
Two New Nematode Species, Desmolaimus magnus sp. nov. (Monhysterida, Linhomoeidae) and Metadesmolaimus robustus sp. nov. (Monhysterida, Xyalidae), from the Yellow Sea, China with Phylogenetic Analyses within Linhomoeidae and Xyalidae †


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Nematode Identification
2.2. DNA Extraction, PCR Amplification, and Phylogenetic Analysis
3. Results and Discussion
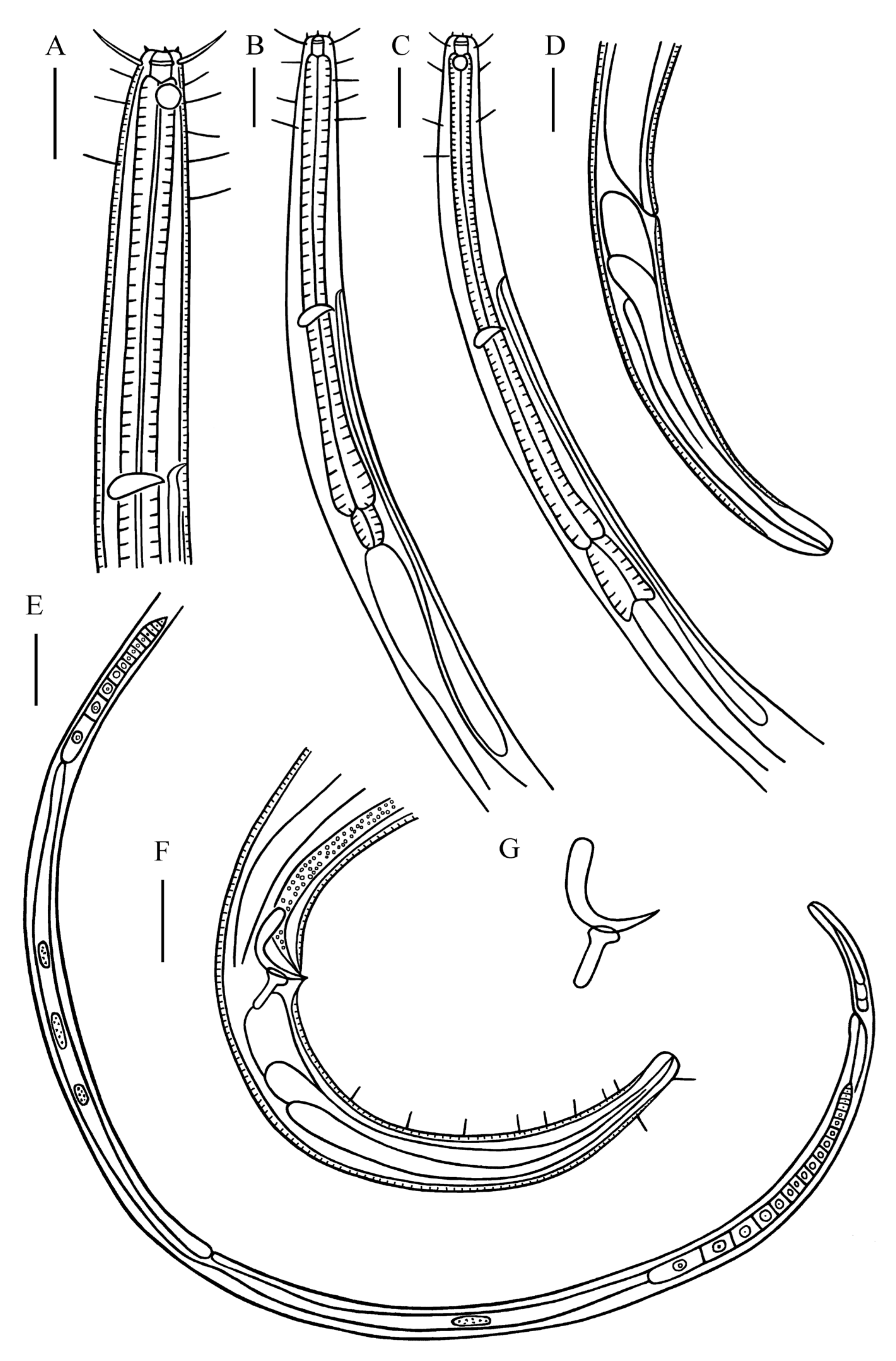
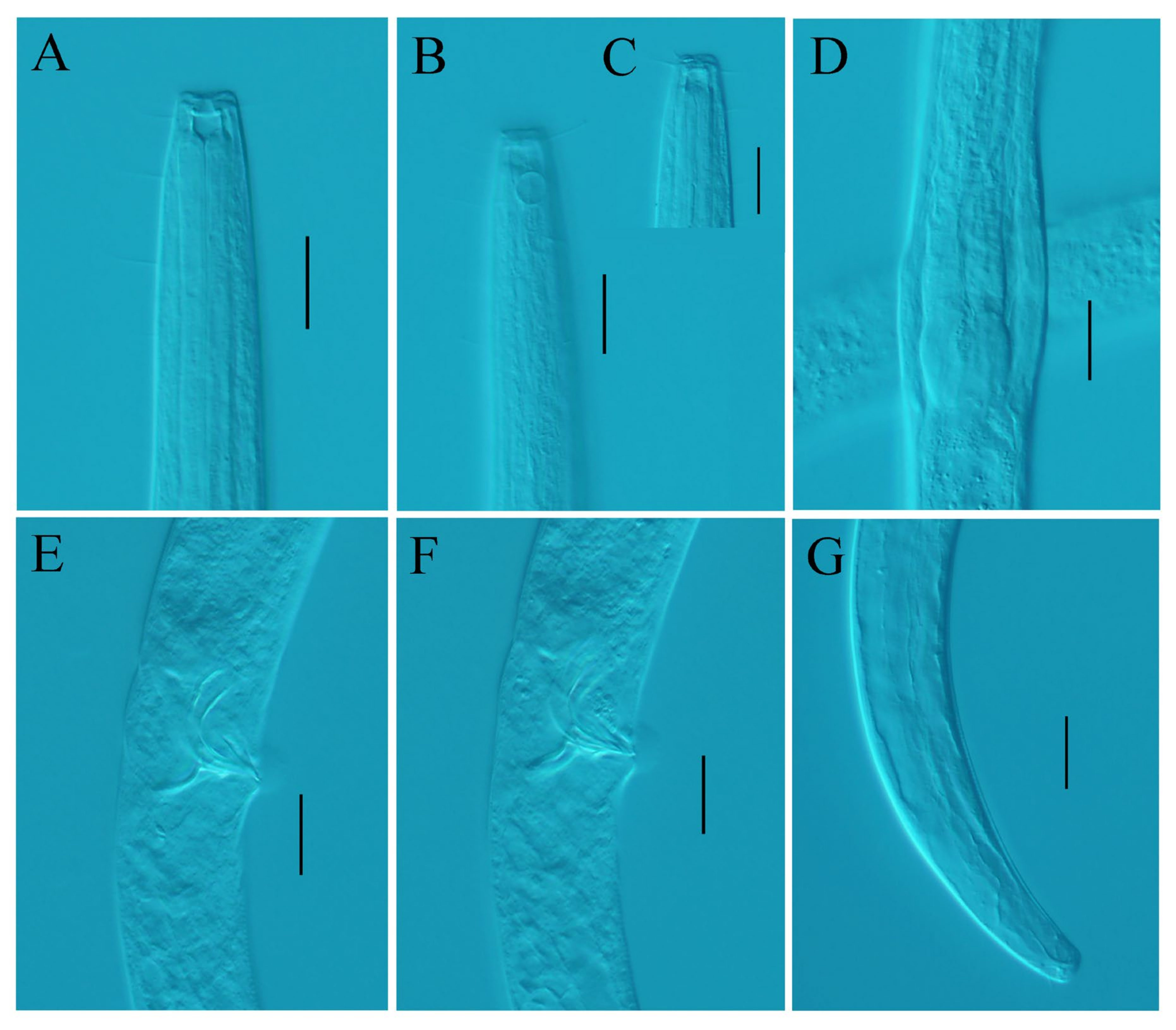
3.1. Taxonomy of Desmolaimus magnus sp. nov.
3.1.1. Description
3.1.2. Differential Diagnosis and Discussion
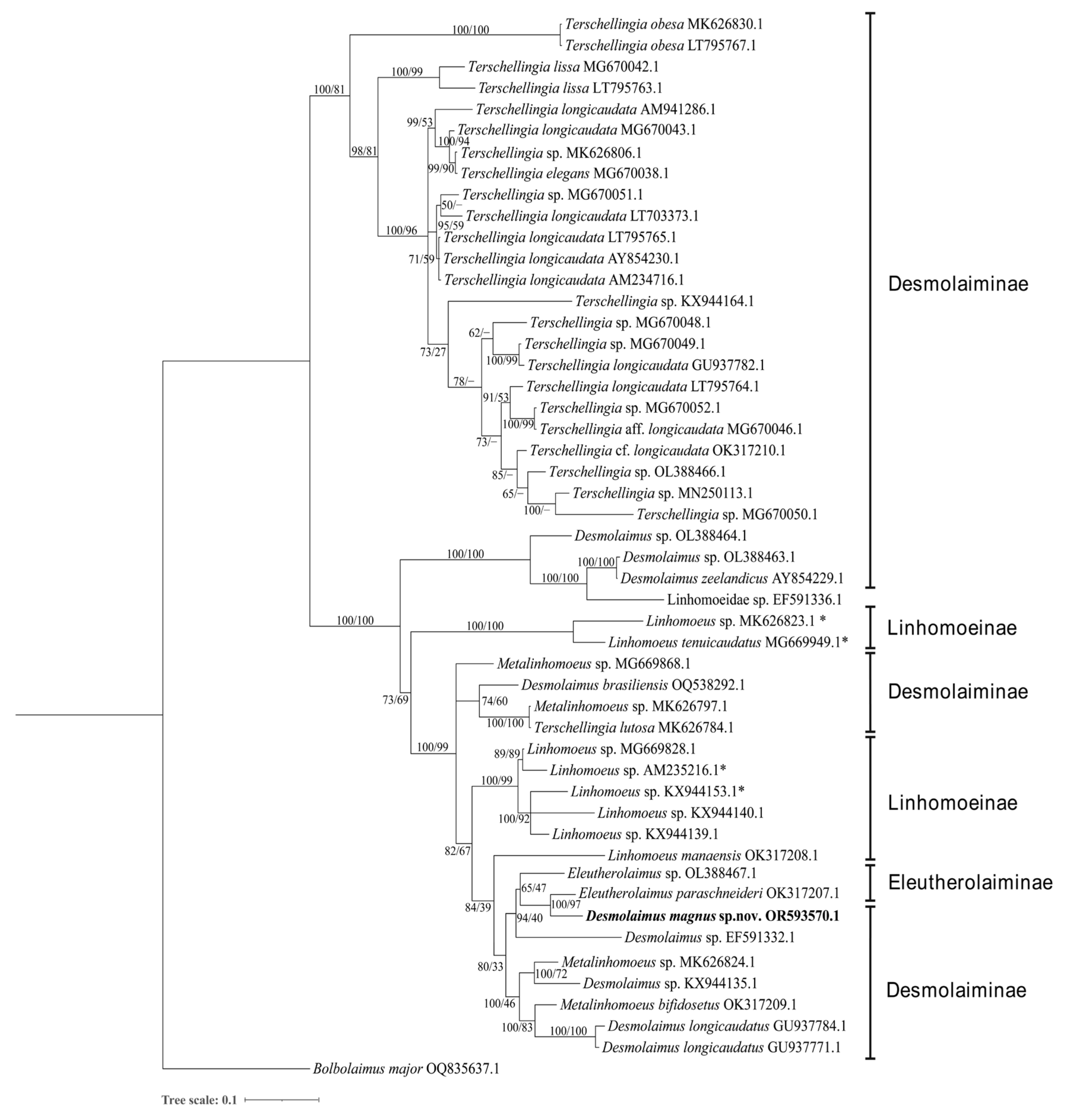
3.1.3. Molecular Phylogenetic Analysis
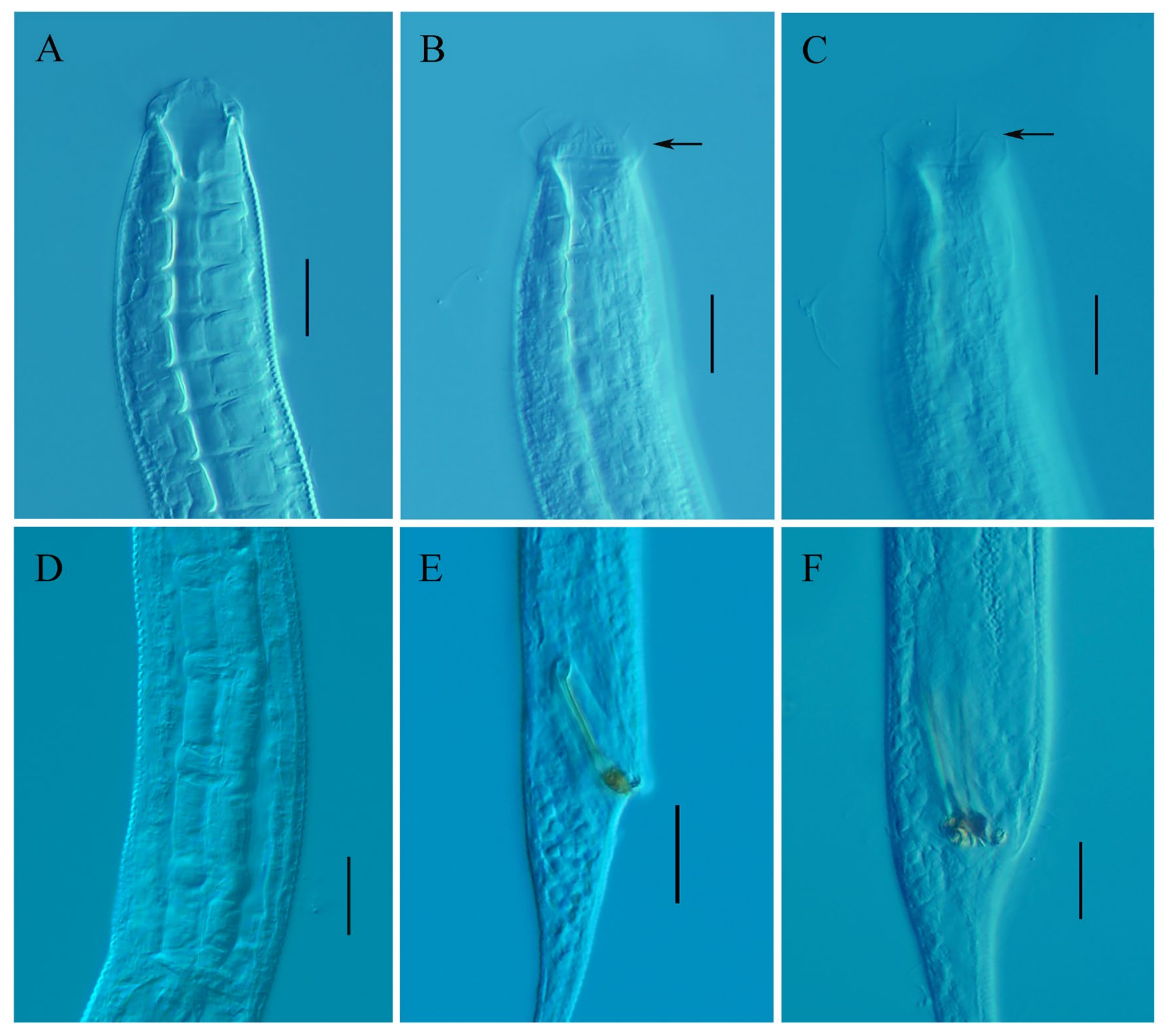
3.2. Taxonomy of Metadesmolaimus robustus sp. nov.
3.2.1. Description
3.2.2. Differential Diagnosis and Discussion
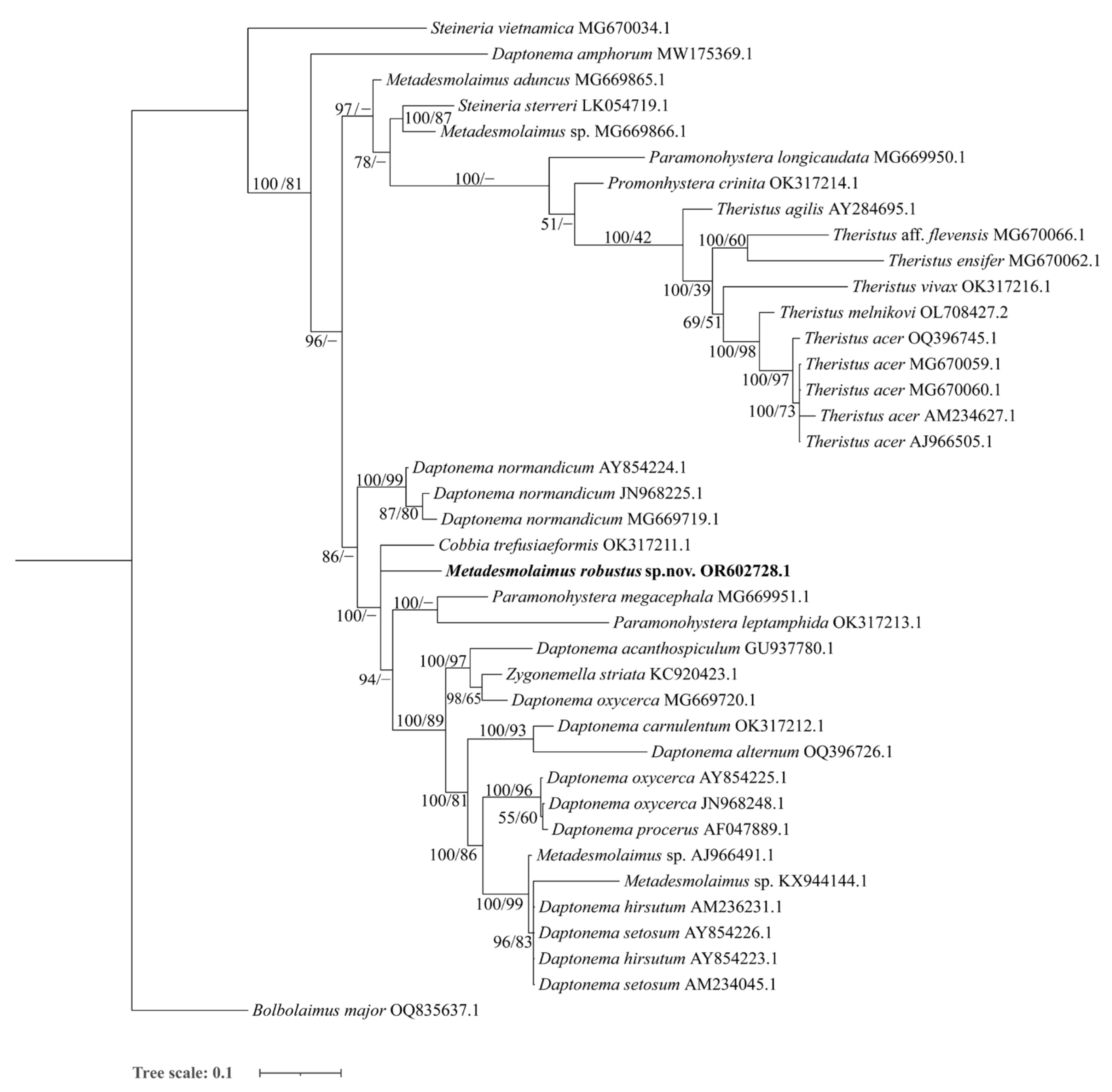
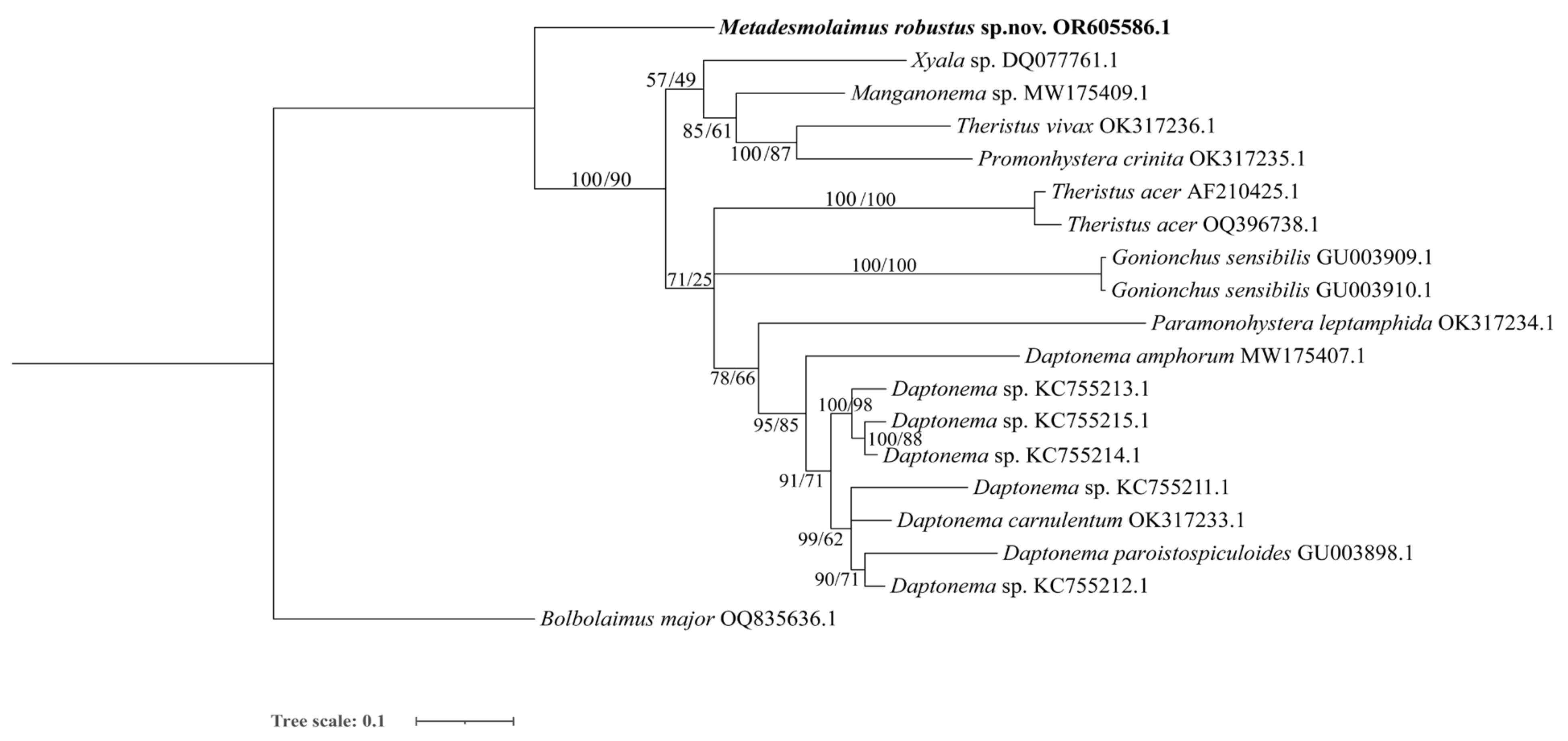
3.2.3. Molecular Phylogenetic Relationships and Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hodda, M. Phylum Nematoda: A classification, catalogue and index of valid genera, with a census of valid species. Zootaxa 2022, 5114, 1–289. [Google Scholar] [CrossRef] [PubMed]
- Lorenzen, S. Die Nematodenfauna im Verklappungsgebiet für Industrieabwässer nordwestlich von Helgoland: I. Araeolaimida und Monhysterida. Zool. Anz. 1971, 187, 223–248. [Google Scholar]
- Tchesunov, A.V.; Miljutin, D.M. Three new free-living nematode species (Monhysterida) from the Arctic abyss, with revision of the genus Eleutherolaimus Filipjev, 1922 (Linhomoeidae). Russ. J. Nematol. 2006, 14, 57–75. [Google Scholar]
- Venekey, V.; Gheller, P.F.; Maria, T.F.; Brustolin, M.C.; Kandratavicius, N.; Vieira, D.C.; Brito, S.; Souza, G.S.; Fonseca, G. The state of the art of Xyalidae (Nematoda, Monhysterida) with reference to the Brazilian records. Mar. Biodivers. 2014, 44, 367–390. [Google Scholar] [CrossRef]
- De Man, J.G. Die einheimischen, frei in der reinen Erde und im süßen Wasser lebende Nematoden monographisch bearbeitet. Vorläufiger Bericht und descriptiv-systematischer Theil. Tijdschr. Ned. Dierkd. Ver. 1880, 5, 1–104. [Google Scholar]
- Kreis, H.A. Freilebende marine Nematoden von der Nordwest-Küste Frankreichs (Trébeurden Côtes du Nord). Capita Zool. 1929, 2, 1–98. [Google Scholar]
- Schneider, W. Neue freilebende Nematoden aus Höhlen und Brunnen. I. Nematoden aus Jugoslawischen Höhlen. Zool. Anz. 1940, 132, 84–94. [Google Scholar]
- Gerlach, S.A. Über freilebende Meeresnematoden. Revision der Linhomoeidae. Zool. Jb. Syst. Bd. 1963, 90, 599–658. [Google Scholar]
- Leduc, D.; Gwyther, J. Description of new species of Setosabatieria and Desmolaimus (Nematoda: Monhysterida) and a checklist of New Zealand free-living marine nematode species. New Zealand J. Mar. Freshw. Res. 2008, 42, 339–362. [Google Scholar] [CrossRef]
- Gagarin, V.G. Two New Species of Free-Living Nematodes of the Family Linhomoeidae (Nematoda, Monhysterida) from Mangrove Forests in Vietnam. Inland Water Biol. 2019, 12, 21–31. [Google Scholar] [CrossRef]
- Schuurmans Stekhoven, J.H. Additional notes to my monographs on the freeliving marine Nemas of the Belgian Coast. I. and II. Mémoires Musée Royal D’histoire Naturelle Belg. 1935, 72, 1–36. [Google Scholar]
- Gerlach, S.A. Die Nematodenbesiedlung des Sandstrandes und des Küstengrundwassers an der italienischen Küste I. Systematischer Teil. Archo. Zool. Ital. 1953, 37, 517–640. [Google Scholar]
- Gerlach, S.A.; Riemann, F. The Bremerhaven Checklist of Aquatic Nematodes. A Catalogue of Nematoda Adenophorea excluding the Dorylaimida. Veröff. Inst. Meeresforsch. Bremerhav. 1973, 4, 1–736. [Google Scholar]
- Lorenzen, S. Revision det Xyalidae (freilebende Nematoden) auf der Grundlage einer kritischen Analyse von 56 Arten aus Nord- und Ostsee. Veroff. Inst. Meeresforsch. Bremerh. 1977, 16, 197–261. [Google Scholar]
- Guo, Y.Q.; Chen, Y.Z.; Liu, M.D. Metadesmolaimus zhanggi sp. nov. (Nematoda: Xyalidae) from East China Sea, with a pictorial key to Metadesmolaimus species. Cah. Biol. Mar. 2016, 5, 73–79. [Google Scholar]
- McIntyre, A.D.; Warwick, R.M. Meiofauna techniques. In Methods for the Study of Marine Benthos; Holme, N.A., McIntyre, A.D., Eds.; Blackwell Scientific Publications: Oxford, UK, 1984; pp. 217–244. [Google Scholar]
- Blaxter, M.L.; De Ley, P.; Garey, J.R.; Liu, L.X.; Scheldemann, P.; Vierstraete, A.; VanXeteren, J.R.; Mackey, L.Y.; Dorris, M.; Frisse, L.M.; et al. A molecular evolutionary framework for the phylum Nematoda. Nature 1998, 392, 71–75. [Google Scholar] [CrossRef] [PubMed]
- Nunn, G.B. Nematode Molecular Evolution. Ph.D. Thesis, University of Nottingham, Nottingham, UK, 1992. [Google Scholar]
- Zhao, Z.; Li, D.; Davies, K.A.; Ye, W. Schistonchus zealandicus n. sp. (Nematoda: Aphelenchoididae) associated with Ficus macrophylla in New Zealand. Nematology 2015, 17, 53–66. [Google Scholar] [CrossRef]
- Allgén, C.A. Freilebende Nematoden aus dem Trondhjemsfjord. Capita Zool. 1933, 4, 1–162. [Google Scholar]
- Leduc, D.; Zhao, Z.Q. The Marine Biota of Aotearoa New Zealand. Ngâ toke o Parumoana: Common free-living Nematoda of Pâuatahanui Inlet, Te-Awarua-o-Porirua Harbour, Wellington. NIWA Biodivers. Mem. 2023, 135, 1–212. [Google Scholar]
- Sahraean, N.; Van Campenhout, J.; Rigaux, A.; Mosallanejad, H.; Leliaert, F.; Moens, T. Lack of population genetic structure in the marine nematodes Ptycholaimellus pandispiculatus and Terschellingia longicaudata in beaches of the Persian Gulf. Mar. Ecol. 2017, 38, e12426. [Google Scholar] [CrossRef]
- Gagarin, V.G. Two new species of the genus Metadesmolaimus Schuurmans Stekhoven, 1935 (Nematoda, Monhysterida) from Goast of Vietnam. Int. J. Nematol. 2013, 23, 119–128. [Google Scholar]
- Gerlach, S.A. Brasilianische Meeres-Nematoden 1. (Ergebnisse eines Studienaufenthaltes na der Universitãt São Paulo). Bol. Do Inst. Ocean. 1956, 5, 3–69. [Google Scholar] [CrossRef]
- Gagarin, V.G. Daptonema obesum sp.n. and Steineria vietnamica sp.n. (Nematoda, Xyalidae) from mangrove forest of Vietnam. Int. J. Nematol. 2013, 23, 129–137. [Google Scholar]
- Neres, P.F.; Fonseca-Genevois, V.G.; Torres, R.A.; Fonseca Cavalcanti, M.; Castro, F.J.V.; Silva, N.R.R.; Rieger, T.T.; Decraemer, W. Morphological and molecular taxonomy of a new Daptonema (Nematoda, Xyalidae) with comments on the systematics of some related taxa. Zool. J. Linnaean Soc. 2010, 158, 1–15. [Google Scholar] [CrossRef]
- Leduc, D.; Zhao, Z.Q. Molecular characterization of free-living nematodes from Kermadec Trench (Nematoda: Aegialoalaimidae, Xyalidae) with description of Aegialoalaimus tereticaudan sp. Zootaxa 2021, 4949, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Cunha, B.P.; Brito, S.; Fonseca, G. Zygonemella: The forgotten genus of the family Xyalidae (Nematoda). Zootaxa 2013, 3669, 179–183. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters | Holotype | Paratypes | Paratypes |
|---|---|---|---|
| Male | Males (n = 4) | Females (n = 4) | |
| Total body length | 4724 | 5373 ± 174 (5116–5500) | 5521 ± 357 (5008–5833) |
| Maximum body diameter | 36 | 38 ± 2 (35–40) | 43 ± 4 (40–49) |
| Head diameter | 15 | 15 ± 1 (14–15) | 15 ± 1 (14–15) |
| Length of outer labial sensilla | 3 | 3 (3–3) | 3 (3–3) |
| Length of cephalic setae | 13 | 13 (13–13) | 12 ± 1 (12–13) |
| Length of subcephalic setae | 11 | 12 ± 1 (11–12) | 12 ± 2 (11–14) |
| Amphidial fovea width | 8 | 8 (8–8) | 8 (8–8) |
| Amphidial fovea from anterior end | 11 | 10 ± 1 (9–11) | 10 ± 1 (9–10) |
| Body diameter at amphidial fovea | 18 | 18 ± 1 (17–18) | 18 (18–18) |
| Nerve ring from anterior end | 147 | 163 ± 10 (150–172) | 160 ± 13 (151–178) |
| Body diameter at nerve ring | 31 | 33 ± 1 (31–34) | 31 ± 2 (29–34) |
| Pharynx length | 266 | 293 ± 13 (277–304) | 289 ± 14 (278–310) |
| Body diameter at base of pharynx | 36 | 36 ± 1 (35–37) | 32 ± 2 (30–35) |
| Anal body diameter | 33 | 37 ± 1 (35–38) | 30 ± 3 (27–33) |
| Spicules length along arc | 40 | 40 ± 1 (39–41) | - |
| Gubernaculum length | 18 | 18 ± 2 (16–20) | - |
| Vulva from anterior end | - | - | 3243 ± 251 (2910–3473) |
| Body diameter at vulva | - | - | 43 ± 4 (40–49) |
| V%, de Man’s ratio | - | - | 58.7 ± 1.4 (57.1–60.1) |
| Tail length | 225 | 250 ± 7 (241–256) | 232 ± 8 (223–239) |
| a, de Man’s ratio | 131.2 | 140.9 ± 10.8 (134.6–157.1) | 129.1 ± 13.1 (114.2–141.2) |
| b, de Man’s ratio | 17.8 | 18.4 ± 1.2 (16.9–19.6) | 19.1 ± 1.4 (17.8–20.3) |
| c, de Man’s ratio | 21.0 | 21.5 ± 1.1 (20.1–22.6) | 23.8 ± 1.3 (22.5–25.6) |
| c’, de Man’s ratio | 6.8 | 6.8 ± 0.4 (6.3–7.3) | 7.7 ± 0.6 (7.1–8.3) |
| Characters | Holotype | Paratypes | Paratype |
|---|---|---|---|
| Male | Males (n = 3) | Female (n = 1) | |
| Total body length | 2084 | 1546 ± 66 (1471–1593) | 1697 |
| Maximum body diameter | 43 | 32 ± 2 (31–34) | 52 |
| Head diameter | 25 | 22 ± 1 (21–23) | 26 |
| Length of inner labial sensilla | 7 | 6 (6–6) | 7 |
| Length of outer labial sensilla | 23 | 21 ± 1 (20–21) | 21 |
| Length of cephalic setae | 15 | 11 ± 1 (11–12) | 12 |
| Nerve ring from anterior end | 142 | 89 ± 1 (88–90) | 132 |
| Body diameter at nerve ring | 42 | 28 ± 2 (27–30) | 39 |
| Pharynx length | 420 | 312 ± 11 (303–324) | 416 |
| Body diameter at base of pharynx | 43 | 31 ± 1 (30–32) | 43 |
| Anal body diameter | 35 | 26 ± 1 (26–27) | 34 |
| Spicules length along arc | 52 | 41 ± 2 (39–42) | – |
| Gubernaculum length | 12 | 10 ± 1 (9–10) | – |
| Vulva from anterior end | – | – | 1233 |
| Body diameter at vulva | – | – | 51 |
| V%, de Man’s ratio | – | – | 72.66% |
| Tail length | 173 | 126 ± 7 (122–134) | 151 |
| a, de Man’s ratio | 48.5 | 47.9 ± 1.2 (46.9–49.2) | 32.6 |
| b, de Man’s ratio | 5.0 | 4.9 ± 0.3 (4.7–5.2) | 4.1 |
| c, de Man’s ratio | 12.0 | 12.3 ± 0.5 (11.9–12.9) | 11.2 |
| c’, de Man’s ratio | 4.9 | 4.8 ± 0.4 (4.5–5.2) | 4.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, W.; Meng, Z.; Wang, C. Two New Nematode Species, Desmolaimus magnus sp. nov. (Monhysterida, Linhomoeidae) and Metadesmolaimus robustus sp. nov. (Monhysterida, Xyalidae), from the Yellow Sea, China with Phylogenetic Analyses within Linhomoeidae and Xyalidae. Diversity 2023, 15, 1130. https://doi.org/10.3390/d15111130
Guo W, Meng Z, Wang C. Two New Nematode Species, Desmolaimus magnus sp. nov. (Monhysterida, Linhomoeidae) and Metadesmolaimus robustus sp. nov. (Monhysterida, Xyalidae), from the Yellow Sea, China with Phylogenetic Analyses within Linhomoeidae and Xyalidae. Diversity. 2023; 15(11):1130. https://doi.org/10.3390/d15111130
Chicago/Turabian StyleGuo, Wen, Zhiyu Meng, and Chunming Wang. 2023. "Two New Nematode Species, Desmolaimus magnus sp. nov. (Monhysterida, Linhomoeidae) and Metadesmolaimus robustus sp. nov. (Monhysterida, Xyalidae), from the Yellow Sea, China with Phylogenetic Analyses within Linhomoeidae and Xyalidae" Diversity 15, no. 11: 1130. https://doi.org/10.3390/d15111130
APA StyleGuo, W., Meng, Z., & Wang, C. (2023). Two New Nematode Species, Desmolaimus magnus sp. nov. (Monhysterida, Linhomoeidae) and Metadesmolaimus robustus sp. nov. (Monhysterida, Xyalidae), from the Yellow Sea, China with Phylogenetic Analyses within Linhomoeidae and Xyalidae. Diversity, 15(11), 1130. https://doi.org/10.3390/d15111130