
Comparative Analysis of Bacteria, Fungi, and Arbuscular Mycorrhizal Fungi in Medicinal Plants Lippia alba and Petiveria alliacea in Colombia

 , , , and
, , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
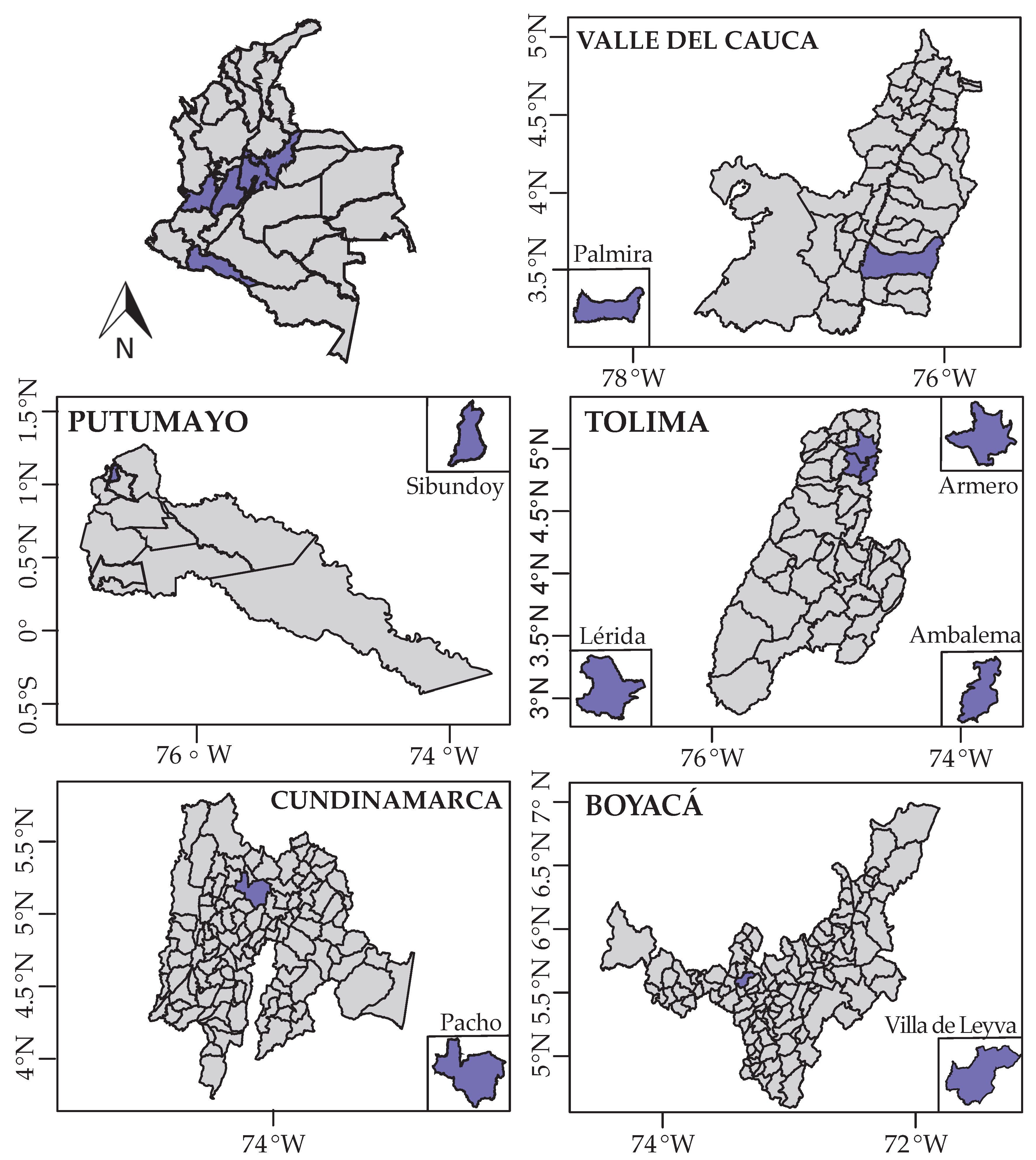
2.1. Soil Sampling of Medicinal Plants
2.2. Physicochemical Measurements of Soils
2.3. DNA Extraction and Metabarcoding from Bacteria, Fungi, and AMF
2.4. Bioinformatic and Diversity Analyses
3. Results
3.1. Soil Physicochemical Analysis
3.2. Comparative Analysis of Soil and Root Microbiome
3.2.1. Rhizosphere Soil and Root Bacteria Associated with Medicinal Plants
3.2.2. Rhizosphere Soil and Root Fungi Associated with Medicinal Plants
3.2.3. Taxonomic Assignation of AMF to Medicinal Plants
3.3. Diversity Analysis by Sample Type and Medicinal Plants
3.4. Multiple Factor Analysis for Bacteria and Fungi
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lee, S.-J.; Kong, M.; St-Arnaud, M.; Hijri, M. Arbuscular Mycorrhizal Fungal Communities of Native Plant Species under High Petroleum Hydrocarbon Contamination Highlights Rhizophagus as a Key Tolerant Genus. Microorganisms 2020, 8, 872. [Google Scholar] [CrossRef] [PubMed]
- Meng, S.; Peng, T.; Liu, X.; Wang, H.; Huang, T.; Gu, J.-D.; Hu, Z. Ecological Role of Bacteria Involved in the Biogeochemical Cycles of Mangroves Based on Functional Genes Detected through GeoChip 5.0. mSphere 2022, 7, e00936-21. [Google Scholar] [CrossRef] [PubMed]
- Vejan, P.; Abdullah, R.; Khadiran, T.; Ismail, S.; Nasrulhaq Boyce, A. Role of Plant Growth Promoting Rhizobacteria in Agricultural Sustainability—A Review. Molecules 2016, 21, 573. [Google Scholar] [CrossRef]
- Banerjee, S.; van der Heijden, M.G.A. Soil Microbiomes and One Health. Nat. Rev. Microbiol. 2023, 21, 6–20. [Google Scholar] [CrossRef]
- Busby, R.R.; Barbato, R.A.; Jung, C.M.; Bednar, A.J.; Douglas, T.A.; Ringelberg, D.B.; Indest, K.J. Alaskan Plants and Their Assembled Rhizosphere Communities Vary in Their Responses to Soil Antimony. Appl. Soil Ecol. 2021, 167, 104031. [Google Scholar] [CrossRef]
- Li, Y.; Yang, Y.; Wu, T.; Zhang, H.; Wei, G.; Li, Z. Rhizosphere Bacterial and Fungal Spatial Distribution and Network Pattern of Astragalus Mongholicus in Representative Planting Sites Differ the Bulk Soil. Appl. Soil Ecol. 2021, 168, 104114. [Google Scholar] [CrossRef]
- Pascale, A.; Proietti, S.; Pantelides, I.S.; Stringlis, I.A. Modulation of the Root Microbiome by Plant Molecules: The Basis for Targeted Disease Suppression and Plant Growth Promotion. Front. Plant Sci. 2020, 10, 1741. [Google Scholar] [CrossRef]
- Zhang, W.; Mason, G.A. Modulating the Rhizosphere Microbiome by Altering the Cocktail of Root Secretions. Plant Physiol. 2022, 188, 12–13. [Google Scholar] [CrossRef]
- Wu, L.; Wang, J.; Huang, W.; Wu, H.; Chen, J.; Yang, Y.; Zhang, Z.; Lin, W. Plant-Microbe Rhizosphere Interactions Mediated by Rehmannia Glutinosa Root Exudates under Consecutive Monoculture. Sci. Rep. 2015, 5, 15871. [Google Scholar] [CrossRef]
- Amani Machiani, M.; Javanmard, A.; Habibi Machiani, R.; Sadeghpour, A. Arbuscular Mycorrhizal Fungi and Changes in Primary and Secondary Metabolites. Plants 2022, 11, 2183. [Google Scholar] [CrossRef]
- Goodwin, P.H. The Rhizosphere Microbiome of Ginseng. Microorganisms 2022, 10, 1152. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Yang, J.; Zhao, S.; Yu, Q.; Weng, L.; Xiao, C. Root Exudates Influence Rhizosphere Fungi and Thereby Synergistically Regulate Panax Ginseng Yield and Quality. Front. Microbiol. 2023, 14, 1194224. [Google Scholar] [CrossRef] [PubMed]
- Castañeda, R.; Cáceres, A.; Velásquez, D.; Rodríguez, C.; Morales, D.; Castillo, A. Medicinal Plants Used in Traditional Mayan Medicine for the Treatment of Central Nervous System Disorders: An Overview. J. Ethnopharmacol. 2022, 283, 114746. [Google Scholar] [CrossRef] [PubMed]
- Montero-Villegas, S.; Crespo, R.; Rodenak-Kladniew, B.; Castro, M.A.; Galle, M.; Cicció, J.F.; García de Bravo, M.; Polo, M. Cytotoxic Effects of Essential Oils from Four Lippia Alba Chemotypes in Human Liver and Lung Cancer Cell Lines. J. Essent. Oil Res. 2018, 30, 167–181. [Google Scholar] [CrossRef]
- Morais, S.M.D.; Sobrinho, A.C.N.; Liberato, H.R.; Pereira, R.D.C.A.; Pessoa, C.; Alves, D.R.; Fontenelle, R.O.D.S. Biotechnological potential of essential oils from different chemotypes of Lippia alba(Mill.) N.E.Br. ex Britton & P. Wilson. Boletín Latinoam. Y Caribe Plantas Med. Y Aromáticas 2022, 21, 725–736. [Google Scholar] [CrossRef]
- García, L.T.; Leal, A.F.; Moreno, É.M.; Stashenko, E.E.; Arteaga, H.J. Differential Anti-Proliferative Effect on K562 Leukemia Cells of Lippia Alba (Verbenaceae) Essential Oils Produced under Diverse Growing, Collection and Extraction Conditions. Ind. Crops Prod. 2017, 96, 140–148. [Google Scholar] [CrossRef]
- Ortiz, N.; Jiménez, M.F.; Chaverri, C.; Cicció, J.F.; Díaz, C. Effect on Cell Growth, Viability and Migration of Geraniol and Geraniol-Containing Essential Oil from Lippia Alba (Verbenaceae) on Gastric Carcinoma Cells. J. Essent. Oil Res. 2022, 34, 65–76. [Google Scholar] [CrossRef]
- Hernández, J.F.; Urueña, C.P.; Cifuentes, M.C.; Sandoval, T.A.; Pombo, L.M.; Castañeda, D.; Asea, A.; Fiorentino, S. A Petiveria Alliacea Standardized Fraction Induces Breast Adenocarcinoma Cell Death by Modulating Glycolytic Metabolism. J. Ethnopharmacol. 2014, 153, 641–649. [Google Scholar] [CrossRef]
- Murillo, N.; Lasso, P.; Urueña, C.; Pardo-Rodriguez, D.; Ballesteros-Ramírez, R.; Betancourt, G.; Rojas, L.; Cala, M.P.; Fiorentino, S. Petiveria Alliacea Reduces Tumor Burden and Metastasis and Regulates the Peripheral Immune Response in a Murine Myeloid Leukemia Model. Int. J. Mol. Sci. 2023, 24, 12972. [Google Scholar] [CrossRef]
- Vaghela, N.; Gohel, S. Medicinal Plant-Associated Rhizobacteria Enhance the Production of Pharmaceutically Important Bioactive Compounds under Abiotic Stress Conditions. J. Basic Microbiol. 2023, 63, 308–325. [Google Scholar] [CrossRef]
- Rizvi, A.; Ahmed, B.; Khan, M.S.; El-Beltagi, H.S.; Umar, S.; Lee, J. Bioprospecting Plant Growth Promoting Rhizobacteria for Enhancing the Biological Properties and Phytochemical Composition of Medicinally Important Crops. Molecules 2022, 27, 1407. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.-S.; Liu, J.-M.; Sun, J.; Huang, Y.-T.; Jin, N.; Li, M.-M.; Liang, Y.-T.; Fan, B.; Wang, F.-Z. Analysis of Endophytic Bacterial Diversity From Different Dendrobium Stems and Discovery of an Endophyte Produced Dendrobine-Type Sesquiterpenoid Alkaloids. Front. Microbiol. 2022, 12, 775665. [Google Scholar] [CrossRef] [PubMed]
- The Jamovi Project Jamovi. Available online: https://www.jamovi.org/ (accessed on 2 October 2023).
- Doyle, J.J.; Doyle, J. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global Patterns of 16S rRNA Diversity at a Depth of Millions of Sequences per Sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef]
- Gardes, M.; Bruns, T.D. ITS Primers with Enhanced Specificity for Basidiomycetes—Application to the Identification of Mycorrhizae and Rusts. Mol. Ecol. 1993, 2, 113–118. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, J.; Innis, M.; Gelfand, D.; Sninsky, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In Pcr Protocols: A Guide to Methods and Applications; Academic Press, Inc.: New York, NY, USA, 1990; Volume 31, pp. 315–322. [Google Scholar]
- Morgan, B.S.T.; Egerton-Warburton, L.M. Barcoded NS31/AML2 Primers for Sequencing of Arbuscular Mycorrhizal Communities in Environmental Samples. Appl. Plant Sci. 2017, 5, 1700017. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME Allows Analysis of High-Throughput Community Sequencing Data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-Resolution Sample Inference from Illumina Amplicon Data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef]
- Öpik, M.; Vanatoa, A.; Vanatoa, E.; Moora, M.; Davison, J.; Kalwij, J.M.; Reier, Ü.; Zobel, M. The Online Database MaarjAM Reveals Global and Ecosystemic Distribution Patterns in Arbuscular Mycorrhizal Fungi (Glomeromycota). New Phytol. 2010, 188, 223–241. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated Estimation of Fold Change and Dispersion for RNA-Seq Data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Fierer, N.; Jackson, R.B. The Diversity and Biogeography of Soil Bacterial Communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Shi, J.-Y.; Yuan, X.-F.; Lin, H.-R.; Yang, Y.-Q.; Li, Z.-Y. Differences in Soil Properties and Bacterial Communities between the Rhizosphere and Bulk Soil and among Different Production Areas of the Medicinal Plant Fritillaria Thunbergii. Int. J. Mol. Sci. 2011, 12, 3770–3785. [Google Scholar] [CrossRef] [PubMed]
- Uroz, S.; Buée, M.; Murat, C.; Frey-Klett, P.; Martin, F. Pyrosequencing Reveals a Contrasted Bacterial Diversity between Oak Rhizosphere and Surrounding Soil. Environ. Microbiol. Rep. 2010, 2, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Ni, T.; Xun, W.; Huang, X.; Huang, Q.; Ran, W.; Shen, B.; Zhang, R.; Shen, Q. Influence of Straw Incorporation with and without Straw Decomposer on Soil Bacterial Community Structure and Function in a Rice-Wheat Cropping System. Appl. Microbiol. Biotechnol. 2017, 101, 4761–4773. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.A.; Jackson, R.B. Toward an Ecological Classification of Soil Bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Ward, N.L.; Challacombe, J.F.; Janssen, P.H.; Henrissat, B.; Coutinho, P.M.; Wu, M.; Xie, G.; Haft, D.H.; Sait, M.; Badger, J.; et al. Three Genomes from the Phylum Acidobacteria Provide Insight into the Lifestyles of These Microorganisms in Soils. Appl. Environ. Microbiol. 2009, 75, 2046–2056. [Google Scholar] [CrossRef]
- Sait, M.; Davis, K.E.R.; Janssen, P.H. Effect of pH on Isolation and Distribution of Members of Subdivision 1 of the Phylum Acidobacteria Occurring in Soil. Appl. Environ. Microbiol. 2006, 72, 1852–1857. [Google Scholar] [CrossRef]
- Navarrete, A.A.; Kuramae, E.E.; de Hollander, M.; Pijl, A.S.; van Veen, J.A.; Tsai, S.M. Acidobacterial Community Responses to Agricultural Management of Soybean in Amazon Forest Soils. FEMS Microbiol. Ecol. 2013, 83, 607–621. [Google Scholar] [CrossRef]
- Hugoni, M.; Luis, P.; Guyonnet, J.; Haichar, F. el Z. Plant Host Habitat and Root Exudates Shape Fungal Diversity. Mycorrhiza 2018, 28, 451–463. [Google Scholar] [CrossRef]
- Zhang, N.; Castlebury, L.; Miller, A.; Huhndorf, S.M.; Schoch, C.; Seifert, K.; Rossman, A.; Rogers, J.; Kohlmeyer, J.; Volkmann-Kohlmeyer, B.; et al. An Overview of the Systematics of the Sordariomycetes Based on a Four-Gene Phylogeny. Mycologia 2006, 98, 1076–1087. [Google Scholar] [CrossRef] [PubMed]
- Buée, M.; De Boer, W.; Martin, F.; van Overbeek, L.; Jurkevitch, E. The Rhizosphere Zoo: An Overview of Plant-Associated Communities of Microorganisms, Including Phages, Bacteria, Archaea, and Fungi, and of Some of Their Structuring Factors. Plant Soil 2009, 321, 189–212. [Google Scholar] [CrossRef]
- Moll, J.; Hoppe, B.; König, S.; Wubet, T.; Buscot, F.; Krüger, D. Spatial Distribution of Fungal Communities in an Arable Soil. PLoS ONE 2016, 11, e0148130. [Google Scholar] [CrossRef] [PubMed]
- Baldrian, P.; Voříšková, J.; Dobiášová, P.; Merhautová, V.; Lisá, L.; Valášková, V. Production of Extracellular Enzymes and Degradation of Biopolymers by Saprotrophic Microfungi from the Upper Layers of Forest Soil. Plant Soil 2011, 338, 111–125. [Google Scholar] [CrossRef]
- Van der Wal, A.; Geydan, T.D.; Kuyper, T.W.; de Boer, W. A Thready Affair: Linking Fungal Diversity and Community Dynamics to Terrestrial Decomposition Processes. FEMS Microbiol. Rev. 2013, 37, 477–494. [Google Scholar] [CrossRef]
- Bhargav, V.; Kumar, A.; Dhiman, H.; Aggarwal, A. Biodiversity of Vesicular-Arbuscular Mycorrhiza in the Rhizosphere of Some Medicinal Plants of Kurukshetra University. Res. J. Biotechnol. 2023, 18, 24–30. [Google Scholar] [CrossRef]
- Peña-Venegas, C.P.; Sterling, A.; Andrade-Ramírez, T.K. Arbuscular Mycorrhization in Colombian and Introduced Rubber (Hevea brasiliensis) Genotypes Cultivated on Degraded Soils of the Amazon Region. Agriculture 2021, 11, 361. [Google Scholar] [CrossRef]
- Marinho, F.; Ramalho da Silva, I.; Oehl, F.; Maia, L. Checklist of Arbuscular Mycorrhizal Fungi in Tropical Forests. Sydowia 2018, 70, 107–127. [Google Scholar] [CrossRef]
- Zheng, Z.; Ma, X.; Zhang, Y.; Liu, Y.; Zhang, S. Soil Properties and Plant Community-Level Traits Mediate Arbuscular Mycorrhizal Fungal Response to Nitrogen Enrichment and Altered Precipitation. Appl. Soil Ecol. 2022, 169, 104245. [Google Scholar] [CrossRef]
- Mendes, R.; Garbeva, P.; Raaijmakers, J.M. The Rhizosphere Microbiome: Significance of Plant Beneficial, Plant Pathogenic, and Human Pathogenic Microorganisms. FEMS Microbiol. Rev. 2013, 37, 634–663. [Google Scholar] [CrossRef]
- Richardson, A.E.; Kawasaki, A.; Condron, L.M.; Ryan, P.R.; Gupta, V.V.S.R. Root Microbiome Structure and Microbial Succession in the Rhizosphere. In Rhizosphere Biology: Interactions between Microbes and Plants; Gupta, V.V.S.R., Sharma, A.K., Eds.; Rhizosphere Biology; Springer: Singapore, 2021; pp. 109–128. ISBN 9789811561252. [Google Scholar]
- Cui, Y.; Bing, H.; Fang, L.; Wu, Y.; Yu, J.; Shen, G.; Jiang, M.; Wang, X.; Zhang, X. Diversity Patterns of the Rhizosphere and Bulk Soil Microbial Communities along an Altitudinal Gradient in an Alpine Ecosystem of the Eastern Tibetan Plateau. Geoderma 2019, 338, 118–127. [Google Scholar] [CrossRef]
- Misra, P.; Maji, D.; Awasthi, A.; Pandey, S.S.; Yadav, A.; Pandey, A.; Saikia, D.; Babu, C.S.V.; Kalra, A. Vulnerability of Soil Microbiome to Monocropping of Medicinal and Aromatic Plants and Its Restoration through Intercropping and Organic Amendments. Front. Microbiol. 2019, 10, 2604. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vélez-Martínez, G.A.; Duque-Zapata, J.D.; Reyes-Ardila, W.L.; Muñoz Flórez, J.E.; Díaz Gallo, S.A.; Díaz Ariza, L.A.; López-Álvarez, D. Comparative Analysis of Bacteria, Fungi, and Arbuscular Mycorrhizal Fungi in Medicinal Plants Lippia alba and Petiveria alliacea in Colombia. Diversity 2023, 15, 1167. https://doi.org/10.3390/d15121167
Vélez-Martínez GA, Duque-Zapata JD, Reyes-Ardila WL, Muñoz Flórez JE, Díaz Gallo SA, Díaz Ariza LA, López-Álvarez D. Comparative Analysis of Bacteria, Fungi, and Arbuscular Mycorrhizal Fungi in Medicinal Plants Lippia alba and Petiveria alliacea in Colombia. Diversity. 2023; 15(12):1167. https://doi.org/10.3390/d15121167
Chicago/Turabian StyleVélez-Martínez, Glever Alexander, Juan Diego Duque-Zapata, Wendy Lorena Reyes-Ardila, Jaime Eduardo Muñoz Flórez, Sergio Alberto Díaz Gallo, Lucia Ana Díaz Ariza, and Diana López-Álvarez. 2023. "Comparative Analysis of Bacteria, Fungi, and Arbuscular Mycorrhizal Fungi in Medicinal Plants Lippia alba and Petiveria alliacea in Colombia" Diversity 15, no. 12: 1167. https://doi.org/10.3390/d15121167
APA StyleVélez-Martínez, G. A., Duque-Zapata, J. D., Reyes-Ardila, W. L., Muñoz Flórez, J. E., Díaz Gallo, S. A., Díaz Ariza, L. A., & López-Álvarez, D. (2023). Comparative Analysis of Bacteria, Fungi, and Arbuscular Mycorrhizal Fungi in Medicinal Plants Lippia alba and Petiveria alliacea in Colombia. Diversity, 15(12), 1167. https://doi.org/10.3390/d15121167