Abstract
We provide a diversity assessment of the agamid genus Phrynocephalus Kaup, 1825. We analyze COI mtDNA barcodes from 385 individuals sampled all over Phrynocephalus range. We apply the ABGD, ASAP, bGMYC, mlPTP and hsPTP species delimitation algorithms to analyze the COI gene fragment variation and assess the species diversity in Phrynocephalus. Nine species groups are revealed in Phrynocephalus in agreement with earlier studies on the phylogenetic relationships of the genus. We demonstrate that the present taxonomy likely underestimates the actual diversity of the genus. Alternative species delimitation algorithms provide a confusingly wide range of possible number of Phrynocephalus species—from 54 to 103 MOTUs (molecular operational taxonomic units). The ASAP species delimitation scheme recognizing 63 MOTUs likely most closely fits the currently recognized taxonomic framework of Phrynocephalus. We also report on 13 previously unknown Phrynocephalus lineages as unverified candidate species. We demonstrate that the ASAP and the ABGD algorithms likely most closely reflect the actual diversity of Phrynocephalus, while the mlPTP and hsPTP largely overestimate it. We argue that species delimitation in these lizards based exclusively on mtDNA markers is insufficient, and call for further integrative taxonomic studies joining the data from morphology, mtDNA and nuDNA markers to fully stabilize the taxonomy of Phrynocephalus lizards.
Keywords:
taxonomy; DNA barcodes; species delimitation; Reptilia; ABGD; ASAP; bGMYC; mlPTP; hsPTP; mtDNA; cryptic species 1. Introduction
Toad-headed agamas of the genus Phrynocephalus Kaup, 1825 represent one of the taxonomically and morphologically most diverse groups of Central Asian reptiles, being an ecologically important component of all major deserts of Middle and Central Asia, and large areas in the Near and the Middle East [1,2]. Despite its long taxonomic history since 1771 when Pallas described the first species of Phrynocephalus as Lacerta helioscopa (Pallas, 1771) [3], systematics of the genus still remain in a state of flux [4]. The extensive taxonomic confusion in the genus Phrynocephalus persisting since the time of active scientific exploration of Central Asia in mid-nineteenth century continues today. For example, as the total number of taxa formally named within Phrynocephalus exceeds 160, more than a hundred of the proposed names are presently regarded as junior synonyms [1]. There is no agreement on the number of species within the genus; according to various estimates, it ranges from 26 to 50 species [1,2,4,5,6,7,8,9]. A number of previous works, e.g., [1,4,7], contained inconsistent taxonomic information or used GenBank sequences derived from misidentified or mislabeled specimens [2]. Finally, the different author teams often tend to use alternative taxonomies, and nomenclature of the genus is still far from stable [1,2,4,10,11].
Based on recent molecular phylogenetic studies on mitochondrial DNA genealogy [12] and a multilocus phylogeny, which included both mitochondrial (mtDNA) and nuclear (nuDNA) protein-coding genes [2], based on the analysis of 51 samples representing ca. 33 species of Phrynocephalus, the genus was divided into the following ten species groups: (1) Microphrynocephalus (encompassing the P. interscapularis group), (2) Phrynosaurus (encompassing the P. scutellatus group), (3) Oreosaura (encompassing the P. vlangalii group), (4) Megalochilus (encompassing the P. mystaceus group), (5) P. maculatus group (referred to as “Arabian group” in [2]), (6) Helioscopus (encompassing the P. helioscopus group), (7) P. raddei group, (8) P. guttatus group, (9) P. versicolor group (referred to as P. przewalskii group herein), and (10) an orphaned species P. axillaris. More recently, [4] analyzed phylogenetic relationships of Phrynocephalus based on mtDNA, nuDNA and allozyme dataset partly overlapping with that of Solovyeva et al. [2], which included 46 individuals representing 29 Phrynocephalus nominal species. The analysis of Macey et al. [4] recovered essentially the same species groups though with a different topology of phylogenetic relationships among them and generally lower support values for most of the basal nodes.
A number of recent studies have focused on molecular taxonomy and phylogenetic relationships of various Phrynocephalus species or species complexes [7,8,13,14,15,16,17,18,19,20,21] and on structure of complete mitochondrial genomes [22,23,24,25,26]. However, there is a lack of comparative phylogenetic works based on large datasets for the whole genus, thus, further studies are needed to access the genetic diversity across the genus Phrynocephalus, estimate the levels of cryptic diversity, and eventually to stabilize the taxonomy of the group.
Scientists are rapidly developing large DNA barcode libraries, DNA sequences of specific genes for species across the tree of life, in order to document and conserve biodiversity [27,28]. DNA barcoding has proven to be an invaluable tool for identification of organisms [27,28,29], and is widely applied in biodiversity surveys [30], conservation [31], collection management [32], taxonomy, including the identification of taxa in need of further systematic study [33] and the discovery of yet undescribed species [34,35,36]. COI DNA barcoding, focused on the sequencing of a single standardized genetic marker (in the case of animals cytochrome oxidase subunit I, COI) has been widely used to study species diversity in many groups of vertebrates [37,38,39,40,41,42,43], including reptiles [44,45,46,47,48]. Although COI barcoding data can, in some cases, be used in studies on phylogenetic relationships and phylogeography, their primary application lies in species discovery and identification [29]. As such, COI barcoding was successfully applied for assessment of cryptic diversity across several species groups of the genus Phrynocephalus [13,14,15,17,18,19,21].
Novel statistical methods for delimitation of the number of MOTUs (molecular operational taxonomic units, roughly corresponding to putative “species”) from DNA-barcode data represent a promising approach in biodiversity studies [49]. It is important to emphasize that species delimitation based on the COI gene data must be combined with other lines of evidence, such as nuclear DNA genetic markers, or the data from the integrative taxonomic analyses (including data on morphology, ecology or distribution), for a more reliable estimate of species numbers [49,50]. In general, when the existence of new species is suggested by DNA barcoding, this should be taken rather as a signal for further investigation, suggesting that data from other independent lines of evidence are required to make the tentative species delimitation more reliable [51,52,53]. The recent progress in computer models for species delimitation based on the Bayesian interference or other algorithms allows to evaluate alternative clustering patterns of the identified mtDNA lineages and to statistically determine the most stable clustering options likely corresponding to the putative species boundaries [50].
Creating comprehensive COI reference databases will help with preliminary identification of species, morphologically cryptic lineages, and specimens in scientific collections. This is particularly important for reptiles, the most species-rich vertebrate group on Earth, yet lacking a comprehensive understanding of the extinction risk, with only 45% of described reptile species have been assessed by IUCN to date [54]. Here we aim to: (i) construct a comprehensive COI reference library providing a solid basis for species identification of a taxonomically challenging lizard genus Phrynocephalus across the Middle East and Central Eurasia (including genetic information from 385 specimens from all over the genus range); (ii) preferably analyze the materials collected from the vicinity of type localities of nominal taxa, thus allowing a more reliable attribution of the mtDNA lineages to the currently recognized species; (iii) investigate the existence of a barcoding gap in Phrynocephalus; and (iv) evaluate the use of five phylogenetic species concept-based approaches (bGMYC, ABGD, ASAP, mlPTP and hsPTP) for delimiting species in Phrynocephalus using the COI barcoding gene region. Based on the results of our analyses we estimate the performance of each species delimitation method from COI barcoding, scrutinize diversity within the genus, and discuss the newly obtained data in the scope of unresolved problems of Phrynocephalus taxonomy.
2. Materials and Methods
2.1. Taxon Sampling
Tissue samples were taken from 285 Phrynocephalus specimens from the herpetological collection of the Zoological museum of Moscow State University (ZMMU) and 24 specimens from other herpetological collections (Institute of Zoology Academy of Sciences of the Republic of Uzbekistan (IZUAS); Kunming Institute of Zoology, Chinese Academy of Sciences, Kunming, Yunnan, China, KIZ) from across the entire range of the genus Phrynocephalus (Figure 1). We obtained biological samples of Phrynocephalus from different scientific collections (see details in BOLD projects DS-ABLRP and DS-NPLRP [55]). Altogether, we analyzed COI sequence data for 385 specimens of Phrynocephalus, including 99 newly generated sequences, 204 sequences obtained during our previous studies of the group [2,12,13,14,15,19,21], and 82 sequences which were downloaded from GenBank (see Supplementary Table S1). The taxonomic framework generally follows Solovyeva et al. [2], Barabanov and Ananjeva [1], and the recent taxonomic reviews of Phrynocephalus [13,14,15,17,18,19,21].
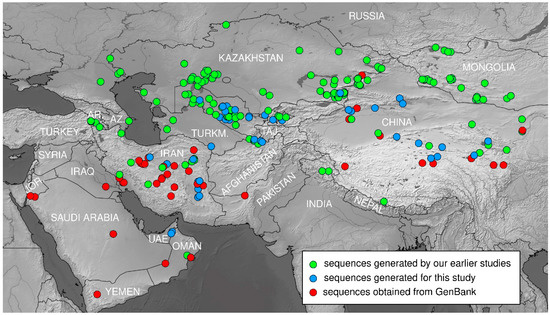
Figure 1.
Map showing distribution of the genus Phrynocephalus and location of studied populations. For the detailed information on the localities and voucher specimens see Supplementary Table S1. References for sequences generated by our earlier studies are following [1,12,13,15,19,21]. Abbreviations: UAE—United Arab Emirates; JOR.—Jordan; AR.—Armenia; AZ.—Azerbaijan; TURKM.—Turkmenistan; UZB.—Uzbekistan; TAJ.—Tajikistan; KYR.—Kyrgyzstan; NEP.—Nepal. Base map created using simplemappr.net.
We analyzed multiple samples and localities for widespread species complexes of Phrynocephalus, and overall, our sampling included COI sequences for representatives of all currently recognized nominal species of the genus with the exception of the following five species: P. clarkorum Anderson and Leviton, P. euptilopus Alcock and Finn, P. golubewii Shenbrot and Semenov, P. luteoguttatus Boulenger, and P. roborowskii Bedriaga. When possible, we included in the analysis materials collected from the type localities of nominal taxa or in their vicinity (radius of 50 km), for a more reliable attribution of the mtDNA lineages to the currently recognized Phrynocephalus species. Altogether our analyses include sequence data from topotypes of 33 Phrynocephalus taxa (see Supplementary Table S1 for details). Sequences of four closely related agamids Paralaudakia caucasia (Eichwald), Par. lehmanni (Nikolsky), Par. microlepis (Blanford), and Stelllagama stellio (Linnaeus) were used as outgroups in all phylogenetic analyses. For a list of the species covered by our sampling, museum voucher and locality information see Supplementary Table S1.
2.2. DNA Extraction, Amplification and Sequencing
We analyzed a 654 bp fragment of the cytochrome oxidase I subunit (COI) mitochondrial DNA. Molecular analysis was conducted (i) at the Department of Vertebrate Zoology, Faculty of Biology, Lomonosov Moscow State University (MSU), Moscow, Russia, and (ii) was outsourced for DNA isolation and sequencing to the core analytical facility at the Canadian Centre for DNA Barcoding (CCDB), Biodiversity Institute of Ontario, University of Guelph, Guelph, Canada. In the MSU laboratory, the total genomic DNA was extracted using the standard phenol-chloroform extraction protocol [56], PCR amplification was performed using MyCycler BioRad under conditions described by Ivanova et al. [57]. Two primers pairs were used for PCR and sequencing: VF1d (5’-TTCTCAACCAACCACAARGAYATYGG-3’, forward) + VR1d (5’-TAGACTTCTGGGTGGCCRAARAAYCA-3’, reverse) [56]; and Rep-COI-F (5’-TNTTMTCAACNAACCACAAAGA-3’, forward) + Rep-COI-R (5’-ACTTCTGGRTGKCCAAARAATCA-3’, reverse) [44]. PCR reaction volume was 20 μL and it contained ca. 100 ng of template DNA, 0.3 pM/μL of each PCR primer, 1xTaq-buffer with 25 mM of MgCl2 (Silex, Moscow, Russia), 0.2 mM dNTPs, and 1 unit of Taq-polymerase (Silex, Moscow, Russia; 5 units/μL). The results of the amplification were examined using electrophoresis in 1% agarose gel in the presence of ethidium bromide.
For processing in CCDB, tissues were arrayed into 96-well microplates (following [58,59,60]) and submitted for molecular analysis. Prior to DNA extraction, each plate well was filled with 50 μL of lysis buffer with Proteinase K and the plates were incubated overnight (12–18 h) at 56 °C, followed by a robotic standard glass fiber DNA extraction protocol [57]. PCR conditions and primers were as described above. Sequences, electrophoresis data (for both successful and unsuccessful amplifications), and other specimen information for the individuals analyzed through the BOLD “Lizards of the Palearctic” projects DS-ABLRP and DS-NPLRP are available on the Barcode of Life website [55]. All sequences have been deposited in GenBank and are detailed in Supplementary Table S1.
2.3. Phylogenetic Analyses
Sequences were aligned using Seqman 5.06 and verified using BioEdit Sequence Alignment Editor 7.1.3.0 [61]. Mean inter- and intraspecific uncorrected genetic p-distances and sequence characteristics were calculated in MEGA X [62]. Histograms showing the distribution of pairwise divergences were generated in LibreOffice Calc (https://www.libreoffice.org/discover/calc/, accessed on 2 August 2022).
We applied the Bayesian inference (BI) and maximum likelihood (ML) approaches to infer gene trees for the full COI dataset containing 385 sequences of Phrynocephalus and four sequences of outgroup Agaminae taxa, and a reduced dataset of the revealed COI haplotypes (163 sequences, including 159 Phrynocephalus haplotypes). The BI approach was implemented using MrBayes v.3.1.2 [63,64] and BEAST v1.8.2 [65] software. PartitionFinder v1.0.1 [66] was used to estimate the optimal nucleotide substitution models, resulting in HKY + G as the best-fit model for all three codon-partitions of the COI gene, as suggested by the Akaike information criterion (AIC). In MrBayes, phylogenetic analysis was performed with two simultaneous runs, each with four chains, for five million generations, 10% of generations were cut as burn in. The convergence of the runs was verified to make sure that the effective sample sizes (ESS) were all above 200 by examining the likelihood plots using TRACER v.1.5 [67].
In BEAST, phylogenetic analysis was run under the Yule coalescent model, assuming a strict molecular clock and fixing the substitution rate to 1. Two runs of ten million generations were conducted to reconstruct an ultrametric phylogenetic tree of haplotypes (N = 159). Parameter convergence was assessed using Tracer; the first 10% of generations were discarded as burn-in. The maximum clade credibility tree was generated using TreeAnnotator v1.8.2, distributed as part of the BEAST package.
The ML tree was generated using the IQ-TREE webserver [68]; preceded by the selection of substitution models using the Bayesian Information Criterion (BIC) in MODELFINDER [69], which supported TrN + I + G for COI codon positions 1 and 2, and GTR + F + I + G for position 3.
Confidence in nodal topology for BI analysis was estimated by calculating posterior probabilities (BI PP), and for ML analysis was assessed by 1000 bootstrap replications (ML BS). The nodes with BI PP values >0.95 and ML BS values 75% or above were a priori regarded as strongly supported; BI PP values between 0.95 and 0.90 and ML BS values between 75% and 50% were regarded as tendencies; while lower values were regarded as indicating not significant node support [70].
2.4. Species Delimitation
We followed the phylogenetic species concept (PSC) to delimit MOTUs as putative species using COI sequences. To assess the number of putative species-level lineages within the genus Phrynocephalus, we implemented the following five alternative PSC-based automated species delineation methods to estimate species boundaries from barcode data: (1) the Automatic Barcode Gap Discovery (ABGD; [32]), (2) Assemble Species by Automatic Partitioning (ASAP; [71]), (3) the Bayesian Generalized Mixed Yule-Coalescent model (bGMYC, [50]), (4) Maximum Likelihood Poisson Tree Process (mlPTP), and (5) heuristic search PTP (hsPTP), (the last two from Zhang et al. [72]) (for details see below). These methods enable the delimitation of independently-evolving species based on genetic data and do not require a priori hypotheses of putative species groupings, thereby limiting potential bias in species delimitation; they also demonstrated their effectiveness in a number of empirical studies [73,74,75,76].
All five species delineation methods implemented herein are based on analyses of a single gene and differ in the use of genetic information embedded in phylogenetic reconstructions. The performance of these approaches varies depending on speciation rates, population and sample sizes, and other parameters, with a tendency for oversplitting of putative species in PTP, and overlumping in ABGD [77]. However, in cases of concordant outcome of these methods, the resulting delimitation appears to be more plausible [77].
The ABGD method is an approach to statistical detection of barcoding gaps in a pairwise genetic distance distribution [32]. Identifying the existence of barcoding gap presumably occurring between intra- and interspecific distances [78,79] is important to accurately choose the species identification method. Barcoding gaps were used to partition the COI dataset into initial partition (species hypotheses). Resulting inferences were then recursively applied to yield finer recursive partitions until no further partitioning was possible. ABGD analysis was run on the COI dataset through a web-based interface (http://www.abi.snv.jussieu.fr/public/abgd/abgdweb.html, accessed on 3 July 2022) [32] using default parameters (ten steps of intraspecific divergence prior from Pmin = 0.001 to Pmax = 0.10, X = 2).
Recently, a more advanced method was developed on the base of ABGD [71]. ASAP—Assemble Species by Automatic Partitioning (ASAP) is a method to build species partitions from single locus sequence alignments. ASAP analysis was run through a web-based interface https://bioinfo.mnhn.fr/abi/public/asap/ using default parameters. Both in ABGD and ASAP analyses we computed a matrix of pairwise distances using the simple-distance model (p-distances).
The Generalized Mixed Yule-Coalescent model (GMYC) is an approach to identifying species ‘boundaries’ associated with shifts in branching rates between intra- and interspecies cladogenetic events, on a time-calibrated ultrametric tree by maximizing the likelihood score of the model [80,81]. This model relies on the expected abrupt changes in branching events between among-species branching events and a neutral coalescent process within-species. We used a Bayesian implementation of this method (bGMYC [50]), for which we obtained the distribution of ultrametric phylogenetic trees of COI haplotypes with BEAST v1.8.4 [65], and then used 100 random phylogenetic trees from the distribution of trees of haplotypes as an input for subsequent bGMYC analysis following [50]. We ran bGMYC for 50,000 generations with burn-in 40,000 and a thinning parameter of 100. We summarized results of bGMYC analyses in a matrix of pairwise co-assignment probabilities for each haplotype, shown as a heatmap (not presented).
The Poisson tree processes (PTP) approach [72] infers putative species boundaries on a given phylogenetic input tree by relying on the branch lengths, assessing the number of substitutions between branching events. The main assumption of this model is that the number of substitutions between species is significantly higher than the number of substitutions within species. We ran a PTP species delimitation analysis on the bPTP web server https://species.h-its.org/ptp/ [72]. We used the phylogeny of the COI dataset obtained through BI analysis as input data and implemented maximum likelihood (mlPTP) and heuristic search (hsPTP) algorithms. Outgroups were discarded before conducting the PTP analyses to avoid bias that could have arisen if some of the outgroup taxa were too distant [72]. We ran the PTP analysis for 500,000 MCMC generations, with a thinning value of 100, a burn in of 25% and we visually confirmed the convergence of the MCMC chain following recommendations by [72].
2.5. Evaluation of Species Delimitation Results
To evaluate results of different species delimitation methods we used LIMES software ver. 1.3 [82]. LIMES automatically calculates indexes for different partitions (one method—one partition), showing which methods are more congruent with each other, and which have a tendency for oversplitting. Along with the results from specific species delimitation software we tested the existence of a barcoding gap in Phrynocephalus, and tested the performance of two arbitrary thresholds of p-distance values of 3.0% and 5.0% of substitutions. We therefore regarded as a putative candidate ‘species’ those MOTUs which were concordantly recovered as a distinct entity by the most consensual species delimitation method.
3. Results
3.1. Sequence Characteristics
We obtained COI barcodes for 385 Phrynocephalus specimens, representing 63 taxa. Overall, 159 COI gene haplotypes for the genus Phrynocephalus were recovered in our dataset (see Supplementary Table S1). The sequenced fragment of the COI gene comprised up to 654 bp in length, among which 311 sites were identified as conservative, 343 as variable and 299 as parsimony-informative. Nucleotide composition analysis showed an anti-G bias: 30.9% (A), 27.6% (T/U), 27.9% (C), and 13.6% (G) typical for mtDNA genes. The transition-transversion bias (R) was estimated to be 4.20 (all data is given for in-group comparisons and for complete sequences only).
3.2. Phylogenetic Trees
Bayesian inference (BI) and maximum likelihood (ML) analyses resulted in essentially identical topologies, differing only in several not supported nodes (Figure 2, Figure 3 and Figure 4; ML-topology with node support values for ML and BI analyses shown in Supplementary Figure S1; ultrametric phylogenetic tree of the revealed COI haplotypes is shown in Supplementary Figure S2). In the resulting COI gene tree for Phrynocephalus, most basal nodes remained unresolved, but the nodes at the level of species groups were generally well-resolved and received strong support both in BI and ML analyses (see Figure 2, Figure 3 and Figure 4). Overall, the recovered topology was generally consistent with previous phylogenetic and phylogeographic studies on Phrynocephalus [2,7,12,13,14,15,17,18,19,21,83,84,85,86,87].
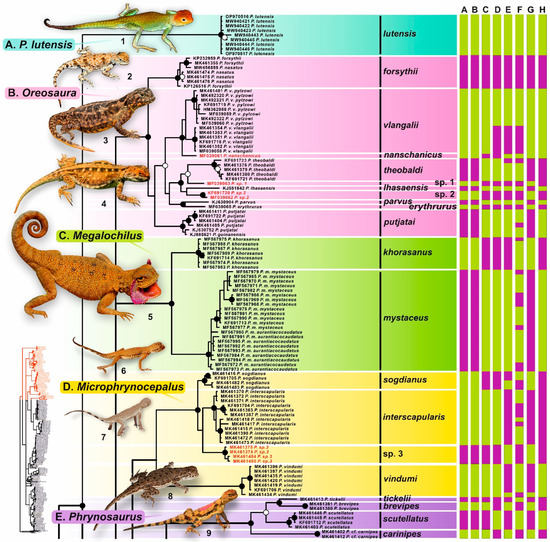
Figure 2.
Results of species delimitation analysis of Phrynocephalus plotted on the Bayesian inference phylogenetic tree. Part I. Black circles correspond to node support values (BI PP) > or = 0.99, white circles correspond to BI PP-values 0.95 < 0.99, no circles indicate nodes with support values lower than BI PP 0.95. Color bars on the right of each clade correspond to the results of species delimitation analyses based on: A—ABGD, B—ASAP1, C—ASAP2, G—bGMYC, E—mlPTP, F—hsPTP, G—3.0% p-distance threshold, and H—5.0% p-distance threshold. For voucher specimen information see Supplementary Table S1. Photos on thumbnails by R. A. Nazarov, E. A. Dunayev, E. N. Solovyeva and N. A. Poyarkov (not to scale): 1—P. lutensis; 2—P. forsythii; 3—P. vlangalii; 4—P. putjatai; 5—P. mystaceus; 6—P. inrescapularis; 7—P. sogdianus; 8—P. vindumi; 9—P. scutellatus.
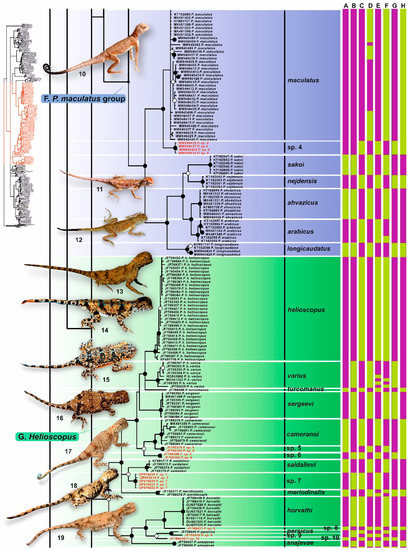
Figure 3.
Results of species delimitation analysis of Phrynocephalus plotted on the Bayesian inference phylogenetic tree (continued). Part II. For the meaning of the node markers and color bars depicting species delimitation analyses results see Figure 2. For voucher specimen information see Supplementary Table S1. Photos on thumbnails by R. A. Nazarov, E. A. Dunayev, E. N. Solovyeva and N. A. Poyarkov (not to scale): 10—P. maculatus; 11—P. sakoi; 12—P. ahvazicus; 13—P. helioscopus; 14—P. varius; 15—P. horvathi; 16—P. cameranoi; 17—P. saidalievi; 18—P. sergeevi; 19—P. persicus.
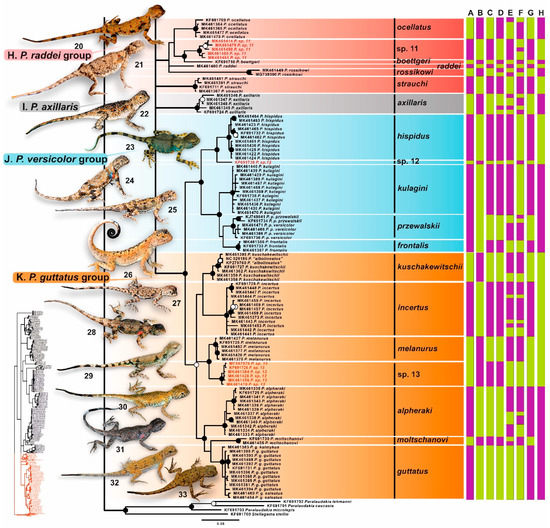
Figure 4.
Results of species delimitation analysis of Phrynocephalus plotted on the Bayesian inference phylogenetic tree (continued). For the meaning of the node markers and color bars depicting species delimitation analyses results see Figure 2. For voucher specimen information see Supplementary Table S1. Photos on thumbnails by R. A. Nazarov, E. A. Dunayev, E. N. Solovyeva and N. A. Poyarkov (not to scale): 20—P. ocellatus; 21—P. boettgeri; 22—P. strauchi; 23—P. axillaris; 24—P. hispidus; 25—P. kulagini; 26—P. przewalskii from China; 27—P. incertus; 28—P. przewalskii from Mongolia (“P. versicolor”); 29—P. melanurus; 30—P. kuschakewitschii; 31—P. alpheraki; 32—P. guttatus; 33—P. moltschanovi.
Monophyly of the genus Phrynocephalus received high support, with P. lutensis strongly suggested as a sister taxon to all remaining Phrynocephalus in all analyses (1.0/99, hereafter nodal support values given for BI PP/ML BS, respectively). Within the remaining Phrynocephalus species the Bayesian tree recognized eight major clades corresponding to the species groups as designated by Solovyeva et al. [2]: Oreosaura, Megalochilus, Phrynosaurus, maculatus group (referred to as ‘Arabian group’ in [2]), Helioscopus, P. axillaris, P. guttatus group, and P. przewalskii group (this group was referred to as P. versicolor group in Solovyeva et al. [2], see below). Monophyly of each species group received significant or high support (Figure 2, Figure 3 and Figure 4). Monophyly of two other species groups designated by Solovyeva et al. [2] was poorly or not supported: Microphrynocephalus and the P. raddei group, with P. strauchi not forming a clade with other members of the P. raddei group (Figure 4). Below we briefly describe the genealogical relationships, genetic differentiation and species group composition in Phrynocephalus as revealed by our analyses.
The basal split within Phrynocephalus is formed by P. lutensis (1.0/100), a recently described species of Phrynocephalus from central Iran [8]. The monophyly of the group including all remaining species of Phrynocephalus was not strongly supported (Figure 2, Figure 3 and Figure 4). Within the subgenus Oreosaura, consisting of the oviparous species of Qinghai-Tibetan Plateau (1.0/100), the clade including P. forsythii + P. nasatus (1.0/100) formed a sister lineage with respect to all other species of the group, represented in our analyses with P. erythrurus, P. theobaldi, P. lhasaensis, P. parvus, P. putjatai, P. vlangallii, P. nanschanicus and two previously unknown phylogenetic lineages, which we tentatively identify here as Phrynocephalus sp. 1–2. Phylogenetic relationships among the species of the Oreosaura clade were insufficiently resolved.
The monotypic subgenus Megalochilus (1.0/100) included the largest psammophilous toad-headed agama species, P. mystaceus, which was represented with two highly divergent reciprocally monophyletic lineages: one of them corresponds to P. mystaceus khorasanus from Iran (1.0/100) and another to the remaining populations from Middle Asia, Kazakhstan and southern Russia (1.0/99) (Figure 2).
Monophyly of the subgenus Microphrynocephalus received low support (0.85/91, see Figure 2). This group encompassed the small-sized psammophilous species from Middle East and Middle Asia; P. vindumi (1/100) from southwest Iran was suggested as sister lineage to the species of the P. interscapularis complex (1/100). Within the latter, three major lineages, corresponding to P. sogdianus, P. interscapularis and a previously unknown lineage which we identified as Phrynocephalus sp. 3.
The species of Phrynosaurus represented with the P. scutellatus complex formed a well-supported clade (1.0/100) with the P. maculatus group from Arabian Peninsula and Near East. Overall, the genealogical relationships within this clade were well-resolved. Within the P. scutellatus complex, our analysis revealed four highly divergent lineages with well-resolved genealogical relationships (1/100, see Figure 2).
The P. maculatus group included P. maculatus sensu stricto from central Iran and a new lineage Phrynocephalus sp. 4 from Abarkouh previously reported by Ebrahimipour et al. [87]. Together these lineages formed a sister clade (1.0/100) with respect to P. longicaudatus and the four species of the P. arabicus complex (including P. arabicus, P. ahvazicus, P. nejdensis and P. sakoi, 1.0/100; Figure 2). Genealogical relationships within the Helioscopus group, which included solid ground-dwelling species from Middle Asia, Iranian Plateau and Transcaucasia, were well-resolved and most nodes received high values of support (Figure 3). Numerous lineages, revealed within this clade, are clustered into two reciprocally monophyletic groups corresponding to the P. helioscopus species complex (with nine lineages from Middle Asia, including previously unidentified lineages Phrynocephalus sp. 5–7; 1.0/100), and to the P. persicus species complex (with six lineages from Iran and Transcaucasia, including three previously unknown lineages Phrynocephalus sp. 8–10; 1.0/100).
Monophyly of the P. raddei group sensu Solovyeva et al. [2] which includes small ground-dwelling species from southern Middle Asia, is not supported (0.65/51); this group includes six major clades with unresolved phylogenetic relationships: P. ocellatus, Phrynocephalus sp. 11, P. raddei, P. boetgeri, P. rossikowi, and P. strauchi; phylogenetic position of the latter species is not resolved (Figure 4). The phylogenetic affinity of P. axillaris, a species from Tarim Basin in western China, is also unresolved; this orphaned species forms a small monotypic clade (1.0/100) with shallow phylogenetic structuring (Figure 4).
The P. przewalskii group and P. guttatus group, joining species from steppes and deserts of the Inner Central Asia and northern Middle Asia, respectively, formed a well-supported clade (1.0/100, Figure 4). Monophyly of the P. przewalskii group was well supported (1.0/100), while monophyly of the P. guttatus group received only moderate support (0.97/94). The P. przewalskii group included the species from China, Mongolia and Tuva Republic in Russia; it consisted of two main subclades joining species from the western (two lineages assigned to P. hispidus and an unnamed lineage Phrynocephaus sp. 12 [2]), and eastern (P. kulagini, P. frontalis, and P. przewalski including P. versicolor) parts of Central Asia. The P. guttatus group included the species from the northern part of Middle Asia, Kazakhstan, and southern Russia forming three major subgroups: the P. guttatus complex (P. guttatus, P. moltschanovi and P. alpheraki; 0.99/96), the P. kuschakewitschii complex (P. incertus and P. kuschakewitschii; 0.96/85), and two lineages assigned to P. melanurus and an unnamed lineage Phrynocephaus sp. 13 (1.0/100) [2].
3.3. Barcoding Gap
A histogram showing the distribution of pairwise genetic p-distances demonstrates the presence of a putative barcoding gap at the level of 3.0% of substitutions in the COI gene, which generally corresponds to the threshold between the ingroup and between-group comparisons (Figure 5A). In some cases, the observed maximum ingroup distances overlapped with the between-group distances of their closest relatives (see Supplementary Table S2 for genetic distances). For example, there was no distinct barcoding gap in P. melanurus (genetic differentiation between P. melanurus and its sister lineage Phrynocephalus sp. 13 comprised p = 2.62%, only slightly higher than the intraspecific distance within P. persicus, p = 2.52%). Another putative gap is observed at the level of 9.0% of substitutions in the COI gene, which corresponds to the comparisons between closely-related species complexes (Figure 5A).
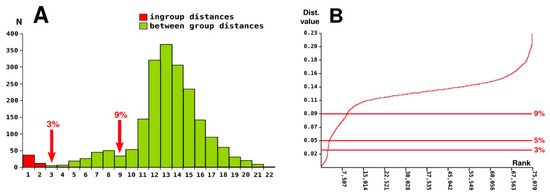
Figure 5.
Pairwise genetic divergence and species delimitation in Phrynocephalus: (A)—histogram showing the distribution of pairwise genetic divergences for COI sequences of Phrynocephalus; (B)—distribution of the ranked pairwise differences calculated with ABGD; the red line indicates the cumulative frequency of the distance values delimited with ABGD (uncorrected p-distances).
A similar picture is observed in the distribution of the ranked pairwise differences, with a sudden increase in slope in the vicinity of the 3.0% of substitutions threshold, and a less pronounced slope at the 9.0% level (Figure 5B). Overall, the only distinct gap in the distribution of pairwise genetic distances in Phrynocephalus was recorded at the level of 3.0% of substitutions.
3.4. Species Delimitation
The number of MOTUs recovered using the five phylogenetic species delimitation methods applied herein (ABGD, ASAP, bGMYC, mlPTP and hsPTP) varied significantly, but in all cases exceeded the species list based on the current taxonomy of the genus Phrynocephalus. The number of MOTU was estimated as 54 for ABGD, 55 or 63 for ASAP (two equally best partitions with equal ASAP coefficient = 5.0), 69 for bGMYC, 87 for mlPTP, and 103 for hsPTP.
As ASAP analysis resulted into two best partitions, we referred to resulting partitions as to ASAP1 (N = 55) and ASAP2 (N = 63) and used them as separate partitions in the subsequent calculation of Ctax and Rtax metrices. In addition, we used the p-distance levels of 3.0% and 5.0% as formal thresholds as separate partitions (following the results of barcode gap estimation presented above, and the standard species-level divergence threshold for the COI gene in reptiles following [44]).
The resolving power of hsPTP was maximal (Rtax = 0.94), the lowest power of resolution belonged to 5.0% p-distance (Rtax = 0.38), detecting only 38% of all species limits indicated by any other method. Other approaches suggested intermediate Rtax values: 0.49 for ABGD, 0.50 for ASAP1 and 0.52 for ASAP2, and a relatively high Rtax = 0.80 for mlPT.
The most congruent pair of species delimitation methods included ABGD and ASAP1 with Ctax = 0.98, i.e., 98% of all species limits inferred by the two methods are in agreement. The most incongruent pair is hsPTP and 5.0% p-distance with Ctax = 0.40, and Ctax values of the other pairwise comparisons range from 0.48 to 0.90.
ASAP1 and ASAP2 partitions were indicated as the most consensual species delimitation methods as compared to all other algorithms, with their mean Ctax values of 0.78. The lowest overall congruence was estimated for hsPTP (0.57).
4. Discussion
4.1. Comparative Performance of Different Approaches to Species Delimitation
Species delimitation is a quickly evolving way of describing biodiversity, and many recent studies apply innovative protocols and approaches for species delimitation [52,88,89]. Since COI-barcoding is presently the most common and widespread approach to species identification and biodiversity assessment [27,29], the development of single-locus delimitation methods are of special importance.
In the present paper, we combined five fast, single-locus distance-based and tree-based delimitation methods based on an almost complete data matrix of a taxonomically challenging genus of lizards. Our results generally confirm the high taxonomic utility of the COI-barcoding region in Phrynocephalus. Despite the recent significant progress in development of novel methods for more accurate species delimitations, no single method can be currently preferred [89]. Our study suggests that the credibility should be given to the approach which combines several alternative species delimitation methods simultaneously, and then objectively chooses the consensus species hypothesis based on their results. The combination of five species delimitation methods implemented in the present work suggested that results of the ASAP analysis generally most closely correspond to the current morphology-based classification of Phrynocephalus. ASAP resulted in two equally best partitions, recognizing 55 and 63 MOTUs (ASAP1 and ASAP2, respectively). Development of reliable DNA barcode libraries requires accurate identification of the revealed mtDNA lineages; therefore, below, we discuss the correspondence of the resulting partitioning schemes to the current Phrynocephalus taxonomy and demonstrate that diversity of the genus still likely remains underestimated. Future in-depth integrative studies including multilocus genetic analyses along with morphological and ecological data are required to clarify the cases, where the results of alternative species delimitation schemes differ. We hope that our analysis will be helpful for further investigations on Phrynocephalus diversity which eventually can improve our understanding of the taxonomy of the genus.
Though COI-barcoding is at present routinely used to delimit taxa including cryptic species, which are distinctly differentiated genetically but not morphologically, it should be acknowledged that as a single-locus method it has certain limitations. As phylogenetics widely acknowledges potential discordance among gene and species trees due to introgressive hybridization [90,91,92,93], or incomplete lineage sorting [94,95], species inferences based on a single locus alone can be misleading (reviewed by [96]). The most of the discordances between the current Phrynocephalus taxonomy and the MOTU lists representing putative ‘species’ resulted from an overestimation of species numbers. This suggests either unrecognized diversity and that the incomplete taxonomy of certain species groups, or reflects the oversplitting tendency of some of the species delimitation methods (especially that of PTP). For instance, the P. helioscopus complex was split into eight (ABGD), nine (ASAP), ten (ASAP, bGMYC), and 14–15 groups (mlPTP, hsPTP) (Figure 2, Figure 3 and Figure 4). The cases of possible species underestimation were few, where currently recognized as valid taxa were collapsed in single barcode units. This includes the case of parapatric P. versicolor from Mongolia and P. przewalskii from China: all methods but PTP lumped these species into a single entity, while ABGD and ASAP1 analyses suggested they are part of a larger entity including other members of the P. przewalskii complex (P. kulagini and P. frontalis) (Figure 4).
Our analyses revealed two cases of discordance between morphology-based classification and COI-barcoding-based species delimitation. The first case includes the P. przewalskii—P. versicolor—P. frontalis complex from Gobi, Qaidam, Alashan and Ordos deserts in southern Mongolia and central China. This complex includes several morphologically distinct morphotypes (see Figure 4) corresponding to different types of substrates (solid clay or gravel soils vs. sand dunes or fixed sands), known as “substrate races” [97]. The detailed morphological [98] and molecular [99,100] studies revealed discordant breaks in mtDNA and nuDNA markers along with clinal variation in morphometric characters in this complex. In our analyses, ABGD and ASAP1 recognized all members of the P. przewalskii—P. versicolor—P. frontalis complex, including P. kulagini from Tuva Republic in southern Siberia, as a single unit, while other methods identified P. kulagini and P. frontalis as independent “species”, while ASAP2 and bGMYC could not distinguish between haplotypes of P. versicolor from Mongolia and P. przewalskii from China (Figure 4). This result may be at least partially explained by historical vicariance which led to initial differentiation in mtDNA and morphology, with consequent hybridization and mtDNA introgression, which was reported earlier for this group of lizards [98,100]. Another case when COI-barcoding failed to recognize morphologically distinct taxa as independent MOTUs is the parapatric P. forsythii and P. nasatus, inhabiting Taklamakan Desert and the adjacent foothills of Tian Shan Mountains in western China. These two species are steadily distinguished in a number of diagnostic morphological characters [14,101], yet all species delimitation methods applied herein failed to recognize them as independent groups (Figure 2). Further studies, including examination of nuDNA-markers, are required to clarify evolutionary relationships and taxonomic status of these species.
Our results provide further evidence that the application of COI-barcoding along with alternative species delimitation methods provides a powerful tool to quickly test the initial species hypothesis in agamid lizards of the genus Phrynocephalus. However, a comparison of the performance of the five species delimitation approaches applied herein is not trivial. The number of MOTUs revealed by five different approaches together with the application of two formal genetic divergence thresholds (p-distance of 3.0% and 5.0%) varied from 42 to 103. No clear barcoding gap was revealed for Phrynocephalus at the level of 5.0% of substitutions in the COI gene, which was previously suggested as a formal indicator of species-level divergence in reptiles [44]. The application of the putative barcoding gap at the level of 3.0% of substitutions (Figure 2), results in detecting 57 MOTUs, which are highly congruent with the currently existing morphology-based taxonomy of Phrynocephalus. At the same time the formal 5.0%-threshold recognized only 42 species and significantly underestimates diversity in the P. arabicus, P. helioscopus, P. przewalskii and P. guttatus species complexes (Figure 2, Figure 3 and Figure 4). The ABGD and ASAP1-based delimitations proposed highly similar sets of MOTUs in Phrynocephalus (54 and 55 species, respectively). At the same time our results suggest that the delimitation schemes resulting from mlPTP and hsPTP demonstrate a clear tendency to oversplitting, recognizing almost twice more MOTUs (87 and 103 species, respectively) than ABGD, ASAP1, or the 5.0%-threshold. As shown in a few theoretical and empirical studies, it is likely that PTP-based species delimitation methods may reflect population splits rather than species divergences, thus often leading to oversplitting [89,102,103,104].
Herein we provided a generalized assessment of diversity in the genus Phrynocephalus, applying different species delimitation approaches to a single-locus COI-barcoding dataset. Our results suggest that ASAP provides the most consensual delimitation scheme, which also most closely reflects the morphology-based taxonomy of Phrynocephalus. Integrating various lines of evidence including mtDNA and nuDNA-markers, morphology, ecology and distribution is required to for a more accurate estimation of species borders in Phrynocephalus. In further studies, our species delimitation scheme for Phrynocephalus should be tested using the application of more informative genomic data, and an integrative taxonomic analysis.
4.2. An Overview of the Phylogenetic Lineages and Taxonomic Implications
Barabanov and Ananjeva [1] reviewed the available species names proposed for the genus Phrynocephalus, revised the taxonomy of the group and recognized 37 valid species within the genus based exclusively on morphological data. Solovyeva et al. [12] and Solovyeva et al. [2] provided a novel molecular phylogenetic hypothesis for Phrynocephalus, delimited subgenera and species groups within the genus. However, the sampling used in the two latter studies was limited and did not allow the estimation of species-level diversity within Phrynocephalus. Macey et al. [4] reported on an updated phylogeny for Phrynocephalus based on mtDNA, nuDNA sequences and allozyme data, which generally supported the results of Solovyeva et al. [2]. Below we discuss the taxonomic implications from our assessment of Phrynocephalus species diversity and revise the taxonomic changes proposed by Macey et al. [4].
Several recent studies [1,12,13,15,20,21] widely applied the subspecies category to the taxonomy of Phrynocephalus. Though there has been a certain skepticism regarding the usage of subspecies in herpetological taxonomy in the past [105,106,107], recently the category of subspecies has been getting more popular in scope of wider application of phylogenomic data allowing a reveal of cases of mito-nuclear discordance due to ongoing or ancient hybridization [108,109,110,111]. Marshall et al. [111] define the subspecies as a geographically circumscribed lineage that may have been temporarily isolated in the past, but which has since merged over broad zones of intergradation that show no evidence of reproductive isolation. Following the existing tradition in Phyrnocephalus systematics, our several prior taxonomic studies on the group have largely applied the subspecies category in describing geographically isolated, genetically and morphologically distinct populations [11,12,19,20,21,97,101,112]. For example, Solovyeva et al. [19,20] revised diversity and taxonomy of the P. helioscopus—P. persicus complex, recognizing two polytypic species P. helioscopus and P. persicus with seven and three subspecies, respectively. However, further molecular studies have demonstrated deep phylogenetic differentiation among the subspecies in this group with the divergence levels often exceeding those of Phrynocephalus taxa traditionally recognized as “good” species [2,4,17]. Several subsequent works raised some of the proposed subspecies to species rank, however not revising taxonomy of the complex as a whole [4,113]. Further integrative taxonomic studies along with the genomic-level data elucidating the existence of gene flow among the lineages are required to evaluate the taxonomic rank of the geographically circumscribed lineages within the wide-ranged species complexes of Phrynocephalus.
Solovyeva et al. [2,19] recognized ten major species groups within the genus Phrynocephalus: (1) Microphrynocephalus (encompassing the P. interscapularis group), (2) Phrynosaurus (encompassing the P. scutellatus group), (3) Oreosaura (encompassing the P. vlangalii group), (4) Megalochilus (encompassing the P. mystaceus group), (5) P. maculatus group, (6) Helioscopus (encompassing the P. helioscopus group), (7) P. raddei group, (8) P. guttatus group, (9) P. przewalskii group (as P. versicolor group), and (10) an orphaned species P. axillaris. Our analysis of the COI data strongly supported the monophyly of six of these species groups (Oreosaura, Megalochilus, Phrynosaurus, P. maculatus group, P. przewalskii group and P. axillaris), while three species groups received moderate support (Microphrynocephalus, Helioscopus, and guttatus group). Phrynocephalus lutensis, which was not included in the analysis of Solovyeva et al. [2,19], with high support represents a sister lineage to all other Phrynocephalus species, supporting the results of Ebrahimipour et al. [87]. However, the P. raddei group received no significant support and the phylogenetic position of P. strauchi remained essentially unresolved; nonetheless we assign this species to the P. raddei group based on the earlier results of Solovyeva et al. [2] and Macey et al. [4]. Below we provide a critical review of the phylogenetic lineages and diversity of Phrynocephalus. The following taxonomy is based on the results of ASAP2 (63 MOTUs) and morphological data published in earlier studies (see below).
4.2.1. Phrynocephalus lutensis
The large-sized and brightly-colored species P. lutensis was recently described from Dasht-e Lut Desert in eastern Iran by Kamali and Anderson [8]. This unique and insufficiently studied species occupies the most basal position in the genus Phrynocephalus, agreeing with earlier results [87] (Figure 2A). Phrynocephalus lutensis also appears to be profoundly different from other congeners in morphology [8], therefore the taxonomic status of this ancient lineage requires further investigation.
Included taxa: one species, P. lutensis Kamali and Anderson.
4.2.2. Microphrynocephalus (Phrynocephalus interscapularis Species Group)
The P. interscapularis species group includes the smallest members of the genus Phrynocephalus, all of which live on loose sands. It was coined as a subgenus Microphrynocephalus by Solovyeva et al. [12]. In our study, this species group includes four MOTUs: three lineages of the P. interscapularis species complex from Uzbekistan and Tajikistan, and P. vindumi from eastern Iran (Figure 2D). Macey et al. [4] obtained a more extensive sampling from eastern Middle East, their analysis included three species from Afghanistan and Pakistan, which were absent in our study: P. clarkorum, P. ornatus, and P. luteoguttatus; in their analyses these species form a clade with sister relationships to the P. interscapularis complex + P. vindumi. Macey et al. [4] also demonstrated that P. ornatus sensu lato is not monophyletic and elevated the subspecies P. ornatus vindumi to a full species status, the taxonomy followed herein. Macey et al. [4] included in their analysis two populations of the P. interscapularis complex, which were indicated as two subspecies (P. i. interscapularis and P. i. sogdianus); they reported on a p = 3.2% genetic divergence between these lineages, along with one fixed allozyme difference in EST locus. On the basis of these differences, Macey et al. [4] proposed a full species status for P. sogdianus. However, the sample used by Macey et al. [4] as P. sogdianus (CAS 182988) comes from Surkhondaryo Prov. of Uzbekistan, and not from the type locality of this taxon in environs of Pyandzh Village in southwestern Tajikistan [114]. Therefore, the conclusion on the full species status of P. sogdianus by Macey et al. [4] is insufficiently justified. In our study we examined a more extensive sampling on the P. interscapularis complex which included several localities for P. interscapularis sensu stricto from Uzbekistan (Bukhara, Termez, Qoraqalpog’iston, and Navoi; type locality—Bukhara, Uzbekistan [1,115], haplotypes it1–it4; see Supplementary Table S1), and two localities for P. sogdianus from Uzbekistan (Surkhondaryo) and southwestern Tajikistan (haplotypes so1–so2; see Supplementary Table S1). According to our data, P. interscapularis is recovered as paraphyletic with respect to P. sogdianus with the population from the Lake Aidarkul (central Uzbekistan; haplotypes sp5.1–sp5.2; see Supplementary Table S1) forming a highly divergent lineage with sister relationships to the clade joining P. interscapularis sensu stricto + P. sogdianus. The divergence between P. interscapularis sensu stricto and P. sogdianus is significant (p = 3.8% in COI gene) and was estimated to take place ca. 2.6 mya [2]; most species delimitation algorithms recognize these lineages as separate entities (see Figure 2). Therefore, our data agree with Macey et al. [4] on recognizing P. sogdianus as a distinct species. At the same time, the population from the Lake Aidarkul in Uzbekistan we herein tentatively indicate as a candidate species Phrynocephalus sp. 3; its taxonomic status should be further verified through an integrative taxonomic analysis.
Included taxa: seven species, including P. clarkorum Anderson and Leviton; P. interscapularis Lichtenstein; P. luteoguttatus Boulenger; P. ornatus Boulenger; P. sogdianus Chernov; P. vindumi Golubev; and Phrynocephalus sp. 3.
4.2.3. Phrynosaurus (Phrynocephalus scutellatus Species Group)
According to Solovyeva et al. [2], the subgenus Phrynosaurus encompasses the single species P. scutellatus, inhabiting mountainous plateaus of central and eastern Iran. Both Solovyeva et al. [12] and Solovyeva et al. [2] included a single specimen of P. scutellatus in their analyses; while Macey et al. [4] included samples from two populations in Kerman and Khorasan provinces of Iran; therefore, these studies failed to analyze differentiation within this group. A recent study by Rahimian et al. [116] examined morphological and molecular variation across the P. scutellatus range and recognized this taxon as a species complex, including the four major species-level clades from southeastern Iran (clade I), mountains of central Iran (clade II), northeastern Iran (clade IV), and a narrow area in borderlands of northeastern Iran and Afghanistan (clade III). However, Rahimian et al. [116] refused taxonomic reassessment of the revealed lineages pending further data. Our study of a broad sampling of the P. scutellatus complex across Iran has confirmed the results of Rahimian et al. [116] in recognizing four major lineages within this group, all of which are suggested as distinct MOTUs according to the species delimitation analyses (Figure 2E). Genetic variation among these lineages is high with p-distances ranging from 5.5% to 12.0% for COI gene (Supplementary Table S2); they can be also distinguished by a number of diagnostic morphological characters (unpublished data). The lineage of P. scutellatus from Esfahan and Semnan provinces of central Iran (haplotypes sc1, sc2, see Supplementary Table S1; clade II of Rahimian et al. [116]) likely corresponds to P. scutellatus sensu stricto (type locality—Mt. Sophia, near Esfahan, Esfahan Province, Iran [1,117]). The lineage inhabiting the Khorasan Province of Iran (haplotype br, see Supplementary Table S1; clade IV of Rahimian et al. [116]) can be confidently assigned to the name P. brevipes Nikolsky, 1907 stat. nov.: Phrynocephalus olivieri var. brevipes Nikolsky, 1907 was originally described from the environs of Naim-Abad in western Khorasan (now in Mazanderan Province of Iran [1]). According to our and previously published data [116] this lineage is widely distributed in northeastern Iran; morphologically specimens of this lineage fit well the original description by Nikolsky [118]. Taxonomic affiliations of the two remaining lineages from the easternmost Khorasan and Sistan-e Baluchestan provinces of Iran remain unclear. Phrynocephalus from Khorasan Province (haplotype ti, see Supplementary Table S1) might correspond to P. tickelii Gray, 1845 originally described from western Afghanistan [1]. Another available name from this group is Phrynocephalus olivieri var. carinipes Nikolsky, 1907; it was originally described from “Pudesch-Kupa” (at present the area between the villages Toodeshk and Kuhpayeh in Isfahan Province of Iran, ca. N 32.71, E 52.56), and highly likely represents a junior synonym of P. scutellatus sensu stricto. We tentatively propose to use this name for Phrynocephalus from Sistan-e Baluchestan province (haplotype ol, see Supplementary Table S1. Further integrative studies along with examination of the type specimens and genetic materials from the respective type localities are required to fully stabilize the taxonomy of the P. scutellatus species complex.
Included taxa: four species, including P. brevipes Nikolsky; P. carinipes Nikolsky; P. scutellatus (Olivier); and P. tickelii Gray.
4.2.4. Phrynocephalus maculatus Species Group
This group inhabits deserts of the Near East from the Arabian Peninsula to Mesopotamia and desert areas of central and western Iran. Our analyses revealed seven distinct MOTUs, three of which were previously assigned to P. maculatus, and four to P. arabicus (Figure 3F). Solovyeva et al. [2,12] demonstrated that P. maculatus sensu lato is paraphyletic with respect to P. arabicus, with P. m. maculatus sensu stricto from central Iran forming a sister lineage to the clade joining P. arabicus and P. m. longicaudatus from the Arabian Peninsula, and therefore elevated P. longicaudatus to the full species status. Macey et al. [4] also reported on the similar topology of phylogenetic relationships in the group and proposed the identical taxonomic changes, however without reference on the works of Solovyeva et al. [2,12]. The split between P. maculatus and the P. longicaudatus + P. arabicus complex was estimated as 4.8 mya [2]. Our results agree with the recent data of Ebrahimipour et al. [87] in recognizing the P. maculatus population from Abarkouh as a distinct MOTU. ASAP2, PTP, bGMYC, and the 3.0%-substitution threshold recognize this lineage as a distinct entity (referred below as Phrynocephalus sp. 4), while ASAP1, ABGD and the formal 5.0%-substitution threshold do not differentiate it from the remaining populations of P. maculatus. Additional investigations are needed to clarify the status of the Abarkouh population.
Macey et al. [4] examined two populations of P. arabicus from Saudi Arabia and Oman, which they identified as P. arabicus 1 and P. arabicus 2 in their study. However, Macey et al. [4] have overlooked the recent significant progress in taxonomy of the P. arabicus species complex made by Melnikov et al. [17,18], who described two new species from Iran (P. ahvazicus) and Oman (P. sakoi), and resurrected P. nejdensis from Saudi Arabia. Our analyses fully agree with the results of Melnikov et al. [17,18] and generally confirm the full-species status of P. arabicus sensu stricto, P. ahvazicus, P. nejdensis, and P. sakoi. However, the genetic distance between P. ahvazicus and P. arabicus is lower than the 3.0% threshold (p = 2.5%, see Supplementary Table S2); morphological characters distinguishing these two taxa are also quite vague [17]. Further multilocus studies are required to clarify the phylogenetic relationships between P. ahvazicus and P. arabicus and their taxonomic status.
Included taxa: seven species, including P. ahvazicus Melnikov, Melnikova, Nazarov, Rajabizadeh, Al-Johany, Amr and Ananjeva; P. arabicus Anderson; P. longicaudatus Haas; P. maculatus Anderson; P. nejdensis Haas; P. sakoi Melnikov, Melnikova, Nazarov, Al-Johany and Ananjeva; and Phrynocephalus sp. 4.
4.2.5. Oreosaura (Phrynocephalus vlangalii Species Group)
The subgenus Oreosaura was established by Barabanov and Ananjeva [1] to encompass the P. vlangalii species group, joining viviparous species inhabiting high elevation deserts of the Qinghai-Tibet Plateau and the Taklamakan Desert in Central Asia. Our analyses distinguished ten distinct MOTUs within this subgenus, which comprise two major groups (Figure 2B). The first group joins the high-elevation species, and the second group includes P. forsythii and P. nasatus, which inhabit lower elevations [14]. The samples of P. forsythii from Taklamakan Desert of China and P. nasatus from the adjacent Tian Shan Mountains showed very shallow differentiation in COI sequences (p = 1.0%, see Supplementary Table S2), with the two taxa recovered as paraphyletic and all analyses recognizing these taxa as a single unit. This brings into question the species status of P. nasatus. This species was originally described from historical collections by Golubev and Dunayev [101], and later was considered as a junior synonym of P. axillaris by Barabanov and Ananjeva [1]; however, the latter authors did not provide any justification for this decision. Recently P. nasatus was rediscovered and redescribed by Dunayev [14], who reported on its parapatric distribution with P. forsythii, and outlined the morphological differences of P. nasatus from other congeners. Our work demonstrates that P. nasatus is closely related to P. forsythii; further multilocus study in progress will likely clarify the status and relationships between these two taxa.
Prynocephalus vlangalii species complex distributed in Qinghai Plateau including Qaidam Desert in our analyses formed three major monophyletic clades with shallow divergence (p = 1.3%, see Supplementary Table S2), of which one specimen (MF039061, from Nanshan Mountains) was separated as a distinct MOTU by PTP and ASAP2 analyses. Following Jin and Brown [86], this population corresponds to P. vlangalii var. nanschanica Bedriaga, 1906; based on the results of species delimitation we herein tentatively refer to it as to P. nanschanicus stat. nov. An earlier study on phylogeography of P. vlangalli revealed three major lineages within this species: a lineage from the headwaters of the Yellow River, a lineage from Qaidam Basin, and a lineage from Suganhu area [119]. Jin et al. [120] and Jin and Brown [86] recognized three subspecies within P. vlangalii complex: P. v. vlangalii, P. v. pylzowi, and P. v. nanschanica (the latter taxon is treated as P. nanschanicus in the present work; the gender of the species name is herein adjusted to fit the masculine gender of Phrynocephalus). Our analyses also confirmed that the populations of P. v. vlangalii and P. v. pylzowi form two reciprocally monophyletic clades, which, however were not recognized as distinct MOTUs only by mlPTP and hsPTP and we treat them here as subspecies (Figure 2B). The most recent phylogenetic study on this complex based on both mitochondrial and nuclear markers [121] revealed four major intraspecific lineages within P. vlangalli. It is difficult to evaluate the correspondence of these lineages to the COI-haplotypes reported in our study without additional studies.
The populations of P. putjatai from the environs of Qinghai Lake form a strongly divergent clade sister to all remaining lineages of highland species of Oreosaura (p > 6.0%, see Supplementary Table S2). Jin and Brown [86] demonstrated that morphologically different P. guinanensis is genetically indistinguishable from P. putjatai (spelled as “P. putjatia” in their work; however “P. putjatai” is the correct spelling according to Barabanov and Ananjeva [1]). Our study agrees with the results of Jin and Brown [86,122] and Jin et al. [123] on the phylogenetic position of P. guinanensis (Figure 2B). The recent multilocus phylogenetic study by Chen et al. [121] reported on three major lineages within P. putjatai. Further studies are needed to clarify the extent of distribution and taxonomic status of these lineages.
Within the populations from Xizang (Tibet) and the Himalaya, our analyses reveal six MOTUs. The samples from Ladakh in Jammu and Kashmir State of India correspond to P. theobaldi (type locality—Tso Morari, Ladakh, Jammu and Kashmir, India, see [124]). One sample of P. cf. theobaldi (MF039063 from Ngari Prefecture of Xizang) is deeply divergent from the topotype specimens of P. theobaldi in COI sequences (p = 4.9%, see Supplementary Table S2). This population was regarded as P. t. theobaldi by Jin and Brown [86] and Jin et al. [125], however these authors lacked topotypic materials on P. theobaldi from Ladakh; our study demonstrated that the Ngari population is likely not conspecific to P. theobaldi sensu stricto, and we tentatively identify it as Phrynocephalus sp. 1 pending further studies. The samples of Phrynocephalus from central Xizang, China, fall into two separate MOTUs. One lineage we provisionally identify as P. lhasaensis (referred to as “P. t. orientalis 2” by Jin and Brown [86], however this name is not available due to homonymy with Phrynocephalus helioscopus orientalis Bedriaga, 1912 according to Barabanov and Ananjeva [1]). The second lineage in this complex (KF691720 from Pelguzo Lake, and MF039062 from Brahmaputra River valley; both localities in Xizang) which was referred to as “P. t. orientalis 1” by Jin and Brown [86] we tentatively identify as Phrynocephalus sp. 2.
Prynocephalus erythrurus from northern Xizang (type locality in Sagus Kul, northwestern Tibet) includes two MOTUs, of which one likely corresponds to P. erythrurus sensu stricto (MF039065), while another one from Tuotuo River in Qinghai Province of China to P. parvus (KJ630904) (regarded as “P. e. parva” by Jin and Brown [86]; the gender of the species name is herein adjusted to fit the masculine gender of Phrynocephalus); this taxonomy is further corroborated by the earlier studies [121,126]. The lineages within P. erythrurus species complex are recognized as separate MOTUs by all species delimitation methods; therefore, we tentatively recognize them as distinct species.
Overall, our results are generally consistent with the phylogeny of Oreosaura proposed by Jin and Brown [86], Jin et al. [120], and Chen et al. [121]. In these studies, the authors achieved much better phylogenetic resolution for Oreosaura, however, their study lacked data on P. theobaldi from India. Additionally, Jin et al. [125] conducted species delimitation analysis for P. theobaldi based on two nuclear markers, and distinguished four clusters within this species; they may partially correspond to P. theobaldi and P. lhasaensis recognized herein. Macey et al. [4] included genetic information from five species-level lineages of Oreosaura in their analysis and did not discuss the status of other lineages revealed in earlier studies. In addition to our sampling, Macey et al. [4] examined P. roborowskii from Qaidam Desert, and P. vlangalii hongyuanensis from northwestern Sichuan, and argued that the latter taxon should be elevated to the species rank as P. hongyuanensis. However, their data are contradictory to the data presented by Jin et al. [120] who demonstrated P. roborowskii to be genetically indistinguishable from P. v. vlangalii. Moreover, Jin et al. [120] included in their analyses four localites of P. v. pylzowi including the population from Waqie, Hongyuan County (the type locality of P. hongyuanensis), and demonstrated that P. hongyuanensis is identical to P. v. pylzowi. Given the density of sampling presented by Jin et al. [120], we follow their taxonomy and do not recognize P. roborowskii and P. hongyuanensis as valid species.
At the same time, our sampling on Oreosaura was quite limited; therefore, all conclusions and species identifications presented herein should be taken cautiously. Despite the recent significant progress in phylogenetic studies on Oreosaura [86,119,120,121,122,123,124,125,126]; for many species of this clade COI-barcodes are still absent. In the present paper we provide our vision on the possible taxonomic assignment of the currently known COI-lineages of Oreosaura, and urge other researchers to provide a COI-barcode library for the Chinese species of Phrynocephalus in their future studies.
Included taxa: ten species, including P. forsythii Anderson; P. lhasaensis Barabanov, Ananjeva, Papenfuss and Wang; P. erythrurus Zugmayer; P. nanschanicus Bedriaga, “1905” 1906; P. parvus Bedriaga; P. putjatai Bedriaga; P. theobaldi Blyth; P. vlangallii Strauch; and Phrynocephalus sp. 1–2.
4.2.6. Megalochilus (Phrynocephalus mystaceus Species Group)
The subgenus Megalochilus was originally established for the largest species of Phrynocephalus—the psammophilous secret toadheaded agama, P. mystaceus [127]. Our analyses agree with the earlier data of Solovyeva et al. [2,21] in recognizing two deeply divergent reciprocally monophyletic lineages within Megalochilus with non-overlapping distributions (Figure 2C): P. mystaceus inhabiting Middle Asia, southern Russia, Kazakhstan and western Xinjiang of China, and the recently described P. m. khorasanus from deserts of northeastern Iran [26]. The divergence between these lineages in COI gene is significant (p = 7.2%, see Supplementary Table S2), it was dated as 2.1 mya [2]. The two MOTUs of Megalochilus are recognized as separate entities by all species delimitation analyses implemented herein (Figure 2). Given the morphological and genetic distinctiveness of these two lineages we propose to treat them as separate species and formally recognize the Iranian lineage as a full species P. khorasanus stat. nov. Within P. mystaceus sensu stricto, sequences of the currently recognized subspecies P. m. mystaceus, P. m. aurantiacocaudatus and P. m. galli are intermixed; further research is needed for a more detailed assessment of interspecific variation within P. mystaceus.
Included taxa: two species, including P. mystaceus (Pallas); and P. khorasanus Solovyeva, Dunayev, Nazarov, Radjabizadeh and Poyarkov.
4.2.7. Helioscopus (Phrynocephalus helioscopus Species Group)
The members of P. helioscopus species group inhabit vast areas from Caucasus to China and from southern Russia to Iran, preferring deserts with solid substrates (clay, gravel, or saline soils). The basal divergence of this group was dated as ca. 6.2 mya [2]. Until recently the group included only two species—P. helioscopus and P. persicus, presently both are recognized as species complexes [19,20,113]. In our analyses, the P. helioscopus species group has the largest number of putatively species-level lineages—13, with ten MOTUs in the P. helioscopus complex and six in the P. persicus complex; both complexes formed two reciprocally monophyletic groups with generally well-resolved phylogenetic relationships (Figure 3G). Solovyeva et al. [19] and Melnikov et al. [113] have revised the P. persicus species complex and recognized P. persicus from northern and central Iran and P. horvathi from southern Caucasus as distinct species. Additionally, Melnikov et al. [113] described the population from Fars Province in southern central Iran as a distinct species P. ananjevae. Our phylogeny fully agree with the results of Solovyeva et al. [19] and Melnikov et al. [113], and additionally report on three new previously unknown candidate species Phrynocephalus sp. 8 from Naxcivan, Azerbaijan (sister to P. horvathi; p = 5.6%), Phrynocephalus sp. 9 from environs of Ardebil in Iran (sister to P. persicus; p = 2.5%, see Supplementary Table S2), and Phrynocephalus sp. 10 from Apsheron Peninsula in Azerbaijan (4.6% to P. persicus and 8.4% to P. horvathi). All members of the P. persicus complex are suggested as distinct taxonomic entities according to species delimitation analyses (Figure 3), although ASAP1 unites Phrynocephalus sp. 9 with P. persicus. The taxonomic status of Phrynocephalus sp. 10 from Apsheron Peninsula is of special concern, since this population may be critically endangered or already extinct in the wild [128,129,130]. Further integrative taxonomic studies are needed to clarify the taxonomy of the P. persicus species complex.
Solovyeva et al. [19,20] revised phylogenetic relationships and taxonomy of the P. helioscopus species complex and recognized eight major lineages within it, which they treated as subspecies: P. h. helioscopus, P. h. varius, P. h. turcomanus, P. sergeevi, P. cameranoi, P. saidalievi, P. meridionalis, and an undescribed lineage from Karatau, southern Kazakhstan. Recently, Macey et al. [4] based on a limited sampling compared populations of the P. helioscopus complex from Aralsk, Kazakhstan (P. h. helioscopus) and from Turkmenbashi (Krasnovodsk), Turkmenistan (P. h. turcomanus), and suggested a full species status for turcomanus based on ‘significant mt-DNA differences’, despite the absence of fixed differences in allozymes [45] (p. 10). However, Macey et al. [4] ignored the phylogenetic results of Solovyeva et al. [19,20] and overlooked the fact that the elevation of P. h. turcomanus to full species would make P. helioscopus paraphyletic, as P. h. turcomanus is deeply nested within the radiation of other P. helioscopus subspecies (Figure 3). The radiation within the P. helioscopus complex likely started approximately 3.4 mya [2]; genetic distances among the lineages of the complex are high and vary from 2.6% to 10.3% (Supplementary Table S2). All species delimitation approaches agree in recognizing eight MOTUs of the P. helioscopus complex as independent entities, including Phrynocephalus from Karatau (here referred to as Phrynocephalus sp. 6). Additionally, nearly all species delimitation approaches supported two previously unknown lineages. One of them is Phrynocephalus sp. 5, which includes the populations originally assigned to P. cameranoi from the left bank of Ili River in eastern Kazakhstan. The second one is Phrynocephalus sp. 7, which represents a sister lineage to P. saidalievi with moderate node support. ASAP1 unites Phrynocephalus sp. 5 with P. cameranoi, though they are regarded as distinct entities based on the results of ASAP2. Phrynocephalus sp. 7 is recognized as a separate candidate species by all species delimitation methods, but not by the formal p-distance thresholds (Figure 3). Therefore, based on genetic [19] and morphological differentiation reviewed in [20], we propose to elevate the aforementioned lineages to species rank, and recognize them as P. helioscopus sensu stricto, P. varius, P. turcomanus, P. cameranoi stat. nov., P. sergeevi stat. nov., P. saidalievi, P. meridionalis stat. nov., and Phrynocephalus sp. 5–7. The taxonomic status of an undescribed candidate species Phrynocephalus sp. 6 from Karatau, southern Kazakhstan, is currently under examination by us.
Macey et al. [4] reported on phylogenetic placement of P. golubewii, originally described as a subspecies of P. maculatus, and demonstrated that based on mtDNA data the sample collected from the type locality of P. golubewii (Bami, Turkmenistan) actually belongs to the P. helioscopus species group and is reconstructed as a sister lineage to the clade including P. helioscopus and P. turcomanus. This result is quite surprising given the profound morphological differences between P. golubewii and the members of the P. helioscopus group. Moreover, in the analysis of a single nuclear DNA marker (RAG1) by Macey et al. [4] P. golubewii is clustered within the P. raddei species group as a sister species to P. rossikowi; this discrepancy was not discussed by Macey et al. [4]. The discordance between the mtDNA and nuDNA markers might result for various reasons, such as an incomplete lineage sorting or introgressive hybridization [96]; it was earlier reported for other members of Phrynocephalus [2]. Since P. golubewii occurs in sympatry with the members of the P. helioscopus complex, a possible misidentification during sample collection by Macey et al. [4] has also to be considered. Therefore, the phylogenetic placement of P. golubewii remains unclear and requires further investigation.
Included taxa: seventeen species, including P. ananjevae Melnikov, Melnikova, Nazarov and Rajabizadeh; P. cameranoi Bedriaga; P. golubewii Shenbrot and Semenov (?—requires confirmation), P. helioscopus (Pallas); P. horvathi Méhely; P. meridionalis Solovyeva, Dunayev and Poyarkov; P. persicus De Filippi; P. saidalievi Sattorov; P. sergeevi Solovyeva, Dunayev and Poyarkov; P. turcomanus Solovyeva, Dunayev and Poyarkov; P. varius Eichwald; and Phrynocephalus sp. 5–10.
4.2.8. Phrynocephalus raddei Species Group
This species group includes a number of gravel-dwelling taxa inhabiting the southern part of Middle Asia (the southernmost Kazakhstan, Turkmenistan, Uzbekistan and Tajikistan); the basal divergence within this group was dated as 4.6 mya according to Solovyeva et al. [2]. Our ABGD and bGMYC analyses recognize six distinct MOTUs within the P. raddei group (Figure 4H). Macey et al. [4] assigned P. strauchi to this species group though based on a poorly supported topology. The multilocus analysis by Solovyeva et al. [2] demonstrated that P. strauchi represents a sister lineage to the members of the P. raddei group based on analysis of mtDNA markers, but represents a distinct lineage with unresolved phylogenetic placement according to the analysis of nuDNA genes. In the present paper P. strauchi forms a distinct lineage with unresolved phylogenetic position (Figure 4). Based on overall morphological similarity and the mtDNA phylogeny of Solovyeva et al. [2] and Macey et al. [4], we assign P. strauchi to the P. raddei group, although further investigations on the phylogenetic placement of this species are desirable.
Macey et al. [4] in their study included three members of the P. raddei group: P. rossikowi, P. bannikovi and P. raddei; moreover P. golubewii was suggested as a member of this species group based on their analysis of RAG1 nuclear DNA gene. Macey et al. [4] also stated that “P. reticulatus from central Uzbekistan was not sampled <…> in the study of Solovyeva et al. (2014)” [4] (p. 35). However, though Macey et al. [4] referred to the work by Golubev [10] on the taxonomy of this group, they failed to report that in this paper the author has demonstrated that P. reticulatus Eichwald represents a subjective junior synonym of P. ocellatus (Lichtenstein) (described from Bukhara, Uzbekistan), and resurrected the latter nomen as a valid species name [10]. Subsequently, this taxonomy was widely accepted for over 25 years [2,11,12,101,112,130,131,132,133]. We also follow this taxonomy in the present paper, and would like to underline that, in opposite to the unjustified statement of Macey et al. [4], P. ocellatus was included in the phylogenetic studies by Solovyeva et al. [2,12], where it was recovered as a member of the P. raddei species group. Additionally, Macey et al. [4] demonstrated that P. bannikovi is a sister species of P. rossikowi, and suggested elevation of the former taxon to a full species rank. We did not sample P. bannikovi in the present study and follow the taxonomy proposed by Macey et al. [4] in recognizing P. bannkovi as a distinct species and as a member of the P. raddei species group.
Our analyses indicated that the present taxonomy of the P. raddei species group is largely incomplete. First, P. raddei was recovered as paraphyletic with respect to P. ocellatus. Presently P. raddei is considered to include two subspecies: P. raddei raddei Boettger (inhabiting southwestern Turkmenistan), and P. raddei boettgeri Bedriaga (from southern Uzbekistan and Tajikistan). In our study, the sample of P. raddei boettgeri is clustered within the P. ocellatus species complex with significant support (1.0/99), while the position of P. raddei raddei and P. rossikowi remains unresolved and the monophyly of the P. ocellatus species complex is essentially not supported (Figure 4H). Phrynocephalus raddei raddei (together with P. bannikovi, which was not included in our analyses) was recovered as a sister lineage to P. rossikowi by Macey et al. [4]. Genetic divergence between P. raddei raddei and P. raddei boettgeri is also high (p = 8.3%, see Supplementary Table S2). Therefore, due to the non-monophyly of P. raddei sensu lato and the significant molecular and morphological divergence, we herein propose to elevate the Uzbekistani taxon to a full species rank as P. boettgeri stat. nov.
Furthermore, P. ocellatus in our analyses is recovered as paraphyletic with respect to P. boettgeri and is represented by two well-supported and genetically divergent lineages (p = 6.2%, see Supplementary Table S2). The lineage from southern and western Uzbekistan corresponds to P. ocellatus sensu stricto (haplotypes oc1–oc3; Supplementary Table S1); the lineage from northern central Uzbekistan (Uchkuduk and Mingbulak) was not reported in earlier studies and is molecularly and morphologically distant from all other members of the P. raddei species group (see Supplementary Table S1). The taxonomic reassessment of the Uchkuduk and Mingbulak lineage is in progress; herein we tentatively identify this lineage as Phrynocephalus sp. 11.
Included taxa: seven species, including P. bannikovi Darevsky, Rustamov and Shammakov; P. boettgeri Bedriaga; P. ocellatus (Lichtenstein); P. raddei Boettger; P. rossikowi Nikolsky; P. strauchi Nikolsky; and Phrynocephalus sp. 11.
4.2.9. Phrynocephalus axillaris
According to Solovyeva et al. [2], the phylogenetic placement of P. axillaris from Taklimakan Desert of China is inconsistent in the analyses of mtDNA and nuDNA data. Based on mtDNA markers this species was suggested as a sister lineage to P. mystaceus though with low topological support [2,4]; while the nuDNA markers confidently placed P. axillaris as a sister lineage to the clade joining the P. guttatus and P. przewalskii species groups [2]. In our analyses of COI mtDNA gene the phylogenetic position of P. axillaris is unresolved, and no geographic lineages were revealed within this species (Figure 4I).
Included taxa: one species, P. axillaris Blanford.
4.2.10. Phrynocephalus guttatus Species Group
In our analyses, the P. guttatus species group included seven distinct MOTUs (Figure 4K). These lineages are distributed from southern European Russia across Kazakhstan and northern Middle Asia through to westernmost China. The divergence within the P. guttatus group started ca. 2.5 mya; the phylogenetic relationships within this group reported in our study correspond well to the earlier results of Solovyeva et al. [2] and Dunayev et al. [13]. Western Kazakhstan, southern Russia and the northernmost Turkmenistan are inhabited with P. guttatus sensu stricto, the Aral Sea region is occupied by P. moltschanovi, while P. alpheraki occurs in the Ili River valley in eastern Kazakhstan and Yining District of Xinjiang Province of China. These three taxa are identified as separate entities by all species delimitation approaches tested in our study (Figure 4K), and together they comprise the P. guttatus species complex. Sequences of the subspecies P. guttatus salsatus and P. guttatus kalmykus were placed among other samples of P. guttatus sensu stricto and show only a shallow genetic differentiation (p < 0.45%; see Supplementary Table S2).
Two species of the P. guttatus species group inhabit the Balkhash Lake region in eastern Kazakhstan: P. kuschakewitschii and P. incertus. These species with parapatric distribution [13] form a poorly-supported clade (0.95/85) and represent two reciprocally monophyletic groups with genetic distance of p = 4.8% (Supplementary Table S2). Phrynocephalus albolineatus Zhao, described from Tacheng County in the westernmost Xinjiang of China, clusters within P. kuschakewitschii and represents a subjective junior synonym of this species, as it was demonstrated earlier [13]. Phrynocephalus kuschakewitschii and P. incertus are supported as separate units by most species delimitation methods (Figure 4), supporting the full species status first suggested by Melville et al. [83].
The desert areas of Junggar Depression in northwestern China and Zaysan and Alakol depressions in eastern Kazakhstan are occupied by members of a distinct clade of the P. guttatus species group, consisting of two lineages. Macey et al. [4] applied the name P. salenskyi Bedriaga, 1907 to the members of this clade, stating that it is “the oldest name available for Phrynocephalus populations of the low elevation Chinese desert—northern Caspian Basin” [4] (p. 35). At the same time, some other authors, e.g., Jin and Brown [86], applied the name P. grumgrzimailoi Bedriaga, 1909 to this taxon. However, these taxonomies are both misleading, since they have obviously ignored the older synonym P. melanurus Eichwald, 1831, which was described from Zaysan Depression and resurrected by Semenov et al. [134]. After its revalidation, this nomen was widely applied for the members of P. guttatus species group from the easternmost Kazakhstan either for a subspecies [1] or a full species [2,12,135,136,137]. Synonymy of P. grumgrzimailoi with P. melanurus was also suggested by Barabanov and Ananjeva [1].
Taxonomy of the P. guttatus species group is rather complicated due to the high morphological variation among the populations adapted to various substrate types (so-called “substrate races” [97]); as a result, numerous taxa were described in this group during the last 150 years of studies [1]. Dunayev et al. [13] have recently reviewed the distribution and molecular differentiation of the lineages within the P. guttatus species group and demonstrated that P. melanurus comprises two major lineages. The first one occurs in Zaysan Depression and likely is found further in the northwestern part of Junggar Depression in Xinjiang Province of China; it corresponds to P. melanurus sensu stricto. The second lineage penetrates to the Alakol Depression in Eastern Kazakhstan from the southwestern part of Junggar Depression. In Solovyeva et al. [2] these lineages were referred to as ‘P. melanurus 1′ and ‘P. melanurus 2′, respectively. We also recover these two lineages in our analyses; they form two reciprocally monophyletic groups with genetic divergence of 2.6% (Supplementary Table S2), and are regarded as separate entities by ASAP2m bGMYC, mlPTP and hsPTP species delimitation analyses, but not by the ABGD and ASAP1 (Figure 4). The ranges of these two lineages seem to be separated by the Tarbagatai Mountains, however the actual extent of their distribution eastwards to the Junggar Depression in China remains unknown [13]. Though there is no doubt that the name P. melanurus is applicable to the lineage from Zaysan Depression and the northern part of Junggar Depression, while P. salenskyi and P. grumgrzimailoi represent its junior synonyms, the taxonomic status of the southern lineage from the Alakol Depression is less clear. Given the significant morphological differences between the specimens from Alakol and the topotype P. melanurus, along with genetic divergence and the results of species delimitation analyses, we herein tentatively indicate this lineage as Phrynocephalus sp. 13. Among the existing nomens, P. isseli Bedriaga (type locality in Saur Mountains, Xinjiang, China) or P. arcellazzii Bedriaga (type locality in Qitai, Xinjiang, China) might represent the available names for this lineage [1]. Further integrative taxonomic assessment along with intensified sampling effort in northwestern China and a careful examination of the name-bearing types of the group are needed to fully stabilize the taxonomy of the P. guttatus species group.
Included taxa: seven species, including P. alpheraki Bedriaga in Nikolsky; P. guttatus (Gmelin); P. incertus Bedriaga in Nikolsky; P. kuschakewitschii Bedriaga in Nikolsky; P. melanurus Eichwald; P. moltschanovi Nikolsky; and Phrynocephalus sp. 13.
4.2.11. Phrynocephalus przewalskii Species Group
In our analyses, the P. przewalskii species group included five distinct MOTUs distributed from southern Siberia (Tuva Republic) across Mongolia to central China (Figure 4J). Taxonomy of this complex has undergone significant changes after the works of Wang and Fu [138], Urquhart et al. [99,100], Jin and Brown [139], and Gozdzik and Fu [98], who overall demonstrated that the populations of P. przewalski and P. frontalis, occurring in Ordos and Alashan deserts of Central China, do not represent separate species due to the clinal variation of morphological characters and a significant amount of gene flow between the populations. However, the abovementioned studies only included samples from China, but not from the rest of the groups’ vast range, including Mongolia and Russia. In agreement with Solovyeva et al. [2] and Dunayev et al. [15], our analyses revealed two major clades within the P. przewalskii species group, corresponding to the populations from the western part of its range (from Junggar Depression and Govi-Altai aimag of Mongolia to Gansu Province of China), and from the eastern and northern parts of its distribution (from central China across Mongolia to Tuva Republic). The western clade corresponds to P. hispidus, recently revalidated by Solovyeva et al. [2]; morphological differences of this species from other members of the P. przewalskii species group were detailed by Dunayev et al. [15]. Additionally, a sample from Gansu Province of China formed a sister lineage to P. hispidus from Mongolia with significant genetic distance among them (p = 3.6%, see Supplementary Table S2); they are recognized as separate units by all species delimitation algorithms but formal 5.0% p-distance threshold (Figure 4). We herein tentatively identify the Gansu population as Phrynocephalus sp. 12 pending further morphological and genetic data. Remarkably, the population from Anxi, Jiuquan Prefecture in Gansu Province of China (CAS:Herp:170914–23) originally reported as ‘P. versicolor’ by Macey et al. [4] actually refers to Phrynocephalus sp. 12; this misidentification explains the discrepant results on the phylogenetic placement of P. versicolor and P. przewalskii in Macey et al. [4] (see below).
The phylogenetic relationships within the northeastern clade of the P. przewalskii species group generally agree with the results reported in Dunayev et al. [15]. Recently Solovyeva et al. [2] demonstrated significant divergence in mtDNA and nuDNA sequences between the subspecies P. versicolor versicolor from Mongolia and P. versicolor kulagini from Tuva Republic, and elevated the latter to a full species as P. kulagini. Morphological differentiation between these species was subsequently reported by Dunayev et al. [15]. In our analyses, P. kulagini is recovered as an independent unit by all species delimitation algorithms but ABGD, ASAP1 and the formal 5.0% p-distance threshold (Figure 4); it demonstrates 3.7–4.0% of sequence divergence from its sister species (Supplementary Table S2). The samples of P. przewalskii from Alashan Desert in China were grouped in one clade with P. versicolor from Mongolia, with a very shallow differentiation among them (p = 1.0%, see Supplementary Table S2); only PTP analyses recognized them as separate entities (Figure 4). The samples of P. frontalis from Ordos Desert in China were slightly divergent from P. przewalskii + P. versicolor (p = 3.2%, see Supplementary Table S2), and bGMYC, ASAP2, mlPTP, hsPTP analyses along with 3.0% p-distance barcoding gap recognized them as separate entities (Figure 4). Considering the previously published data [98,99,100] which demonstrated the existence of gene flow among the Chinese populations of P. przewalskii and P. frontalis, the taxonomy of this species complex remains obscure and cannot be resolved based on the mtDNA data alone. As P. przewalskii was reported from southern Gobi Desert in Inner Mongolia of China [98,99,100], it is not surprising that its distribution continues northwards to central Mongolia, where this lineage is known under the name P. versicolor. All three names available for this complex (P. przewalskii, P. versicolor, and P. frontalis) were proposed by Strauch [140] in the same publication. As Wang and Fu [138] were the first to review this question and have chosen to give priority to P. przewalskii over P. versicolor and P. frontalis, in case these three taxa would be eventually lumped in a single species, the name P. przewalskii should be used for it. Further integrative and multilocus studies on a denser sampling from both China and Mongolia are needed to clarify the taxonomy of the P. przewalskii—P. versicolor species complex.
Included taxa: five species, including P. frontalis Strauch; P. hispidus Bedriaga; P. kulagini Bedriaga; P. przewalskii Strauch (including P. versicolor Strauch); and Phrynocephalus sp. 12.
4.2.12. Phrynocephalus Incertae Sedis
Due to the significant recent progress in molecular studies on the genus Phrynocephalus, there is only one currently recognized species left which lacks phylogenetic information. This is the enigmatic P. euptilopus described from Afghanistan; from its characteristic morphology and large body size this species may be phylogenetically related to P. mystaceus [141].
Included taxa: one species, P. euptilopus Alcock and Finn.
Overall, our analyses indicate that despite the progress made for over two centuries of taxonomic studies of Phrynocephalus, the species-level diversity of the genus likely remains largely underestimated. The review by Barabanov and Ananjeva [1] recognized 37 valid species in the genus Phrynocephalus, however their taxonomy has not been followed consistently, e.g., the Reptile Database currently recognizes just 34 valid species in this genus [9]. Our species delimitation analyses have revealed at least 49 lineages representing nominal species of Phrynocephalus. We also reported on 13 putative candidate species, which may either correspond to yet undescribed taxa or to the available names currently recognized as synonyms. Together with eight species of Phrynocephalus not included in our study, but confirmed as distinct species by earlier works [4,141], the total number of nominal species in Phrynocephalus comprises 56, and along with 13 candidate species reported in the present study reaches 71. We also demonstrate that the ASAP species delimitation analysis provides the scheme most closely reflecting the currently recognized diversity of Phrynocephalus (with 55–63 MOTUs recovered by this method); the ABGD analysis provided highly concordant species delimitation scheme (with 54 MOTUs). At the same time the bGMYC, the mlPTP, and the hsPTP analyses obviously overestimated the diversity of Phrynocephalus (recognizing 69, 87, and 103 MOTUs, respectively). The formal threshold of 5.0% of substitutions in COI gene results in 42 MOTUs, thus slightly underestimating the currently recognized diversity of Phrynocephalus. At the same time the application of the threshold of 3.0% of substitutions resulted in 57 MOTUs, which closely approaches the ASAP species delimitation results.
Taxonomic studies on Phrynocephalus are challenging, in particular due to the complicated nomenclature history with over 160 species-level names proposed for the members of the genus [1,9]; for most taxa, the detailed descriptions of the type specimens are lacking. At the same time, the high morphological and ecological plasticity of many Phrynocephalus species which evolved different morphotypes on various types of substrates [97] often complicates species diagnostics from the morphological characters alone. Finally, the likely relatively widespread interspecific hybridization leading to introgression and the discordance in phylogenetic signals from mitochondrial and nuclear genomes further hampers species delimitation in Phrynocephalus. Remarkably, the intermixture between different lineages of toad-headed agamas is not only reported among the recently diverged closely related taxa [97,98,99,100], but also likely happened during the early stages of Phrynocephalus evolution [2]. Overall, this makes the species delimitation based exclusively on mtDNA markers in this group unreliable, and only an integrative taxonomic approach joining data from morphology, mtDNA and nuDNA markers, and natural history can provide exhaustive evidence for a species status of Phrynocephalus lineages. Therefore, we recognize the taxonomic implications discussed above as hypothetical; further integrative studies are needed to test the hypotheses put in the present paper.
Given the comparatively little number of recent phylogenetic and taxonomic studies on Phrynocephalus as compared to the significant number of works on these lizards published in the end of XIX–early XX centuries, the further progress on taxonomy of the genus can be achieved only through a careful analysis of the available literature, museum collections, and the published data. Macey et al. [4] provided an updated phylogenetic hypothesis for the genus Phrynocephalus which resulted in a number of taxonomic recommendations. However, as demonstrated above, in many cases these authors have overlooked a number of existing recent and old publications on Phrynocephalus, and as a result the taxonomy of the genus they proposed is largely misleading. For example, Macey et al. [4] ignored the data on phylogenetic relationships of Phrynocephalus and the resulting taxonomic acts published by previous authors [2,12], solely based on the fact that, as they stated, they failed to obtain the collection information on the specimens used in these studies [4]. We stress herein that in this age of internet communication and biodiversity crises, international cooperation is crucial for taxonomic practice in studies of herpetofaunal diversity in Asia, especially for such a complicated and insufficiently studied group as Phrynocephalus [29]. Not only is this paramount for elaboration of an adequate taxonomy, any comparative analyses, it is now also becoming a fundamental cornerstone of biodiversity conservation.
5. Conclusions
In the present study, we constructed an extensive COI barcoding library for Phrynocephalus, a taxonomically challenging genus of Agamidae, representing a key element of arid habitats in Middle and Central Asia, Middle and Near East. We have successfully recovered barcodes from 385 specimens representing over 90% of the nominal Phrynocephalus species, covering the intraspecific diversity of most species. We use a number of different approaches of single locus molecular species delimitation to infer the fit of the recovered MOTUs to the current taxonomy of the genus. Developing a reliable DNA barcode library for Phrynocephalus is especially important, as numerous previous works, e.g., [1,4,7,23], and sequences published in GenBank contain inconsistent taxonomic information or are based on misidentifications. Misidentified specimens or mislabeled sequences can lead to profound mistakes in taxonomy [142] and result in under- and over-estimation of genetic diversity between and within species [143,144]. Therefore, the goal of our manuscript is to scrutinize the current knowledge on Phrynocephalus diversity, thereby setting the ground for further in-depth integrative studies on systematics and phylogenetic relationships of this genus based on multilocus phylogenetic approaches and analyses of additional types of data, such as morphology, distribution, and ecology.
Our paper provides further evidence for the effectiveness of the multi-tier species delimitation analyses within an integrative taxonomic framework [89]. We demonstrate that alternative species delimitation algorithms may provide a confusingly wide range of possible number of MOTUs (from 54 to 103 in our empirical study). We argue that the multi-tier approach applied herein, which combines various methods of species delimitation resulting in a consensus species hypothesis, is crucial for a more accurate species delimitation in this taxonomically challenging group of lizards. Though the analysis of morphological data on Phrynocephalus lies outside the scope of our present study, further studies combining our results with the lines of evidence from nuclear genome (preferably on a genomic level), along with the data from morphology and distribution, will likely result in a robust taxonomy of the agamid genus Phrynocephalus. It should be noted that the use of DNA barcoding can fail in case of cryptic species, when hybridization is occurring between species, or when incipient speciation is taking place. These processes were previously documented for a number of Phrynocephalus species complexes [14,98,100,101], making the application of DNA barcoding challenging in these groups. However, while it is important to not over-interpret results from DNA barcoding, this method still can be helpful in detecting groups or populations in which the abovementioned processes may be occurring, thereby highlighting areas for future investigations. We stress herein the need to evaluate numerous lines of taxonomic evidence for an objective, repeatable hypothesis of species limits in future studies of Phrynocephalus lizards.
Finally, our results pinpoint cryptic diversity that requires further molecular and taxonomic evaluation, suggesting that the present taxonomy underestimates the actual diversity of the genus Phrynocephalus. Our study tentatively recognizes 63 MOTU within the genus and also reports on 13 previously unknown and currently unnamed Phrynocephalus lineages which we identify here as candidate species. Among all species delimitation algorithms tested in the present study, the ASAP algorithm most closely reflects the current taxonomy of Phrynocephalus, while the mlPTP and hsPTP analyses greatly overestimate the number of species in this group. While we cannot rely on mtDNA markers alone to delimit and describe new species, uncovering high levels of intraspecific mtDNA diversity provides important clues for targeting putative cryptic taxa that should be further investigated using genomic-level data. We further discuss that the species delimitation in Phrynocephalus based exclusively on mtDNA markers is insufficient due to the mito-nuclear discordance resulting from interspecific hybridization likely often recorded in this group. We call for international cooperation and further integrative taxonomic studies joining the data from morphology, mtDNA and nuDNA markers to fully stabilize the taxonomy of Phrynocephalus lizards.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d15020149/s1, Figure S1: Maximum Likelihood tree. Values near the nodes represent bootstrap support from ML/posterior probabilities from BI; Figure S2: Bayesian inference phylogenetic tree of COI gene haplotypes of the genus Phrynocephalus. Black circles correspond to node support values (BI PP) > or = 0.99, white circles correspond to BI PP-values 0.95 < 0.99, no circles indicate nodes with support values lower than BI PP 0.95. For details on haplotype information see Supplementary Table S1; Table S1: List of sequences and samples used in molecular analysis; Table S2: Uncorrected p-distances (%) for sequences of COI mtDNA for Phrynocephalus between different forms (below diagonal). Values on the diagonal correspond to uncorrected interspecific p-distances. Standard error estimates are shown above the diagonal. References [2,12,13,15,18,19,21,22,23,24,25,26,86,87,145,146,147,148,149] are cited in the Supplementary Materials.
Author Contributions
Conceptualization, methodology E.N.S. and N.A.P.; sample preparation, E.N.S., E.A.D., R.A.N., D.A.B. and N.A.P.; molecular analysis, E.N.S.; data analysis, E.N.S.; writing—original draft, E.N.S. and N.A.P.; writing—review and editing, E.N.S., E.A.D., D.A.B., R.A.N. and N.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
Specimen storage and identification was conducted within the frameworks of the Russian State Project (AAAA-A16-116021660077-3). Study was completed with financial support of Russian Science Foundation grant no. 22-14-00037 (molecular and phylogenetic analyses, specimen collection) and with support of Russian Foundation for Basic Research grant no. 20-54-56033 and Iran National Science Foundation (INSF) grant no. 99003440 (specimen collection, specimen examination).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data presented in this study are available in GenBank (https://www.ncbi.nlm.nih.gov/ (accessed on 29 August 2022).
Acknowledgments
We are grateful to Alex Borisenko, Natalia V. Ivanova and Paul D. N. Hebert for the opportunity to process our samples of Phrynocephalus within the BOLD project in Canada, and to colleagues, who took part in providing samples and discussion of results: Marina A. Chirikova, Tatyana N. Duysebayeva, Anna A. Bannikova, Vladimir S. Lebedev, Pavel V. Kvartalnov, and Leonid A. Neymark. For permission to study specimens under her care and for permanent support, we thank Valentina F. Orlova (ZMMU). We are grateful to Christopher Joldnall for proofreading and linguistic help. We would like to thank two anonymous reviewers for their comments on the earlier version of the manuscript, and especially to Yuanting Jin for the constructive criticism, sharing his original data on mitogenomes of Chinese Phrynocephalus, and for numerous comments which helped us to improve this study.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Barabanov, A.V.; Ananjeva, N.B. Catalogue of the available scientific species-group names for lizards of the genus Phrynocephalus Kaup, 1825 (Reptilia, Sauria, Agamidae). Zootaxa 2007, 1399, 1–56. [Google Scholar] [CrossRef]
- Solovyeva, E.N.; Lebedev, V.S.; Dunayev, E.A.; Nazarov, R.A.; Bannikova, A.A.; Che, J.; Murphy, R.W.; Poyarkov, N.A. Cenozoic aridization in Central Eurasia shaped diversification of toad-headed agamas (Phrynocephalus; Agamidae, Reptilia). PeerJ. 2018, 6, e4543. [Google Scholar] [CrossRef] [PubMed]
- Pallas, P.S. Reise Durch Verschiedene Provinzen des Russischen Reichs. Bd. I; Kayserliche Academie der Wissenschaften: St. Petersburg, Russia, 1771; 540p. [Google Scholar]
- Macey, J.R.; Schulte, J.A.; Ananjeva, N.B.; Van Dyke, E.T.; Wang, Y.; Orlov, N.; Shafiei, S.; Robinson, M.D.; Dujsebayeva, T.; Freund, G.S.; et al. A molecular phylogenetic hypothesis for the Asian agamid lizard genus Phrynocephalus reveals discrete biogeographic clades implicated by plate tectonics. Zootaxa 2018, 4467, 1–81. [Google Scholar] [CrossRef] [PubMed]
- Wermuth, H. Liste der rezenten Amphibien und Reptilien. Agamidae. Tierreich 1967, 86, 127. [Google Scholar]
- Moody, S.M. Phylogenetic and Historical Biogeographical Relationships of the Genera in the Family Agamidae (Reptilia: Lacertilia). Ph.D. Thesis, University of Michigan, Ann Arbor, MI, USA, 1980; 373p. [Google Scholar]
- Guo, X.; Wang, Y. Partitioned Bayesian analyses, dispersal–vicariance analysis, and the biogeography of Chinese toad-headed lizards (Agamidae: Phrynocephalus): A re-evaluation. Mol. Phylogenet. Evol. 2007, 45, 643–662. [Google Scholar] [CrossRef]
- Kamali, K.; Anderson, S. A new Iranian Phrynocephalus (Reptilia: Squamata: Agamidae) from the hottest place on earth and a key to the genus Phrynocephalus in southwestern Asia and Arabia. Zootaxa 2015, 3904, 249–260. [Google Scholar] [CrossRef]
- Uetz, P.; Freed, P.; Hošek, J. The Reptile Database. Available online: http://reptiledatabase.reptarium.cz (accessed on 27 August 2022).
- Golubev, M.L. About the name Agama ocellata Lichtenstein in Eversmann, 1823 (Reptilia, Agamidae) with redescription of the types. Herpetol. Res. 1991, 1, 12–17, (In Russian with English summary). [Google Scholar]
- Dunayev, E.A. Nomenclature and distribution of toad-agams, Phrynocephalus (Reptilia, Agamidae) in Iliyskaya Hollow. Bull. Mosc. Soc. Nat. Biol. Ser. 1996, 101, 36–41, (In Russian with English summary). [Google Scholar]
- Solovyeva, E.N.; Poyarkov, N.A.; Dunayev, E.A.; Nazarov, R.A.; Lebedev, V.S.; Bannikova, A.A. Phylogenetic relationships and subgeneric taxonomy of toad-headed agamas Phrynocephalus (Reptilia, Squamata, Agamidae) based on mitochondrial DNA sequence data. Dokl. Biol. Sci. 2014, 455, 119–124. [Google Scholar] [CrossRef]
- Dunayev, E.A.; Solovyeva, E.N.; Poyarkov, N.A. Taxonomy, Phylogeny and Distribution of Phrynocephalus (superspecies guttatus) (Reptilia: Agamidae). Curr. Stud. Herpetol. 2020, 20, 16–34. (In Russian) [Google Scholar] [CrossRef]
- Dunayev, E.A. History of study, taxonomy, distribution, and ecology of Phrynocephalus nasatus Golubev et Dunayev, 1995 (Reptilia: Agamidae). Russ. J. Herpetol. 2020, 27, 87–96. [Google Scholar] [CrossRef]
- Dunayev, E.A.; Solovyeva, E.N.; Poyarkov, N.A. Systematics, phylogeny and evolution of Phrynocephalus (superspecies przewalskii) (Reptilia: Agamidae). Russ. J. Herpetol. 2021, 28, 43–59. [Google Scholar] [CrossRef]
- Hu, C.; Wu, Y.; Ma, L.; Chen, Y.-J.; Ji, X. Genetic and morphological divergence among three closely related Phrynocephalus species (Agamidae). BMC Evol. Biol. 2019, 19, 114. [Google Scholar] [CrossRef]
- Melnikov, D.; Melnikova, E.; Nazarov, R.; Rajabizadeh, M.; Al-Johany, A.; Amr, Z.S.; Ananjeva, N.B. Taxonomic revision of Phrynocephalus arabicus Anderson, 1984 complex with description of a new species from Ahvaz, south-western Iran. Russ. J. Herpetol. 2014, 21, 149–159. [Google Scholar]
- Melnikov, D.; Melnikova, E.; Nazarov, R.; Al-Johany, A.; Ananjeva, N. A new species of Phrynocephalus (Agamidae, Sauria) from Al Sharqiyah sands, Northeastern Oman, dedicated to the memory of Sako Tuniyev (1983–2015). Russ. J. Herpetol. 2015, 22, 301–309. [Google Scholar]
- Solovyeva, E.N.; Poyarkov, N.A.; Dunaev, E.A.; Duysebayeva, T.N.; Bannikova, A.A. Molecular differentiation and taxonomy of the sunwatcher toad headed agama species complex Phrynocephalus superspecies helioscopus (Pallas 1771) (Reptilia: Agamidae). Russ. J. Genet. 2015, 47, 842–856. [Google Scholar]
- Solovyeva, E.N.; Dunayev, E.A.; Poyarkov, N.A. Interspecific taxonomy of sunwatcher toadhead agama species complex (Phrynocephalus helioscopus, Squamata). Zool. Z. 2012, 91, 1377–1396. [Google Scholar]
- Solovyeva, E.N.; Dunayev, E.A.; Nazarov, R.A.; Radjabizadeh, M.; Poyarkov, N.A. Molecular and morphological differentiation of Secret Toad-headed agama, Phrynocephalus mystaceus, with the description of a new subspecies from Iran (Reptilia, Agamidae). ZooKeys 2018, 748, 97–129. [Google Scholar] [CrossRef]
- Shao, M.; Ma, L.; Wang, Z. The complete mitochondrial genome of the toad-headed lizard, Phrynocephalus forsythii (Reptilia, Squamata, Agamidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 3147–3148. [Google Scholar] [CrossRef] [PubMed]
- Shao, M.; Ma, L.; Zhang, G.; Wang, Z. The complete mitochondrial genome of the toad-headed lizard, Phrynocephalus albolineatus (Reptilia, Squamata, Agamidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2017, 28, 137–138. [Google Scholar] [CrossRef]
- Song, S.; Li, D.; Zhang, C.; Jiang, K.; Zhang, D.; Chang, C. The complete mitochondrial genome of the color changeable toad-headed agama, Phrynocephalus versicolor (Reptilia, Squamata, Agamidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2014, 27, 1121–1122. [Google Scholar] [CrossRef]
- Liao, P.; Jin, Y. The complete mitochondrial genome of the toad-headed lizard subspecies, Phrynocephalus theobaldi orientalis (Reptilia, Squamata, Agamidae). Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Liao, P.; Tong, H.; Jin, Y. The complete mitochondrial genome of the subspecies, Phrynocephalus erythrurus parva (Reptilia, Squamata, Agamidae), a toad-headed lizard dwell at highest elevations of any reptile in the world. Mitochondrial DNA Part A DNA Mapp. Seq. Anal. 2016, 27, 703–704. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.; Ratnasingham, S.; De Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270 (Suppl. 1), S96–S99. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [PubMed]
- Murphy, R.W.; Crawford, A.J.; Bauer, A.M.; Che, J.; Donnellan, S.C.; Fritz, U.; Haddad, C.F.B.; Nagy, Z.T.; Poyarkov, N.A.; Vences, M.; et al. Cold Code: The global initiative to DNA barcode amphibians and nonavian reptiles. Mol. Ecol. Resour. 2013, 13, 161–167. [Google Scholar] [CrossRef]
- Crawford, A.J.; Lips, K.R.; Bermingham, E. Epidemic disease decimates amphibian abundance, species diversity, and evolutionary history in the highlands of central Panama. Proc. Natl. Acad. Sci. USA 2010, 107, 13777–13782. [Google Scholar] [CrossRef]
- Begerow, D.; Nilsson, H.; Unterseher, M.; Maier, W. Current state and perspectives of fungal DNA barcoding and rapid identification procedures. Appl. Microbiol. Biotechnol. 2010, 87, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Acha, G. ABGD, Automatic Barcode Gap Discovery for primary species delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef] [PubMed]
- Vieites, D.R.; Wollenberg, K.C.; Andreone, F.; Köhler, J.; Glaw, F.; Vences, M. Vast underestimation of Madagascar’s biodiversity evidenced by an integrative amphibian inventory. Proc. Natl. Acad. Sci. USA 2009, 106, 8267–8272. [Google Scholar] [CrossRef]
- Neigel, J.; Domingo, A.; Stake, J. DNA barcoding as a tool for coral reef conservation. Coral Reefs 2007, 26, 487. [Google Scholar] [CrossRef]
- Vernooy, R.; Haribabu, E.; Muller, M.R.; Vogel, J.H.; Hebert, P.D.; Schindel, D.E.; Shimura, J.; Singer, G.A. Barcoding life to conserve biological diversity: Beyond the taxonomic imperative. PLoS Biol. 2010, 8, e1000417. [Google Scholar] [CrossRef]
- Crawford, A.J.; Cruz, C.; Griffith, E.; Ross, H.; Ibáñez, R.; Lips, K.R.; Driskell, A.C.; Bermingham, E.; Crump, P. DNA barcoding applied to ex situ tropical amphibian conservation programme reveals cryptic diversity in captive populations. Mol. Ecol. Resour. 2013, 13, 1005–1018. [Google Scholar] [PubMed]
- Smith, M.A.; Poyarkov, N.A.; Hebert, P.D.N. DNA barcoding: CO1 DNA barcoding amphibians: Take the chance, meet the challenge. Mol. Ecol. Resour. 2008, 8, 235–246. [Google Scholar] [CrossRef]
- Vences, M.; Nagy, Z.T.; Sonet, G.; Verheyen, E. DNA barcoding amphibians and reptiles. In DNA Barcodes; Humana Press: Totowa, NJ, USA, 2012; pp. 79–107. [Google Scholar]
- Vasconcelos, R.; Montero-Mendieta, S.; Simó-Riudalbas, M.; Sindaco, R.; Santos, X.; Fasola, M.; Llorente, G.; Razzetti, E.; Carranza, S. Unexpectedly high levels of cryptic diversity uncovered by a complete DNA barcoding of reptiles of the Socotra Archipelago. PLoS ONE 2016, 11, e0149985. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Yang, C.; Ke, D. DNA barcoding and phylogenetic relationships in Anatidae. Mitochondrial DNA Part A 2014, 27, 1042–1044. [Google Scholar] [CrossRef]
- Huang, Z.; Ruan, R. DNA barcodes and insights into the phylogenetic relationships of Corvidae (Aves: Passeriformes). Mitochondrial DNA Part A 2018, 29, 529–534. [Google Scholar] [CrossRef]
- Shen, Y.; Guan, L.; Wang, D.; Gan, X. DNA barcoding and evaluation of genetic diversity in Cyprinidae fish in the midstream of the Yangtze River. Ecol. Evol. 2016, 6, 2702–2713. [Google Scholar] [CrossRef] [PubMed]
- Tizard, J.; Patel, S.; Waugh, J.; Tavares, E.; Bergmann, T.; Gill, B.; Norman, J.; Christidis, L.; Scofield, P.; Haddrath, O.; et al. DNA barcoding a unique avifauna: An important tool for evolution, systematics and conservation. BMC Evol. Biol. 2019, 19, 52. [Google Scholar] [CrossRef] [PubMed]
- Nagy, Z.T.; Sonet, G.; Glaw, F.; Vences, M. First Large-scale DNA barcoding assessment of reptiles in the biodiversity hotspot of Madagascar, based on newly designed COI primers. PLoS ONE 2012, 7, e34506. [Google Scholar] [CrossRef]
- Hartmann, T.; Geissler, P.; Poyarkov, N.A.; Ihlow, F.; Galoyan, E.A.; Rödder, D.; Böhme, W. A new species of the genus Calotes Cuvier, 1817 (Squamata: Agamidae) from southern Vietnam. Zootaxa 2013, 3599, 246–260. [Google Scholar] [CrossRef]
- Nazarov, R.; Poyarkov, N.A.; Orlov, N.L.; Phung, T.M.; Nguyen, T.T.; Hoang, D.M.; Ziegler, T. Two new cryptic species of the Cyrtodactylus irregularis complex (Squamata: Gekkonidae) from southern Vietnam. Zootaxa 2012, 3302, 1–24. [Google Scholar] [CrossRef]
- Nazarov, R.A.; Poyarkov, N.A.; Orlov, N.L.; Nguyen, N.S.; Milto, K.D.; Martynov, A.A.; Konstantinov, E.L.; Chulisov, A.S. A review of Cyrtodactylus (Reptilia: Sauria: Geckonidae) fauna of Laos with description of four new species. Proc. Zool. Inst. RAS 2014, 318, 391–423. [Google Scholar] [CrossRef]
- Neang, T.; Chan, S.; Poyarkov, N.A. A new species of smooth skink (Squamata: Scincidae: Scincella) from Cambodia. Zool. Res. 2018, 38, 214–233. [Google Scholar] [CrossRef]
- Puillandre, N.; Macpherson, E.; Lambourdière, J.; Cruaud, C.; Boisselier-Dubayle, M.C.; Samadi, S. Barcoding type specimens helps to identify synonyms and an unnamed new species in Eumunida Smith, 1883 (Decapoda: Eumunididae). Invertebr. Syst. 2011, 25, 322–333. [Google Scholar] [CrossRef]
- Reid, N.M.; Carstens, B.C. Phylogenetic estimation error can decrease the accuracy of species delimitation: A Bayesian implementation of the general mixed Yule-coalescent model. BMC Evol. Biol. 2012, 12, 196. [Google Scholar] [CrossRef]
- De Salle, R.O. Species discovery versus species identification in DNA barcoding efforts: Response to Rubinoff. Conserv. Biol. 2006, 20, 1545–1547. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 16. [Google Scholar] [CrossRef]
- Wiemers, M.; Fiedler, K. Does the DNA barcoding gap exist?—A case study in blue butterflies (Lepidoptera: Lycaenidae). Front. Zool. 2007, 4, 8. [Google Scholar] [CrossRef] [PubMed]
- Tingley, R.; Meiri, S.; Chapple, D.G. Addressing knowledge gaps in reptile conservation. Biol. Conserv. 2016, 204(Part A), 1–5. [Google Scholar] [CrossRef]
- Barcoding of Life Project. Available online: www.boldsystems.org (accessed on 1 June 2022).
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbour Laboratory Press: New York, NY, USA, 1989; 385p. [Google Scholar]
- Ivanova, N.V.; DeWaard, J.; Hebert, P.D.N. An inexpensive, automation friendly protocol for recovering high quality DNA. Mol. Ecol. Notes 2006, 6, 998–1002. [Google Scholar] [CrossRef]
- Borisenko, A.V.; Lim, B.K.; Ivanova, N.V.; Hanner, R.H.; Hebert, P.D. DNA barcoding in surveys of small mammal communities: A field study in Suriname. Mol. Ecol. Resour. 2008, 8, 471–479. [Google Scholar] [CrossRef]
- Borisenko, A.V.; Sones, J.E.; Hebert, P.D. The front-end logistics of DNA barcoding: Challenges and prospects. Mol. Ecol. Resour. 2009, 9, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Kruskop, S.V.; Borisenko, A.V.; Ivanova, N.V.; Lim, B.K.; Eger, J.L. Genetic diversity of northeastern Palaearctic bats as revealed by DNA barcodes. Acta Chiropterologica 2012, 14, 1–4. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. In Nucleic Acids Symposium Series; Information Retrieval Ltd.: London, UK, 1999; pp. 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Huelsenbeck, J. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef]
- Rambaut, A.; Drummond, A.J. Tracer v1. 5. 2007. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 3 July 2022).
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef]
- Huelsenbeck, J.P.; Hillis, D.M. Success of phylogenetic methods in the four-taxon case. Syst. Biol. 1993, 42, 247–264. [Google Scholar] [CrossRef]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble species by automatic partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.K.; Leaché, A.D.; Burbrink, F.T.; McGuire, J.A.; Moritz, C. Coalescent-based species delimitation in an integrative taxonomy. Trends Ecol. Evol. 2012, 27, 480–488. [Google Scholar] [CrossRef]
- Dellicour, S.; Flot, J.F. Delimiting species-poor data sets using single molecular markers: A study of barcode gaps, haplowebs and GMYC. Syst. Biol. 2015, 64, 900–908. [Google Scholar] [CrossRef]
- Eyer, P.A.; Seltzer, R.; Reiner-Brodetzki, T.; Hefetz, A. An integrative approach to untangling species delimitation in the Cataglyphis bicolor desert ant complex in Israel. Mol. Phylogenet. Evol. 2017, 115, 128–139. [Google Scholar] [CrossRef]
- Monaghan, M.T.; Wild, R.; Elliot, M.; Fujisawa, T.; Balke, M.; Inward, D.J.; Lees, D.C.; Ranaivosolo, R.; Eggleton, P.; Barraclough, T.G.; et al. Accelerated species inventory on Madagascar using coalescent-based models of species delineation. Syst. Biol. 2009, 58, 298–311. [Google Scholar] [CrossRef]
- Dellicour, S.; Flot, J.F. The hitchhiker’s guide to single-locus species delimitation. Mol. Ecol. Resour. 2018, 18, 1234–1246. [Google Scholar] [CrossRef]
- Meyer, C.P.; Paulay, G. DNA barcoding: Error rates based on comprehensive sampling. PLoS Biol. 2005, 3, e422. [Google Scholar] [CrossRef]
- Gorin, V.A.; Solovyeva, E.N.; Hasan, M.; Okamiya, H.; Karunarathna, D.M.S.S.; Pawangkhanant, P.; de Silva, A.; Juthong, W.; Milto, K.D.; Nguyen, L.T.; et al. A little frog leaps a long way: Compounded colonizations of the Indian Subcontinent discovered in the tiny Oriental frog genus Microhyla (Amphibia: Microhylidae). PeerJ. 2020, 8, e9411. [Google Scholar] [CrossRef] [PubMed]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [PubMed]
- Fujisawa, T.; Barraclough, T.G. Delimiting species using single-locus data and the Generalized Mixed Yule Coalescent approach: A revised method and evaluation on simulated data sets. Syst. Biol. 2013, 62, 707–724. [Google Scholar] [CrossRef] [PubMed]
- Ducasse, J.; Ung, V.; Lecointre, G.; Miralles, A. LIMES: A tool for comparing species partition. Bioinformatics 2020, 36, 2282–2283. [Google Scholar] [CrossRef]
- Melville, J.; Hale, J.; Mantziou, G.; Ananjeva, N.B.; Milto, K.; Clemann, N. Historical biogeography, phylogenetic relationships and intraspecific diversity of agamid lizards in the Central Asian deserts of Kazakhstan and Uzbekistan. Mol. Phylogenet. Evol. 2009, 53, 99–112. [Google Scholar] [CrossRef]
- Pang, J.; Wang, Y.; Zhong, Y.; Hoelzele, A.R.; Papenfuss, T.J.; Zeng, X.; Ananjeva, N.B.; Zhang, Y. A phylogeny of Chinese species in the genus Phrynocephalus (Agamidae) inferred from mitochondrial DNA sequences. Mol. Phylogenet. Evol. 2003, 27, 398–409. [Google Scholar] [CrossRef]
- Barabanov, A.M.; Ananjeva, N.B.; Papenfuss, T.J.; Wang, Y. A new name for Phrynocephalus theobaldi orientalis Wang, Papenfuss et Zeng, 1999. Russ. J. Herpetol. 2002, 9, 80. [Google Scholar]
- Jin, Y.T.; Brown, R. Partition number, rate priors and unreliable divergence times in Bayesian phylogenetic dating. Cladistics 2018, 34, 568–573. [Google Scholar] [CrossRef]
- Ebrahimipour, F.; Rastegar-Pouyani, N.; Rastegar Pouyani, E.; Hosseinian Yousefkhani, S.S.; Kamali, K. Molecular phylogenetic relationships within the Phrynocephalus maculatus–arabicus species complex (Sauria: Agamidae) in Iran. J. Zool. Syst. Evol. Res. 2021, 59, 1583–1588. [Google Scholar] [CrossRef]
- Schlick-Steiner, B.C.; Steiner, F.M.; Seifert, B.; Stauffer, C.; Christian, E.; Crozier, R.H. Integrative taxonomy: A multisource approach to exploring biodiversity. Annu. Rev. Entomol. 2010, 55, 421–438. [Google Scholar] [CrossRef]
- Xu, X.; Kuntner, M.; Bond, J.E.; Ono, H.; Liu, F.; Yu, L.; Li, D. A multi-tier species delimitation approach resolves conflicts in delineating the primitively segmented spider genus Heptathela endemic to Japanese islands. BioRxiv 2019. [Google Scholar] [CrossRef]
- Babik, W.; Branicki, W.; Crnobrnja-Isailović, J.; Cogălniceanu, D.; Sas, I.; Olgun, K.; Poyarkov, N.A.; Garcia-París, M.; Arntzen, J.W. Phylogeography of two European newt species—Discordance between mtDNA and morphology. Mol. Ecol. 2005, 14, 2475–2491. [Google Scholar] [CrossRef]
- Renoult, J.P.; Geniez, P.; Bacquet, P.; Benoit, L.; Crochet, P.-A. Morphology and nuclear markers reveal extensive mitochondrial introgressions in the Iberian Wall Lizard species complex. Mol. Ecol. 2009, 18, 4298–4315. [Google Scholar] [CrossRef]
- Leache, A.D.; Cole, C.J. Hybridization between multiple fence lizard lineages in an ecotone: Locally discordant variation in mitochondrial DNA, chromosomes, and morphology. Mol. Ecol. 2007, 16, 1035–1054. [Google Scholar] [CrossRef] [PubMed]
- Toews, D.P.; Brelsford, A. The biogeography of mitochondrial and nuclear discordance in animals. Mol. Ecol. 2012, 21, 3907–3930. [Google Scholar] [CrossRef] [PubMed]
- McGuire, J.A.; Linkem, C.W.; Koo, M.S.; Hutchison, D.W.; Lappin, A.K.; Orange, D.I.; Lemos-Espinal, J.; Riddle, B.R.; Jaeger, J.R. Mitochondrial introgression and incomplete lineage sorting through space and time: Phylogenetics of Crotaphytid lizards. Evolution 2007, 61, 2879–2897. [Google Scholar] [CrossRef]
- Nguyen, S.N.; Zhou, W.-W.; Le, T.-N.T.; Tran, A.-D.T.; Jin, J.-Q.; Vo, B.D.; Nguyen, L.T.; Nguyen, T.T.; Nguyen, T.Q.; Hoang, D.D.; et al. Cytonuclear discordance, cryptic diversity, complex histories, and conservation needs in Vietnamese bent-toed geckos of the Cyrtodactylus irregularis species complex. Russ. J. Herpetol. 2017, 24, 133–154. [Google Scholar] [CrossRef]
- Fisher-Reid, M.C.; Wiens, J.J. What are the consequences of combining nuclear and mitochondrial data for phylogenetic analysis? Lessons from Plethodon salamanders and 13 other vertebrate clades. BMC Evol. Biol. 2011, 11, 300. [Google Scholar] [CrossRef]
- Dunayev, E.A. Systematics and paleogeography: Conceptual synthesis by the example of Phrynocephalus (superspecies guttatus) (Reptilia: Agamidae). In Evolution and Systematics: Lamarck and Darwin in Modern Studies. Archiver of the Zoological Museum of Moscow State University; Sviridov, A.V., Shatalkin, A.I., Eds.; KMK Scientific Press: Moscow, Russia, 2009; Volume 50, pp. 274–298. (In Russian) [Google Scholar]
- Gozdzik, A.; Fu, J. Are toad-headed lizards Phrynocephalus przewalskii and frontalis (family Agamidae) the same species? Defining species boundaries with morphological and molecular data. Russ. J. Herpetol. 2009, 16, 107–118. [Google Scholar]
- Urquhart, J.; Bi, K.E.; Gozdzik, A.; Fu, J. Isolation and characterization of microsatellite DNA loci in the toad-headed lizards, Phrynocephalus przewalskii complex. Mol. Ecol. Notes 2005, 5, 928–930. [Google Scholar] [CrossRef]
- Urquhart, J.; Wang, Y.; Fu, J. Historical vicariance and male-mediated gene flow in the toad-headed lizards Phrynocephalus przewalskii. Mol. Ecol. 2009, 18, 3714–3729. [Google Scholar] [CrossRef] [PubMed]
- Golubev, M.L.; Dunayev, E.A. Phrynocephalus nasatus (Reptilia, Agamidae), a new species of toad agama from Western China. Russ. J. Herpetol. 1995, 2, 5–9. [Google Scholar]
- Grummer, J.A.; Bryson, R.W.; Reeder, T.W. Species delimitation using Bayes factors: Simulations and application to the Sceloporus scalaris species group (Squamata: Phrynosomatidae). Syst. Biol. 2014, 63, 119–133. [Google Scholar] [CrossRef] [PubMed]
- Firkowski, C.R.; Bornschein, M.R.; Ribeiro, L.F.; Pie, M.R. Species delimitation, phylogeny and evolutionary demography of co-distributed, montane frogs in the southern Brazilian Atlantic Forest. Mol. Phylogenet. Evol. 2016, 100, 345–360. [Google Scholar] [CrossRef]
- Pie, M.R.; Bornschein, M.R.; Ribeiro, L.F.; Faircloth, B.C.; McCormack, J.E. Phylogenomic species delimitation in microendemic frogs of the Brazilian Atlantic Forest. Mol. Phylogenet. Evol. 2019, 141, 106627. [Google Scholar] [CrossRef]
- Wilson, E.O.; Brown, W.L. The subspecies concept and its taxonomic application. Syst. Zool. 1953, 2, 97–111. [Google Scholar] [CrossRef]
- Frost, D.R.; Hillis, D.M. Species in concept and practice: Herpetological applications. Herpetologica 1990, 46, 87–104. [Google Scholar]
- Frost, D.R.; Kluge, A.G.; Hillis, D.M. Species in contemporary herpetology: Comments on phylogenetic inference and taxonomy. Herpetol. Rev. 1992, 23, 46–54. [Google Scholar]
- Kindler, C.; Fritz, U. Phylogeography and taxonomy of the barred grass snake (Natrix helvetica), with a discussion of the subspecies category in zoology. Vertebr. Zool. 2018, 68, 269–281. [Google Scholar]
- De Queiroz, K. An updated concept of subspecies resolves a dispute about the taxonomy of incompletely separated lineages. Herpetol. Rev. 2020, 51, 459–461. [Google Scholar]
- Hillis, D.M. New and not-so-new conceptualizations of species and subspecies: A reply to the “It’s species all the way down” view. Herpetol. Rev. 2021, 52, 49–50. [Google Scholar]
- Marshall, T.L.; Chambers, E.A.; Matz, M.V.; Hillis, D.M. How mitonuclear discordance and geographic variation have confounded species boundaries in a widely studied snake. Mol. Phylogenet. Evol. 2021, 162, 107194. [Google Scholar] [CrossRef]
- Dunayev, E.A. Phylogeny of lizards of Phrynocephalus genus (Reptilia: Agamidae): History of study and methodical approaches. In Voprosy Gerpetologii, Proceedings of the 3rd Meeting of the Nikolsky Herpetological Society, Pushchino, Russia, 9–13 October 2006; Zoological Institute: St. Petersburg, Russia, 2008; pp. 117–126. (In Russian) [Google Scholar]
- Melnikov, D.; Melnikova, E.; Nazarov, R.; Rajabizadeh, M. Taxonomic revision of Phrynocephalus persicus De Filippi, 1863 complex with description of a new species from Zagros, southern Iran. Curr. Stud. Herpetol. 2013, 13, 34–46. [Google Scholar]
- Chernov, S.A. Reptiles—Reptilia. In The Animals of the USSR. Volume 2: The Desert Zone; Pavlovsky, E.N., Vinogradova, B.S., Eds.; USSR Academy of Sciences: Moscow-Leningrad, Russia, 1948; pp. 127–161. (In Russian) [Google Scholar]
- Lichtenstein, H. Nomenclator Reptilium et Amphibiorum Musei Zoologici Berolinensis. Namenverzeichniss der in der zoologischen Sammlung der Koniglichen Universitat zu Berlin aufgestellten Arten von Reptilien und Amphibien nach ihren Ordnungen, Familien und Gattungen; Akademie der Wissenschaften: Berlin, Germany, 1856; 48p. [Google Scholar]
- Rahiamian, H.; Shafiei, S.; Rastegar Pouyani, N.; Rastegar Pouyani, E. Phylogenetic relationships of the gray-toad agama, Phrynocephalus scutellatus (Olivier, 1807), species complex from Iran. Zootaxa 2015, 3990, 369–380. [Google Scholar] [CrossRef] [PubMed]
- Olivier, G.A. Voyage dans l’Empire Ottoman, l’Egypte et al Perse; Agasse: Paris, France, 1807; Volume 3. [Google Scholar]
- Nikolsky, A.M. Reptiles and amphibians collected by N.A. Zarudnyi in Persia in 1903-1904. Annu. Musée Zool. Acad. Imp. Sci. St. Pétersb. 1907, 10, 260–301. (In Russian) [Google Scholar]
- Guo, X.; Li, L.; Wang, Y. Phylogeography of the Phrynocephalus vlangalii Species Complex in the Upper Reaches of the Yellow River Inferred from mtDNA ND4-tRNA(LEU) Segments. Asian Herpetol. Res. 2012, 3, 52–68. [Google Scholar] [CrossRef]
- Jin, Y.; Brown, R.P.; Liu, N. Cladogenesis and phylogeography of the lizard Phrynocephalus vlangalii (Agamidae) on the Tibetan plateau. Mol. Ecol. 2008, 17, 1971–1982. [Google Scholar] [CrossRef]
- Chen, Y.J.; Zhu, L.; Wu, Q.N.; Hu, C.C.; Qu, Y.F.; Ji, X. Geological and climatic influences on population differentiation of the Phrynocephalus vlangalii species complex (Sauria: Agamidae) in the northern Qinghai-Tibet Plateau. Mol. Phylogenet. Evol. 2022, 169, 107394. [Google Scholar] [CrossRef]
- Jin, Y.; Brown, R. Morphological species and discordant mtDNA: A genomic analysis of Phrynocephalus lizard lineages on the Qinghai-Tibetan Plateau. Mol. Phylogenet. Evol. 2019, 139, 106523. [Google Scholar] [CrossRef]
- Jin, Y.; Yang, Z.; Brown, R.P.; Liao, P.; Liu, N. Intraspecific lineages of the lizard Phrynocephalus putjatia from the Qinghai-Tibetan Plateau: Impact of physical events on divergence and discordance between morphology and molecular markers. Mol. Phylogenet. Evol. 2014, 71, 288–297. [Google Scholar] [CrossRef] [PubMed]
- Blyth, E. Report of the Curator, Zoological Department. Proc. Asiat. Soc. Bengal 1863, 32, 79–90. [Google Scholar]
- Jin, Y.; Liu, N.; Brown, R. The geography and timing of genetic divergence in the lizard Phrynocephalus theobaldi on the Qinghai-Tibetan plateau. Sci. Rep. 2017, 7, 2281. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, N. Phylogeography of Phrynocephalus erythrurus from the Qiangtang Plateau of the Tibetan Plateau. Mol. Phylogenet. Evol. 2010, 54, 933–940. [Google Scholar] [CrossRef] [PubMed]
- Ananjeva, N.B. On the validity of Megalochilus mystaceus (Pallas, 1776). In Systematics and Ecology of Amphibians and Reptiles; Ananjeva, N.B., Borkin, L.J., Eds.; Zoological Institute USSR Academy of Sciences: Leningrad, Russia, 1987; Volume 157, pp. 4–13. (In Russian) [Google Scholar]
- Alekperov, A.M.; Galaeva, N.M. Pholidosis of Persian toad-headed agama Phrynocephalus helioscopus persicus De Filippi. Scientific proceedings of Azerbaijan University. Biol. Sci. 1975, 11, 54–57. (In Russian) [Google Scholar]
- Alekperov, A.M. Amphibians and Reptiles of Azerbaijan; Elm Press: Baku, Azerbeijan, 1978; p. 264. (In Russian) [Google Scholar]
- Manilo, V.V. Description of Karyotypes of Some Species and Subspecies of the Genus Phrynocephalus (Sauria, Agamidae) from Central Asia. Vestn. Zool. 2000, 34, 113–118. (In Russian) [Google Scholar]
- Manilo, V.V. Cytogenetic review and evolution of karyotypes in the species of the genus Phrynocephalus Kaup, 1825 (Sauria, Agamidae) from the Eastern Palaearctic. Hamadryad 2001, 26, 227–234. [Google Scholar]
- Manilo, V.V.; Golubev, M.L. Karyotype information on some toad agamas of the Phrynocephalus species group (Sauria, Agamidae) of the former USSR. Asiat. Herpetol. Res. 1993, 5, 105–108. [Google Scholar]
- Szczerbak, N.N. Zoogeographic Analysis of Reptiles of Turkmenistan. In Biogeography and Ecology of Turkmenistan; Springer: Dordrecht, The Netherlands, 1994; pp. 307–328. [Google Scholar]
- Semenov, D.V.; Brushko, Z.K.; Kubykin, R.A.; Shenbrot, G.I. Taxonomic position and protective status of the round-headed lizard (Reptilia, Agamidae) in the territory of the USSR. Zool. Z. 1987, 66, 98–109, (In Russian with English Summary). [Google Scholar]
- Ananjeva, N.B.; Borkin, L.J.; Darevsky, I.S.; Orlov, N.L. Amphibians and Reptiles. Encyclopedia of the Nature in Russia; ABF: Moscow, Russia, 1998; p. 574. (In Russian) [Google Scholar]
- Ananjeva, N.B.; Orlov, N.L.; Khalikov, R.G.; Darevsky, I.S.; Ryabov, S.A.; Barabanov, A.V. The Reptiles of Northern Eurasia; Pensoft Publishers: Sofia, Bulgaria, 2006; p. 245. [Google Scholar]
- Barts, M.; Wilms, T. Die Agamen der Welt. Draco 2003, 4, 4–23. [Google Scholar]
- Wang, Y.; Fu, J. Cladogenesis and vicariance patterns in the toad-headed lizard Phrynocephalus versicolor species complex. Copeia 2004, 2, 199–206. [Google Scholar] [CrossRef]
- Jin, Y.T.; Brown, R. Species history and divergence times of viviparous and oviparous Chinese toad-headed sand lizards (Phrynocephalus) on the Qinghai-Tibetan Plateau. Mol. Phylogenet. Evol. 2013, 68, 259–268. [Google Scholar] [CrossRef] [PubMed]
- Strauch, A.A. Part III. Reptilia and Amphibia. In Mongolia and the Tangut Lands, a Three-Year Journey in Eastern High Asia. Vol. II, Reptiles and Amphibians; Przevalsky, N.M., Ed.; Imperial Russian Geographical Society: St. Petersburg, Russia, 1876; pp. 1–55. (In Russian) [Google Scholar]
- Alcock, A.W.; Finn, F. An account of the Reptilia collected by Dr. F. Maynard, Captain, A.H. McMahon, C.I.E., and the members of the Afghan–Baluch Boundary Comission of 1896. J. Asiat. Soc. Bengal 1897, 65, 550–566. [Google Scholar]
- Kitchener, A.C.; Machado, F.A.; Hayssen, V.; Moehlman, P.D.; Viranta, S.; Esselstyn, J. Consequences of the misidentification of museum specimens: The taxonomic status of Canis lupaster soudanicus. J. Mammology 2020, 101, 1148–1150. [Google Scholar] [CrossRef]
- Chambers, E.A.; Hebert, P.D. Assessing DNA barcodes for species identification in North American reptiles and amphibians in natural history collections. PLoS ONE 2016, 11, e0154363. [Google Scholar] [CrossRef] [PubMed]
- Mulcahy, D.G.; Ibáñez, R.; Jaramillo, C.A.; Crawford, A.J.; Ray, J.M.; Gotte, S.W.; Jacobs, J.F.; Wynn, A.H.; Gonzalez-Porter, G.P.; McDiarmid, R.W.; et al. DNA barcoding of the National Museum of Natural History reptile tissue holdings raises concerns about the use of natural history collections and the responsibilities of scientists in the molecular age. PLoS ONE 2022, 17, e0264930. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, J.; Chen, M.; Xu, R. Phrynocephalus nasatus Mitochondrion, Complete Genome. Available online: https://www.ncbi.nlm.nih.gov/nuccore/MW456559.1 (accessed on 1 July 2022).
- Chen, D.; Zhou, T.; Guo, X. The complete mitochondrial genome of Phrynocephalus forsythii (Reptilia, Squamata, Agamidae), a toad-headed agama endemic to the Taklamakan Desert. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 4046–4048. [Google Scholar] [CrossRef]
- Tong, H.; Jin, Y. The complete mitochondrial genome of an agama, Phrynocephalus putjatia (Reptilia, Squamata, Agamidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 1028–1029. [Google Scholar] [CrossRef]
- Fu, C.; Chen, W.; Jin, Y. The complete mitochondrial genome of Phrynocephalus guinanensis (Reptilia, Squamata, Agamidae). Mitochondrial DNA A DNA Mapp. Seq. Anal. 2016, 27, 1103–1104. [Google Scholar] [CrossRef]
- Gu, H.F.; Xia, Y.; Penga, R.; Mo, B.H.; Li, L.; Zenga, X.M. Authentication of Chinese crude drug gecko by DNA barcoding. Nat. Prod. Commun. 2011, 6, 67–71. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).