



The Orchids of Wetland Vegetation in the Central Balkans

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Overview of the Orchid Flora of Wetland Vegetation in the Central Balkans
- species occurs exclusively in wetland vegetation types (at 100% of its sites, it is found in wetland vegetation types);
- species grows mainly in wetland vegetation types and rarely occurs in other vegetation types (at 50–100% of its sites, it is found in wetland vegetation types);
- species occurs in wetland vegetation types but mostly inhabits other vegetation types (at 10–50% of its sites, it is found in wetland vegetation types);
- species rarely occurs in wetland vegetation types and mostly inhabits other vegetation types (at < 10% of its sites, it is found in wetland vegetation types).
2.1. Richness of Orchid Taxa and Taxonomic Analysis
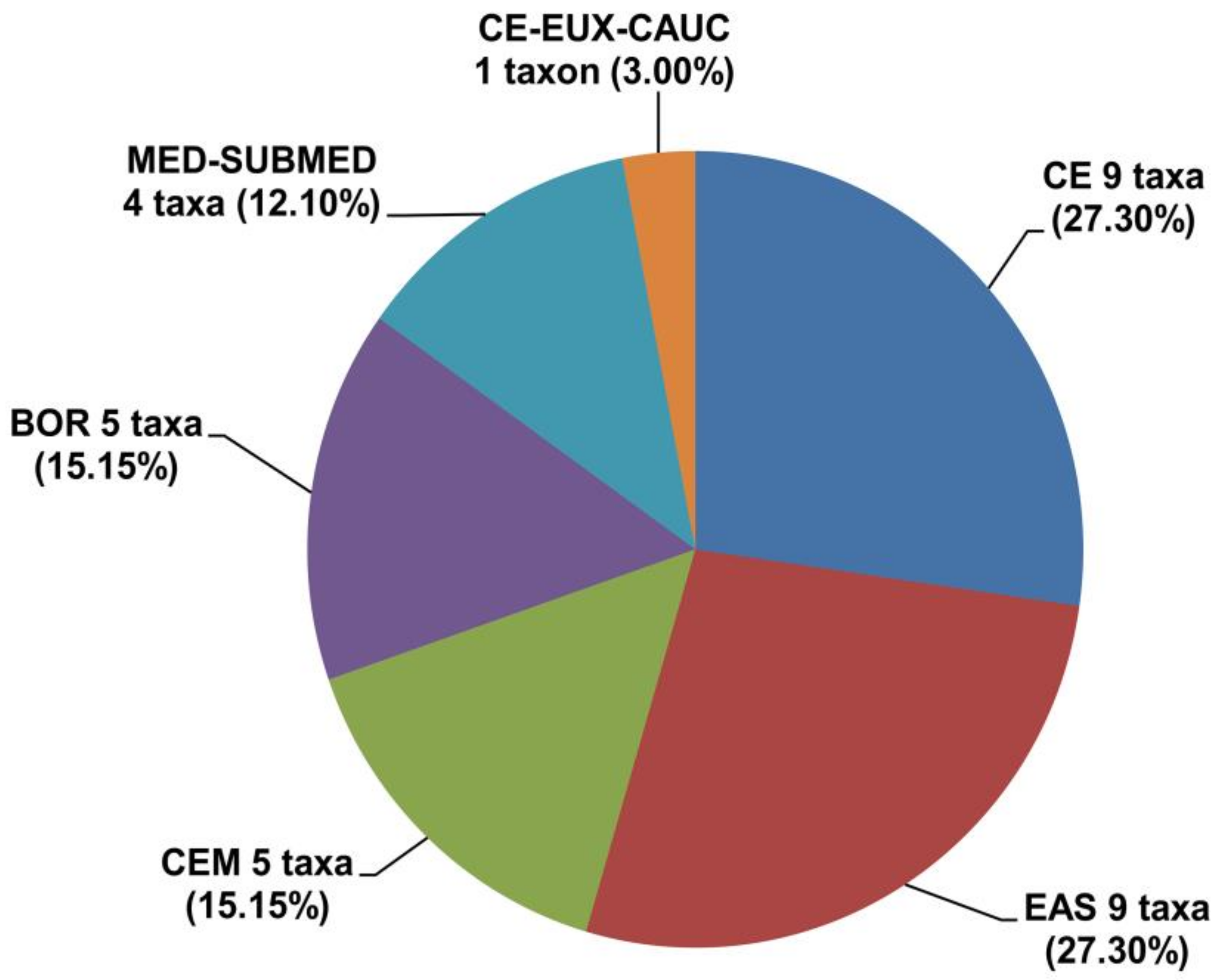
2.2. Phytogeographical Analysis
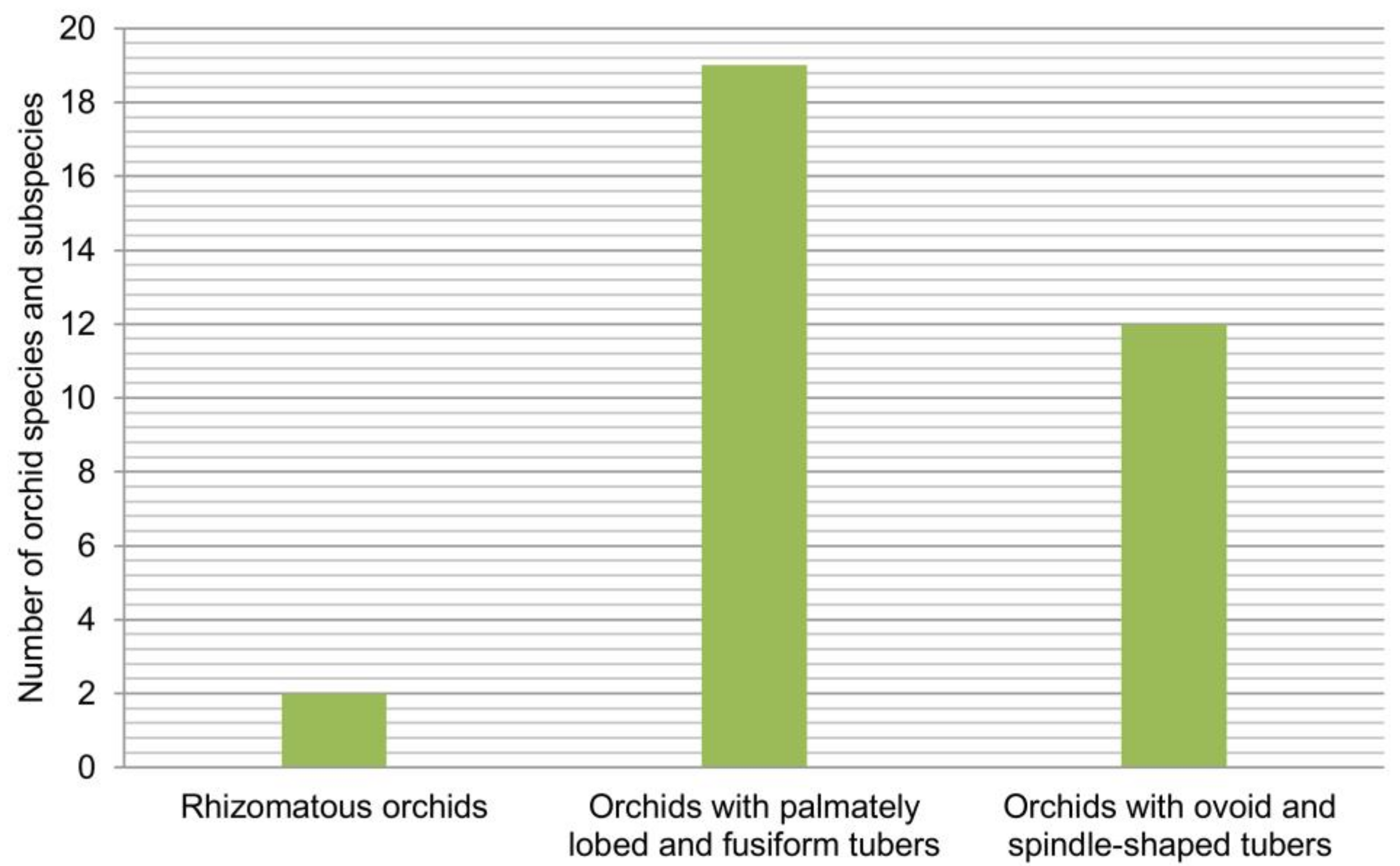
2.3. Life Forms
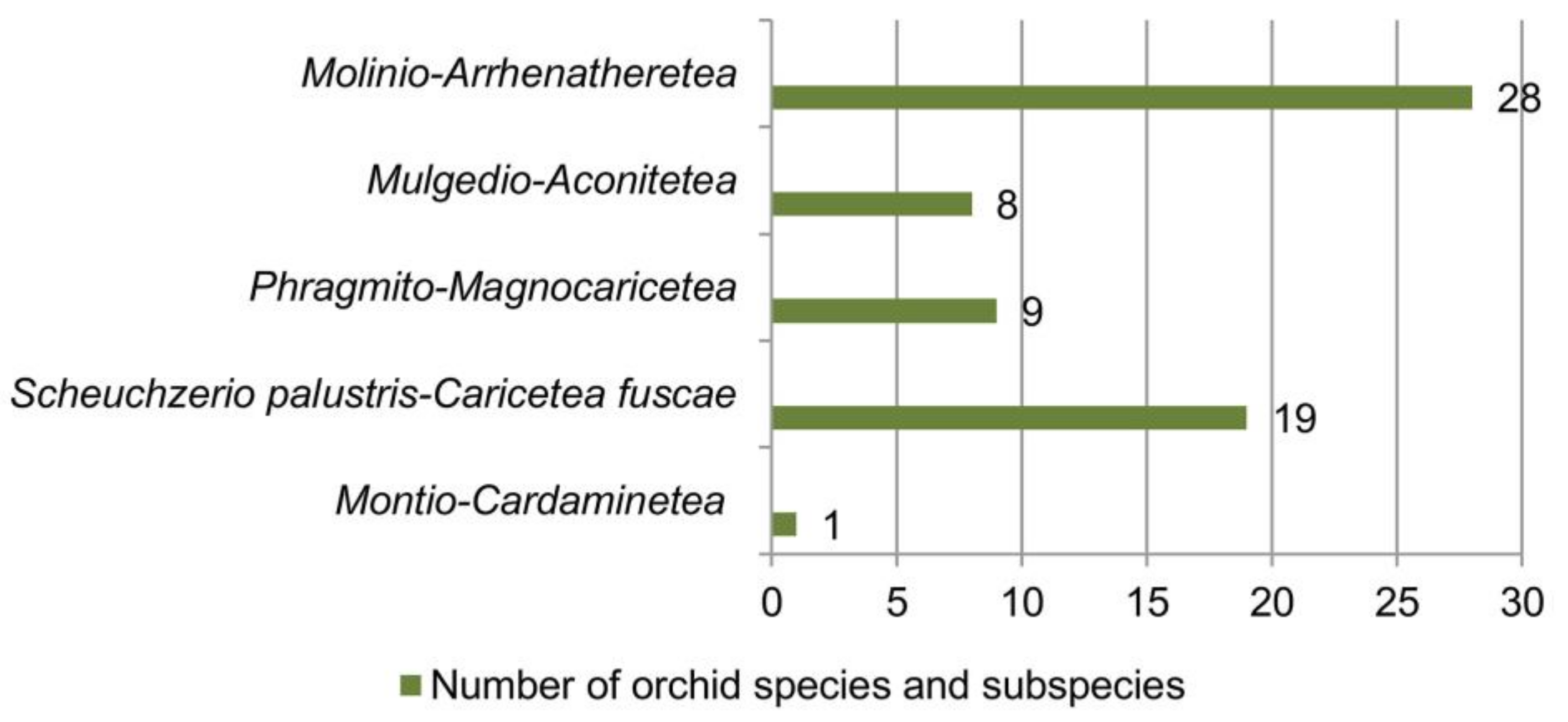
3. Wetland Vegetation
3.1. Wet Meadows
3.2. Tall-Herb Vegetation along Mountain Streams and Springs
3.3. Marshland Vegetation
3.4. Vegetation of Bogs and Fens
3.5. Vegetation of Springs
4. Geological Substrates
5. Threat Factors and Conservation Priorities
5.1. Threat Factors
5.2. Conservation Priorities
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Doucette, A.; Caro, G.G.; McDaniel, J.; Clements, M.A.; et al. Orchid historical biogeography, diversification, Antarctica and the paradox of orchid dispersal. J. Biogeogr. 2016, 43, 1905–1916. [Google Scholar] [CrossRef]
- Waterman, R.J.; Bidartondo, M.I. Deception above, deception below: Linking pollination and mycorrhizal biology of orchids. J. Exp. Bot. 2008, 59, 1085–1096. [Google Scholar] [CrossRef]
- Swarts, N.D.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef] [PubMed]
- Whigham, D.F.; Willems, J.H. Demographic studies and life-history strategies of temperate terrestrial orchids as a basis for conservation. In Orchid Conservation; Dixon, K.W., Kell, S.P., Barrett, R.L., Cribb, P.J., Eds.; Natural History Publications: Kota Kinabaluk, Malaysia, 2003; pp. 137–158. [Google Scholar]
- Hágsater, E.; Dumont, V. (Eds.) Orchids: Status, Survey and Conservation Action Plan; IUCN: Gland, Switzerland; Cambridge, UK, 1996. [Google Scholar]
- Delforge, P. Orchids of Europe, North Africa and the Middle East; A. & C. Black: London, UK, 2006. [Google Scholar]
- Djordjević, V.; Tsiftsis, S. The Role of Ecological Factors in Distribution and Abundance of Terrestrial Orchids. In Orchids Phytochemistry, Biology and Horticulture; Reference Series in Phytochemistry; Mérillon, J.-M., Kodja, H., Eds.; Springer Nature: Cham, Switzerland, 2022; pp. 3–72. [Google Scholar]
- Duffy, K.J.; Scopece, G.; Cozzolino, S.; Fay, M.F.; Smith, R.J.; Stout, J.C. Ecology and genetic diversity of the dense-flowered orchid, Neotinea maculata, at the centre and edge of its range. Ann. Bot. 2009, 104, 507–516. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.; Passalacqua, N.G.; Bartram, S.; Schatz, B.; Croce, A.; Carey, P.D.; Kraudelt, H.; Jeltsch, F. Conservation priorities differ at opposing species borders of a European orchid. Biol. Conserv. 2010, 143, 2207–2220. [Google Scholar] [CrossRef]
- Sagarin, R.D.; Gaines, S.D. The ‘abundant centre’ distribution: To what extent is it a biogeographical rule? Ecol. Lett. 2002, 5, 137–147. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Hutchings, M.J. Biological flora of the British Isles: Epipactis palustris. J. Ecol. 2014, 102, 1341–1355. [Google Scholar] [CrossRef]
- Jersáková, J.; Malinová, T.; Jeřábková, K.; Dötteri, S. Biological Flora of the British Isles: Pseudorchis albida (L.) Á. & D. Löve. J. Ecol. 2011, 99, 1282–1298. [Google Scholar]
- Meekers, T.; Hutchings, M.J.; Honnay, O.; Jacquemyn, H. Biological Flora of the British Isles: Gymnadenia conopsea s.l. J. Ecol. 2012, 100, 1269–1288. [Google Scholar] [CrossRef]
- Jersáková, J.; Traxmandlová, I.; Ipser, Z.; Matthias, K.; Pellegrino, G.; Schatz, B.; Djordjević, V.; Kindlmann, P.; Renner, S.S. Biological flora of Central Europe: Dactylorhiza sambucina (L.) Soó. Perspect. Plant Ecol. Evol. Syst. 2015, 17, 318–329. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Orchid species richness and composition in relation to vegetation types. Wulfenia 2020, 27, 183–210. [Google Scholar]
- Kirillova, I.A.; Dubrovskiy, Y.A.; Degteva, S.V.; Novakovskiy, A.B. Ecological and habitat ranges of orchids in the northernmost regions of their distribution areas: A case study from Ural Mountains, Russia. Plant Divers. 2022. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I.; Karagiannakidou, V.; Alifragis, D. Niche analysis and conservation of the orchids of east Macedonia (NE Greece). Acta Oecol. 2008, 33, 27–35. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Jovanović, S.; Stevanović, V. Factors affecting the distribution and abundance of orchids in grasslands and herbaceous wetlands. Syst. Biodivers. 2016, 14, 355–370. [Google Scholar] [CrossRef]
- Wotavová, K.; Balounová, Z.; Kindlmann, P. Factors affecting persistence of terrestrial orchids in wet meadows and implications for their conservation in a changing agricultural landscape. Biol. Conserv. 2004, 118, 271–279. [Google Scholar] [CrossRef]
- Janečková, P.; Wotavová, K.; Schödelbauerová, I.; Jersáková, J.; Kindlmann, P. Relative effects of management and environmental conditions on performance and survival of populations of a terrestrial orchid, Dactylorhiza majalis. Biol. Conserv. 2006, 129, 40–49. [Google Scholar] [CrossRef]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J.H. Does nectar reward affect rarity and extinction probabilities of orchid species? An assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 121, 257–263. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Munoz, F.; Richard, F.; Schatz, B. Recent declines and range changes of orchids in Western Europe (France, Belgium and Luxembourg). Biol. Conserv. 2015, 190, 133–141. [Google Scholar] [CrossRef]
- Kull, T.; Selgis, U.; Pecina, M.V.; Metsare, M.; Ilves, A.; Tali, K.; Shefferson, R.P. Factors influencing IUCN threat levels to orchids across Europe on the basis of national red lists. Ecol. Evol. 2016, 6, 6245–6265. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Štípková, Z.; Kindlmann, P. Role of way of life, latitude, elevation and climate on the richness and distribution of orchid species. Biodivers. Conserv. 2019, 28, 75–96. [Google Scholar] [CrossRef]
- Tsiftsis, S. The complex effect of heterogeneity and isolation in determining alpha and beta orchid diversity on islands in the Aegean archipelago. Syst. Biodivers. 2020, 18, 281–294. [Google Scholar] [CrossRef]
- Averyanov, L. A review of the genus Dactylorhiza. In Orchid Biology—Reviews and Perspectives; Arditti, J., Ed.; V. Timber Press Inc.: Portland, OR, USA, 1990; pp. 159–206. [Google Scholar]
- Lazarević, P. Fens of the class Scheuchzerio-Caricetea fuscae (Nordh. 1936) R. Tx. 1937. in Serbia—Floristic and Vegetation Characteristics, Threats and Protection. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 2016. (In Serbian). [Google Scholar]
- Aćić, S. Synecological and Phytocoenological Study of Grassland Vegetation of Serbia. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 2018. [Google Scholar]
- Kojić, M.; Popović, R.; Karadžić, B. Syntaxonomic Review of the Vegetation of Serbia; Institute for Biological Research Siniša Stanković: Belgrade, Serbia, 1998. (In Serbian) [Google Scholar]
- Aćić, S.; Šilc, U.; Lakušić, D.; Vukojičić, S.; Dajić-Stevanović, Z. Typification and correction of syntaxa from the class Molinio-Arrhenatheretea Tx. 1937 in Serbia. Hacquetia 2013, 12, 39–54. [Google Scholar] [CrossRef]
- Tomović, G.; Niketić, M.; Lakušić, D.; Ranđelović, V.; Stevanović, V. Balkan endemic plants in Central Serbia and Kosovo regions: Distribution patterns, ecological characteristics and centres of diversity. Bot. J. Linn. Soc. 2014, 176, 173–202. [Google Scholar] [CrossRef]
- Čarni, A.; Ćuk, M.; Zelnik, I.; Franjić, J.; Igić, R.; Ilić, M.; Krstonošić, D.; Vukov, D.; Škvorc, Z. Wet Meadow Plant Communities of the Alliance Trifolion pallidi on the Southeastern Margin of the Pannonian Plain. Water 2021, 13, 381. [Google Scholar] [CrossRef]
- Djordjević, V.; Niketić, M.; Stevanović, V. Orchids of Serbia: Taxonomy, Life Forms, Pollination Systems, and Phytogeographical Analysis. In Orchidaceae: Characteristics, Distribution and Taxonomy; Djordjević, V., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2021; pp. 57–163. [Google Scholar]
- Euro+Med. The Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. Available online: http://ww2.bgbm.org/EuroPlusMed/query.asp (accessed on 18 October 2022).
- Pillon, Y.; Fay, M.F.; Shipunov, A.B.; Chase, M.W. Species diversity versus phylogenetic diversity: A practical study in the taxonomically difficult genus Dactylorhiza (Orchidaceae). Biol. Conserv. 2006, 129, 4–13. [Google Scholar] [CrossRef]
- Djordjević, V.; Tsiftsis, S. Patterns of orchid species richness and composition in relation to geological substrates. Wulfenia 2019, 26, 1–21. [Google Scholar]
- Djordjević, V.; Jovanović, S.; Stevanović, V. Dactylorhiza fuchsii (Orchidaceae), a new species in the flora of Serbia. Arch. Biol. Sci. 2014, 66, 1227–1232. [Google Scholar] [CrossRef]
- Šabanović, E.; Djordjević, V.; Milanović, Đ.; Boškailo, A.; Šarić, Š.; Huseinović, S.; Randjelović, V. Checklist of the Orchidaceae of Bosnia and Herzegovina. Phyton-Ann. Rei Bot. 2021, 61, 83–95. [Google Scholar]
- Niketić, M.; Tomović, G.; Perić, R.; Zlatković, B.; Anačkov, G.; Djordjević, V.; Jogan, N.; Radak, B.; Duraki, Š.; Stanković, M.; et al. Material on the Annotated Checklist of Vascular Flora of Serbia. Nomenclatural, taxonomic and floristic notes I. Bull. Nat. Hist. Mus. Belgr. 2018, 11, 101–180. [Google Scholar] [CrossRef]
- Radak, B. Morphological Variability of Species of the Genus Anacamptis Rich. (Orchidoideae, Orchidaceae) in the Balkan Peninsula and the Pannonian Plain. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 2019; pp. 1–284. [Google Scholar]
- Djordjević, V. The Orchid Flora (Orchidaceae) of Western Serbia; Serbian Academy of Sciences and Art: Belgrade, Serbia, 2021; pp. 1–467. (In Serbian) [Google Scholar]
- Bartók, A.; Csergő, A.-M.; Balázs, Ö.; Hurdu, B.-I.; Jakab, G. A Gymnadenia frivaldii Hampe ex Griseb. Újrafelfedezése areája északihatárán (Keleti Kárpátok, Románia). Kitaibelia 2016, 21, 213–220. [Google Scholar] [CrossRef][Green Version]
- Djordjević, V.; Lakušić, D.; Jovanović, S.; Stevanović, V. Distribution and conservation status of some rare and threatened orchid taxa in the central Balkans and the southern part of the Pannonian Plain. Wulfenia 2017, 24, 143–162. [Google Scholar]
- Milanović, Đ.; Stupar, V.; Šabanović, E.; Djordjević, V.; Brujić, J.; Boškailo, A.; Ranđelović, V. On the distribution and conservation status of some rare orchid taxa (Orchidaceae) in Bosnia and Herzegovina (Western Balkans). Hacquetia 2022, 21, 327–346. [Google Scholar] [CrossRef]
- Kotilínek, M.; Těšitelová, T.; Jersáková, J. Biological Flora of the British Isles: Neottia ovata. J. Ecol. 2015, 103, 1354–1366. [Google Scholar] [CrossRef]
- Meusel, H.; Jäger, E.; Weinert, E. Comparative Chorology of Central European Flora 1 (Vergleichende Chorologie der Zentraleuropäischen Flora 1); Gustav Fischer: Jena, Germany, 1965. [Google Scholar]
- Meusel, H.; Jäger, E.; Weinert, E. Comparative Chorology of Central European flora 2 (Vergleichende Chorologie der Zentraleuropäischen Flora 2); Gustav Fischer: Jena, Germany, 1978. [Google Scholar]
- Meusel, H.; Jäger, E. Vergleichende Chorologie der Zentraleuropäischen Flora 3; Gustav Fischer: Stuttgart, Germany, 1992. [Google Scholar]
- Stevanović, V. Floristic division of the territory of Serbia with an overview of higher chorion and appropriate floral elements. In The Flora of Serbia I; Sarić, M.R., Ed.; Serbian Academy of Sciences and Arts: Belgrade, Serbia, 1992; pp. 49–65. [Google Scholar]
- Tsiftsis, S.; Antonopoulos, Z. Atlas of the Greek Orchids; Mediterraneo Editions: Rethymno, Greece, 2017; Volume 1. [Google Scholar]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geograph; Clarendon: Oxford, UK, 1934. [Google Scholar]
- Ellenberg, H.; Mueller-Dambois, D. A key to Raunkiaer plant life froms with revised subdivisions. Ber. Des Geobot. Inst. Der Eidg. Techn. Hochsch. Stift. Rübel 1967, 37, 56–73. [Google Scholar]
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. Distribution of Orchids with Different Rooting Systems in the Czech Republic. Plants 2021, 10, 632. [Google Scholar] [CrossRef] [PubMed]
- Djordjević, V.; Tsiftsis, S.; Kindlmann, P.; Stevanović, V. Orchid diversity along an altitudinal gradient in the central Balkans. Front. Ecol. Evol. 2022, 10, 929266. [Google Scholar] [CrossRef]
- Bernardos, S.; García-Barriuso, M.; Sánchez-Anta, M.A.; Amich, F. Composition, geographical affinities and endemism of the Iberian Peninsula orchid flora. Nord. J. Bot. 2007, 25, 227–237. [Google Scholar] [CrossRef]
- Vakhrameeva, M.G.; Tatarenko, I.V.; Varlygina, T.I.; Torosyan, G.K.; Zagulski, M.N. Orchids of Russia and Adjacent Countries (Within the Borders of the Former USSR); Gantner Verlag: Ruggell, Liechtenstein, 2008. [Google Scholar]
- Mucina, L.; Bültmann, H.; Dierßen, K.; Theurillat, J.-P.; Raus, T.; Čarni, A.; Šumberová, K.; Willner, W.; Dengler, J.; Gavilán García, R.; et al. Vegetation of Europe: Hierarchical floristic classification system of plant, lichen, and algal communities. Appl. Veg. Sci. 2016, 19, 3–264. [Google Scholar] [CrossRef]
- Peterka, T.; Hájek, M.; Jiroušek, M.; Jiménez-Alfaro, B.; Aunina, L.; Bergamini, A.; Dítě, D.; Felbaba-Klushyna, L.; Graf, U.; Hájková, P.; et al. Formalized classification of European fen vegetation at the alliance level. Appl. Veg. Sci. 2017, 20, 124–142. [Google Scholar] [CrossRef]
- Mišić, V.; Jovanović-Dunjić, R.; Popović, M.; Borisavljević, L.; Antić, M.; Dinić, A.; Danon, J.; Blaženčić, Ž. Plant Communities and Habitats of Stara Planina; Serbian Academy of Sciences and Arts: Belgrade, Serbia, 1978; pp. 1–389. [Google Scholar]
- Petković, B. Marsh vegetation in the area of Tutin. Bull. Inst. Jard. Bot. Univ. Belgrade 1983, 17, 61–102. (In Serbian) [Google Scholar]
- Ranđelović, V.; Zlatković, B. Vegetation of the alliance Calthion in southeastern Serbia. Ekologija 1994, 28–29, 19–31. (In Serbian) [Google Scholar]
- Ranđelović, V.; Zlatković, B. Flora and Vegetation of the Vlasina Plateau; Faculty of Science, University of Niš: Niš, Serbia, 2010; pp. 1–448. (In Serbian) [Google Scholar]
- Košanin, N. Vegetation of Jakupica mountain in Macedonia. Glas. Srp. Kralj. Akad. 1911, 85, 184–242. (In Serbian) [Google Scholar]
- Pavlović, Z. Vegetation of the Mountain Zlatibor. Zb. Rad. Inst. Ekol. I Biogeogr. SANU 1951, 2, 115–182. (In Serbian) [Google Scholar]
- Stjepanović-Veseličić, L. Vegetation of Deliblato Sands. Inst. Ecol. Biogeogr. SANU 1953, 4, 1–113. (In Serbian) [Google Scholar]
- Tatić, B.; Veljović, V.; Petković, B.; Stefanović, M.; Radotić, S. Ass. Lathyreto-Molinietum Coerulae—A new community of meadow vegetation from the Pešter plateau—southwestern Serbia. Bull. Inst. Jard. Bot. Univ. Belgrade 1988, 12, 31–38. [Google Scholar]
- Butorac, B.; Hulo, I. Phytocoenological, floristic and ornithological values of the area Selevenjska pustara as the basis for protection. Zaštita Prir. 1992, 45, 65–76. (In Serbian) [Google Scholar]
- Butorac, B.; Hulo, I. Contribution to Knowledge of Marsch Meadows around the Kereš River; Book of Abstracts of XXIV Tiszakutató Ankét; Szegedi Ökologógiai Napok: Segedin, Hungary, 1993; p. 6. [Google Scholar]
- Lazarević, P. Floristic-Ecological Study of the Peštersko Polje Mire in Southwestern Serbia; University of Belgrade: Belgrade, Serbia, 2009. (In Serbian) [Google Scholar]
- Šabanović, E. Family Orchidaceae in the flora of Bosnia and Herzegovina. Ph.D. Thesis, University of Tuzla, Tuzla, Bosnia and Herzegovina, 2022; pp. 1–301. (In Bosnian). [Google Scholar]
- Babić, N. Lowland meadows in Podunavlje. Rad Vojvođanskih Muz. 1955, 4, 155–156. (In Serbian) [Google Scholar]
- Cincović, T. Types of meadows in Posavina. Proc. Fac. Agric. Belgrade 1956, 4, 1–25. (In Serbian) [Google Scholar]
- Ranđelović, N. Phytocenological-Ecological Characteristics of the Mountain Grasslands of Southeastern Serbia. Ph.D. Thesis, University of Zagreb, Zagreb, Croatia, 1978. (In Serbian). [Google Scholar]
- Ranđelović, V. Flora and Vegetation of the Vlasin Plateau. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 2002. (In Serbian). [Google Scholar]
- Radak, B.; Hristovski, S.; Matevski, V.; Anačkov, G. New orchid taxa for North Macedonia. In Proceedings of the 6th Congress of Ecologists of the Republic of North Macedonia; Ohrid, North Macedonia, 15–18 October 2022, Macedonian Ecological Society: Ohrid, North Macedonia, 2022; p. 82. [Google Scholar]
- Cincović, T. Meadow Vegetation in River Valleys of Western Serbia. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 1959; pp. 1–62. (In Serbian). [Google Scholar]
- Parabućski, S.; Stojanović, S. Oenanthe (banatica)-Alopecuretum pratensis ass. nova. Matica Srp. J. Nat. Sci. 1988, 74, 71–78. (In Serbian) [Google Scholar]
- Gajić, M.; Karadžić, D. Flora of flat Srem with Special Reference to Obedska Bara; Šumarski Fakultet: Belgrade, Serbia; Šumsko Gazdinstvo: Sremska Mitrovica, Serbia, 1991; pp. 1–437. [Google Scholar]
- Micevski, K. Typological classification of lowland meadow and swamp vegetation in Macedonia. Folia Balc. 1957, 1, 29–33. [Google Scholar]
- Jovanović, R. Types of valley meadows of Jasenica. Arch. Biol. Sci. 1957, 9, 1–14. (In Serbian) [Google Scholar]
- Jovanović-Dunjić, R. Typology, Ecology and Dynamics of Swamp and Meadow Vegetation in the Velika Morava Valley. Ph.D. Thesis, University of Belgrade, Belgrade, Serbia, 1965; pp. 1–399. (In Serbian). [Google Scholar]
- Jovanović, V. Kukavica Mountain in southeastern Serbia and the vegetation of its northern part. Leskovački Zb. 1977, 17, 271–299. (In Serbian) [Google Scholar]
- Ranđelović, N.; Rexhepi, F.; Jovanović, V. Plant communities of Southeast Kosovo. In Proceedings of the 2nd Congress of Ecologists of Yugoslavia, Zadar, Plitvice, Yugoslavia, 1–7 October 1979; pp. 957–995. [Google Scholar]
- Jovanović, V. Meadow Vegetation of Southeastern Serbia—Mount Radan, Goljak, Part of Kukavica and Their Surroundings. Ph.D. Thesis, University of Novi Sad, Novi Sad, Serbia, 1979. (In Serbian). [Google Scholar]
- Hundozi, B. Vegetation of Lowland Meadows in Kosovo. Ph.D. Thesis, University of Zagreb, Zagreb, Croatia, 1980; pp. 1–173. (In Serbian). [Google Scholar]
- Jovanović-Dunjić, R.; Stefanović, K.; Popović, R.; Dimitrijević, J. A contribution to the knowledge of meadow ecosystems in the Veliki Jastrebec area. Bull. Inst. Jard. Bot. Univ. Belgrade 1986, 20, 7–31. (In Serbian) [Google Scholar]
- Milanović, Đ.; (Faculty of Forestry, University of Banja Luka, Banja Luka, Bosnia and Herzegovina). Personal communication, 2022.
- Danon, J. Phytocoenological Investigations of Meadows in the Vicinity of Krivi Vir with Special Reference to Nutritional Value of the Hay; Faculty of Natural Sciences, University in Belgrade: Belgrade, Serbia, 1960. (In Serbian) [Google Scholar]
- Mišić, V.; Dinić, A. The vegetation of nature preserves on the Stara Planina Mt. In Proceedings of the 5th Symposium on the Flora of Southeast Serbia, Zaječar, Serbia, 5–9 June 1997; pp. 129–138. [Google Scholar]
- Berisha, N.; Rizani, K.L.; Kadriaj, B.; Millaku, F. Notes on the distribution, ecology, associated vegetation and conservation status of Gymnadenia (Orchidaceae) in Kosovo. Ital. Bot. 2021, 12, 1–27. [Google Scholar] [CrossRef]
- Petković, B.; Tatić, B. Ass. Scirpeto-Phragmitetum Koch. W. 1926, around the Ubavac stream on Fruška Gora. Bull. Nat. Hist. Mus. Belgrade 1978, 33B, 55–68. (In Serbian) [Google Scholar]
- Ranđelović, V. Marsh Vegetation along the Upper Reaches of the South Morava. Diploma Thesis, University of Novi Sad, Novi Sad, Serbia, 1988. (In Serbian). [Google Scholar]
- Ponert, J. Contributions to the orchids of Republic of Macedonia and Serbia. J. Eur. Orch. 2014, 46, 561–577. [Google Scholar]
- Obratov-Petković, D.; Popović, I.; Dajić-Stevanović, Z. Diversity of the vascular flora of Mt. Zlatar (Southwest Serbia). Eurasia J. Biosci. 2007, 5, 35–47. [Google Scholar]
- Perišić, S. Flora and Vegetation of Blačko Lake. Master’s Thesis, University of Belgrade, Belgrade, Serbia, 2002; pp. 1–201. (In Serbian). [Google Scholar]
- Čolić, D. New localities of round-leaved sundew (Drosera rotundifolia L.) on Mt. Stara Planina—Eastern Serbia. Zaštita Prir. 1965, 29–30, 523. (In Serbian) [Google Scholar]
- Petković, B. The new community of matgrass, ass. Carici-Nardetum strictae, from the area of southwestern Serbia. Bull. Nat. Hist. Mus. Belgrade 1985, 40, 89–95. [Google Scholar]
- Stevanović, V.; Niketić, M.; Lakušić, D. Chorological additions to the flora of eastern Yugoslavia. Flora Mediterr. 1991, 1, 121–142. [Google Scholar]
- Lakušić, D. The high mountain flora of Kopaonik—an ecological-phytogeographical study. Master’s Thesis, University of Belgrade, Belgrade, Serbia, 1993; pp. 1–230. (In Serbian). [Google Scholar]
- Zlatković, B.; Ranđelović, V.; Ranđelović, N. Material for the flora of southeastern Serbia. In Flora and Vegetation, Proceedings of the 3rd Symposium on the Flora of Southeast Serbia, Leskovac-Pirot, Serbia, 1–4 June 1993; Ranđelović, N., Ed.; University of Niš: Niš, Serbia; Faculty of Technology in Leskovac: Leskovac, Serbia, 1993; pp. 95–110. [Google Scholar]
- Petković, B.; Krivošej, Z.; Veljić, M. Selaginello-Eriophoretum latifoli—ass. nova from Mount Ošljak (Serbia, Kosovo). Bull. Inst. Jard. Bot. Univ. Belgrade 1996, 30, 89–95. (In Serbian) [Google Scholar]
- Ranđelović, V.; Zlatković, B.; Amidžić, L. Flora and vegetation of the high mountain mires of the Mts. Šar planina. Zaštita Prir. 1998, 50, 377–397. (In Serbian) [Google Scholar]
- Todorović, T. Phytocenological analysis of endangered plant species of the Vlasina plateau. Master’s Thesis, University of Niš, Niš, Serbia, 2013; pp. 1–58. [Google Scholar]
- Rudski, I. Plant communities in the high mountains of southern Serbia. Šumarski List. 1938, 61, 611–623. (In Serbian) [Google Scholar]
- Djordjević, V.; Tsiftsis, S.; Lakušić, D.; Stevanović, V. Niche analysis of orchids of serpentine and non-serpentine areas: Implications for conservation. Plant Biosyst. 2016, 150, 710–719. [Google Scholar] [CrossRef]
- Budzhak, V.V.; Chorney, I.I.; Tokariuk, A.I.; Kuzemko, A.A. Numeric syntaxonomical analysis of the communities with participation of species from Molinia caerulea complex in the southwest of Ukraine. Hacquetia 2016, 15, 63–78. [Google Scholar] [CrossRef]
- Oberdorfer, E. Suddeutsche Pflanzengesellschaften (Plant Communities of Southern Germany). III [Secalietea–Molinio-Arrhenatheretea]; Gustav Fischer: Stuttgart, Germany, 1983. (In German) [Google Scholar]
- Dijk, E.; Grootjans, A.P. Performance of four Dactylorhiza species over a complex trophic gradient. Acta Bot. Neerl. 1998, 47, 351–368. [Google Scholar]
- Schrautzer, J.; Fichtner, A.; Huckauf, A.; Rasran, L.; Jensen, K. Long-term population dynamics of Dactylorhiza incarnata (L.) Soó after abandonment and re-introduction of mowing. Flora 2011, 206, 622–630. [Google Scholar] [CrossRef]
- Šilc, U.; Aćić, S.; Škvorc, Ž.; Krstonošić, D.; Franjić, J.; Dajić Stevanović, Z. Grassland vegetation of the Molinio-Arrhenatheretea class in the NW Balkan Peninsula. Appl. Veg. Sci. 2014, 17, 591–603. [Google Scholar] [CrossRef]
- Škvorc, Ž.; Ćuk, M.; Zelnik, I.; Franjić, J.; Igić, R.; Ilić, M.; Krstonošić, D.; Vukov, D.; Čarni, A. Diversity of wet and mesic grasslands along a climatic gradient on the southern margin of the Pannonian Basin. Appl. Veg. Sci. 2020, 23, 676–697. [Google Scholar] [CrossRef]
- Illyés, Z.; Halász, K.; Rudnóy, S.; Ouanphanivanh, N.; Garay, T.; Bratek, Z. Changes in the diversity of the mycorrhizal fungi of orchids as a function of the water supply of the habitat. J. Appl. Bot. Food. 2009, 83, 28–36. [Google Scholar]
- Hrivnák, R.; Gömöry, D.; Cvachová, A. Inter-annual variabilty of the abundance and morphology of Dactylorhiza majalis (Orchidaceae-Orchideae) in two permanent plots of a mire in Slovakia. Phyton-Ann. Rei Bot. 2006, 46, 27–44. [Google Scholar]
- Molnár, A. (Ed.) Atlas of Hungarian Orchids; Kossuth Kiadó: Budapest, Hungary, 2011. (In Hungarian) [Google Scholar]
- Urban, D. Characteristics of the locality of Hammarbya paludosa (L.) O. Kuntze on the Łęczna-Włodawa Plain (West Polesie). Teka Komis Ochr. Środow. Przyr. 2013, 10, 448–454. [Google Scholar]
- Blinova, I.V. Spatial population structure of rare orchid species in rich fens in the central part of Murmansk oblast. Russ. J. Ecol. 2016, 47, 234–240. [Google Scholar] [CrossRef]
- Jermakowicz, E.; Brzosko, E.; Kotowicz, J.; Wróblewska, A. Genetic diversity of orchid Malaxis monophyllos over European range as an effect of population properties and postglacial colonization. Pol. J. Ecol. 2017, 65, 69–86. [Google Scholar]
- Dijk, E.; Willems, J.H.; van Andel, J. Nutrient responses as a key factor to the ecology of orchid species. Acta Bot. Neerl. 1997, 46, 339–363. [Google Scholar] [CrossRef]
- Bowles, M.; Zettler, L.; Bell, T.; Kelsey, P. Relationship between soil characteristics, distribution and restoration potential of the federal threatened eastern prairie fringed orchid, Platanthera leucophaea (Nutt.) Lindl. Am. Midl. Nat. 2005, 154, 273–285. [Google Scholar] [CrossRef]
- Leuschner, C.; Ellenberg, H. Ecology of Central European Non-Forest Vegetation: Coastal to Alpine, Natural to Man-Made Habitats: Vegetation Ecology of Central Europe; Springer International Publishing: Cham, Switzerland, 2017. [Google Scholar]
- Hrivnák, M.; Slezák, M.; Galvánek, D.; Vlčko, J.; Belanová, E.; Rízová, V.; Senko, D.; Hrivnák, R. Species Richness, Ecology, and Prediction of Orchids in Central Europe: Local-Scale Study. Diversity 2020, 12, 154. [Google Scholar] [CrossRef]
- Tali, K.; Foley, M.J.Z.; Kull, T. Biological flora of the British Isles, 232. Orchis ustulata L. J. Ecol. 2004, 92, 174–184. [Google Scholar]
- Beyrle, H.; Penningsfeld, F.; Hock, B. The role of nitrogen concentration in determining the outcome of the interaction between Dactylorhiza incarnata (L.) Soó and Rhizoctonia sp. New Phytol. 1991, 117, 665–672. [Google Scholar] [CrossRef]
- Dijk, E.; Olff, H. Effects of nitrogen, phosphorus and potassium fertilization on field performance of Dactylorhiza majalis. Acta Bot. Neerl. 1994, 43, 383–392. [Google Scholar] [CrossRef]
- Silvertown, J.; Wells, D.A.; Gillman, M.; Dodd, M.E.; Robertson, H.; Lakhani, K.H. Short-term effects and long-term aftereffects of fertilizer application on the flowering population of green-winged orchid Orchis morio. Biol. Conserv. 1994, 69, 191–197. [Google Scholar] [CrossRef]
- Hornemann, G.; Michalski, S.G.; Durka, W. Short-term fitness and long-term population trends in the orchid Anacamptis morio. Plant Ecol. 2012, 213, 1583–1595. [Google Scholar] [CrossRef]
- Stevanović, V.; Tan, K.; Iatrou, G. Distribution of the endemic Balkan flora on serpentine I. Obligate serpentine endemics. Plant Syst. Evol. 2003, 242, 149–170. [Google Scholar] [CrossRef]
- Chiari, M.; Djerić, N.; Garfagnoli, F.; Hrvatović, H.; Krstić, M.; Levi, N.; Malasoma, A.; Marroni, M.; Menna, F.; Nirta, G.; et al. The geology of the Zlatibor-Maljen area (western Serbia): A geotraverse accross theophiolites of the Dinaric-Hellenic collisional belt. Ofioliti 2011, 36, 139–166. [Google Scholar]
- Gawlick, H.-J.; Sudar, M.; Missoni, S.; Suzuki, H.; Lein, R.; Jovanović, D. Triassic-Jurassic geodynamic evolution of the Dinaridic Ophiolite Belt (Inner Dinarides, SW Serbia). J. Alpine Geol. 2017, 55, 1–167. [Google Scholar]
- Perry, E.C.; Lefticariu, L. Formation and geochemistry of Precambrian cherts. Treatise Geochem. 2003, 7, 99–113. [Google Scholar]
- Djordjević, V.; University of Belgrade, Faculty of Biology, Institute of Botany and Botanical Garden, Belgrade, Serbia. Unpublished work. 2022.
- Duraki, Š. Vascular flora of the high mountain ridge of Kobilica on Šar mountain. Master’s Thesis, University of Belgrade, Belgrade, Serbia, 2008. (In Serbian). [Google Scholar]
- Dzulynski, S.; Walton, E.K. Sedimentary Features in Flysch and Greywackes. Developments in Sedimentology; Elsevier: Amsterdam, The Netherlands, 1965; Volume 7. [Google Scholar]
- Duncan, R.P.; Clemants, S.E.; Corlett, R.T.; Hahs, A.K.; McCarthy, M.A.; McDonnell, M.J.; Schwartz, M.W.; Thompson, K.; Vesk, P.A.; Williams, N.S.G. Plant traits and extinction in urban areas: A meta-analysis of 11 cities. Glob. Ecol. Biogeogr. 2011, 20, 509–519. [Google Scholar] [CrossRef]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publications Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Ballantyne, M.; Pickering, C. Ecotourism as a threatening process for wild orchids. J. Ecotourism. 2012, 11, 34–47. [Google Scholar] [CrossRef]
- Light, M.; MacConaill, M. Effects of trampling on a terrestrial orchid environment. Lankesteriana 2007, 7, 294–298. [Google Scholar] [CrossRef][Green Version]
- Pickering, C.M.; Hill, W.; Newsome, D.; Leung, Y.-F. Comparing hiking, mountain biking and horse riding impacts on vegetation and soils in Australia and the United States of America. J. Environ. Manag. 2010, 91, 551–562. [Google Scholar] [CrossRef]
- Wraith, J.; Pickering, C. Quantifying anthropogenic threats to orchids using the IUCN Red List. Ambio 2018, 47, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Grlić, L. Encyclopedia of Wild Edible Plants; August Cesarec: Zagreb, Croatia, 1986. (In Serbian) [Google Scholar]
- Arditti, J. Fundamentals of Orchid Biology; Wiley Interscience: New York, NY, USA, 1992. [Google Scholar]
- IPCC. The Physical Science Basis; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, MA, USA, 2021. [Google Scholar]
- Kolanowska, M. The future of a montane orchid species and the impact of climate change on the distribution of its pollinators and magnet species. Glob. Ecol. Conserv. 2021, 32, e01939. [Google Scholar] [CrossRef]
- Hájek, M.; Těšitel, J.; Tahvanainen, T.; Peterka, T.; Jiménez-Alfaro, B.; Jansen, F.; Pérez-Haase, A.; Garbolino, E.; Carbognani, M.; Kolari, T.H.M.; et al. Rising temperature modulates pH niches of fen species. Glob. Change Biol. 2022, 28, 1023–1037. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Degree of Representation | Chorological Group | Life Form |
|---|---|---|---|
| Anacamptis coriophora (L.) R.M.Bateman, Pridgeon & M.W.Chase subsp. coriophora | 3 | CE | T |
| Anacamptis laxiflora (Lam.) R.M.Bateman, Pridgeon & M.W.Chase subsp. laxiflora | 1 | MED-SUBMED | T |
| Anacamptis morio (L.) R.M.Bateman, Pridgeon & M.W.Chase subsp. morio | 4 | CE | T |
| Anacamptis palustris (Jacq.) R.M.Bateman, Pridgeon & M.W.Chase subsp. palustris | 1 | EAS | T |
| Anacamptis palustris subsp. elegans (Heuff.) R.M.Bateman, Pridgeon & M.W.Chase | 1 | EAS | T |
| Anacamptis pyramidalis (L.) Rich. | 4 | MED-SUBMED | T |
| Coeloglossum viride (L.) Hartm. | 3 | BOR | I |
| Dactylorhiza cordigera (Fr.) Soó subsp. cordigera | 2 | CEM | I |
| Dactylorhiza cordigera subsp. bosniaca (Beck) Soó | 2 | CEM | I |
| Dactylorhiza fuchsii (Druce) Soó subsp. fuchsii | 3 | BOR | I |
| Dactylorhiza incarnata (L.) Soó subsp. incarnata | 1 | BOR | I |
| Dactylorhiza kalopissii subsp. macedonica (J.Hölzinger & Künkele) Kreutz | 1 | MED-SUBMED | I |
| Dactylorhiza maculata (L.) Soó subsp. maculata | 2 | BOR | I |
| Dactylorhiza maculata subsp. transsilvanica (Schur) Soó | 2 | CE | I |
| Dactylorhiza majalis (Rchb.) P.F.Hunt & Summerh. subsp. majalis | 1 | CE | I |
| Dactylorhiza saccifera (Brongn.) Soó subsp. saccifera | 3 | MED-SUBMED | I |
| Dactylorhiza sambucina (L.) Soó | 4 | CE | I |
| Epipactis palustris (L.) Crantz | 1 | EAS | R |
| Gymnadenia conopsea (L.) R.Br. | 3 | EAS | I |
| Gymnadenia densiflora (Wahlenb.) A.Dietr. | 2 | EAS | I |
| Gymnadenia frivaldii Hampe ex Griseb. | 1 | CEM | I |
| Gymnadenia odoratissima (L.) Rich. | 4 | CE | I |
| Herminium monorchis (L.) R.Br. | 2 | EAS | T |
| Neotinea ustulata (L.) R.M.Bateman, Pridgeon & M.W.Chase | 3 | CE | T |
| Neottia ovata (L.) Bluff & Fingerh. | 3 | EAS | R |
| Nigritella rhellicani Teppner & E.Klein | 4 | CEM | I |
| Orchis mascula subsp. speciosa (Mutel) Hegi | 4 | CE-EUX-CAUC | T |
| Orchis militaris L. subsp. militaris | 4 | EAS | T |
| Platanthera bifolia (L.) Rich. | 3 | EAS | I |
| Platanthera chlorantha (Custer) Rchb. | 3 | CE | I |
| Pseudorchis albida (L.) Á.Löve & D.Löve subsp. albida | 3 | BOR | I |
| Spiranthes spiralis (L.) Chevall. | 4 | CE | T |
| Traunsteinera globosa (L.) Rchb. | 3 | CEM | T |
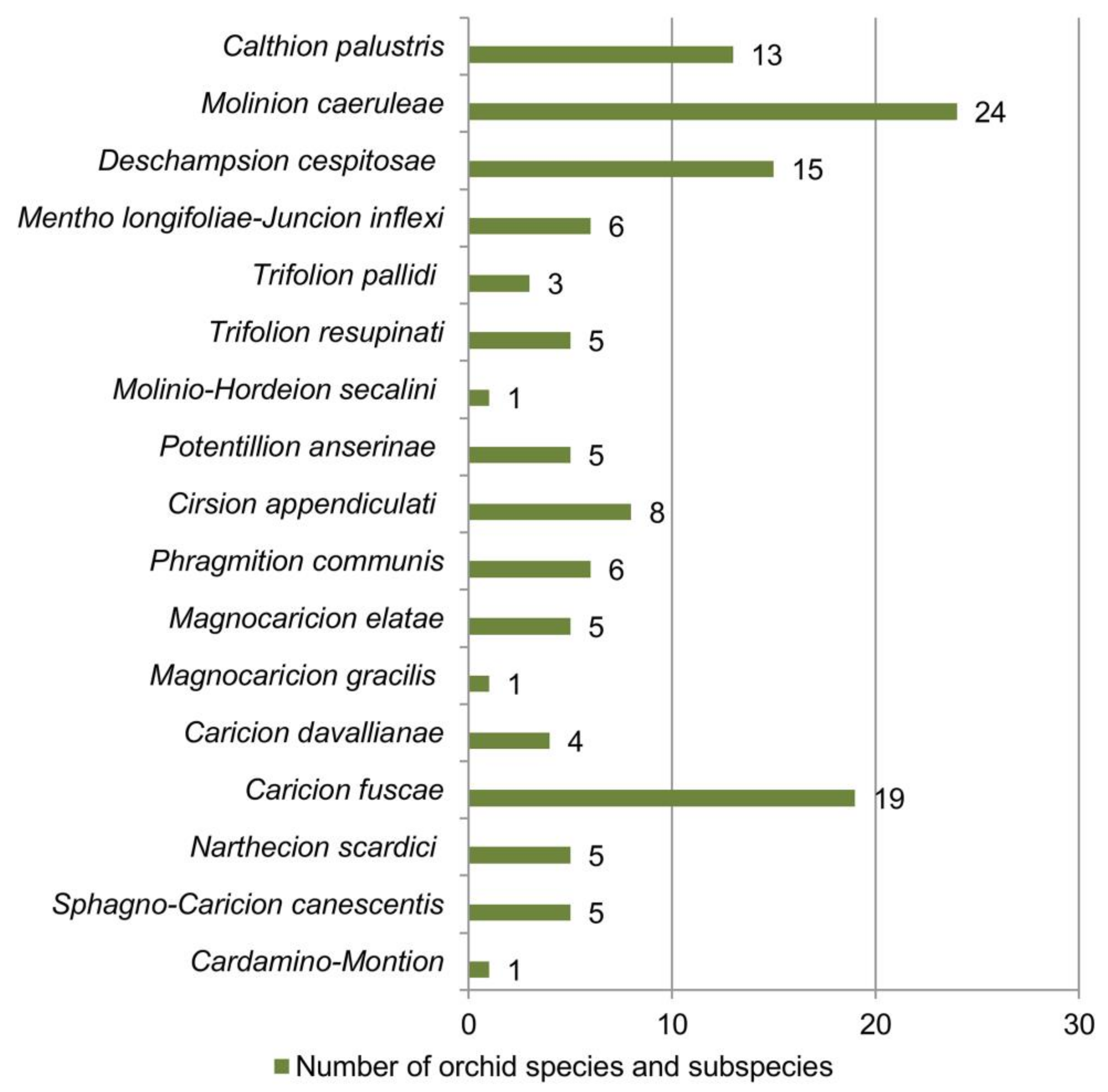
| Vegetation Class | Vegetation Order | Vegetation Alliance | Orchid Species and Subspecies | Literature Sources |
|---|---|---|---|---|
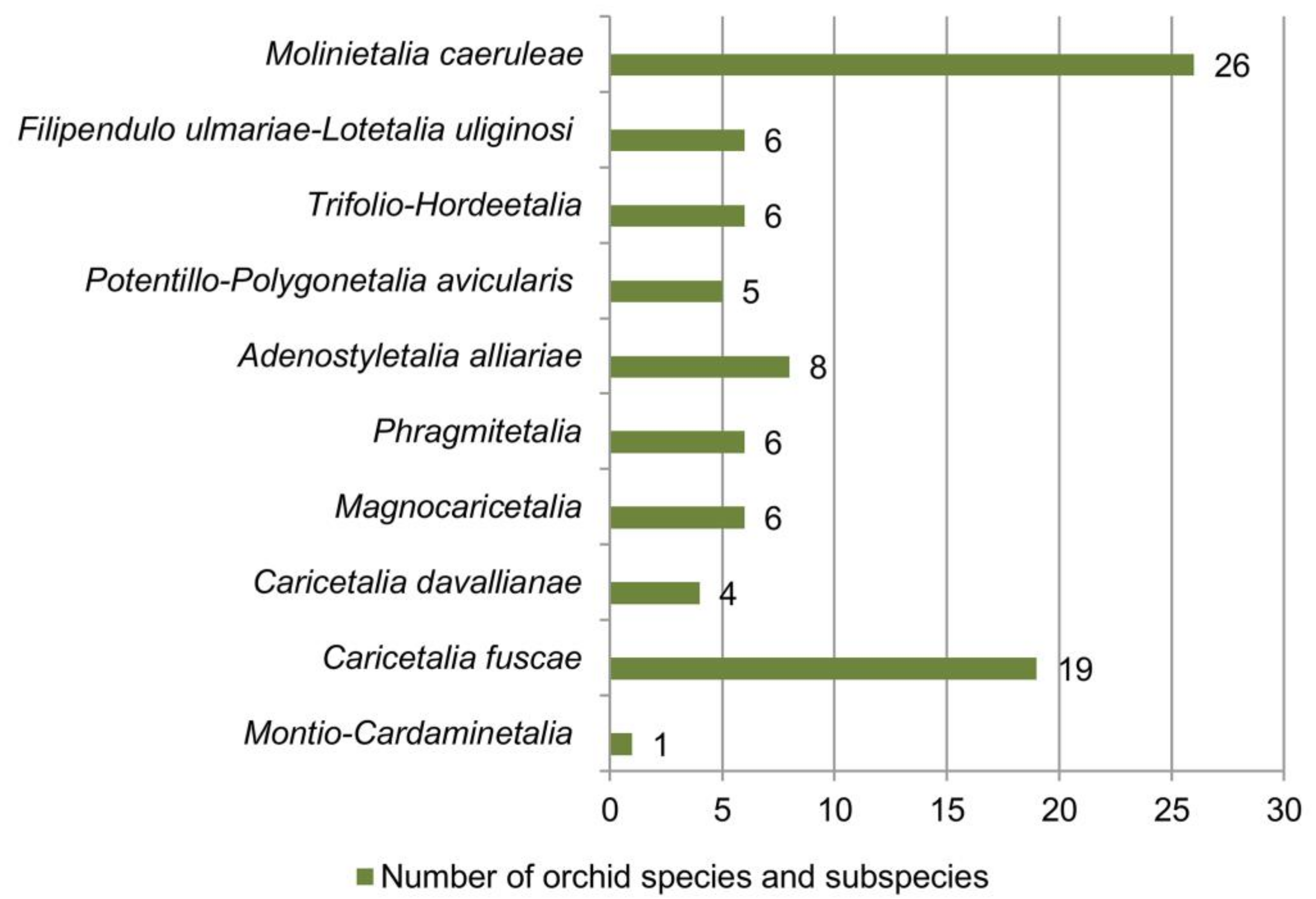
| Molinio- Arrhenatheretea Tx. 1937 | Molinietalia caeruleae Koch 1926 | Calthion palustris Tx. 1937 | Anacamptis palustris subsp. elegans, Anacamptis morio subsp. morio, Dactylorhiza incarnata subsp. incarnata, Dactylorhiza maculata subsp. maculata, Dactylorhiza maculata subsp. transsilvanica, Dactylorhiza cordigera subsp. cordigera, Dactylorhiza cordigera subsp. bosniaca, Dactylorhiza saccifera, Epipactis palustris, Gymnadenia conopsea, Neottia ovata, Platanthera bifolia, Traunsteinera globosa | [15,18,41,59,60,61,62] |
| Molinio- Arrhenatheretea Tx. 1937 | Molinietalia caeruleae Koch 1926 | Molinion caeruleae Koch 1926 | Anacamptis coriophora subsp. coriophora, Anacamptis morio subsp. morio, Anacamptis palustris subsp. palustris, Anacamptis palustris subsp. elegans, Anacamptis pyramidalis, Dactylorhiza fuchsii, Dactylorhiza incarnata subsp. incarnata, Dactylorhiza maculata subsp. maculata, Dactylorhiza maculata subsp. transsilvanica, Dactylorhiza majalis subsp. majalis, Dactylorhiza saccifera, Dactylorhiza sambucina, Epipactis palustris, Gymnadenia conopsea, Herminium monorchis, Neotinea ustulata, Neottia ovata, Orchis mascula subsp. speciosa, Orchis militaris, Platanthera bifolia, Platanthera chlorantha, Pseudorchis albida, Spiranthes spiralis, Traunsteinera globosa | [15,18,41,60,63,64,65,66,67,68,69,70] |
| Molinio- Arrhenatheretea Tx. 1937 | Molinietalia caeruleae Koch 1926 | Deschampsion cespitosae Horvatić 1930 | Anacamptis coriophora subsp. coriophora, Anacamptis morio subsp. morio, Anacamptis palustris subsp. palustris, Anacamptis palustris subsp. elegans, Dactylorhiza cordigera subsp. cordigera, Dactylorhiza incarnata subsp. incarnata, Dactylorhiza maculata subsp. maculata, Dactylorhiza maculata subsp. transsilvanica, Dactylorhiza saccifera, Epipactis palustris, Gymnadenia conopsea, Neotinea ustulata, Neottia ovata, Platanthera bifolia, Traunsteinera globosa | [15,18,40,41,60,62,66,71,72,73,74] |
| Molinio- Arrhenatheretea Tx. 1937 | Filipendulo ulmariae-Lotetalia uliginosi Passarge 1975 | Mentho longifoliae-Juncion inflexi T. Müller et Görs ex de Foucault 2009 | Dactylorhiza incarnata subsp. incarnata, Dactylorhiza saccifera, Epipactis palustris, Gymnadenia conopsea, Gymnadenia densiflora, Platanthera bifolia | [15,18,41,75] |
| Molinio- Arrhenatheretea Tx. 1937 | Trifolio-Hordeetalia Horvatić 1963 | Trifolion pallidi Ilijanić 1969 | Anacamptis morio subsp. morio, Anacamptis palustris subsp. palustris, Anacamptis palustris subsp. elegans | [15,18,41,76,77,78] |
| Molinio- Arrhenatheretea Tx. 1937 | Trifolio-Hordeetalia Horvatić 1963 | Trifolion resupinati Micevski 1957 | Anacamptis coriophora subsp. coriophora, Anacamptis laxiflora, Anacamptis morio subsp. morio, Anacamptis palustris subsp. elegans, Gymnadenia conopsea | [74,79,80,81,82,83,84,85,86] |
| Molinio- Arrhenatheretea Tx. 1937 | Trifolio-Hordeetalia Horvatić 1963 | Molinio-Hordeion secalini Horvatić 1934 | Anacamptis laxiflora | [87] |
| Molinio- Arrhenatheretea Tx. 1937 | Potentillo- Polygonetalia avicularis Tx. 1947 | Potentillion anserinae Tx. 1947 | Anacamptis coriophora subsp. coriophora, Anacamptis palustris subsp. palustris, Anacamptis palustris subsp. elegans, Dactylorhiza incarnata subsp. incarnata, Orchis militaris | [68,71,88] |
| Mulgedio-Aconitetea Hadač et Klika in Klika et Hadač 1944c 1944 | Adenostyletalia alliariae Br.-Bl. 1930 | Cirsion appendiculati Horvat et al. 1937 | Dactylorhiza maculata subsp. maculata, Dactylorhiza sambucina, Dactylorhiza cordigera subsp. cordigera, Dactylorhiza saccifera, Gymnadenia frivaldii, Gymnadenia odoratissima, Gymnadenia conopsea, Nigritella rhellicani | [74,89,90] |
| Phragmito- Magnocaricetea Klika in Klika et Novák 1941 | Phragmitetalia Koch 1926 | Phragmition communis Koch 1926 | Anacamptis palustris subsp. palustris, Anacamptis palustris subsp. elegans, Dactylorhiza incarnata subsp. incarnata, Dactylorhiza kalopissii subsp. macedonica, Epipactis palustris, Gymnadenia conopsea | [15,18,41,60,91,92,93] |
| Phragmito- Magnocaricetea Klika in Klika et Novák 1941 | Magnocaricetalia Pignatti 1953 | Magnocaricion elatae Koch 1926 | Anacamptis coriophora subsp. coriophora, Anacamptis morio subsp. morio, Dactylorhiza incarnata subsp. incarnata, Gymnadenia conopsea, Traunsteinera globosa | [15,41,60,94] |
| Phragmito- Magnocaricetea Klika in Klika et Novák 1941 | Magnocaricetalia Pignatti 1953 | Magnocaricion gracilis Géhu 1961 | Anacamptis palustris subsp. elegans | [86,95] |
| Scheuchzerio palustris-Caricetea fuscae Tx. 1937 | Caricetalia davallianae Br.-Bl. 1950 | Caricion davallianae Klika 1934 | Dactylorhiza cordigera subsp cordigera, Dactylorhiza cordigera subsp. bosniaca, Epipactis palustris, Gymnadenia frivaldii | [15,27,41,69] |
| Scheuchzerio palustris-Caricetea fuscae Tx. 1937 | Caricetalia fuscae Koch 1926 | Caricion fuscae Koch 1926 | Anacamptis coriophora subsp. coriophora, Anacamptis morio subsp. morio, Dactylorhiza cordigera subsp. bosniaca, Dactylorhiza cordigera subsp. cordigera, Dactylorhiza incarnata subsp. incarnata, Dactylorhiza kalopissii subsp. macedonica, Dactylorhiza maculata subsp. maculata, Dactylorhiza maculata subsp. transsilvanica, Dactylorhiza majalis subsp. majalis, Dactylorhiza saccifera, Epipactis palustris, Gymnadenia conopsea, Gymnadenia frivaldii, Herminium monorchis, Neottia ovata, Nigritella rhellicani, Platanthera bifolia, Pseudorchis albida, Traunsteinera globosa | [15,18,27,41,59,62,74,89,93,96,97,98,99,100,101,102,103] |
| Scheuchzerio palustris-Caricetea fuscae Tx. 1937 | Caricetalia fuscae Koch 1926 | Narthecion scardici Horvat ex Lakušić 1968 | Gymnadenia frivaldii, Gymnadenia conopsea, Dactylorhiza cordigera subsp. cordigera, Dactylorhiza cordigera subsp. bosniaca, Pseudorchis albida | [27,90,102,104] |
| Scheuchzerio palustris-Caricetea fuscae Tx. 1937 | Caricetalia fuscae Koch 1926 | Sphagno-Caricion canescentis Passarge (1964) 1978 | Dactylorhiza cordigera subsp. cordigera, Dactylorhiza incarnata subsp. incarnata, Dactylorhiza maculata subsp. maculata, Dactylorhiza maculata subsp. transsilvanica, Epipactis palustris | [15,27,41,103] |
| Montio-Cardaminetea Br.-Bl. et Tx. ex Klika et Hadač 1944 | Montio- Cardaminetalia Pawłowski et al. 1928 | Cardamino- Montion Br.-Bl. 1926 | Dactylorhiza cordigera subsp. cordigera | [89] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djordjević, V.; Aćić, S.; Kabaš, E.; Lazarević, P.; Tsiftsis, S.; Lakušić, D. The Orchids of Wetland Vegetation in the Central Balkans. Diversity 2023, 15, 26. https://doi.org/10.3390/d15010026
Djordjević V, Aćić S, Kabaš E, Lazarević P, Tsiftsis S, Lakušić D. The Orchids of Wetland Vegetation in the Central Balkans. Diversity. 2023; 15(1):26. https://doi.org/10.3390/d15010026
Chicago/Turabian StyleDjordjević, Vladan, Svetlana Aćić, Eva Kabaš, Predrag Lazarević, Spyros Tsiftsis, and Dmitar Lakušić. 2023. "The Orchids of Wetland Vegetation in the Central Balkans" Diversity 15, no. 1: 26. https://doi.org/10.3390/d15010026
APA StyleDjordjević, V., Aćić, S., Kabaš, E., Lazarević, P., Tsiftsis, S., & Lakušić, D. (2023). The Orchids of Wetland Vegetation in the Central Balkans. Diversity, 15(1), 26. https://doi.org/10.3390/d15010026