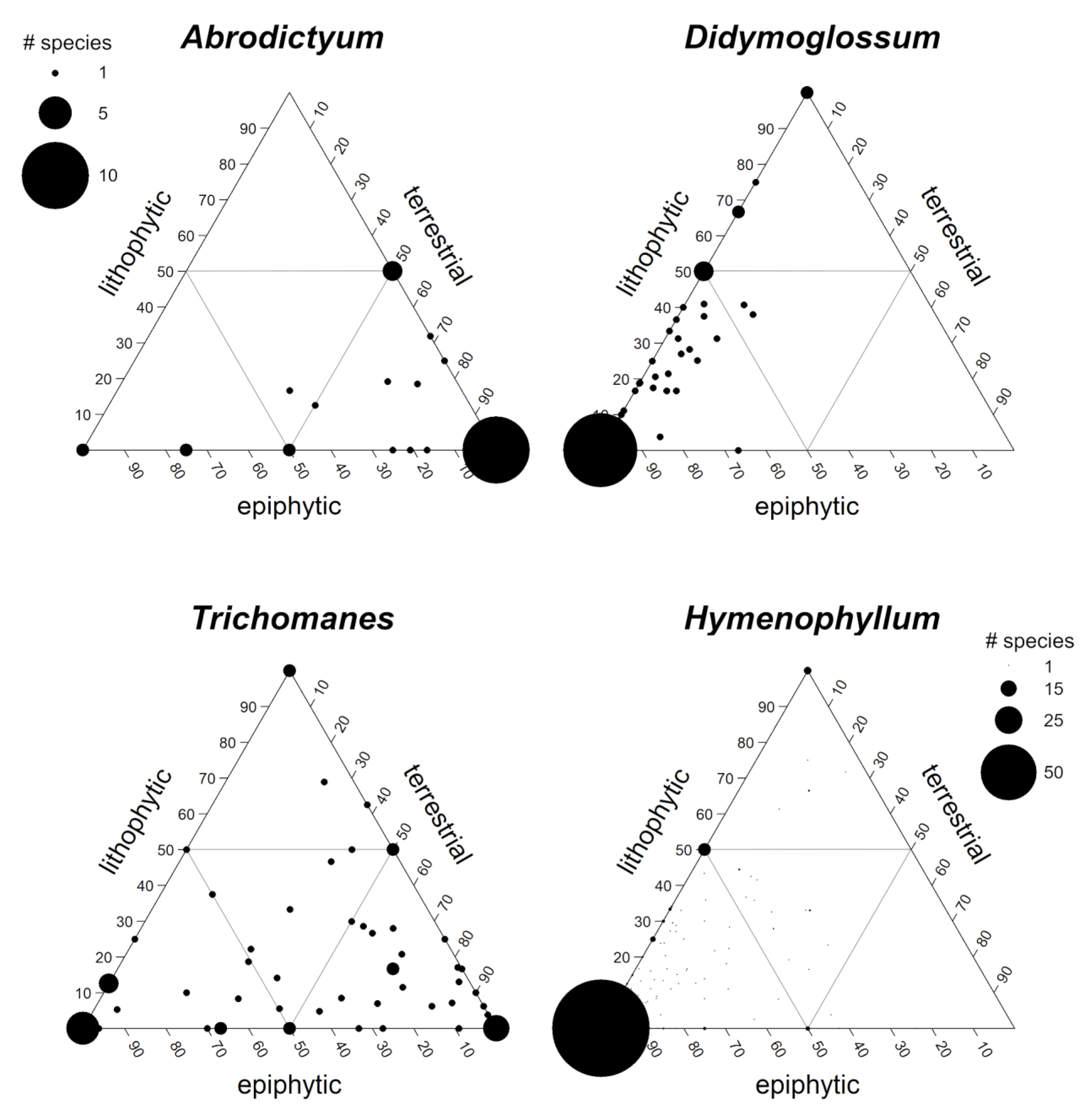
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zotz, G. Plants on Plants. The Biology of Vascular Epiphytes; Springer International Publishing Switzerland: Cham, Switzerland, 2016; p. 282. [Google Scholar]
- Schimper, A.F.W. Die Epiphytische Vegetation Amerikas; Gustav Fischer: Jena, Germany, 1888; Volume 2, p. 162. [Google Scholar]
- Benzing, D.H. The evolution of epiphytism. In Vascular Plants as Epiphytes: Evolution and Ecophysiology, 76th ed.; Lüttge, U., Ed.; Ecological Studies; Springer: Heidelberg, Germany, 1989; pp. 15–41. [Google Scholar]
- Ibisch, P.L. Neotropische Epiphytendiversität-das Beispiel Bolivien; Martina Galunder-Verlag: Wiehl, Germany, 1996; Volume 1, p. 357. [Google Scholar]
- Burns, K.C. How arboreal are epiphytes? A null model for Benzing’s classifications. N. Z. J. Bot. 2010, 48, 185–191. [Google Scholar] [CrossRef]
- Hoeber, V.; Zotz, G. Accidental epiphytes: Ecological insights and evolutionary implications. Ecol. Monogr. 2022, 92, e1527. [Google Scholar] [CrossRef]
- de Laubenfels, D.J.; Silba, J. Notes on Asian-Pacific Podocarpaceae I (Podocarpus). Phytologia 1988, 64, 290–292. [Google Scholar]
- de Laubenfels, D.J. New sections and species of Podocarpus based on the taxonomic status of P. neriifolius (Podocarpaceae) in tropical Asia. Novon A J. Bot. Nomencl. 2015, 24, 133–152. [Google Scholar] [CrossRef]
- Zotz, G. Gefässepiphyten in temperaten Wäldern. Bauhinia 2002, 16, 13–22. [Google Scholar]
- Johnson, D.S. Polypodium vulgare as an epiphyte. Bot. Gaz. 1921, 72, 237–244. [Google Scholar] [CrossRef]
- Klinghardt, M.; Zotz, G. Abundance and seasonal growth of epiphytic ferns at three sites along a rainfall gradient in Western Europe. Flora 2021, 274, 151749. [Google Scholar] [CrossRef]
- Barberis, I.M.; Mogni, V.Y.; Oakley, L.J.; Vogt, C.; Prado, D.E. Biogeography of different life forms of the southernmost neotropical tank bromeliad. J. Biogeogr. 2021, 48, 2085–2097. [Google Scholar] [CrossRef]
- Grigore, M.-N.; Ivanescu, L.; Toma, C. Halophytes: An. Integrative Anatomical Study; Springer International Publishing Switzerland: Cham, Switzerland, 2014. [Google Scholar]
- Winter, K.; Smith, J.A.C. CAM photosynthesis: The acid test. New Phytol. 2022, 233, 599–609. [Google Scholar] [CrossRef]
- Ogburn, R.M.; Edwards, E.J. The ecological water-use strategies of succulent plants. Adv. Bot. Res. 2010, 55, 179–225. [Google Scholar]
- Zotz, G.; Weigelt, P.; Kessler, M.; Kreft, H.; Taylor, A. EpiList 1.0-A global checklist of vascular epiphytes. Ecology 2021, 102, e03326. [Google Scholar] [CrossRef]
- Wester, S.; Zotz, G. Growth and survival of Tillandsia flexuosa on electricity cables in Panama. J. Trop. Ecol. 2010, 26, 123–126. [Google Scholar] [CrossRef]
- Correa, M.D.; Galdames, C.; de Stapf, M.S. Catálogo de las Plantas Vasculares de Panamá; Universidad de Panamá & Instituto Smithsonian de Investigaciones Tropicales: Panama City, Panama, 2004. [Google Scholar]
- Ebihara, A.; Dubuisson, J.-Y.; Iwatsuk, K.; Hennequin, S.; Ito, M. A taxonomic revision of Hymenophyllaceae. Blumea 2006, 51, 221–280. [Google Scholar] [CrossRef]
- Dubuisson, J.Y.; Hennequin, S.; Rakotondrainibe, F.; Schneider, H. Ecological diversity and adaptive tendencies in the tropical fern Trichomanes L. (Hymenophyllaceae) with special reference to climbing and epiphytic habits. Bot. J. Linn. Soc. 2003, 142, 41–63. [Google Scholar] [CrossRef]
- Lehnert, M.; Krug, M. Evolution of substrate specificity and fungal symbiosis in filmy ferns (Hymenophyllaceae): A Bayesian approach for ambiguous character state reconstruction. Symbiosis 2019, 78, 141–147. [Google Scholar] [CrossRef]
- Brazilian Flora 2020 in Construction. Available online: http://floradobrasil.jbrj.gov.br/ (accessed on 15 May 2022).
- Freitas, L.; Salino, A.; Menini Neto, L.; Almeida, T.; Mortara, S.; Stehmann, J.; Amorim, A.M.; Guimaraes, E.; Nadruz Coelho, M.; Zanin, A.; et al. A comprehensive checklist of vascular epiphytes of the Atlantic Forest reveals outstanding endemic rates. PhytoKeys 2016, 58, 65–79. [Google Scholar] [CrossRef]
- Carvajal-Hernández, C.I.; Silva-Mijangos, L.; Kessler, M.; Lehnert, M. Additions to the pteridoflora of Tabasco, Mexico: The importance of the humid montane forest. Acta Bot. Mex. 2018, 124, 7–18. [Google Scholar] [CrossRef]
- Hennequin, S.; Schuettpelz, E.; Pryer, K.M.; Ebihara, A.; Dubuisson, J.Y. Divergence times and the evolution of epiphytism in filmy ferns (Hymenophyllaceae) revisited. Int. J. Plant Sci. 2008, 169, 1278–1287. [Google Scholar] [CrossRef]
- Werner, F.; Homeier, J.; Gradstein, S.R. Diversity of vascular epiphytes on isolated remnant trees in the montane forest belt of southern Ecuador. Ecotropica 2005, 11, 21–40. [Google Scholar]
- Hassler, M. World Ferns. Synonymic Checklist and Distribution of Ferns and Lycophytes of the World. Version 14.4; Last Update December 4th, 2022. Available online: www.worldplants.de/ferns/ (accessed on 12 December 2022).
- Bidin, A. A preliminary survey of the fern flora of Langkawi Islands. Gard. ´S Bull. Singap. 1987, 40, 77–102. [Google Scholar]
- Lellinger, D.B. Hymenophyllaceae. In Flora of the Guianas; Rijn, A.R.A.G.-v., Ed.; Fasc. 3; Koeltz Scientific Books: Königstein, Germany, 1994; Volume Ser. B, pp. 1–66. [Google Scholar]
- Khanina, L.; Bobrovsky, M. Value of large Quercus robur fallen logs in enhancing the species diversity of vascular plants in an old-growth mesic broad-leaved forest in the Central Russian Upland. For. Ecol. Manag. 2021, 491, 119172. [Google Scholar] [CrossRef]
- Ough, K. Regeneration of wet forest flora a decade after clear-felling or wildfire-is there a difference? Austr. J. Bot. 2001, 49, 645–664. [Google Scholar] [CrossRef]
- Franco, M.; Silvertown, J. Life history variation in plants: An exploration of the fast-slow continuum hypothesis. Phil. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1341–1348. [Google Scholar]
- Lemon, J. Plotrix: A package in the red light district of R. R-News 2006, 6, 8–12. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing Vienna: Vienna, Austria, 2013; Available online: http://www.R-project.org (accessed on 12 December 2022).
- Kress, W.J. The systematic occurrence of vascular epiphytes. In Vascular Plants as Epiphytes: Evolution and Ecophysiology, 76th ed.; Lüttge, U., Ed.; Ecological Studies; Springer: Heidelberg, Germany, 1989; pp. 234–261. [Google Scholar]
- Hokche, O.; Berry, P.E.; Huber, O. (Eds.) Nuevo Catálogo de la Flora Vascular de Venezuela; Fundación Instituto Botánico de Venezuela: Caracas, Venezuela, 2008; p. 859. [Google Scholar]
- Steyermark, J.A.; Berry, P.E.; Holst, B.K.; Yatskievych, K. (Eds.) Flora of the Venezuelan Guayana: Pteridophytes, Spermatophytes Acanthaceae-Araceae; Timber Press: Portland, OR, USA, 1995. [Google Scholar]
- Winter, K. Ecophysiology of constitutive and facultative CAM photosynthesis. J. Exp. Bot. 2019, 70, 6495–6508. [Google Scholar] [CrossRef]
- Flora of the Marquesas Islands. Available online: https://naturalhistory2.si.edu/botany/marquesasflora/index.htm. (accessed on 12 December 2022).
- Mellado-Mansilla, D.; Diaz, I.A.; Godoy-Guinao, J.; Ortega-Solis, G.; Moreno-Gonzalez, R. Bosque Pehuen Park’s Flora: A contribution to the knowledge of the Andean montane forests in the Araucania region, Chile. Nat. Areas J. 2018, 38, 298–311. [Google Scholar] [CrossRef]
- Moreno, R.; le Quesne, C.; Díaz, I.; Rodríguez, R. Flora vascular del Parque Futangue, Region de Los Rios (Chile). Gayana Bot. 2013, 70, 121–135. [Google Scholar] [CrossRef]
- Chen, C.-C.; Hyvönen, J.; Schneider, H. Re-terrestrialization in the phylogeny of epiphytic plant lineages: Microsoroid ferns as a case study. J. Syst. Evol. 2022. [Google Scholar] [CrossRef]
- Kessler, M.; Smith, A.R. Prodromus of a fern flora for Bolivia. X. Hymenophyllaceae. Phytotaxa 2017, 328, 201–226. [Google Scholar] [CrossRef]
- Espejo-Serna, A.; López-Ferrari, A.R.; Mendoza-Ruiz, A.; García-Cruz, J.; Ceja-Romero, J.; Pérez-García, B. Mexican vascular epiphytes: Richness and distribution. Phytotaxa 2021, 503, 1–124. [Google Scholar] [CrossRef]
- Acebey, A.R.; Krömer, T.; Kessler, M. Species richness and vertical distribution of ferns and lycophytes along an elevational gradient in Los Tuxtlas, Veracruz, Mexico. Flora 2017, 235, 83–91. [Google Scholar] [CrossRef]
- Bøgh, A. Composition and distribution of the vascular epiphyte flora of an Ecuadorian montane rain forest. Selbyana 1992, 13, 25–34. [Google Scholar]
- Zotz, G.; Almeda, F.; Bautista-Bello, A.P.; Eskov, A.K.; Giraldo-Cañas, D.; Hammel, B.E.; Harrison, R.D.; Köster, N.; Krömer, T.; Lowry, P.P., II; et al. Hemiepiphytes revisited. Perspect. Plant Ecol. Evol. Syst. 2021, 51, 125620. [Google Scholar] [CrossRef]
- Nitta, J.H.; Epps, M.J. Hemi-epiphytism in Vandenboschia collariata (Hymenophyllaceae). Brittonia 2009, 61, 392–397. [Google Scholar] [CrossRef]
- Watts, J.L.; Moran, R.C.; Watkins, J.E., Jr. Hymenasplenium volubile: Documentation of its gametophytes and the first record of a hemiepiphyte in the Aspleniaceae. Ann. Bot. 2019, 124, 829–835. [Google Scholar] [CrossRef]
- Watkins, J., Jr. E.; Moran, R.C. Gametophytes of the fern genera Dracoglossum and Lomariopsis (Lomariopsidaceae) and their phylogenetic significance. Int. J. Pl. Sci. 2019, 180, 1004–1015. [Google Scholar] [CrossRef]
- Testo, W.; Sundue, M. Primary hemiepiphytism in Colysis ampla (Polypodiaceae) provides new insight into the evolution of growth habit in ferns. Int. J. Plant Sci. 2014, 175, 526–536. [Google Scholar] [CrossRef]
- Zizka, A.; Azevedo, J.; Leme, E.; Neves, B.; da Costa, A.F.; Caceres, D.; Zizka, G. Biogeography and conservation status of the pineapple family (Bromeliaceae). Divers. Distrib. 2020, 26, 183–195. [Google Scholar] [CrossRef]
- Khine, P.K.; Fraser-Jenkins, C.; Lindsay, S.; Middleton, D.; Miehe, G.; Thomas, P.; Kluge, J. A contribution toward the knowledge of ferns and lycophytes from Northern and Northwestern Myanmar. Am. Fern J. 2017, 107, 219–256. [Google Scholar] [CrossRef]
- Krömer, T.; Kessler, M. Filmy ferns (Hymenophyllaceae) as high-canopy epiphytes. Ecotropica 2006, 12, 57–63. [Google Scholar]
- Zotz, G.; Büche, M. The epiphytic filmy ferns of a tropical lowland forest-species occurrence and habitat preferences. Ecotropica 2000, 6, 203–206. [Google Scholar]
- Roux, J.P. Synopsis of the Lycopodiophyta and Pteridophyta of Africa, Madagascar and neighbouring islands. Strelitzia 2009, 23, 227–255. [Google Scholar]
- Brownlie, G. The pteridophyte flora of Fiji. Nova Hedwig Beih. 1977, 55, 1–397. [Google Scholar]
- Iwatsuki, K. Taxonomic studies of pteridophyta II. Acta Phytotax. Geobot. 1958, 17, 161–166. [Google Scholar]
- Game, J.C.; Fawcett, S.E.; Smith, A.R. New pteridophyte records for Taveuni (Fiji) and a new species of Chingia (Thelypteridaceae). N. Z. J. Bot. 2018, 56, 26–37. [Google Scholar] [CrossRef]
- Nitta, J.H. Distribution, Ecology, and Systematics of the Filmy Ferns (Hymenophyllaceae) of Moorea, French Polynesia. Water Resources Center Archives. Biology and Geomorphology of Tropical Islands. 2006. Available online: https://escholarship.org/uc/item/6vt6p2w8 (accessed on 12 December 2022).
- Liu, J.X.; Zhang, Q.Y.; Ebihara, A.; Iwatsuki, K. Hymenophyllaceae. In Flora of China, Vol. 2–3 (Pteridophytes); Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MO, USA, 2013; pp. 93–109. [Google Scholar]
- Cámara-Leret, R.; Frodin, D.G.; Adema, F.; Anderson, C.; Appelhans, M.S.; Argent, G.; Arias Guerrero, S.; Ashton, P.; Baker, W.J.; Barfod, A.S.; et al. New Guinea has the world’s richest island flora. Nature 2020, 584, 579–583. [Google Scholar] [CrossRef]
- Christensen, C. Botanical results of the Archbold Expedition No. 8: New and noteworthy Papuan ferns. Brittonia 1937, 2, 265–317. [Google Scholar] [CrossRef]
- Murdock, A.G.; Smith, A.R. Pteridophytes of Moorea, French Polynesia, with a new species, Tmesipteris gracilis (Psilotaceae). Pacific Sci. 2003, 57, 253–265. [Google Scholar] [CrossRef]
- Pichi Sermolli, R.E.G. A contribution to the knowledge of the Pteridophyta of Rwanda, Burundi, and Kivu (Zaire). I. Bull. Jard. Bot. Natl. Belg. Bull. Van Natl. Plantentuin Van Belg. 1983, 53, 177–284. [Google Scholar] [CrossRef]
- Roux, J.P. Conspectus of Southern African Pteridophyta: An Enumeration of the Pteridophyta of Angola, Botswana, Lesotho, Malawi, Mozambique, Namibia, South. Africa (Including the Marion Island Group), Swaziland, Zambia and Zimbabwe; Project Coordinator, Southern African Botanical Diversity Network; c/o National Botanical Institute: Pretoria, South Africa, 2001. [Google Scholar]
- Roux, J.P.; Network, S.A.B.D. Swaziland Ferns and Fern Allies; National Botanical Institute, SABONET: Pretoria, Pretoria, 2003; Volume 19. [Google Scholar]
- Clarkson, B.D. The vegetation of the Kaitake Range Egmont National Park, New Zealand. N. Z. J. Bot. 1985, 23, 15–31. [Google Scholar] [CrossRef]
- Ebihara, A.; Matsumoto, S.; Iwashina, T.; Sugimura, K.; Iwatsuki, K. Hymenophyllaceae (Pteridophytes) of Vanuatu. Ann. Tsukuba Bot. Gard. 2002, 21, 61–72. [Google Scholar]
- Dubuisson, J.Y.; Bauret, L.; Grall, A.; Li, T.; Ebihara, A.; Hennequin, S. Discussion on the taxonomy of African fern Abrodictyum rigidum (Sw.) Ebihara & Dubuisson and description of two new Abrodictyum C.Presl species (Hymenophyllaceae, Polypodiidae) for the Afro-Malagasy region. Phytotaxa 2016, 284, 151–168. [Google Scholar] [CrossRef]
- Benl, G. The pteridophyta of Bioko (Fernando Po):(Contributions to a Flora of the island): II: Marattiaceae, Hymenophyllaceae, Adiantaceae, Hemionitidaceae. Acta Bot. Barcinonensia 1980, 32, 3–34. [Google Scholar]
- Harley, W.J. The ferns of Liberia. Contrib. Gray Herb. Harv. Univ. 1955, 177, 58–101. [Google Scholar] [CrossRef]
- Velayos, M.; Aedo, C.; Cabezas, F.; Estrella, M. XIII-Hymenophyllaceae. In Flora de Guinea Ecuatorial, I; Velayos, M., Aedo, C., Cabezas, F., Estrella, M., Eds.; 2008; p. 28. Available online: https://digital.csic.es/bitstream/10261/117484/3/013-HYMENOPHYLLACEAE.pdf (accessed on 12 December 2022).
- Iwatsuki, K.; Ebihara, A.; Kato, M. Taxonomic studies of pteridophytes of Ambon and Seram (Moluccas) collected on Indonesian-Japanese botanical expeditions 1983-1986. XIII. Hymenophyllaceae. PhytoKeys 2019, 119, 107–115. [Google Scholar] [CrossRef]
- Turner, I.M. A Catalogue of the vascular plants of Malaya. Gard. Bull. Singap. 1995, 47, 1–757. Available online: https://books.google.com.au/books/about/A_Catalogue_of_the_Vascular_Plants_of_Ma.html?id=4zXgswEACAAJ&redir_esc=y (accessed on 12 December 2022).
- Piggott, A.G. Ferns of Malaysia in Colour; Tropical Press: Kuala Lumpur, Malaysia, 1988. [Google Scholar]
- Becker, M. A preliminary survey of the fern flora of Penang Hill. Flora Males. Bull. 1994, 11, 293–300. [Google Scholar]
- Iwatsuki, K.; Price, M.G. The pteridophytes of Mt Burnay and vicinity, Northern Luzon. Southeast. Asian Stud. 1977, 14, 540–572. [Google Scholar]
- Lindsay, S.; Middleton, D.J. Ferns of Thailand, Laos and Cambodia. Available online: http://rbg-web2.rbge.org.uk/thaiferns/ (accessed on 12 December 2022).
- Popelka, O.; Dančák, M.; Sukri, R.S.; Metali, F. Novitates Bruneienses, 10. Filmy ferns (Hymenophyllaceae) of Kuala Belalong. Brunei Darussalam 2018, 70, 123–154. [Google Scholar]
- Sledge, W.A. The Hymenophyllaceae of Ceylon. J. Linn. Soc. Lond. Bot. 1968, 60, 289–308. [Google Scholar] [CrossRef]
- Saïd, A.H.; Hennequin, S.; Rouhan, G.; Dubuisson, J.Y. Disentangling the diversity and taxonomy of Hymenophyllaceae (Hymenophyllales, Polypodiidae) in the Comoros. Eur. J. Taxon. 2017, 313. [Google Scholar] [CrossRef]
- Beentje, H.J. Hymenophyllaceae; Royal Botanic Gardens: Kew, Thailand, 2008. [Google Scholar]
- Blum, C.T.; Roderjan, C.V.; Galvao, F. Composição florística e distribuição altitudinal de epífitas vasculares da Floresta Ombrófila Densa na Serra da Prata, Morretes, Paraná, Brasil. Biota Neotrop. 2011, 11, 141–159. [Google Scholar] [CrossRef]
- Cetzal-Ix, W.; Noguera-Savelli, E.; Ramirez-Marcial, N. New records of ferns for Tabasco, Mexico. Rev. Mex. Biodiv. 2013, 84, 977–982. [Google Scholar] [CrossRef]
- Christenhusz, M.J.M. Index Pteridophytorum Guadalupensium or a revised checklist to the ferns and club mosses of Guadeloupe (French West Indies). Bot. J. Linn. Soc. 2009, 161, 213–277. [Google Scholar] [CrossRef]
- Dittrich, V.A.d.O.; Waechter, J.L.; Salino, A. Species richness of pteridophytes in a montane Atlantic rain forest plot of Southern Brazil. Acta Bot. Bras. 2005, 19, 519–525. [Google Scholar] [CrossRef]
- Ferreira de Lima, R.A.; de Oliveira Dittrich, V.A.; de Souza, V.C.; Salino, A.; Breier, T.B.; de Aguiar, O.T. Flora vascular do Parque Estadual Carlos Botelho, São Paulo, Brasil. Biota Neotrop. 2011, 11, 173–214. [Google Scholar] [CrossRef]
- Gasper, A.L.d.; Sevegnani, L. Lycophyta e samambaias do Parque Nacional da Serra do Itajaí, Vale do Itajaí, SC, Brasil. Hoehnea 2010, 37, 755–767. [Google Scholar] [CrossRef]
- Grayum, M.H.; Churchill, H.W. The vascular flora of La Selva Biological Station, Costa Rica- Polypodiophyta. Selbyana 1989, 11, 66–118. [Google Scholar]
- Haber, W. Appendix 1-Vascular plants of Monteverde. In Monteverde: Ecology and Conservation of a Tropical Cloud Forest; Nadkarni, N.M., Whellwright, N., Eds.; Oxford University Press: New York, NY, USA, 2000; pp. 457–518. [Google Scholar]
- Hyde, M.A.; Wursten, B.T.; Ballings, P.; Coates Palgrave, M. Flora of Zimbabwe: Home page. 2019. Available online: https://www.zimbabweflora.co.zw/index.php (accessed on 12 December 2022).
- Kornás, J. Life-forms and seasonal patterns in the pteridophytes in Zambia. Acta Soc. Bot. Pol. 1977, 46, 669–690. [Google Scholar] [CrossRef]
- Lellinger, D.B. The Ferns and Fern-Allies of Costa Rica, Panama, and the Chocó (Part. I: Psilotaceae through Dicksoniaceae); American Fern Society: Washington, DC, USA, 1989; Volume 2a, p. 364. [Google Scholar]
- Mori, S.A.; Cremers, G.; Gracie, C.; de Granville, J.-J.; Hoff, M.; Mitchell, J.D. Pteridophytes, Gymnosperms, and Monocotyledons; New York Botanical Garden: New York, NY, USA, 1997; Volume 1, p. 422 S. [Google Scholar]
- Nitta, J.H.; Ebihara, A.; Smith, A.R. A taxonomic and molecular survey of the pteridophytes of the Nectandra Cloud Forest Reserve, Costa Rica. PLoS ONE 2020, 15. [Google Scholar] [CrossRef]
- Nóbrega, G.A.; Aidar, M.P.M.; Paciencia, M.; Prado, J. Identification key for lycophytes and ferns from the Picinguaba and Santa Virginia Nuclei, Parque Estadual da Serra do Mar, Ubatuba, SP, Brazil. Biota Neotrop. 2016, 16. [Google Scholar] [CrossRef]
- Shreve, F. Studies on Jamaican Hymenophyllaceae. Bot. Gaz. 1911, 51, 184–209. [Google Scholar] [CrossRef]
- Windisch, P.G. Pteridófitas do Estado de Mato Grosso: Hymenophyllaceae. Bradea 1996, 6, 400–423. [Google Scholar]
- Schippers, R.R. Pteridophytes of Tanzania with special reference to Pare and Usambara Mountains. Fern Gaz. 1993, 14, 171–192. [Google Scholar]
- Windisch, P.G. Hymenophyllaceae (Polypodiopsida) no estado do Rio Grande do Sul. Pesqui. Botânica 2014, 65, 15–48. [Google Scholar]
- Graham, B.M.; Rickard, M.H. A revised list of the pteridophytes of Nevis. Fern Gaz. 1992, 14, 85–95. [Google Scholar]
- Peña-Chocorro, M.C.; Jiménez, B.; Marín, G.; Knapp, S. Checklist of the pteridophytes of the Mbaracayú Forest Nature Reserve, Paraguay. Fern Gaz. 1999, 15, 221–258. [Google Scholar]
- Peña-Chocarro, M.; Espada-Mateos, C.; Vera, M.; Céspedes, G.; Knapp, S. Updated checklist of vascular plants of the Mbaracayú Forest Nature Reserve (Reserva Natural del Bosque Mbaracayú), Paraguay. Phytotaxa 2010, 12, 1–224. [Google Scholar] [CrossRef]
- Monro, A.K.; Santamaria-Aguilar, D.; Gonzalez, F.; Chacon, O.; Solano, D.; Rodriguez, A.; Zamora, N.; Fedele, E.; Correa, M. A first checklist to the vascular plants of La Amistad International Park (PILA), Costa Rica-Panama. Phytotaxa 2017, 322, 1–283. [Google Scholar] [CrossRef]
- Wilson, H.D. Field Guide Stewart Island Plants; Manuka Press: Christchurch, New Zealand, 1994. [Google Scholar]
- Lellinger, D.B. Notes on neotropical Hymenophyllaceae. Am. Fern J. 1991, 81, 24–36. [Google Scholar] [CrossRef]
- Mildawati; Sobir, S.; Sulistijorini, S.; Chikmawati, T. The diversity of pteridophytes in Siberut National Park, Mentawai Islands, West Sumatra, Indonesia. Biodiversitas 2020, 21, 3200–3208. [Google Scholar] [CrossRef]
- Chambers, T.C.; Jermy, A.C.; Crabbe, J.A. A collection of ferns from Espiritu Santo, New Hebrides. Fern Gaz. 1971, 10, 175. [Google Scholar]
- Flora of the Hawaiian Islands. Available online: https://naturalhistory2.si.edu/botany/hawaiianflora/index.htm (accessed on 12 November 2022).
- Barcelona, J.F. The taxonomy and ecology of the pteridophytes of Mt. Iraya and vicinity, Batan Island, Batanes Province, Northern Philippines. In Pteridology in the New Millenium; Springer: Dordrecht, The Netherlands, 2003; pp. 299–325. [Google Scholar]
- Cadet, T. La Végétation de l’Île de la Réunion. Etude Phytoécologique et Phytosociologique. Ph.D. Thesis, Université d’Aix-Marseille, St. Denis, La Rèunion, 1977. [Google Scholar]
- Dubuisson, J.Y.; Rouhan, G.; Grall, A.; Hennequin, S.; Senterre, B.; Pynee, K.; Ebihara, A. New insights into the systematics and evolution of the filmy fern genus Crepidomanes (Hymenophyllaceae) in the Mascarene Archipelago with a focus on dwarf species. Acta Bot. Gall. 2013, 160, 173–194. [Google Scholar] [CrossRef]
- Yuyen, Y.; Boonkerd, T. Pteridophyte flora of Huai Yang waterfall National Park, Prachuap Khiri Khan province, Thailand. Nat. Hist. J. Chulalongkorn Univ. 2002, 2, 39–49. [Google Scholar]
- Chapman, J.D.; Chapman, H.M. The forests of Taraba and Adamawa States, Nigeria. An. Ecological Account and Plant Species Checklist; University of Canterbury: Christchurch, New Zealand, 2001; pp. 146–175. [Google Scholar]
- Fischer, E.; Rembold, K.; Althof, A.; Obholzer, J.; Malombe, I.; Mwachala, G.; Onyango, J.C.; Dumbo, B.; Theisen, I. Annotated checklist of the vascular plants of Kakamega Forest, Western Province, Kenya. J. East. African Nat. Hist. 2010, 99, 129–226. [Google Scholar] [CrossRef]
- Flora of Australia. Australian Biological Resources Study, Canberra. Available online: http://www.ausflora.org.au (accessed on 12 December 2022).
- Game, J.C.; Pene, S.; Smith, A.R. A new specimen-based checklist of ferns and lycophytes from Rotuma (Fiji). N. Z. J. Bot. 2021, 59, 137–153. [Google Scholar] [CrossRef]
- Hameed, C.A.; Madhusoodanan, P.V. A new species of filmy fern (Hymenophyllaceae: Pteridophyta) from South India. SIDA Contr. Bot. 1998, 18, 519–522. [Google Scholar]
- Madhusoodanan, P.V.; Hameed, C.A. Crepidomanes agasthianum, a new filmy fern species (Hymenophyllaceae—Pteridophyta) from India. Nord. J. Bot. 1998, 18, 169–170. [Google Scholar] [CrossRef]
- Farrar, D.R. Trichomanes intricatum: The independent trichomanes gametophyte in the Eastern United States. Am. Fern J. 1992, 82, 68–74. [Google Scholar] [CrossRef]
- Trigoboff, N.; Li, F.W. Crepidomanes intricatum (Hymenophyllaceae), a sporophyte-less filmy fern new to Central New York. Am. Fern J. 2022, 112, 139–141. [Google Scholar]
- Patil, S.M.; Hande, P.R.; Dongare, M.M. A filmy fern: Crepidomanes latealatum (Bosch) Copel. new to Northern Western Ghats of India. Indian For. 2014, 140, 939–940. [Google Scholar]
- Chen, C.W.; Hsu, T.C.; Chao, Y.S.; Lu, P.F.; Li, C.W.; Tram, N.K.T.; Nguyen, I.; Dang, M.T.; Luu, H.T.; Parris, B. A newly recorded genus and twelve newly recorded species of ferns for Vietnam from Lang Biang Plateau. Phytotaxa 2020, 443, 121–143. [Google Scholar] [CrossRef]
- Grieve, G.R.H.; Downs, C.T. A checklist of the plants of the forests and grasslands in the Weza district, southern KwaZulu-Natal and a review of their status in the Red Data List. Koedoe 2015, 57, 1237. [Google Scholar] [CrossRef]
- Pynee, K.; Hennequin, S.; Echternacht, L.; Dubuisson, J.-Y. A new local variety of Crepidomanes minutum (Hymenophyllaceae) in the Mascarene Archipelago (Indian Ocean) and a new record for Mauritius. Phytotaxa 2012, 62, 25–30. [Google Scholar] [CrossRef]
- Faden, R.B. A new species of Trichomanes from Eastern Africa. Am. Fern J. 1977, 67, 5–10. [Google Scholar] [CrossRef]
- Price, M.G. Notes on Philippine ferns. Fern Gaz. 1972, 10, 253–262. [Google Scholar]
- Joseph, J.; Singh, S. Trichomanes saxifragoides Presl (Hymenophyllaceae)-A new record of fern for North Eastern India from Tirap District, Arunachal Pradesh. Bull. Bot. Surv. India 1981, 21, 213–214. [Google Scholar]
- Lee, C.S.; Lee, K.; Hwang, Y. First record of Crepidomanes schmidtianum (Zenker ex Tasch.) K. Iwats. (Hymenophyllaceae) from Korea. Korean J. Plant Tax. 2014, 44, 1–5. [Google Scholar] [CrossRef]
- Bir, S.S.; Irudayaraj, V.; Manickam, V.S. Cytology of some ferns from the Nilgiris, South India III. Fern Gaz. 1996, 15, 141–149. [Google Scholar]
- Da Silva, W.R.; Ferreira, A.W.C.; Ilkiu-Borges, A.L.; Fernandes, R.S. Ferns and lycophytes of remnants in Amazonia Maranhense, Brazil. Biota Neotrop. 2020, 20. Available online: https://www.scielo.br/j/bn/a/nDDPNQNktNz8VhpQyFGrLxx/?lang=en (accessed on 12 December 2022). [CrossRef]
- Matos, F.B.d. Samambaias e Licófitas da RPPN Serra Bonita, Município de Camacan, sul da Banhia, Brasil. 2009. Available online: https://www.researchgate.net/publication/26980966_Samambaias_e_licofitas_da_RPPN_Serra_Bonita_municipio_de_Camacan_sul_da_Bahia_Brasil (accessed on 12 December 2022).
- Wessels Boer, J.G. The new world species of Trichomanes sect. Didymoglossum and Microgonium. Acta Bot. Neerl. 1962, 11, 277–330. [Google Scholar] [CrossRef]
- Pichi Sermolli, R.E.G. Microgonium erosum and related species (Hymenophyllaceae) in continental Africa. Webbia 1982, 35, 241–260. [Google Scholar] [CrossRef]
- Senterre, B.; Rouhan, G.; Morel, C.; Dubuisson, J.Y. New species and records in the fern genus Didymoglossum for the flora of Seychelles, with notes on the Southeast Asian D. motleyi complex (Hymenophyllaceae). Phytotaxa 2017, 292, 201–217. [Google Scholar] [CrossRef]
- Croxall, J.P. Microgonium (Hymenophyllaceae) in Malesia, with special reference to Peninsular Malaysia. Kew Bull. 1986, 41, 519–531. [Google Scholar] [CrossRef]
- Palacios-Rios, M. Pteridophytes of the state of Veracruz, Mexico: New records. Fern Gaz. 1992, 14, 119–122. [Google Scholar] [CrossRef]
- Sánchez, C. A new filmy fern species and new unispecific section of Trichomanes (Hymenophyllaceae) ("filmy ferns") from Cuba. Willdenowia 2001, 31, 125–127. [Google Scholar] [CrossRef]
- Croat, T.B. Flora of Barro Colorado Island; Stanford University Press: Stanford, CA, USA, 1978; p. 943. [Google Scholar]
- Christensen, C. XIX.-On the ferns of the Seychelles and the Aldabra Group. Trans. Linn. Soc. Lond. -Second. Ser. Bot. 1912, VII, 409–424. [Google Scholar] [CrossRef]
- Senterre, B.; Gerlach, J.; Mougal, J.; Matatiken, D. Old growth mature forest types and their floristic composition along the altitudinal gradient on Silhouette Island (Seychelles)-the telescoping effect on a continental mid-oceanic island. Phytocoen 2009, 39, 157–174. [Google Scholar] [CrossRef]
- Cascante-Marín, A.; Estrada-Chavarría, A. Las plantas vasculares de El Rodeo, Costa Rica. Brenesia 2012, 77, 71–128. [Google Scholar]
- Hameed, C.A.; Madhusoodanan, P.V. Trichomanes vamana, a new filmy fern species (Hymenophyllaceae-Pteridophyta) from India. Nord. J. Bot. 2005, 23, 437–439. [Google Scholar] [CrossRef]
- Acebey, A.R.; Lopez-Acosta, J.C.; Tejero-Diez, J.D.; Krömer, T. Richness and composition of ferns and lycophytes in three areas of humid montane forest in Los Tuxtlas, Veracruz, Mexico. Rev. Mex. Biodiv. 2017, 88, 625–635. [Google Scholar] [CrossRef]
- Arana, M.D.; Larsen, C.; Ponce, M.M. Revisión y análisis panbiogeográfico de las Hymenophyllaceae de las Yungas meridionales de Argentina (Selva Tucumano-Boliviana). Rodriguésia 2016, 67, 55–76. [Google Scholar] [CrossRef]
- Trusty, J.L.; Kesler, H.C.; Delgado, G.H. Vascular flora of Isla del Coco, Costa Rica. Proc. Calif. Acad. Sci. 2006, 57, 247–355. [Google Scholar]
- Kornás, J. Notes on African Hymenophyllaceae 2. Trichomanes lenormandii v. d. Bosch New to Continental Africa. Bull. Jard. Bot. Natl. Belg. Bull. Van Natl. Plantentuin Van Belg. 1976, 46, 393–397. [Google Scholar] [CrossRef]
- Bragança Guarnier, G.; da Silva Sylvestre, L.; Abraão Araújo da Silva, I. Checklist preliminar da biodiversidade de pteridófitas do Parque Natural Municipal do Curió. In Parque do Curió, Frago, M.E., Ed.; Missão Asa Editorial: 2020; pp. 43–54. Available online: https://www.researchgate.net/profile/Eduardo-Silva-Neto/publication/348390297_Solos_e_Fragilidade_Ambiental_do_Parque_Natural_Municipal_do_Curio_Paracambi_RJ/links/5ffc58bd299bf1408888fb2c/Solos-e-Fragilidade-Ambiental-do-Parque-Natural-Municipal-do-Curio-Paracambi-RJ.pdf (accessed on 12 December 2022).
- Lellinger, D.B. A Field Manual to the Ferns and Fern-Allies of the United States and Canada; Smithsonian Institution Press: Washington, DC, USA, 1985; p. 389. [Google Scholar]
- Prado, J.; Moran, R.C. Checklist of the ferns and lycophytes of Acre State, Brazil. Fern Gaz. 2009, 18, 230–263. [Google Scholar]
- Tardieu-Blot, M.L. Sur quelques Hymenophyllaceae des îles Mascareignes. Adansonia sér. 2 1977, 17, 147–150. [Google Scholar]
- Farrar, D.R.; Parks, J.C.; McAlpin, B.W. The fern genera Vittaria and Trichomanes in the northeastern United States. Rhodora 1983, 85, 83–92. [Google Scholar]
- Pinson, J.B.; Chambers, S.M.; Sessa, E.B. Vittaria graminifolia (Pteridaceae) and Didymoglossum petersii (Hymenophyllaceae) in Broxton Rocks, GA. Am. Fern J. 2017, 107, 257–264. [Google Scholar] [CrossRef]
- Martínez-Meléndez, N.; Perez-Farrera, M.A.; Martinez-Camilo, R. The vascular epiphyte flora of El Triunfo biosphere reserve, Chiapas, México. Rhodora 2009, 111, 503–535. [Google Scholar] [CrossRef]
- Fosberg, F.R.; Sachet, M.-H. Systematic studies of Micronesian plants. Smiths. Contr. Bot. 1980, 1–40. Available online: https://repository.si.edu/bitstream/handle/10088/6955/scb-0045.pdf?sequence=1&isAllowed=y (accessed on 12 December 2022).
- León, B. Hymenophyllaceae endémicas del Perú. Rev. Peru. Biol. 2006, 13, 903–904. [Google Scholar] [CrossRef]
- Kessler, M.; Smith, A.R.; Sundue, M. Three new Andean species of Hymenophyllum (Hymenophyllaceae–Pteridophyta). Blumea 2005, 50, 555–559. [Google Scholar] [CrossRef]
- Kirby, C. Field Guide to New Zealand’s Epiphytes, Vines & Mistletoes; Environmental Research Institute, University of Waikato: Hamilton, NZ, USA, 2014. [Google Scholar]
- Oliver, W.R.B. New Zealand epiphytes. J. Ecol. 1930, 18, 1–50. [Google Scholar] [CrossRef]
- Gonzatti, F.; Windisch, P.G. Flora of Espírito Santo: Hymenophyllum (Hymenophyllaceae). Rodriguésia 2018, 69, 611–629. [Google Scholar] [CrossRef]
- Benson, D.; McDougall, L. Ecology of Sydney plant species Part 1: Ferns, fern-allies, cycads, conifers and dicotyledon families Acanthaceae to Asclepiadaceae. Cunninghamia 1993, 3, 257–422. [Google Scholar]
- Jarman, S.J.; Kantvilas, G.; Brown, M.J. The ecology of pteridophytes in Tasmanian cool temperate rainforest. Fern Gaz. 1986, 13, 77–86. [Google Scholar]
- Brass, L.J. Ferns of a New Guinea gully. Am. Fern J. 1953, 43, 150–158. [Google Scholar] [CrossRef]
- Nwe, T.Y.; Zhang, X.-C. Taxonomic studies of lycophytes and ferns from the Pan-Himalaya (I): Hymenophyllum (Hymenophyllaceae). Turczaninowia 2017, 20, 75–96. [Google Scholar]
- Chen, C.W.; Ebihara, A.; Cheng, K.Y.; Hsu, T.C.; Huang, Y.M.; Dang, V.D.; Luu, H.T.; Le, V.S.; Li, C.W. Studies of Vietnamese pteridophyte flora 1. Syst. Bot. 2021, 46, 573–581. [Google Scholar] [CrossRef]
- Larsen, C.; Ponce, M.M.; Scataglini, M.A. Revisión de las especies de Hymenophyllum (Hymenophyllaceae) del sur de Argentina y Chile. Gayana. Bot. 2013, 70, 275–330. [Google Scholar] [CrossRef]
- Carvalho, F.A.; Salino, A.; Zartman, C.E. New country and regional records from the Brazilian side of Neblina Massif. Am. Fern J. 2012, 102, 228–232. [Google Scholar] [CrossRef]
- Gonzatti, F.; Windisch, P.G.; Scariot, F.J.; Echeverrigaray, S.; Ritter, M.R. Revision of Hymenophyllum subg. Sphaerocionium (Hymenophyllaceae) in the Atlantic Forest domain (Brazil), based on molecular and morphological evidence. Syst. Bot. 2020, 45, 707–748. [Google Scholar] [CrossRef]
- Kornás, J. Filmy ferns (Hymenophyllaceae) of Central Africa (Zaire, Rwanda, Burundi). 1. Hymenophyllum. Fragmenta Flor. Geobot. Polonica 1993, 38, 3–19. [Google Scholar]
- Diem, J.; Lichtenstein, J.S.D. Las Himenofiláceas del área argentino-chilena del sur. Darwiniana 1959, 11, 611–760. [Google Scholar] [CrossRef]
- Larsen, C.; Gonzatti, F.; Acosta, J.M.; Ponce, M.M. Morphological and molecular evidence to segregate a disjunct species of Hymenophyllum (Hymenophyllaceae) from Southern South America. Syst. Bot. 2020, 45, 439–449. [Google Scholar] [CrossRef]
- Pincheira-Ulbrich, J.; Zambrano, U.; Urrutia-Estrada, J. New record of Hymenophyllum caudatum Bosch (Polypodiopsida, Hymenophyllaceae) extends the mainland distribution in the coastal Mediterranean Forest of South America. Biodiv. Data J. 2022, 10. [Google Scholar] [CrossRef]
- Larsen, C.; Arana, M.D.; Acosta, J.M.; Ponce, M.M. Two new species segregated from Hymenophyllum tunbrigense (Hymenophyllaceae) in southern South America, based on morphological, anatomical, molecular and distributional evidence. Phytotaxa 2017, 303, 218–232. [Google Scholar] [CrossRef]
- Carvajal-Hernández, C.I.; Krömer, T. Riqueza y distribución de helechos y licófitos en el gradiente altitudinal del Cofre de Perote, centro de Veracruz, México. Bot. Sci. 2015, 93, 601–614. [Google Scholar] [CrossRef]
- Christenhusz, M.J.; Schwartsburd, P.B.; Labiak, P.H. Hymenophyllum filmenofilicum (Hymenophyllaceae, Pteridophyta): A new epipetric filmy fern from Paraná, southern Brazil. Kew Bull. 2009, 64, 175–178. [Google Scholar] [CrossRef]
- Manickam, V.S. Rare and endangered ferns of the Western Ghats of South India. Fern Gaz. 1995, 15, 1–10. [Google Scholar]
- Hsu, T.C.; Huang, Y.F.; Chao, Y.S. Systematics of Hymenophyllum subgenus Mecodium (Hymenophyllaceae) in Taiwan. Syst. Bot. 2019, 44, 753–767. [Google Scholar] [CrossRef]
- Godley, E.J. The flora of Antipodes Island. N. Z. J. Bot. 1989, 27, 531–564. [Google Scholar] [CrossRef]
- Olivera Tarifeño, C.M.; Ramos Sandoval, J.D. Determinación Taxonomica, Distribucion y Descripcion de los Helechos (Div. Pteridophyta) en el Parque Nacional de Cutervo; Cutervo–Region Cajamarca. Lic.: Lambayeque, Peru, 2014. [Google Scholar]
- Singh, B.; Singh, V.N.; Phukan, S.J.; Sinha, B.K.; Borthakur, S.K. Contribution to the pteridophytic flora of India: Nokrek Biosphere Reserve, Meghalaya. J. Threatened Taxa 2012, 4, 2277–2294. [Google Scholar] [CrossRef]
- Fowler, R.L. Annotated list of ferns of the Kilauea-Mauna Loa Section of Hawaii National Park. Am. Fern J. 1940, 30, 9–18. [Google Scholar] [CrossRef]
- Ebihara, A. Hymenophyllaceae collected in the Mt. Kinabalu Area in 2000 and 2007. Mem. Nat. Mus. Nat. Sci. 2008, 45, 105–110. [Google Scholar]
- Copeland, E.B. Papuan pteridophytes collected for the Arnold Arboretum by LJ Brass. J. Arnold Arb. 1929, 10, 174–182. [Google Scholar] [CrossRef]
- Gibby, M.; Lovis, J.D. New ferns of Madeira. Fern Gaz. 1989, 13, 285–290. [Google Scholar]
- Fineran, B.A. A botanical survey of seven mutton-bird islands, south-west Stewart Island. J. R. Soc. N. Z. 1973, 3, 475–525. [Google Scholar] [CrossRef]
- Ríos Saldaña, R.; Rincón, G.; Rafael, R.; Santos Mido, M. Guía de Helechos y Plantas Afines del PARQUE Nacional Volcán Barú; SENACYT: Clayton, Panama, 2016. [Google Scholar]
- Rio, C.D.; Hennequin, S.; Rouhan, G.; Ebihara, A.; Lowry, P.P., II; Dubuisson, J.-Y.; Gaudeul, M. Origins of the fern genus Hymenophyllum (Hymenophyllaceae) in New Caledonia: Multiple independent colonizations from surrounding territories and limited in situ diversification. Taxon 2017, 66, 1041–1064. [Google Scholar] [CrossRef]
- Barcelona, J.F. Preliminary report on the ferns and fern allies (pteridophytes) of Mt. Bali-it, Balbalasang-Balbalan National Park, Kalinga, Northern Luzon, Philippines. Sylvatrop 2003, 13, 81–92. [Google Scholar]
- Ebihara, A.; Iwatsuki, K.; Ohsawa, T.A.; Ito, M. Hymenophyllum paniense (Hymenophyllaceae), a new species of filmy fern from New Caledonia. Syst. Bot. 2003, 28, 228–235. [Google Scholar]
- Lindsay, S.; Middleton, D.J.; Saengrit, S. Hymenophyllum pilosissimum C. Chr.(Hymenophyllaceae), a new record for Thailand. Thai For. Bull. (Bot.) 2014, 42, 48–51. [Google Scholar]
- Moore, S.-J.; Chen, C.-M.; Wang, J.-C. Hymenophyllum pilosissimum C. Chr. (Hymenophyllaceae), a new recorded fern from Taiwan. Am. Fern J. 2010, 100, 180–183. [Google Scholar] [CrossRef]
- Perrie, L.R.; Shepherd, L.D.; de Lange, P.J.; Batty, E.L.; Ohlsen, D.J.; Bayly, M.J.; Brownsey, P.J. Hymenophyllum pluviatile, a new and uncommon fern from New Zealand. N. Z. J. Bot. 2013, 51, 308–320. [Google Scholar] [CrossRef]
- Fall, P.L.; Drezner, T.D. Vascular plant species of the Kingdom of Tonga by vegetation type, species origin, growth form, and dispersal mechanism. Ecology 2020, 101, e02902. [Google Scholar] [CrossRef]
- Janzen, D.H.; Liesner, R. Annotated check-list of plants of lowland Guanacaste Province, Costa Rica, exclusive of grasses and non-vascular cryptogams. Brenesia 1980, 18, 15–90. [Google Scholar]
- Klein, V.P.; Demarchi, L.O.; Quaresma, A.C.; Cruz, J.d.; Piedade, M.T.F. The vascular epiphyte flora in a white-sand ecosystem of the Uatumã Sustainable Development Reserve, Central Amazon. Check List 2022, 18, 157–186. [Google Scholar] [CrossRef]
- Meave, J.A.; Rincon-Gutierrez, A.; Ibarra-Manriquez, G.; Gallardo-Hernandez, C.; Romero-Romero, M.A. Checklist of the vascular flora of a portion of the hyper-humid region of La Chinantla, Northern Oaxaca Range, Mexico. Bot. Sci. 2017, 95, 722–759. [Google Scholar] [CrossRef]
- Sánchez Villaverde, C. Tres nuevas especies de Hymenophyllaceae para Cuba. Rev. Jard. Bot. Nac. 1987, 8, 9–16. [Google Scholar]
- Perrie, L.R.; Shepherd, L.D.; Brownsey, P.J.; Larraín, J.; Shaw, B.; Thouvenot, L.; von Konrat, M. Rediscovery and reinstatement of the New Caledonian endemic filmy fern Hymenophyllum pumilio Rosenst. N. Z. J. Bot. 2016, 54, 1–10. [Google Scholar] [CrossRef]
- Dubuisson, J.Y.; Hennequin, S.; Droissart, V. Hymenophyllum senterreanum Dubuisson & Deblauwe, sp. nov.(Hymenophyllaceae) and its relatives in western Central Africa. Phytotaxa 2016, 257, 287–294. [Google Scholar]
- Rojas-Alvarado, A.F. Una especie nueva de Hymenophyllum y una variedad nueva de Trichomanes collariatum Bosch (Filicales: Hymenophyllaceae) en Costa Rica. Lankesteriana 2004, 4, 143–148. [Google Scholar] [CrossRef]
- Ratcliffe, D.A.; Birks, H.J.B.; BIRKS, H.H. The ecology and conservation of the Killarney fern Trichomanes speciosum WILLD. in Britain and Ireland. Biol. Conserv. 1993, 66, 231–247. [Google Scholar] [CrossRef]
- Richards, P.W.; Evans, G.B. Biological flora of the British Isles: Hymenophyllum. J. Ecol. 1972, 60, 245–268. [Google Scholar] [CrossRef]
- Wilmanns, O.; Rasbach, H. Observations on the pteridophytes of Sao Miguel, Acores. Fern Gaz. 1973, 10, 315–329. [Google Scholar]
- Willmot, A. An ecological survey of the ferns of the Killarney District, Co. Kerry, Ireland. Brit. Fern Gaz. 1983, 12, 249–265. [Google Scholar]
- Aguiar, S.; Herrero, A.; Quintanilla, L.G. Confirmación citológica de la presencia de Hymenophyllum wilsonii en España. Lazaroa 2006, 27, 129–131. [Google Scholar]
- Sánchez Velázquez, T. Hymenopyllum wilsonii Hook. (Pteridophyta, Hymenophyllaceae). Confirmación de su presencia en las Islas Canarias. Bot. Macaronésica 2003, 24, 207–2011. [Google Scholar]
- Rasbach, H.; Rasbach, K.; Bennert, H.W. New records and new cytological results for the fern flora of Madeira. Brit. Fern Gaz. 1990, 13, 391–395. [Google Scholar]
- Lee, C.S.; Lee, K.; Lee, S.G.; Ebihara, A. A new taxon of Hymenophyllum (Hymenophyllaceae): H. wrightii f serratum. Korean J. Plant Taxon. 2014, 44, 233–237. [Google Scholar] [CrossRef]
- Mallmann, I.T.; Schmitt, J.L. Richness and floristic composition of the fern community in riparian forest of the river ‘Cadeia’, in Rio Grande do Sul state, Brazil. Cienc. Florest. 2014, 24, 97–109. [Google Scholar] [CrossRef]
- Ponce, M.M.; del Rio, C.; Ebihara, A.; Dubuisson, J.Y. Discussion on taxonomy of the fern genera Crepidomanes and Polyphlebium (Hymenophyllaceae) in Argentina and south-eastern South America, and description of a new local variety for Crepidomanes pyxidiferum. Bot. Lett. 2017, 164, 5–18. [Google Scholar] [CrossRef]
- Croxall, J.P. The Hymenophyllaceae of Queensland. Austr. J. Bot. 1975, 23, 509–547. [Google Scholar] [CrossRef]
- Windisch, P.G. Trichomanes crispum L. (Pteridophyta, Hymenophyllaceae) and allied species. Bradea 1992, VI, 78–117. [Google Scholar]
- Bauret, L.; Grall, A.; Senterre, B.; Rouhan, G.; Hennequin, S.; Ebihara, A.; Dubuisson, J.-Y. New circumscription of Trichomanes cupressoides Desvaux (Hymenophyllaceae), an endemic filmy fern from the Seychelles (Indian Ocean), and new insights into the genus Abrodictyum C.Presl in the western Indian Ocean. Phytotaxa 2015, 202, 14. [Google Scholar] [CrossRef]
- Rojas-Alvarado, A.F. Novelties in Trichomanes l. From Colombia. Caldasia 2009, 31, 9–11. [Google Scholar]
- Moran, R.C. Trichomanes resinosum (Hymenophyllaceae), a new species from southern Venezuela and adjacent Guyana. Brittonia 2000, 52, 238–240. [Google Scholar] [CrossRef]
- Pacheco, L. Trichomanes ribae (Hymenophyllaceae), a new filmy fern from Costa Rica and Panama. Am. Fern J. 2002, 92, 292, 294–295. [Google Scholar] [CrossRef]
- Ebihara, A.; Matsumoto, S.; Ito, M. Taxonomy of the reticulate Vandenboschia radicans complex (Hymenophyllaceae) in Japan. Acta Phytotax.Geobot. 2009, 60, 26–40. [Google Scholar]
- Bidin, A.A.; Jaman, R. The pteridophytes of Tawau Hills Park, Sabah. ASEAN Rev. Biodiv. Environ. Cons. (ARBEC) 1999, 1, 1–11. [Google Scholar]
- Ward, C.M. The pteridophytes of Flores (Açores): A survey with bibliography. Brit. Fern Gaz. 1970, 10, 119–126. [Google Scholar]
- Vumba Nature. Available online: http://www.vumba-nature.com/ (accessed on 12 October 2022).
- Available online: herbario.zamorano.edu/handle/11036/1677 (accessed on 10 October 2022).
- Available online: https://fm-digital-assets.fieldmuseum.org/1140/158/C0628078F.jpg (accessed on 12 October 2022).




{kind=link}
{kind=link}
{kind=link}
| Key to Quantify Verbal Information | % Epiphytic Occurrence (=EV) |
|---|---|
| epiphyte and lithophyte (50% to L)/epiphyte or terrestrial/facultative epiphyte (50% to T) | 50 |
| epiphyte, lithophyte, or terrestrial (33.3% to L and T) | 33.4 |
| holoepiphyte/hemiepiphyte/epiphyte | 100 |
| obligatory epiphyte | 100 |
| rarely/seldom | 5 |
| very rarely/accidental epiphyte/exceptionally | 1 |
| also as/sometimes/occasionally/uncommonly | 15 |
| commonly/generally/ mainly/mostly/preferentially/primarily/usually | 85 |
| often/frequently/most frequently | 67 |
| less frequently | 33 |
| Genus | Description in [19] | EV | LV | TV | EV ≥ 0.5 | % Epiphytes in EpiList 1.0 |
|---|---|---|---|---|---|---|
| Abrodictyum | Terrestrial in humid places, or epiphytic on tree trunks | 19 | 14 | 67 | 21 | 37 |
| Callistopteris | Usually epilithic, occasionally epiphytic | 1 | 8 | 91 | 0 | 40 |
| Cephalomanes | Terrestrial or epilithic | 5 | 15 | 80 | 0 | 20 |
| Crepidomanes | Mostly epiphytic | 64 | 32 | 4 | 80 | 71 |
| Didymoglossum | Epilithic or low-epiphytic | 71 | 24 | 5 | 85 | 92 |
| Hymenophyllum | Usually low- to middle-epiphytes on tree trunks, sometimes canopy sun epiphytes, occasionally epilithic or terrestrial | 75 | 18 | 7 | 85 | 89 |
| Polyphlebium | Epilithic or epiphytic | 78 | 15 | 7 | 85 | 88 |
| Trichomanes | Usually terrestrial in humid places, sometimes epiphytic on tree trunks | 36 | 15 | 49 | 34 | 57 |
| Vandenboschia | Hemi-epiphytic on tree trunks or epilithic, occasionally terrestrial | 64 | 24 | 12 | 74 | 92 |
| whole family | 63 | 19 | 18 | 71 | 72 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zotz, G.; Einzmann, H.J.R. How Epiphytic Are Filmy Ferns? A Semi-Quantitative Approach. Diversity 2023, 15, 270. https://doi.org/10.3390/d15020270
Zotz G, Einzmann HJR. How Epiphytic Are Filmy Ferns? A Semi-Quantitative Approach. Diversity. 2023; 15(2):270. https://doi.org/10.3390/d15020270
Chicago/Turabian StyleZotz, Gerhard, and Helena J. R. Einzmann. 2023. "How Epiphytic Are Filmy Ferns? A Semi-Quantitative Approach" Diversity 15, no. 2: 270. https://doi.org/10.3390/d15020270
APA StyleZotz, G., & Einzmann, H. J. R. (2023). How Epiphytic Are Filmy Ferns? A Semi-Quantitative Approach. Diversity, 15(2), 270. https://doi.org/10.3390/d15020270