Abstract
We studied how range sizes and shifts in species ranges depend on niche breadth in European dragonflies. We measured range sizes and shifts over a 22-year period (1988–2010) and grouped species into those reproducing in permanent running (perennial lotic) water, permanent standing (perennial lentic) water, and temporary (running or standing) water. Running water species are more specialized and have narrower niches with a more fixed niche position than standing water species. Temporary water species are more generalist and have broader niches without a fixed niche position as clear as permanent water species because they may utilize both temporary and permanent habitats. Running water species have smaller ranges, and some of them have contracted their ranges more than species reproducing in standing or temporary waters; that is, they are especially at risk of habitat loss and climate change because of the joint effects of their narrow niches and small range sizes. Temporary water species track climate changes better than permanent water species. This suggests that ecological specialization may cause contemporary range shifts to lag behind changes in climate and resources. Furthermore, it indicates that recent changes in climate and human land use cause biotic homogenization, where specialists are outperformed and replaced by generalists.
1. Introduction
Climate change strongly impacts the distributions of dragonflies, and it seems that, at least at temperate latitudes, it has become one of the most important driving forces behind distributional changes in this insect order [1,2,3,4,5,6,7,8,9,10,11,12,13,14,15,16]. Climate change causes dramatic shifts in geographical distributions of species. Species that are limited by their lower thermal tolerance threshold and are capable of at least partially tracking changes in climate experience significant range expansions [17]. Species from a variety of ecological systems [17], across [18] and within broad taxonomic groups [10], exhibit northward range expansions, consistent with a warming climate. Apart from their climatic envelopes, the occurrence of species is also determined by the availability of suitable habitats [19,20]. Species that cannot keep pace with climate change [17] or adapt to new environmental conditions [21,22] by utilizing formerly unavailable habitats or new habitats will lag behind changes in climate and resources and eventually experience range contraction or local to regional extinction [23,24].
The ongoing climate changes cause habitat alterations and threaten certain aquatic habitats [25]. A 20% decrease in summer precipitation has been projected for all areas around the Mediterranean Basin, and increased evapotranspiration due to rising air and water temperatures will also reduce water availability [25]. Higher proportions of winter precipitation will be received as rain, and snow will accumulate in smaller quantities and melt earlier in the season, leading to reduced summer flows [25]. Such shifts in the form and timing of precipitation and runoff may disrupt the life cycle of species adapted to permanent running water by causing unstable flow regimes and summertime desiccation of streams and rivers [12]. Drying has already been observed in southern Europe, where increasing numbers of standing water bodies dry out or become temporarily dry [2]. Human population growth and industrial and agricultural development further intensify the pressures on aquatic habitats and increase the levels of water scarcity [12]. In this context, change in climate and human land use are expected to have a negative impact on the distribution of species dependent on aquatic habitats, especially species adapted to permanent running water habitats.
Dragonflies reproduce in most running and standing water habitats, and their larvae cannot freely migrate in search of suitable conditions in the same way as adults can [26]. Many dragonfly species inhabit water bodies that dry out periodically, and dragonflies’ adaptations to that condition include prolonged egg stage [27], drought-resistant eggs [28,29], or larvae that can survive drought periods for as long as 8 months in moist cracks in the sediment [30]. Some species have a rapid larval development and a shortened larval stage [27], and some are able to accelerate larval development in response to rapid water loss [31]. However, not all species have these adaptations, and the development of most dragonfly larvae lasts from 1 to 3 years, but depending on the latitude/altitude and habitat suitability, it may last up to 6 years [15,26]. One of the effects of climate change is therefore caused by the selective pressure that shift in water permanence constitutes on species assemblages.
A major challenge in conservation ecology is to understand how different species respond to climate and environmental changes; that is, what enables some species to persist while others decline? Several studies have linked responses to climate change to species traits, such as niche breadth [8,11,32,33,34,35,36,37,38]. Local species diversity often increases when the temperature rises mainly due to the arrival of ecological generalists that can respond quickly because they can utilize a large variety of habitats [34,36,39]. This is, however, accompanied by loss of habitat specialists that only have the ability to utilize a narrow range of habitats [19,36]. Consequently, ecological specialists with narrow niches are replaced by broad niche generalists, and this in turn causes biotic homogenization, where species from a variety of taxonomic groups and geographic regions are lost due to recent climate change [19,40,41,42]. Hence, the dynamics of range shifts are constrained not only by climatic boundaries but also by the niche breadth of species. Functional species traits are therefore important when determining and understanding how species respond to climate change [8,11,33,36,37,38]. This has led to the hypothesis that key traits, such as niche breadth, can be used to predict temporal changes in species range.
We study how niche breadth in a larval habitat influences the response of European dragonfly species to climate change. The larvae of species adapted to permanent running (perennial lotic) waters have a narrower niche breadth with a more fixed niche position than permanent standing (perennial lentic) water and temporary (running or standing) water species. Species adapted to life in temporary waters have the broadest niche breadth without a fixed niche position as they may be found not only in temporary water but also in various permanent habitats, sometimes including both running and standing waters (Figure 1). According to the niche breadth of a larvae habitat, we divided species into three groups: species reproducing in (1) permanent running water, (2) permanent standing water, and (3) temporary water. Instead of a single focus on the running- to standing-water gradient [8,11,33], we chose to include the complete range of habitat resources by also implementing the temporary water species as an independent functional group. First, we chose this because the larvae of many dragonfly species may utilize different aquatic habitats along both a running- to standing-water gradient and a permanent- to temporary-water gradient [26]. As the transitions from permanent to temporary water bodies affects different dragonfly species in different ways [9], it specifically allows us to test how species with different niche breadths within the complete range of habitat resources react to climatic change. Second, climate change is known to cause biotic homogenization in terrestrial organisms [19,40,41,42], and as the three ecological habitat categories also represent a gradient from habitat specialist to habitat generalist, it allows us to evaluate whether this also applies to organisms whose larvae are strictly aquatic. The larvae of species adapted to permanent running waters are more specialized and habitat constrained than species that can also live in standing waters. Species adapted to life in temporary waters can be considered as more generalist than permanent water species as they may reproduce not only in temporary water but also in various permanent habitats, sometimes including both running and standing waters (Figure 1). Third, by including temporary habitats, it allows us to evaluate the relative importance of temporary ponds that are expected to have an increasing functional significance as an aquatic resource. This is important because among the gradients of water bodies, temporary habitats constitute an essential freshwater ecosystem as underlined by the inclusion of the Mediterranean temporary ponds as a priority habitat for conservation (code 3170) in the European Union (EU) Habitats Directive (92/43/EEC) [43]. Due to their heterogeneity, temporary waters support high diversity and act as stepping stones for the dispersal of species [44].
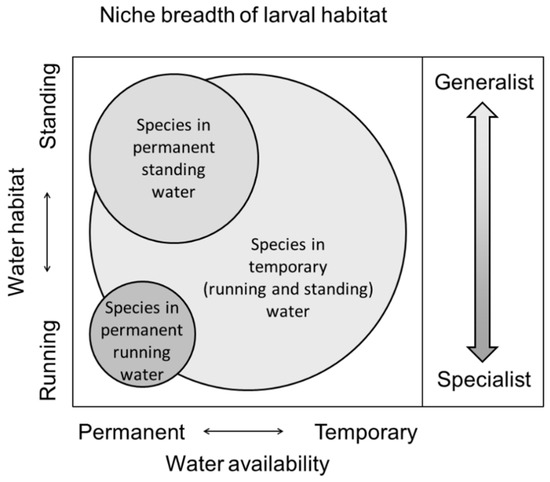
Figure 1.
Size of the ecological niche space of the aquatic larvae of the European dragonfly (Odonata) species separated in three habitat categories: permanent running (perennial lotic) water, permanent standing (perennial lentic) water, and temporary (running or standing) water. Color scale to the right represents a gradient going from habitat specialist with a narrow niche breadth to habitat generalist with a broad niche breadth as also illustrated with the size of the ecological niche space of three habitat categories.
We use distribution maps for European dragonflies from 1988 [45] and 2010 [12] to determine differences in range sizes and follow how both latitudinal and northern range borders of the species have changed over a 22-year period. As documented above, there is a gap in our knowledge of how a key species trait, such as niche breadth, of the larval habitat of European dragonfly species influences their response to climate change. Especially, studies that evaluate the specialist–generalist gradient and test for climate-driven homogenization in aquatic communities and organisms that represent both aquatic and terrestrial ecosystems are important. In this context, we specifically ask: (1) Do species reproducing in permanent standing water and temporary water have larger ranges than species reproducing in permanent running water? (2) Do species reproducing in temporary habitats track changes better than species in permanent habitats? (3) Do species reproducing in permanent running water contract more or expand less than species in permanent standing water and temporary water? (4) Do generalist species outperform specialists with a narrow niche breadth? Answering these questions will help explain how environmental changes affect species differently according to their ability to utilize various types of aquatic habitats.
2. Materials and Methods
2.1. Study Area
Our study area included Europe, which, as the westernmost peninsula of Eurasia, is limited to the north by the Arctic Ocean, to the west by the Atlantic Ocean, and to the south by the Mediterranean Sea. The eastern border of the study area follows a combination of the 35° E longitude and the eastern margin used in outline range maps in Askew [45]. All larger European islands in the Mediterranean were included (Figure 2).
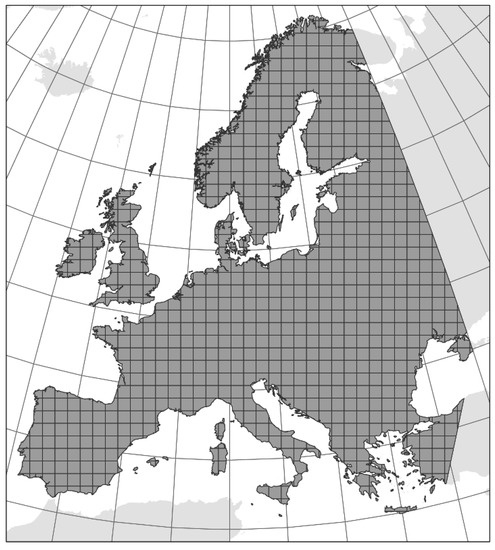
Figure 2.
Study area covers the westernmost peninsula of Eurasia. The gridded map with 880 cells of 100 × 100 km highlighted in grey.
2.2. Data
Distribution ranges of dragonflies in Europe were taken from outline maps in Askew [45] and in the online species summary in Kalkman et al. [12]. Maps from Askew [45] were georeferenced in ArcGIS 10.2 [46] based on scanned TIFF images, whereas maps from Kalkman et al. [12] were provided as shape files from the Freshwater Biodiversity Unit under the IUCN Global Species Program. The maps do not always represent the full distribution of a species, but only the part that falls within the Eurasian peninsula, including western Russia and Africa north of the Sahara. We excluded data from east of 35° E and south of the Mediterranean Sea because dragonfly occurrences in these regions are not well documented (e.g., Dijkstra and Lewington) [47]. Each distribution map from the 1988 and 2010 data was cut with the same European coastline layer in ArcGIS 10.2 [46] to ensure that species ranges followed the same extent of land cover to make them directly comparable.
Of the 130 species of dragonflies known to occur within the study area, we constructed outline range maps for 123 species after excluding vagrant species and species without range maps in one or both data sets (see Table S1 for a list of excluded species, taxonomic and nomenclatural notes, and modifications to species ranges). Of the 123 species included, 4 colonized Europe between 1988 and 2010, whereas 119 species occurred in both data sets (see Table S1 for a list of the 4 and 119 species, respectively). All parts of species ranges categorized as extant were included, whereas range parts where the species was scored as extinct were omitted.
Based on a combination of scientific and popular species names and the wordings—ephemeral water, ephemeral pond, temporary water, temporary pond, seasonal water, and seasonal pond—we searched in the key book references on Eurasian dragonfly species [45,47,48,49,50,51] and peer-reviewed literature found in, for example, Web of Science [27,52,53,54,55] (per March 2016), for which species can occur in temporary water bodies in Europe, regions of origin for species colonizing Europe, or similar latitudes east of Europe and adjacent southern regions. The larval habitat of species not reported to sometimes occur in temporary water bodies was subsequently reviewed to determine whether they had a preference for standing over running water. By this, we assigned a niche breadth of the larval habitat for each species with a primary emphasis on whether they are able to reproduce successfully in temporary (running and standing) water bodies and a secondary emphasis on the remaining species’ ability to utilize permanent standing waters over permanent running ones. The 123 species were divided into three ecological habitat categories—permanent running waters (n = 37), permanent standing waters (n = 49), and temporary (running or standing) waters (n = 37)—based on references in Table S2.
2.3. Data Analysis
We transformed the outline distributions into gridded maps with 880 cells of 100 × 100 km in ArcGIS 10.2 [46] to estimate distributional range as an occupancy of grid cells. The large grid resolution allowed us to minimize artefacts from outline range maps, including false absences or more commonly false presences, thus overestimating the extent of occurrence of species. For each species, we calculated both overall range size within the study area expressed as the number of occupied grid cells and latitudinal range expressed as the latitudinal extent of distributional range between northern and southern range borders in 1988 and 2010, respectively.
Change in distributional range was measured for each species as shifts in both latitudinal range and northern range border. The former was calculated by subtracting the latitudinal extent of distributional range in 1988 from the range in 2010 and the latter by subtracting the maximum latitude in 1988 from the values in 2010. Range expansions and contractions were expressed with positive and negative values, respectively. Species experiencing contraction in latitudinal range or northern range border were divided into two subsets to test for correlations between the extent of their range contraction and their ability to utilize habitats.
2.4. Statistical Analysis
Differences within a year in overall range size and latitudinal range were analyzed in two-way combinations of the three unmatched test groups—permanent running water, permanent standing water, and temporary (running or standing) water—with Mann–Whitney–Wilcoxon tests.
Shift in latitudinal range and northern range border was analyzed in two-way combinations of the three unmatched test groups—permanent running water, permanent standing water, and temporary (running or standing) water—with Mann–Whitney–Wilcoxon tests.
All statistical tests were performed using R [56].
3. Results
3.1. Range Size
Species reproducing in permanent standing water and temporary water had, on average, larger ranges in 1988 and 2010, than permanent running water species (Figure 3, Table S3). Differences between species in permanent running water and permanent standing water were highly significant for both overall range and latitudinal range in 1988 and 2010, respectively (Figure 3, Table S3). In contrast, differences between species in permanent running water and temporary water were only significant for overall range in 2010 and not latitudinal range (Figure 3, Table S3).
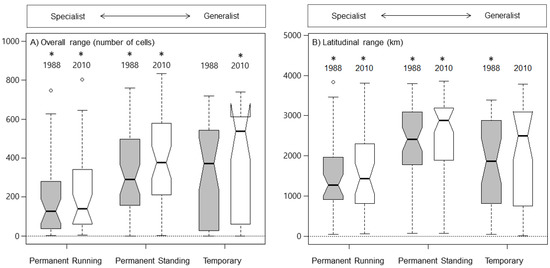
Figure 3.
Range sizes in European dragonfly (Odonata) species adapted to permanent running (perennial lotic) water, permanent standing (perennial lentic) water, and temporary (running and standing) water habitats in 1988 (grey) and 2010 (white), respectively, with overall range size expressed as number of occupied 100 × 100 km grid cells, and latitudinal range expressed as the latitudinal extent of distributional range between northern and southern range borders separated by habitat utilization. The box-and-whisker plots illustrate the spread and skewness of the data through their quartiles. The whiskers extending from the box show data variability outside the upper and lower quartiles. Outlier points that differ significantly from the rest of the dataset are plotted as individual points (empty circles) beyond the whiskers. The box notches narrows around the median (thick black middle line) and offers a visual guide on the significance of the difference of medians; if the notches of two boxes do not overlap, this will provide evidence of a statistically significant difference between the medians. Significant variables at α = 0.05 in the within-year analysis of differences in overall range size and latitudinal range of the three unmatched test groups—permanent running water, permanent standing water, and temporary (running or standing) water—are indicated with *. Top scale represents a gradient going from habitat specialist with a narrow niche breadth to habitat generalist with broad niche breadth.
Differences between species reproducing in permanent standing water and temporary water were only significant for latitudinal range in 1988, despite the latter having a larger median overall range and a smaller median latitudinal range in 1988 and 2010 compared with permanent standing water species (Figure 3, Table S3).
3.2. Range Shifts
On average, the overall range and latitudinal range expanded, and the northern range border shifted farther north from 1988 to 2010 in all the three ecological habitat categories (Table S4). The median change in the overall range in the number of 100 × 100 km grid cells was 35, the median change in the latitudinal range was 130 km, and the median shift in the northern range margin was 61 km for all species.
Species reproducing in permanent standing water and temporary water had, on average, expanded their latitudinal range and northern range border between 1988 and 2010 to a larger extent than species in permanent running water (Figure 4, Table S4). Differences between species in permanent running water and permanent standing water were, however, not significant for a shift in either latitudinal range or northern range border (Figure 4, Table S4). In contrast, differences between species in permanent running water and temporary water and between permanent standing water and temporary water were highly significant for both range shift measures; that is, temporary water species had expanded both their latitudinal range and northern range border to a much larger extent than species in permanent (both running and standing) waters (Figure 4, Table S4).
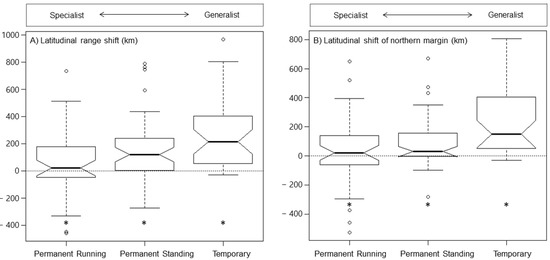
Figure 4.
Range shift in European dragonfly (Odonata) species adapted to permanent running (perennial lotic) water, permanent standing (perennial lentic) water, or temporary (running and standing) water habitats between 1988 and 2010 with a shift in latitudinal range expressed as the change in latitudinal extent of distributional range and a shift in northern range border expressed as the change in the maximum of distributional range latitude separated by habitat utilization. Range expansion and contraction is expressed with positive and negative values, respectively. The box-and-whisker plots illustrate the spread and skewness of the data through their quartiles. The whiskers extending from the box show data variability outside the upper and lower quartiles. Outlier points that differ significantly from the rest of the dataset are plotted as individual points (empty circles) beyond the whiskers. The box notches narrow around the median (thick black middle line) and offer a visual guide of the significance of the difference of medians; if the notches of two boxes do not overlap, this will provide evidence of a statistically significant difference between the medians. Significant variables at α = 0.05 in the analysis of a shift in latitudinal range and northern range border of the three unmatched test groups —permanent running water, permanent standing water, and temporary (running or standing) water—are indicated with *. Top scale represents a gradient going from habitat specialist with a narrow niche breadth to habitat generalist with a broad niche breadth.
Of the species experiencing contraction in either latitudinal range or northern range border between 1988 and 2010, those in permanent running water had, on average, contracted latitudinal range and northern range border to a much larger extent than species in permanent standing water and temporary water (Table S4). Differences in range contraction between the three habitat categories were, however, not significant for either of the range shift measures (Table S4). Nevertheless, the percentages of species in each category that had experienced range contraction were different, with relatively many more range contracting species in the permanent running water group than in the other two groups (Figure 5).
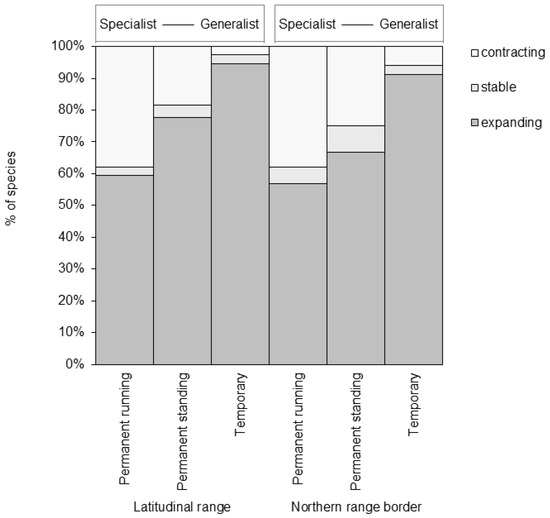
Figure 5.
Percentage of European dragonfly (Odonata) species in three ecological habitat categories—permanent running (perennial lotic) water, permanent standing (perennial lentic) water, and temporary (running and standing) water habitats—that have experienced expanding, stable, or contracting latitudinal range or northern range border between 1988 and 2010. Top scale represents a gradient going from habitat specialist with a narrow niche breadth to habitat generalist with a broad niche breadth.
4. Discussion
4.1. Data Quality
Dragonflies are among the taxa with the best data record in space and time across Europe [47], and range maps that are expert-drawn outline maps are the best currently available data for addressing macroecological questions. One drawback of range maps is that such maps may include false absences or more commonly false presences [57], thus overestimating the extent of occurrence of species [58]. Moreover, a common critique is that outline maps only represent the knowledge on distribution of the respective authors rather than the true species distribution. Some critics suggest that analysis will not show true patterns, but only changes in authors’ knowledge. Nevertheless, multiple studies on various taxonomic groups within plants, vertebrates, and invertebrates have used outline distribution to find differences in species ranges or species richness (e.g., Hawkins et al. [59]), including macroecological studies on European dragonflies similar to ours (e.g., Grewe et al. [8], Hof et al. [11], and Hof et al. [33]). Furthermore, of the 85 reviewed analyses of species richness in Hawkins et al. [59], 69% were based on range maps. We acknowledge that our maps represent rough approximations of the distribution of European dragonflies, but as demonstrated in Hurlbert et al. [60], this problem can be counteracted by using a sufficiently large grid resolution. Other range map skeptics suggest that range expansions are simply the outcome of a higher number of records. Although we cannot rule out sampling heterogeneity on continental scales, the most significant northern range border shifts were found for species that invaded Central Europe from the Mediterranean or extended their previous northern range border in Central Europe northwards. As Central Europe is one of the best sampled regions [47], we assume that a lower sampling intensity in other parts of Europe should not affect these observed range shifts. Moreover, a bias in the distribution estimates should only matter if there were strong differences in mapping accuracy between species in the three ecological habitat categories. Finally, more local studies that have reported smaller-scale range shifts in European dragonfly species based on point data of range shifts support our findings (e.g., Riservato et al. [2], Knijf et al. [3], Hassall et al. [9], Hickling et al. [10], Ott [13], Suhling et al. [15], and Termaat et al. [16]).
4.2. Do Species Reproducing in Permanent Standing Water and Temporary Water Have Larger Ranges Than Species Reproducing in Permanent Running Water?
Dragonflies in general have high dispersal ability [26], and as a result, their distribution should mainly be driven by changes in current climate and resources because their flying capabilities allow them to track changing climatic and environmental conditions. However, species distributions are not maintained in a quasi-equilibrium state with climate, and we found that species that have adapted to permanent standing water and temporary water, and that are thus more generalists in their habitat utilization than species in permanent running water, have much larger ranges than species in permanent running water. That species with broad niches have larger ranges than species with narrow niches clearly demonstrates that range dynamics in European dragonflies are heavily influenced by habitat specialization or linked dispersal limitation. The fact that permanent running water species have relatively small ranges compared with generalists could arise from their extreme habitat specialization rather than from their reduced dispersal ability. As proposed by Baselga et al. [61], habitat specialization and dispersal ability may, however, be like ‘two sides of the same coin’, where dispersal ability and habitat specialization may represent a relationship filtering species response to changes in climate and resources [62,63,64].
Compared with generalist species that reproduce in standing waters, larval adaptations to running water, which occasionally can be fast-flowing and scouring during the wet season, may include a modified body shape and a modified morphology of body parts [26]. This includes a flattened body and an enlarged contact surface with the substrate, narrow and long caudal appendages, reduced or lost swimming ability, reduced number of antenna segments, and leg spurs that allow the larvae to burrow into sandy-stone substrates [15,26]. Due to these adaptations, some running water species are strictly linked to certain microhabitats within different types of permanent running water [26]. This produces more separate and less mobile populations, thus reducing dispersal success and range expansion and thereby causing range filling in these specialist species to lag behind changes in climate and resources [8,11]. In contrast, the landscape has been, and still is, more permeable to generalist species that are adapted to either permanent standing water or temporary water habitats because migration is not restricted or blocked by unsuitable habitats as in specialists [23]. Generalists have a larger proportion of suitable habitats available and, therefore, have larger ranges that are more interconnected, which should further increase both dispersal success and range expansion. Further, as standing waters and especially temporary waters are less persistent in time compared with running waters, these habitats may also have selected for a higher dispersal ability in dragonflies adapted to standing water [11]. Our results clearly support this as generalist species with the broadest niche breadth tend to have larger ranges than permanent standing water species at the same time as they exhibit the most climate-driven response by being most prone to increase their ranges. Similarly, Bota-Sierra et al. [65] found that generalist species in the western Andes in Colombia colonized ecosystems recently created by human activities, while specialist species remain in forest ecosystems.
Due to the European topography, postglacial invasions of dragonflies may have been confined to only a few routes where especially mountain chains acted as barriers, while geotectonic depressions and wide river plains likely played important roles as corridors [66]. Current ranges of dispersal limited organisms often represent partially incomplete postglacial colonization from southern glacial refugia [61] because habitat specialization has prevented colonization in species with narrow niches. Here, the historical landscape may have filtered distributions differentially through the continuous glacial races [66,67,68]. However, besides a stepwise loss of species with increasing latitude, we find little evidence of postglacial colonization lag of European dragonflies on a continental scale [68,69]. The proportion of standing water species increases with latitude, and in addition to their larger latitudinal ranges, they have more northern distribution centers and range boundaries than running water species [33]. The differences in range size between generalist and specialist species may therefore have ecological explanations, namely, dispersal limitations connected to habitat specialization and the availability of a suitable habitat. This is especially relevant for those dragonflies that have adapted to life in permanent running water and that rely on rivers and streams for successful dispersal among water as their adaptations prevent them from utilizing other habitat types, even only as a stepping stone during colonization.
Some species that we categorized as being able to utilize temporary habitats have relatively small ranges compared with permanent standing water species and are even similar in size to some permanent running water species. This is due to the fact that temporary water species may include not only species adapted to standing water, but also species with a preference for running water that occasionally are also found in seasonal habitats. Furthermore, especially temporary water species include several Afrotropical species, which have only recently established populations north of the Mediterranean Sea and, therefore, only have a restricted current distribution from where they are colonizing southern Europe. At the moment, they are strongly dispersal limited as an effect of their small range size and not so much because they experience habitat loss and are vulnerable to climate change.
4.3. Do Species Reproducing in Temporary Habitats Track Changes Better Than Species in Permanent Habitats?
Besides an overall increase in ranges, species adapted to temporary habitats expanded their latitudinal ranges and northern range borders the most. Consequently, temporary water species migrate north much more than species bound to permanent (both running and standing) waters. This is illustrated by some of the most significant shifts in northern range borders seen in European dragonflies, such as in Anax imperator Leach, 1815; Anax parthenope (Selys, 1839); Aeshna affinis Vander Linden, 1820; Sympetrum fonscolombii (Selys, 1840); and Crocothemis erythraea (Brullé, 1832) which are all generalist species with the ability to utilize temporary water and which are known to have either invaded Central Europe from the Mediterranean or extended their previous northern range boundaries in Central Europe northwards [13,14]. Common to them, they utilize not only temporary waters, but also various permanent habitats, sometimes including both running and standing waters. Further, they can respond quickly because they can utilize the broadest range of niches, which allows them to persist during times of climate change and habitat loss through the efficient relocation to suitable habitats. The ephemeral nature of temporary habitats selects for high dispersal ability, where their shortened larval stage and rapid larval development [27] clearly provide an advantage in a rapidly changing environment compared with larval stages lasting from 1 to 6 years as in permanent water species [15,26].
4.4. Do Species Reproducing in Permanent Running Water Contract More or Expand Less Than Species in Permanent Standing Water and Temporary Water?
Of those species that experience range contraction, we found an overweight of running water species, which, on average, also showed larger contractions than species that inhabit permanent standing water or temporary water. We suggest that these larger contractions are not driven by a decrease in their realized climatic envelopes as much as by habitat loss and degradation; that is, the declines in habitat specialists are not believed to be linked to climate as much as to differential land use or eutrophication effects. Life forms are even and ever more directly affect by a continuous decline in the availability of a suitable habitat [12]. Changes (of which climate effects are just a part) due to human land use, drainage of wetland areas, pollution, eutrophication, and overgrowth of habitats may cause synergistic treats, where local populations are driven to extinction and habitat fragmentation is prompted to escalate [12]. This is believed to be the reason why Nehalennia speciosa is declining and has already gone regionally extinct in many areas across its European range [70]. Contemporary land use is believed to produce more separate and less mobile populations. Especially, distance between suitable habitats in dragonfly specialists is important for their chances of at least partially tracking changes in habitats and resources. However, migratory routes already seem to be partly interrupted over long distances due to loss of habitats, and the connections to distribution centers are also broken in some species [66]. Without suitable dispersal corridors, species responses to changes in climate and resources may not be realized [20,23]. Combined with the dramatic changes to running water environments that prompt them to dry up, especially during the dry season [12], our results suggest that permanent running water species, in particular, could face severe challenges in response to such changes. However, this effect may apply not only to species with a small range size and a narrow niche breadth, but also to less specialized species that cannot persist in temporary dry habitats.
4.5. Do Generalist Species Outperform Specialists with a Narrow Niche Breadth?
We found that specialist species in permanent running waters could be most vulnerable to habitat loss and climate change, likely due to interacting effects of a narrow niche and a small range size. Furthermore, the fact that generalist species have the largest ranges and probably track climate changes the best, and that relatively more specialist than generalist species contract ranges, could indicate that generalists are outperforming specialists. This could provide an early warning of biotic homogenization in European dragonflies similar to what has been found in several studies in which climate change response has been related to niche breadth. It is seen in plants [35], butterflies [34,37,38], frogs [32], reptiles [40], birds [19], and mammals [42], but also in other studies on dragonflies, as in North America [71] and the British Isles [36].
5. Conclusions
Habitat-defined groups of dragonfly species differ in their response to climate and environmental changes, and we suggest that species with narrow habitat requirements will be most affected by ongoing climate change and differential land use due to synergistic effects of a narrow niche and small range size. We find that species that are adapted to habitats that become temporarily dry move north to a much larger extent than species in permanent water habitats, including both running and standing waters. By this, we suggest that temporary waters support dragonfly diversity and act as stepping stones for the dispersal of generalist species. Species adapted to permanent standing water or temporary water habitats, which are less persistent in time and space than running water, disperse better than species adapted to permanent running water habitats. Species in permanent standing water and temporary water have larger ranges than specialist species in permanent running water. Additionally, relatively more species with a small range size and a narrow niche breadth living in permanent running water contract their ranges than less specialized species. This suggests that ecological specialization or dispersal limitations connected to ecological specialization may have prevented postglacial colonization in certain regions. It also suggests that specializations may cause contemporary range shifts to lag behind changes in climate and resources because the landscape may be less permeable to specialized species. Furthermore, this could provide an early indication that recent changes in climate and human land use cause biotic homogenization in the European dragonflies, where ecological specialists are outperformed and replaced by generalists.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/d14090719/s1; Table S1: List of species excluded from the analysis, where modifications to species range have been applied, which have colonized Europe after 1988, and with taxonomic and nomenclatural changes between 1988 and 2010 [71,72,73,74]; Table S2: List of species included in the analysis with individual measures of range size and range shift and ecological habitat category with references. See methods for detailed descriptions of the entries on species range in this table; Table S3: Different measures of range size and test results by ecological habitat category; Table S4: Different measures of range shift and test results for all species and range contracting species by ecological habitat category.
Author Contributions
Conceptualization, K.O., J.-C.S. and H.B.; methodology, K.O.; formal analysis, K.O.; investigation and data curation, K.O.; writing—original draft preparation, K.O.; writing—review and editing, K.O., J.-C.S. and H.B. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the Danish Ministry of Culture (TAKT2010-035), the Graduate School of Science and Technology of Aarhus University, and the Natural History Museum, Aarhus. This study is a contribution to JCS’s VILLUM Investigator project “Biodiversity Dynamics in a Changing World” funded by VILLUM FONDEN (grant 16549) and the Center for Informatics Research on Complexity in Ecology (CIRCE), funded by Aarhus University and the Aarhus University Research Foundation under the AU IDEAS program.
Institutional Review Board Statement
Not applicable.
Data Availability Statement
Data are provided in the paper and Supplementary Tables S1–S4.
Acknowledgments
We thank Brody Steven Sandel for the help with R scripts and Peder Klith Bøcher and Lonnie Mikkelsen for the ArcGIS assistance. We are particularly grateful to Melanie Bilz and Jemma Able from the Freshwater Biodiversity Unit under the IUCN Global Species Program for providing range maps from 2010 as ArcGIS shapefiles.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Bell, N.; Dylmer, E.; Olsen, K. Status of Aeshna Serrata in Denmark. Brachytron 2014, 16, 38–42. [Google Scholar]
- Riservato, E.; Boudot, J.; Ferreira, S.; Jović, M.; Kalkman, V.; Schneider, W.; Samraoui, B.; Cuttelod, A. The Status and Distribution of Dragonflies of the Mediterranean Basin; IUCN: Gland, Switzerland; Malaga, Spain, 2009. [Google Scholar]
- de Knijf, G.; Anselin, A. When South Goes North: Mediterranean Dragonflies (Odonata) Conquer Flanders (North-Belgium). BioRisk 2010, 5, 141–153. [Google Scholar] [CrossRef]
- de Kniff, G.; Anselin, A.; Goffart, P. Changes in Ranges: Invertebrates on the Move. In Proceedings of the 13th International Colloquium of the European Invertebrate Survey, Leiden, Germany, 2–5 September 2001; Reemer, M., van Helsdingen, P., Kleukers, R., Eds.; European Invertebrate Survey: Leiden, Germany, 2003; pp. 33–38. [Google Scholar]
- de Knijf, G.; Flenker, U.; Vanappelghem, C.; Manci, C.O.; Kalkman, V.J.; Demolder, H. The Status of Two Boreo-Alpine Species, Somatochlora Alpestris and S. Arctica, in Romania and Their Vulnerability to the Impact of Climate Change (Odonata: Corduliidae). Int. J. Odonatol. 2011, 14, 111–126. [Google Scholar] [CrossRef]
- Dingemanse, N.J.; Kalkman, V.J. Changing Temperature Regimes Have Advanced the Phenology of Odonata in the Netherlands. Ecol. Entomol. 2008, 33, 394–402. [Google Scholar] [CrossRef]
- Gonseth, Y.; Monnerat, C. Recent Changes in Distribution of Dragonflies in Switzerland (Odonata). In Proceedings of the 13th International Colloquium of the European Invertebrate Survey, Leiden, Germany, 2–5 September 2001; Reemer, M., van Helsdingen, P., Kleukers, R., Eds.; European Invertebrate Survey: Leiden, Germany, 2003; pp. 23–31. [Google Scholar]
- Grewe, Y.; Hof, C.; Dehling, D.M.; Brandl, R.; Brändle, M. Recent Range Shifts of European Dragonflies Provide Support for an Inverse Relationship between Habitat Predictability and Dispersal. Glob. Ecol. Biogeogr. 2013, 22, 403–409. [Google Scholar] [CrossRef]
- Hassall, C.; Thompson, D.J. The Effects of Environmental Warming on Odonata: A Review. Int. J. Odonatol. 2008, 11, 131–153. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Thomas, C.D. A Northward Shift of Range Margins in British Odonata. Glob. Change Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Hof, C.; Brändle, M.; Dehling, D.M.; Munguía, M.; Brandl, R.; Araújo, M.B.; Rahbek, C. Habitat Stability Affects Dispersal and the Ability to Track Climate Change. Biol. Lett. 2012, 8, 639–643. [Google Scholar] [CrossRef]
- Kalkman, V.J.; Boudot, J.-P.; Bernard, R.; Conze, K.-J.; de Knijf, G.; Dyatlova, E.; Ferreira, S.; Jović, M.; Ott, J.; Riservato, E.; et al. European Red List of Dragonflies; Publications Office of the European Union: Luxembourg, 2010. [Google Scholar]
- Ott, J. Dragonflies and Climatic Change—Recent Trends in Germany and Europe. BioRisk 2010, 5, 253–286. [Google Scholar] [CrossRef]
- Ott, J. Expansion of Mediterranean Odonata in Germany and Europe—Consequences of Climatic Changes. In Fingerprints of Climate Change—Adapted Behaviour and Shifting Species Ranges; Walther, G., Burga, C., Edwards, P., Eds.; Kluwer Academic; Plenum Publication: New York, NY, USA, 2001; pp. 89–111. [Google Scholar]
- Suhling, F.; Sahlén, G.; Gorb, S.; Kalkman, V.; Dijkstra, K.-D.; van Tol, J. Order Odonata. In Ecology and General Biology: Thorp and Covich’s Freshwater Invertebrates; Thorp, J., Rogers, D., Eds.; Academic Press: San Diego, CA, USA, 2015; pp. 893–932. [Google Scholar]
- Termaat, T.; Kalkman, V.; Bouwman, J. Changes in the Range of Dragonflies in the Netherlands and the Possible Role of Temperature Change. BioRisk 2010, 5, 155–173. [Google Scholar] [CrossRef]
- Lenoir, J.; Svenning, J.C. Climate-Related Range Shifts—A Global Multidimensional Synthesis and New Research Directions. Ecography 2015, 38, 15–28. [Google Scholar] [CrossRef]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Fox, R.; Thomas, C.D. The Distributions of a Wide Range of Taxonomic Groups Are Expanding Polewards. Glob. Change Biol. 2006, 12, 450–455. [Google Scholar] [CrossRef]
- Davey, C.M.; Chamberlain, D.E.; Newson, S.E.; Noble, D.G.; Johnston, A. Rise of the Generalists: Evidence for Climate Driven Homogenization in Avian Communities. Glob. Ecol. Biogeogr. 2012, 21, 568–578. [Google Scholar] [CrossRef]
- Hill, J.K.; Thomas, C.D.; Huntley, B. Climate and Habitat Availability Determine 20th Century Changes in a Butterfly’s Range Margin. Proc. R. Soc. B Biol. Sci. 1999, 266, 1197–1206. [Google Scholar] [CrossRef]
- Lancaster, L.T.; Dudaniec, R.Y.; Hansson, B.; Svensson, E.I. Latitudinal Shift in Thermal Niche Breadth Results from Thermal Release during a Climate-Mediated Range Expansion. J. Biogeogr. 2015, 42, 1953–1963. [Google Scholar] [CrossRef]
- Thomas, C.D.; Bodsworth, E.J.; Wilson, R.J.; Simmons, A.D.; Davies, Z.G.; Musche, M.; Conradt, L. Ecological and Evolutionary Processes at Expanding Range Margins. Nature 2001, 411, 577–581. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 59–71. [Google Scholar] [CrossRef]
- Channell, R.; Lomollno, M.v. Dynamic Biogeography and Conservation of Endangered Species. Nature 2000, 403, 84–86. [Google Scholar] [CrossRef]
- IPCC Climate Change 2021: The Physical Science Basis: Contribution of Working Group I to the Sixth Assessment; Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Campridge, UK; New York, NY, USA, 2021.
- Corbet, P.S. Dragonflies: Behavior and Ecology of Odonata; Harley Books: Colchester, UK, 2004. [Google Scholar]
- Corbet, P.S.; Suhling, F.; Soendgerath, D. Voltinism of Odonata: A Review. Int. J. Odonatol. 2006, 9, 1–44. [Google Scholar] [CrossRef]
- Cham, S.A. Ovipositing Behaviour and Observations on Eggs and Prolarva of Ischnura Pumilio. J. Br. Dragonfly Soc. 1992, 8, 6–10. [Google Scholar]
- de Block, M.; McPeek, M.A.; Stoks, R. Life-History Evolution When Lestes Damselflies Invaded Vernal Ponds. Evol. Ecol. 2008, 62, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Wiggins, G.; Mackay, R.; Smith, I. Evolutionary and Ecological Strategies of Animals in Annual Temporary Pools. Arch. Für Hydrobiol. Suppl. 1980, 58, 97–206. [Google Scholar]
- Fischer, Z. Food Composition and Food Preference in Larvae of Lestes sponsa (L.) in Astatic Water Environment. Pol. Arch. Hydrobiol. 1967, 14, 59–71. [Google Scholar]
- Botts, E.A.; Erasmus, B.F.N.; Alexander, G.J. Small Range Size and Narrow Niche Breadth Predict Range Contractions in South African Frogs. Glob. Ecol. Biogeogr. 2013, 22, 567–576. [Google Scholar] [CrossRef]
- Hof, C.; Brändle, M.; Brandl, R. Lentic Odonates Have Larger and More Northern Ranges than Lotic Species. J. Biogeogr. 2006, 33, 63–77. [Google Scholar] [CrossRef]
- Menéndez, R.; Megías, A.G.; Hill, J.K.; Braschler, B.; Willis, S.G.; Collingham, Y.; Fox, R.; Roy, D.B.; Thomas, C.D. Species Richness Changes Lag behind Climate Change. Proc. R. Soc. B Biol. Sci. 2006, 273, 1465–1470. [Google Scholar] [CrossRef]
- Ozinga, W.A.; Colles, A.; Bartish, I.V.; Hennion, F.; Hennekens, S.M.; Pavoine, S.; Poschlod, P.; Hermant, M.; Schaminée, J.H.J.; Prinzing, A. Specialists Leave Fewer Descendants within a Region than Generalists. Glob. Ecol. Biogeogr. 2013, 22, 213–222. [Google Scholar] [CrossRef]
- Powney, G.D.; Cham, S.S.A.; Smallshire, D.; Isaac, N.J.B. Trait Correlates of Distribution Trends in the Odonata of Britain and Ireland. PeerJ 2015, 3, e1410. [Google Scholar] [CrossRef]
- Pöyry, J.; Luoto, M.; Heikkinen, R.K.; Kuussaari, M.; Saarinen, K. Species Traits Explain Recent Range Shifts of Finnish Butterflies. Glob. Chang Biol. 2009, 15, 732–743. [Google Scholar] [CrossRef]
- Warren, M.S.; Hill, J.K.; Thomas, J.A.; Asher, J.; Fox, R.; Huntley, B.; Roy, D.B.; Telfer, M.G.; Jeffcoate, S.; Harding, P.; et al. Rapid Responses of British Butterflies to Opposing Forces of Climate and Habitat Change. Nature 2001, 414, 65–69. [Google Scholar] [CrossRef]
- Angert, A.L.; Crozier, L.G.; Rissler, L.J.; Gilman, S.E.; Tewksbury, J.J.; Chunco, A.J. Do Species’ Traits Predict Recent Shifts at Expanding Range Edges? Ecol. Lett. 2011, 14, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Foufopoulos, J.; Kilpatrick, A.M.; Ives, A.R. Climate Change and Elevated Extinction Rates of Reptiles from Mediterranean Islands. Am. Nat. 2011, 177, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Hill, J.K.; Thomas, C.D.; Fox, R.; Telfer, M.G.; Willis, S.G.; Asher, J.; Huntley, B. Responses of Butterflies to Twentieth Century Climate Warming: Implications for Future Ranges. Proc. R. Soc. B Biol. Sci. 2002, 269, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Myers, P.; Lundrigan, B.L.; Hoffman, S.M.G.; Haraminac, A.P.; Seto, S.H. Climate-Induced Changes in the Small Mammal Communities of the Northern Great Lakes Region. Glob. Change Biol. 2009, 15, 1434–1454. [Google Scholar] [CrossRef]
- Ruiz, E. Management of Natura 2000 Habitats. 3170 *Mediterranean Temporary Ponds. Directive 92/43/EEC on the Conservation of Natural Habitats and of Wild Fauna and Flora; European Commission: Luxembourg, 2008. [Google Scholar]
- Pérez-Bilbao, A.; Benetti, C.; Garrido, J. Biodiversity and Conservation of Temporary Ponds—Assessment of the Conservation Status of “Veiga de Ponteliñares”, NW Spain (Natura 2000 Network), Using Freshwater Invertebrates. In Biodiversity in Ecosystems—Linking Structure and Function; Blanco, J., Lo, Y.-H., Eds.; InTech: London, UK, 2015; pp. 241–269. [Google Scholar]
- Askew, R.R. The Dragonflies of Europe; Harley Books (BH & A Harley Ltd.): Colchester, UK, 1988. [Google Scholar]
- ESRI ArcGIS Desktop v. 10.2 [GIS Software]; Environmental Systems Research Institute: Redlands, CA, USA, 2010.
- Dijkstra, K.-D.; Lewington, R. Field Guide to the Dragonflies of Britain and Europe; British Wildlife Publishing: Totnes, UK, 2006. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2015-3. Available online: www.iucnredlist.org (accessed on 17 May 2016).
- Sternberg, K.; Buchwald, R. Die Libellen Baden-Württembergs: Grosslibellen (Anisoptera); Eugen Ulmer: Stuttgart, Germany, 2000; Volume 2. [Google Scholar]
- Sternberg, K.; Buchwald, R. Die Libellen Baden-Württembergs: Allgemeiner Teil, Kleinlibellen (Zygoptera); Eugen Ulmer: Stuttgart, Germany, 1999; Volume 1. [Google Scholar]
- Beschovski, V.; Marinov, M. Fauna, Ecology, and Zoogeography of Dragonflies (Insecta: Odonata) of Bulgaria. In Biogeography and Ecology of Bulgaria; Fet, V., Popov, A., Eds.; Springer: Dordrecht, Germany, 2007; pp. 199–231. [Google Scholar]
- Carchini, G.; della Bella, V.; Solimini, A.G.; Bazzanti, M. Relationships between the Presence of Odonate Species and Environmental Characteristics in Lowland Ponds of Central Italy. Ann. Limnol. Int. J. Limnol. 2007, 43, 81–87. [Google Scholar] [CrossRef]
- Koli, V.K.; Bhatnagar, C.; Shekhawat, D.S. Diversity and Species Composition of Odonates in Southern Rajasthan, India. Proc. Zool. Soc. 2015, 68, 202–211. [Google Scholar] [CrossRef]
- Suhling, F.; Jödicke, R.; Schneider, W. Odonata of African Arid Regions—Are There Desert Species? Cimbebasia 2003, 18, 207–224. [Google Scholar]
- Florencio, M.; Díaz-Paniagua, C. Presencia de Lestes Macrostigma (Eversmann, 1836) (Odonata: Lestidae) En Las Lagunas Temporales Del Parque Nacional de Doñana (Sudoeste de España). Boletín Soc. Entomológica Aragonesa 2012, 50, 579–581. [Google Scholar]
- R Core TeAm. R: A Language and Environment for Statistical Computing; R Core TeAm: Vienna, Austria, 2022. [Google Scholar]
- Rondinini, C.; Wilson, K.A.; Boitani, L.; Grantham, H.; Possingham, H.P. Tradeoffs of Different Types of Species Occurrence Data for Use in Systematic Conservation Planning. Ecol. Lett. 2006, 9, 1136–1145. [Google Scholar] [CrossRef]
- Jetz, W.; Sekercioglu, C.H.; Watson, J.E.M. Ecological Correlates and Conservation Implications of Overestimating Species Geographic Ranges. Conserv. Biol. 2008, 22, 110–119. [Google Scholar] [CrossRef]
- Hawkins, B.A.; Field, R.; Cornell, H.V.; Currie, D.J.; Guégan, J.F.; Kaufman, D.M.; Kerr, J.T.; Mittelbach, G.G.; Oberdorff, T.; O’Brien, E.M.; et al. Energy, Water, and Broad-Scale Geographic Patterns of Species Richness. Ecology 2003, 84, 3105–3117. [Google Scholar] [CrossRef]
- Hurlbert, A.H.; Jetz, W. Species Richness, Hotspots, and the Scale Dependence of Range Maps in Ecology and Conservation. Proc. Natl. Acad. Sci. USA 2007, 104, 13384–13389. [Google Scholar] [CrossRef] [PubMed]
- Baselga, A.; Lobo, J.M.; Svenning, J.C.; Aragón, P.; Araújo, M.B. Dispersal Ability Modulates the Strength of the Latitudinal Richness Gradient in European Beetles. Glob. Ecol. Biogeogr. 2012, 21, 1106–1113. [Google Scholar] [CrossRef]
- Jocque, M.; Field, R.; Brendonck, L.; de Meester, L. Climatic Control of Dispersal-Ecological Specialization Trade-Offs: A Metacommunity Process at the Heart of the Latitudinal Diversity Gradient? Glob. Ecol. Biogeogr. 2010, 19, 244–252. [Google Scholar] [CrossRef]
- Abbott, J.C.; Bota-Sierra, C.A.; Guralnick, R.; Kalkman, V.; González-Soriano, E.; Novelo-Gutiérrez, R.; Bybee, S.; Ware, J.; Belitz, M.W. Diversity of Nearctic Dragonflies and Damselflies (Odonata). Diversity 2022, 14, 575. [Google Scholar] [CrossRef]
- Bota-Sierra, C.A.; García-Robledo, C.; Escobar, F.; Novelo-Gutiérrez, R.; Londoño, G.A. Environment, Taxonomy and Morphology Constrain Insect. Thermal Physiology along Tropical Mountains. Funct. Ecol. 2022, 36, 1924–1935. [Google Scholar] [CrossRef]
- Bota-Sierra, C.A.; Flórez-V, C.; Escobar, F.; Sandoval-H, J.; Novelo-Gutiérrez, R.; Londoño, G.A.; Cordero-Rivera, A. The Importance of Tropical Mountain Forests for the Conservation of Dragonfly Biodiversity: A Case from the Colombian Western Andes. Int. J. Odonatol. 2021, 24, 233–247. [Google Scholar] [CrossRef]
- Sternberg, K. Die Postglaziale Besiedlung Mitteleuropas Durch Libellen, Mit Besonderer Berucksichtigung Sudwestdeutschlands (Insecta, Odonata). The Postglacial Colonization of Central Europe by Dragonflies, with Special Reference to Southwestern Germany (Insecta, Odonata). J. Biogeogr. 1998, 25, 319–337. [Google Scholar] [CrossRef]
- Corser, J.D.; White, E.L.; Schlesinger, M.D. Odonata Origins, Biogeography, and Diversification in an Eastern North American Hotspot: Multiple Pathways to High Temperate Forest Insect. Diversity. Insect. Conserv. Divers. 2014, 7, 393–404. [Google Scholar] [CrossRef]
- Heiser, M.; Schmitt, T. Do Different Dispersal Capacities Influence the Biogeography of the Western Palearctic Dragonflies (Odonata)? Biol. J. Linn. Soc. 2010, 99, 177–195. [Google Scholar] [CrossRef]
- Keil, P.; Simova, I.; Hawkins, B.A. Water-Energy and the Geographical Species Richness Pattern of European and North African Dragonflies (Odonata). Insect. Conserv. Divers. 2008, 1, 142–150. [Google Scholar] [CrossRef]
- Bernard, R.; Wildermuth, H. Nehalennia Speciosa (Charpentier, 1840) in Europe: A Case of a Vanishing Relict (Zygoptera: Coenagrionidae). Odonatologica 2005, 34, 335–378. [Google Scholar]
- Ball-Damerow, J.E.; M’Gonigle, L.K.; Resh, V.H. Changes in Occurrence, Richness, and Biological Traits of Dragonflies and Damselflies (Odonata) in California and Nevada over the Past Century. BioDivers. Conserv. 2014, 23, 2107–2126. [Google Scholar] [CrossRef]
- Boudot, J.-P.; Kalkman, V.J.; Azpilicueta Amorin, M.; Bogdanović, T.; Cordero Rivera, A.; Degabriele, G.; Dommanget, J.-L.; Ferreira, S.; Garrigos, B.; Jović, M.; et al. Atlas of the Odonata of the Mediterranean and North Africa. Libellula Suppl. 2009, 9, 1–256. [Google Scholar]
- Aguesse, P. Les Odonates de l’Europe Occidentale, Du Nord de l’Afrique et Des Îles Atlantiques. In Faune de l’Europe et du Bassin Méditerranéen; Masson et Cie: Paris, France, 1968; Volume 4, pp. 1–258. [Google Scholar]
- Pilgrim, E.M.; von Dohlen, C.D. Molecular and Morphological Study of Species-Level Questions within the Dragonfly Genus Sympetrum (Odonata: Libellulidae). Ann. Entomol. Soc. Am. 2007, 100, 688–702. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).