
Deep-Sea Epibenthic Megafaunal Assemblages of the Falkland Islands, Southwest Atlantic


Abstract
:1. Introduction
2. Materials and Methods
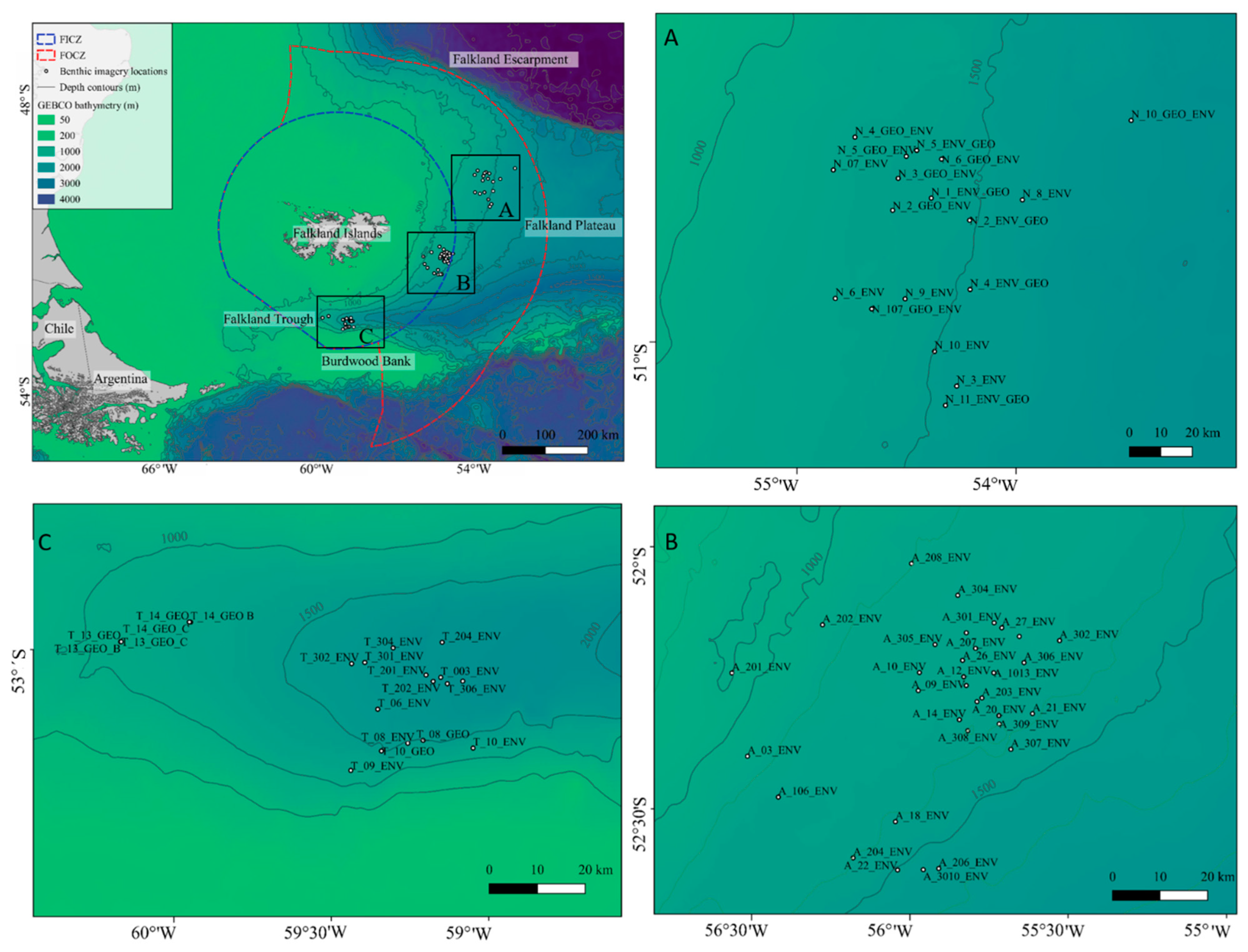
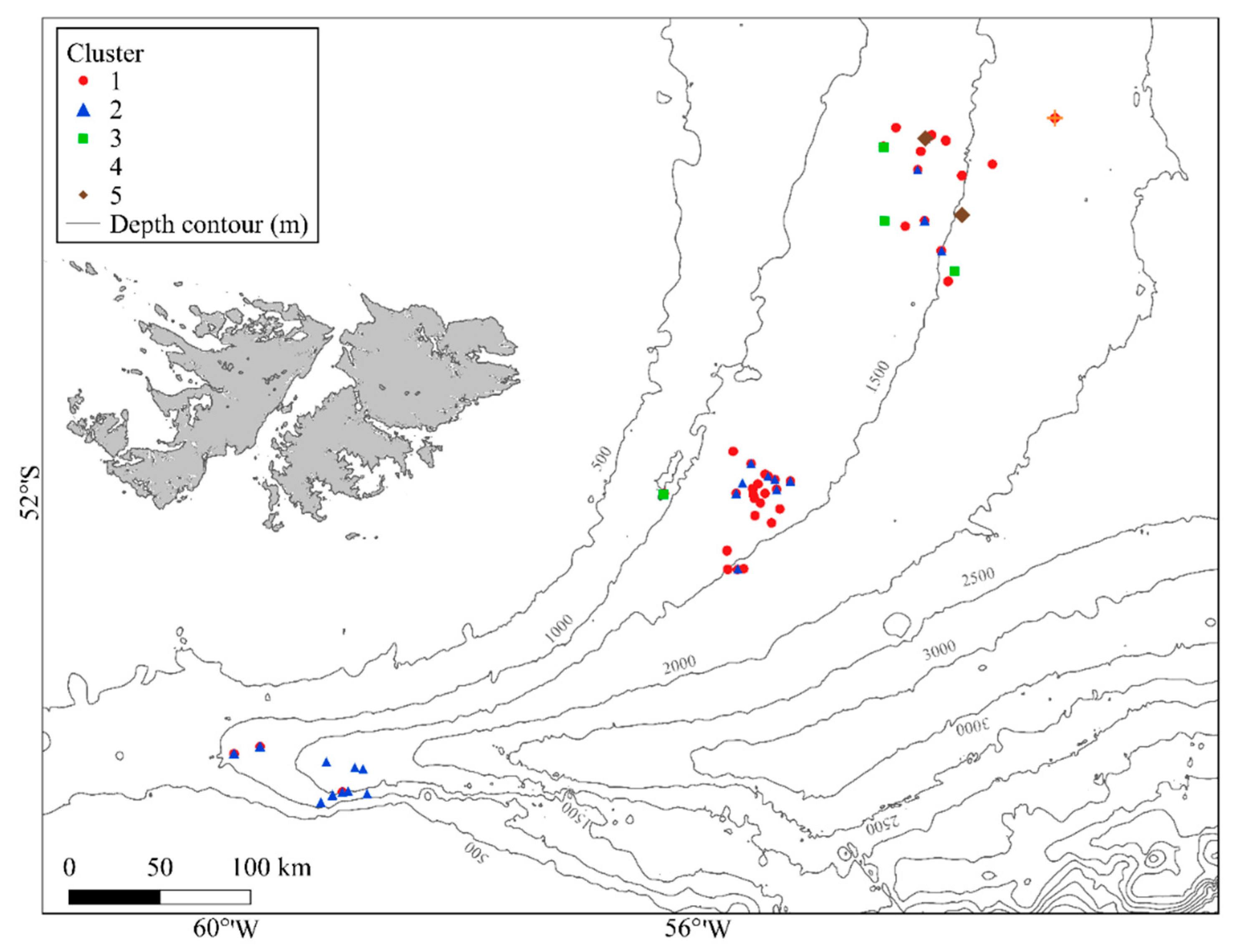
2.1. Study Area
2.2. Quantitative Image Analysis
2.3. Data Analysis
3. Results
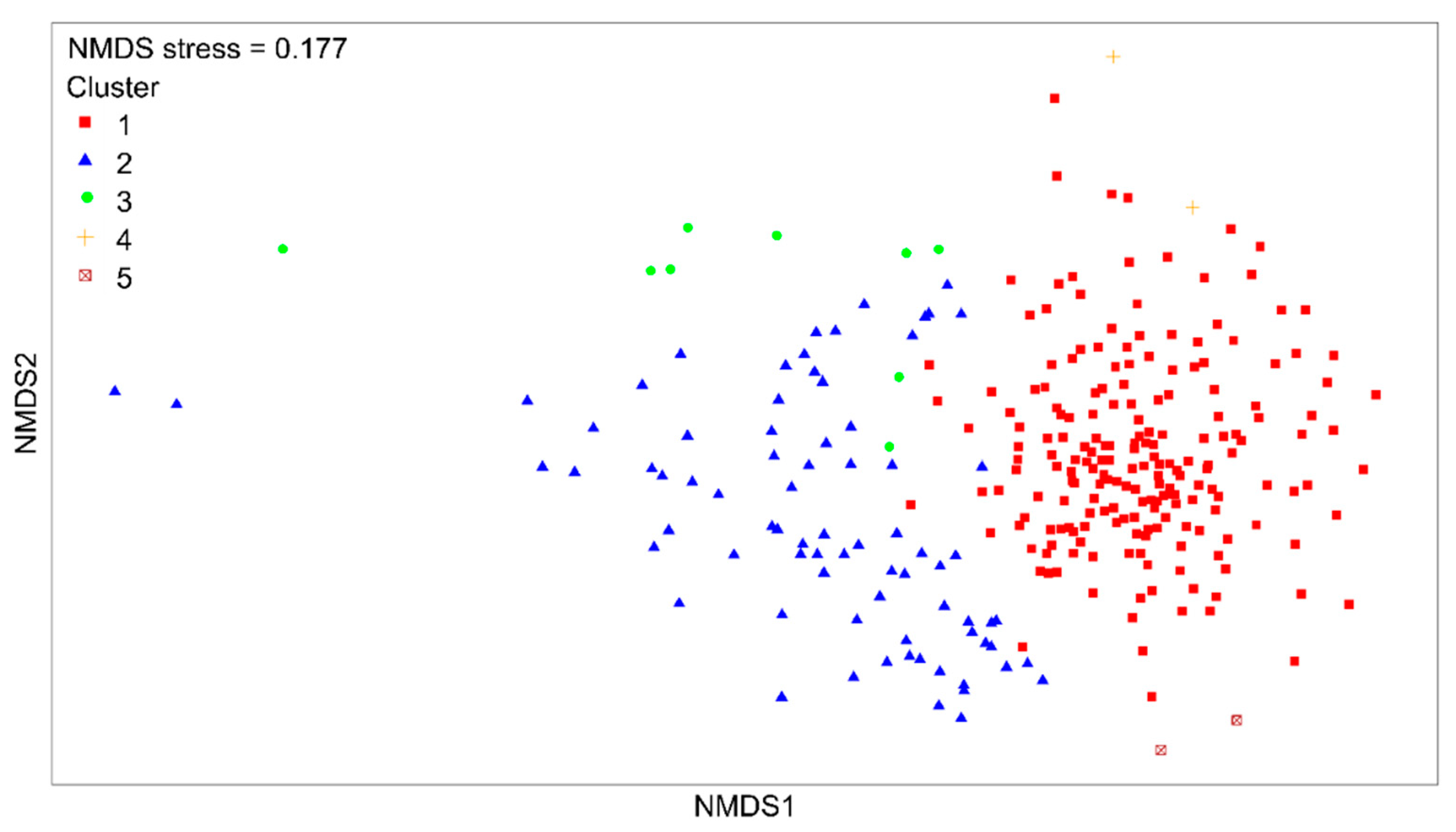
3.1. Quantitative Analysis of Epibenthic Megafaunal Assemblages
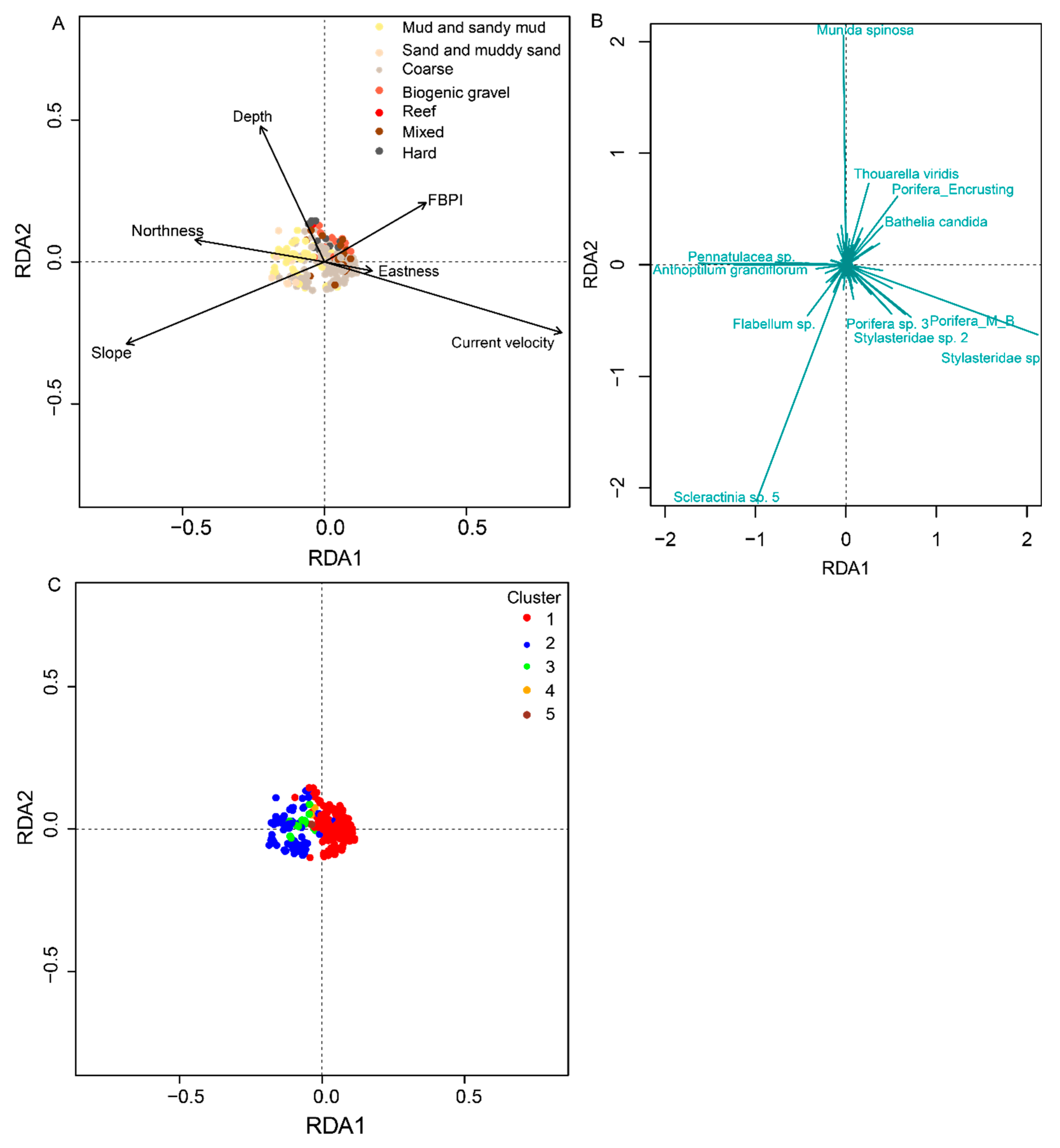
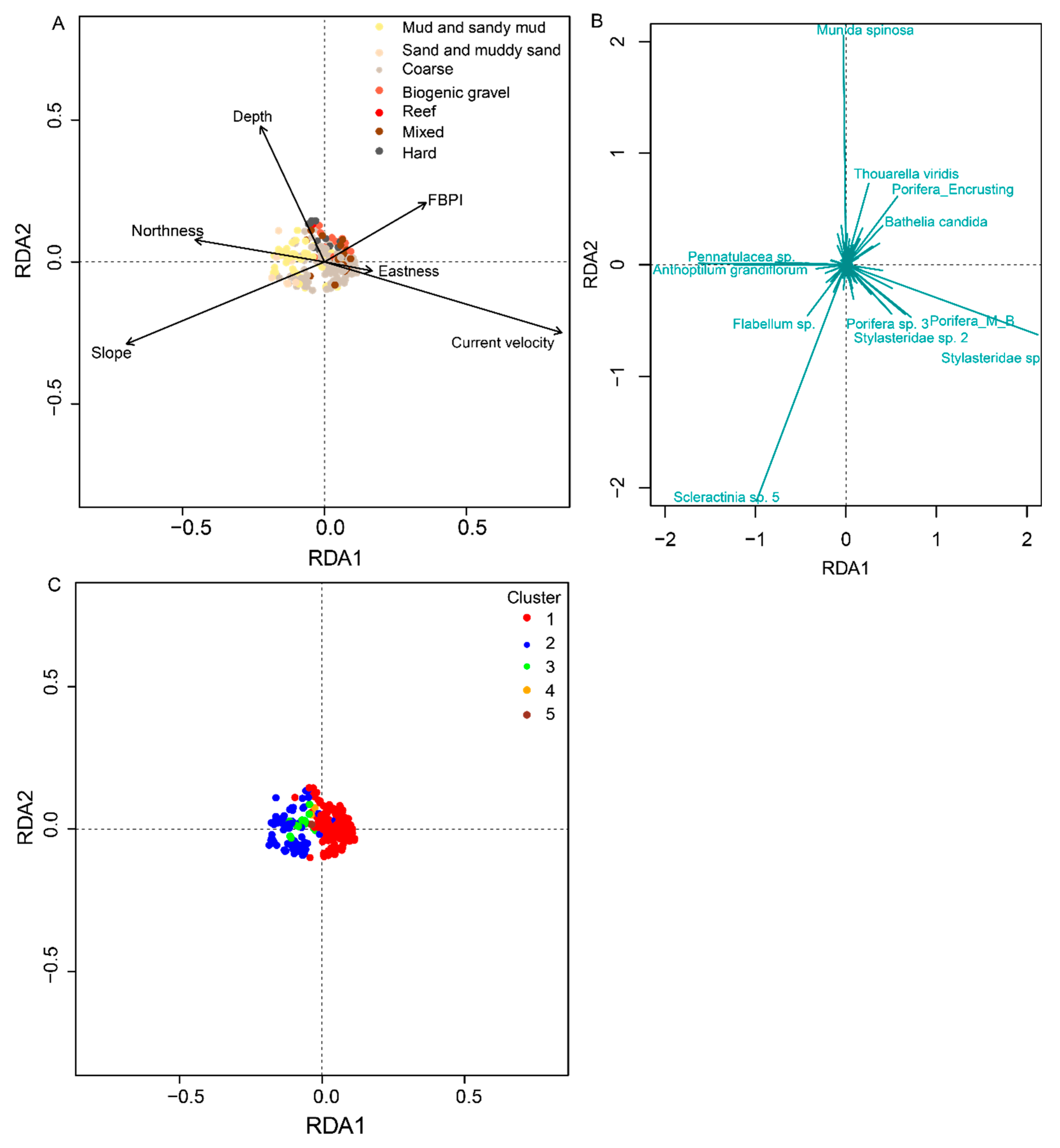
3.2. Environmental Drivers of Faunal Assemblages
4. Discussion
4.1. Deep-Sea Epibenthic Megafaunal Assemblages of the Falkland Islands
4.2. Environmental Drivers of Faunal Assemblages
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davies, A.; Roberts, M.; Hall-Spencer, J. Preserving Deep-Sea Natural Heritage: Emerging Issues in Offshore Conservation and Management. Biol. Conserv. 2007, 138, 229–312. [Google Scholar] [CrossRef]
- Puig, P.; Canals, M.; Company, J.; Martin, J.; Amblas, A.; Lastras, G.; Palanques, A.; Calafat, A. Ploughing the Deep Sea Floor. Nature 2012, 489, 286–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dover, C.L.; Aronson, J.; Pendleton, L.; Smith, S.; Arnaud-Haond, S.; Moreno-Mateos, D.; Barbier, E.; Billett, D.; Bowers, K.; Danovaro, R.; et al. Ecological Restoration in the Deep Sea: Desiderata. Mar. Policy 2014, 44, 98–106. [Google Scholar] [CrossRef]
- Clark, M.R.; Althaus, F.; Schlacher, T.A.; Williams, A.; Bowden, D.A.; Rowden, A.A. The Impacts of Deep-Sea Fisheries on Benthic Communities: A Review. ICES J. Mar. Sci. 2016, 73, i51–i69. [Google Scholar] [CrossRef]
- Miller, K.A.; Thompson, K.F.; Johnston, P.; Santillo, D. An Overview of Seabed Mining Including the Current State of Development, Environmental Impacts, and Knowledge Gaps. Front. Mar. Sci. 2018, 4, 418. [Google Scholar] [CrossRef]
- Danovaro, R.; Dell’Anno, A.; Fabiano, M.; Pusceddu, A.; Tselepides, A. Deep-Sea Ecosystem Response to Climate Changes: The Eastern Mediterranean Case Study. Trends Ecol. Evol. 2001, 16, 505–510. [Google Scholar] [CrossRef]
- Ramirez-Llodra, E.; Tyler, P.A.; Baker, M.C.; Bergstad, O.A.; Clark, M.R.; Escobar, E.; Levin, L.A.; Menot, L.; Rowden, A.A.; Smith, C.R.; et al. Man and the Last Great Wilderness: Human Impact on the Deep Sea. PLoS ONE 2011, 6, e22588. [Google Scholar] [CrossRef] [Green Version]
- Sweetman, A.K.; Thurber, A.R.; Smith, C.R.; Levin, L.A.; Mora, C.; Wei, C.; Gooday, A.J.; Jones, D.O.B.; Rex, M.; Yasuhara, M.; et al. Major Impacts of Climate Change on Deep-Sea Benthic Ecosystems. Elem. Sci. Anthr. 2017, 5, 4. [Google Scholar] [CrossRef]
- Levin, L.A.; Wei, C.L.; Dunn, D.C.; Amon, D.J.; Ashford, O.S.; Cheung, W.W.L.L.; Colaço, A.; Dominguez-Carrió, C.; Escobar, E.G.; Harden-Davies, H.R.; et al. Climate Change Considerations Are Fundamental to Management of Deep-Sea Resource Extraction. Glob. Chang. Biol. 2020, 26, 4664–4678. [Google Scholar] [CrossRef]
- Danovaro, R.; Fanelli, E.; Canals, M.; Ciuffardi, T.; Marie-claire, F.; Taviani, M. Archimer Towards a Marine Strategy for the Deep Mediterranean Sea: Analysis of Current Ecological Status. Mar. Policy 2020, 112, 103781. [Google Scholar] [CrossRef]
- UNGA United Nations General Assembly. Sustainable Fisheries, Including through the 1995 Agreement for the Implementation of the Provisions of the United Nations Convention on the Law of the Sea of 10 December 1982 Relating to the Conservation and Management of Straddling Fish Stocks and Highly Migratory Fish Stocks, and Related Instruments; General Assembly Resolution 64/72, A/RES/64/72; UNGA: New York, NY, USA, 2009. [Google Scholar]
- FAO. International Guidelines for the Management of Deep-Sea Fisheries in the High Seas; FAO: Rome, Italy, 2009; 73p. [Google Scholar]
- Doxa, A.; Almpanidou, V.; Katsanevakis, S.; Queiros, A.M.; Kaschner, K.; Garilao, C.; Kesner-Reyes, K.; Mazaris, A.D. 4D Marine Conservation Networks: Combining 3D Prioritization of Present and Future Biodiversity with Climatic Refugia. Glob. Chang. Biol. 2022, 28, 4577–4588. [Google Scholar] [CrossRef] [PubMed]
- Combes, M.; Vaz, S.; Grehan, A.; Morato, T. Systematic Conservation Planning at an Ocean Basin Scale: Identifying a Viable Network of Deep-Sea Protected Areas in the North Atlantic and the Systematic Conservation Planning at an Ocean Basin Scale: Identifying a Viable Network of Deep-Sea Protected. Front. Mar. Sci. 2021, 8, 611358. [Google Scholar] [CrossRef]
- Danovaro, R.; Company, J.B.; Corinaldesi, C.; Onghia, G.D.; Galil, B.; Gambi, C.; Gooday, A.J.; Lampadariou, N.; Luna, G.M.; Morigi, C.; et al. Deep-Sea Biodiversity in the Mediterranean Sea: The Known, the Unknown, and the Unknowable. PLoS ONE 2010, 5, e11832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martinez Arbizu, P.; Menot, L.; Buhl-Mortensen, P.; et al. Deep, Diverse and Definitely Different: Unique Attributes of the World’s Largest Ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef] [Green Version]
- Schejter, L.; Bremec, C.; Genzano, G.; Gaitán, E.; Perez, C. Benthic Communities in the Southwest Atlantic Ocean: Conservation Value of Animal Forests at the Burdwood Bank Slope. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 426–439. [Google Scholar] [CrossRef]
- Bernal, M.C.; Cairns, S.D.; Penchaszadeh, P.E.; Lauretta, D. Stylasterids (Hydrozoa: Stylasteridae) from Mar Del Plata Submarine Canyon and Adjacent Area (Southwestern Atlantic), with a Key to the Species off Argentina. Zootaxa 2021, 4969, 401–452. [Google Scholar] [CrossRef]
- Lauretta, D.; Martinez, M.I. Corallimorpharians (Anthozoa: Corallimorpharia) from the Argentinean Sea. Zootaxa 2019, 4688, 249–263. [Google Scholar] [CrossRef]
- Fernandez, M.O.; Collins, A.G.; Marques, A.C. Gradual and Rapid Shifts in the Composition of Assemblages of Hydroids (Cnidaria) along Depth and Latitude in the Deep Atlantic Ocean. J. Biogeogr. 2020, 47, 1541–1551. [Google Scholar] [CrossRef]
- Cordeiro, R.T.; Neves, B.M.; Kitahara, M.V.; Arantes, R.C.; Perez, C.D. First Assessment on Southwestern Atlantic Equatorial Deep-Sea Coral Communities. Deep. Res. Part I Oceanogr. Res. Pap. 2020, 163, 103344. [Google Scholar] [CrossRef]
- Cairns, S.D. Antarctic and Subantarctic Scleractinia. Antarct. Res. Ser. 1983, 34, 1–74. [Google Scholar] [CrossRef]
- Cairns, S.D.; Polonio, V. New Records of Deep-Water Scleractinia off Argentina and the Falkland Islands. Zootaxa 2013, 3691, 58–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinmann, L.; Baques, M.; Wenau, S.; Schwenk, T.; Spiess, V.; Piola, A.R.; Bozzano, G. Discovery of a Giant Cold-Water Coral Mound Province along the Northern Argentine Margin and Its Link to the Regional Contourite Depositional System and Oceanographic Setting. Mar. Geol. 2020, 427, 106223. [Google Scholar] [CrossRef]
- Sumida, G.; Yukio, M.; Antonio, L.; Madureira, S.; Hovland, M.; Sumida, P.Y.G.; Yoshinaga, M.Y.; Madureira, S.; Hovland, M. Seabed Pockmarks Associated with Deepwater Corals off SE Brazilian Continental Slope, Santos Basin. Mar. Geol. 2004, 207, 159–167. [Google Scholar] [CrossRef]
- Arantes, R.C.M.; Castro, C.B.; Pires, D.O.; Seoane, J.C.S. Depth and Water Mass Zonation and Species Associations of Cold-Water Octocoral and Stony Coral Communities in the Southwestern Atlantic. Mar. Ecol. Prog. Ser. 2009, 397, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Sumida, P.Y.G.; Bernardino, A.F.; Léo De, F.C. Brazilian Deep-Sea Biodiversity; Springer: Cham, Switzerland, 2020; ISBN 2520-1077. [Google Scholar]
- Saeedi, H.; Bernardino, A.F.; Shimabukuro, M.; Falchetto, G.; Sumida, P.Y. Macrofaunal Community Structure and Biodiversity Patterns Based on a Wood-Fall Experiment in the Deep South-West Atlantic. Deep Sea Res. Part I Oceanogr. Res. Pap. 2019, 145, 73–82. [Google Scholar] [CrossRef]
- Muñoz, A.; Cristobo, J.; Rios, P.; Druet, M.; Polonio, V.; Uchupi, E.; Acosta, J.; Group, A. Sediment Drifts and Cold-Water Coral Reefs in the Patagonian Upper and Middle Continental Slope. Mar. Pet. Geol. 2012, 36, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Bremec, C.; Scientific, N.; Schejter, L.; Giberto, D. Benthic Invertebrates By-Catch of Demersal Fisheries: A Comparison between Subantactic and Antarctic Shelf Waters (45° S–57° S). Ber. Polar-Meeresforsch. 2005, 507, 179–181. [Google Scholar]
- Oliver, M. Informe de la Campana de Investigacion Pesquera Atlantis 2009; Vigo Oceanographic Center, Spanish Institute of Oceanography: Madrid, Spain, 2009. [Google Scholar]
- Brewin, P.E.; Farrugia, T.J.; Jenkins, C.; Brickle, P. Straddling the Line: High Potential Impact on Vulnerable Marine Ecosystems by Bottom-Set Longline Fishing in Unregulated Areas beyond National Jurisdiction. ICES J. Mar. Sci. 2020, 78, 2132–2145. [Google Scholar] [CrossRef]
- Schejter, L.; Albano, M. Benthic Communities at the Marine Protected Area Namuncurá/Burdwood Bank, SW Atlantic Ocean: Detection of Vulnerable Marine Ecosystems and Contributions to the Assessment of the Rezoning Process. Polar Biol. 2021, 44, 2023–2037. [Google Scholar] [CrossRef]
- Gabriel, E.; Tatia, M.; Lo, J.; Perez, C.D.; Cordeiro, R.T.; Bremec, C.S.; Schejter, L.; Rimondino, C.; Chiesa, I.; Díaz de Astarloa, J.M.; et al. Namuncurá Marine Protected Area: An Oceanic Hot Spot of Benthic Biodiversity at Burdwood Bank, Argentina. Polar Biol. 2016, 39, 2373–2386. [Google Scholar] [CrossRef] [Green Version]
- Schejter, L.; Acuña, F.H.; Garese, A.; Cordeiro, R.T.S.; Pérez, C.D. Sea Pens (Cnidaria: Pennatulacea) from Argentine Waters: New Distributional Records and First Report of Associated Anemones. Pan-Am. J. Aquat. Sci. 2018, 13, 292–301. [Google Scholar]
- Cristobo, J.; Portela, J.R.; Acosta, J.; Ríos, P.; Luis, J.; Yepes, J.A. La Investigación de Los Ecosistemas Marinos Vulnerables En Aguas Internacionales Del Atlántico Suroccidental y de Las Posibles Interacciones Con Las Actividades Pesqueras: El Proyecto ATLANTIS. In Proceedings of the XVI Simposio Ibérico de Estudios de Biología Marina, Alicante, Spain, 6–10 September 2010. [Google Scholar]
- Downie, A.L.; Vieira, R.P.; Hogg, O.T.; Darby, C. Distribution of Vulnerable Marine Ecosystems at the South Sandwich Islands: Results from the Blue Belt Discovery Expedition 99 Deep-Water Camera Surveys. Front. Mar. Sci. 2021, 8, 652. [Google Scholar] [CrossRef]
- Portela, J.; Acosta, J.; Cristobo, J.; Muoz, A.; Parra, S.; Ibarrola, T.; Del Ro, J.L.; Vilela, R.; Ros, P.; Blanco, R.; et al. Management Strategies to Limit the Impact of Bottom Trawling on VMEs in the High Seas of the SW Atlantic. In Marine Ecosystems; IntECH: Rijeka, Croatia, 2012; pp. 199–228. [Google Scholar]
- Portela, J.; Cristobo, J.; Ríos, P.; Acosta, J.; Parra, S. A First Approach to Assess the Impact of Bottom Trawling over Vulnerable Marine Ecosystems on the High Seas of the Southwest Atlantic. In Biodiversity in Ecosystems: Linking Structure and Function; IntECH: Rijeka, Croatia, 2015; pp. 271–298. [Google Scholar] [CrossRef] [Green Version]
- del Rio, J.; Acosta, J.; Cristobo, J.; Portela, J.; Al, E. Estudio de Los Ecosistemas Marinos Vulnerables en Aguas Internacionales del Atlántico Sudoccidental; Instituto Español de Oceanografía: Madrid, Spain, 2012. [Google Scholar]
- Teso, V.; Urteaga, D.; Pastorino, G. Assemblages of Certain Benthic Molluscs along the Southwestern Atlantic: From Subtidal to Deep Sea. BMC Ecol. 2019, 19, 49. [Google Scholar] [CrossRef] [Green Version]
- Schejter, L.; Bremec, C.S.; Cairns, S.D. Scleractinian corals recorded in the Argentinean Antarctic expeditions between 2012 and 2014, with comments on Flabellum (Flabellum) areum Cairns, 1982. Polar Res. 2016, 35, 29762. [Google Scholar] [CrossRef] [Green Version]
- OBIS. Available online: http://www.iobis.org/ (accessed on 14 July 2021).
- Williams, A.; Althaus, F.; Schlacher, T.A. Towed Camera Imagery and Benthic Sled Catches Provide Different Views of Seamount Benthic Diversity. Limnol. Oceanogr.-Methods 2015, 13, 62–73. [Google Scholar] [CrossRef] [Green Version]
- Schejter, L.; Bremec, C.S.; Ocean, S.W.A.; Schejter, L.; Bremec, C.S. Stony Corals (Anthozoa:Scleractinia) of Burdwood Bank and Neighbouring Stony Corals (Anthozoa:Scleractinia) of Burdwood Bank and Neighbouring Areas, SW Atlantic Ocean. Sci. Mar. 2019, 83, 247–260. [Google Scholar] [CrossRef]
- Neal, L.; Paterson, G.L.J.J.; Blockley, D.; Scott, B.; Sherlock, E.; Huque, C.; Glover, A.G. Biodiversity Data and New Species Descriptions of Polychaetes from Offshore Waters of the Falkland Islands, an Area Undergoing Hydrocarbon Exploration. Zookeys 2020, 938, 1–86. [Google Scholar] [CrossRef]
- Perez, C.D.; Zamponi, M.O. New Records of Octocorals (Cnidaria, Anthozoa) from the South Western Atlantic Ocean, with Zoogeographic Considerations. Zootaxa 2004, 630, 1–12. [Google Scholar] [CrossRef]
- Kitahara, M.V. Species Richness and Distribution of Azooxanthellate Scleractinia in Brazil Species Richness and Distribution of Azooxanthellate Scleractinia in Brazil. Bull. Mar. Sci. 2015, 81, 497–518. [Google Scholar]
- Portela, J.; Acosta, J.; Parra-Descalzo, S.; Cristobo, J.; Tel, E.; Río-Iglesias, J.L.; Pereiro-Muñoz, J.A.; Elvira, E.; Patrocinio, T.; Ríos, P.; et al. Proyecto ATLANTIS: Informe Preliminar Sobre Ecosistemas Marinos Vulnerables en Aguas Internacionales del Atlántico Sudoccidental y de las Posibles Interacciones con la Actividad Pesqueras; Programa Pesquerías Lejanas: Madrid, Spain, 2009. [Google Scholar]
- Durden, J.; Schoening, T.; Althaus, F.; Friedman, A.; Garcia, R.; Glover, A.; Greinert, J.; Stout, N.; Jones, D.; Jordt, A.; et al. Perspectives in Visual Imaging for Marine Biology and Ecology: From Acquisition to Understanding. Oceanogr. Mar. Biol. 2016, 54, 510. [Google Scholar]
- Price, D.M.; Robert, K.; Callaway, A.; Lo Lacono, C.; Hall, R.A.; Huvenne, V.A.I. Using 3D Photogrammetry from ROV Video to Quantify Cold-Water Coral Reef Structural Complexity and Investigate Its Influence on Biodiversity and Community Assemblage. Coral Reefs 2019, 38, 1007–1021. [Google Scholar] [CrossRef] [Green Version]
- Bridges, A.E.H.; Barnes, D.K.A.; Bell, J.B.; Ross, R.E.; Howell, K.L. Benthic Assemblage Composition of South Atlantic Seamounts. Front. Mar. Sci. 2021, 8, 1530. [Google Scholar] [CrossRef]
- Davies, J.S.; Howell, K.L.; Stewart, H.A.; Guinan, J.; Golding, N. Defining Biological Assemblages (Biotopes) of Conservation Interest in the Submarine Canyons of the South West Approaches (Offshore United Kingdom) for Use in Marine Habitat Mapping. Deep. Res. Part II Top. Stud. Oceanogr. 2014, 104, 208–229. [Google Scholar] [CrossRef] [Green Version]
- Almond, P.M.; Linse, K.; Dreutter, S.; Grant, S.M.; Griffiths, H.J.; Whittle, R.J.; Mackenzie, M.; Reid, W.D.K.K. In-Situ Image Analysis of Habitat Heterogeneity and Benthic Biodiversity in the Prince Gustav Channel, Eastern Antarctic Peninsula. Front. Mar. Sci. 2021, 8, 614496. [Google Scholar] [CrossRef]
- Van Audenhaege, L.; Broad, E.; Hendry, K.R.; Huvenne, V.A.I. High-Resolution Vertical Habitat Mapping of a Deep-Sea Cliff Offshore Greenland. Front. Mar. Sci. 2021, 8, 621. [Google Scholar] [CrossRef]
- Robert, K.; Jones, D.O.B.; Huvenne, V.A.I. Megafaunal Distribution and Biodiversity in a Heterogeneous Landscape: The Megafaunal Distribution and Biodiversity in a Heterogeneous Landscape: The Iceberg-Scoured Rockall Bank, NE Atlantic. Mar. Ecol. Prog. Ser. 2014, 501, 67–88. [Google Scholar] [CrossRef] [Green Version]
- Orejas, C.; Gori, A.; Lo Iacono, C.; Puig, P.; Gili, J.M.; Dale, M.R.T.; Lo Iacono, C.; Puig, P. Cold-Water Corals in the Cap de Creus Canyon, Northwestern Mediterranean: Spatial Distribution, Density and Anthropogenic Impact. Mar. Ecol. Prog. Ser. 2009, 397, 37–51. [Google Scholar] [CrossRef]
- Orejas, C.; Wienberg, C.; Titschack, J.; Tamborrino, L. Madrepora Oculata Forms Large Frameworks in Hypoxic Waters off Angola (SE Atlantic). Sci. Rep. 2021, 11, 15170. [Google Scholar] [CrossRef]
- Beazley, L.; Kenchington, E.; Yashayaev, I.; Murillo, F.J. Drivers of epibenthic megafaunal composition in the sponge grounds of the Sackville Spur, northwest Atlantic. Deep Sea Res. Part I Oceanogr. Res. Pap. 2017, 98, 102–114. [Google Scholar] [CrossRef]
- NOBLE. Noble Energy. South Falklands Basin Environmental and Geochemical Program 2014—FIST Environmental Baseline and Habitat Survey Report; NOBLE: London, UK, 2014. [Google Scholar]
- NOBLE. Noble Energy. South Falklands Basin Environmental and Geochemical Program 2014—FISA Environmental Baseline and Habitat Survey Report; NOBLE: London, UK, 2014. [Google Scholar]
- NOBLE. Noble Energy. South Falklands Basin Environmental and Geochemical Program 2014—FINA Environmental Baseline and Habitat Survey Report; NOBLE: London, UK, 2014. [Google Scholar]
- FOGL Falkland Oil and Gas Ltd. Scotia East Site Survey March 2011 Survey Report 8668; FOGL Falkland Oil and Gas Ltd.: London, UK, 2011. [Google Scholar]
- FOGL Falkland Oil and Gas Ltd. Loligo Northwest Site Survey March 2011 Survey Report 8751; FOGL Falkland Oil and Gas Ltd.: London, UK, 2011. [Google Scholar]
- FOGL Falkland Oil and Gas Ltd. Hero Site Survey March 2011 Survey Report 8667; FOGL Falkland Oil and Gas Ltd.: London, UK, 2011. [Google Scholar]
- Schejter, L. Unveiling the Submarine Landscape of the Namuncurá Marine Protected Area, Burdwood Bank, SW Atlantic Ocean; P248-253 Instituto Nacional de Investigación y Desarrollo Pesquero: Mar del Plata, Argentina, 2017; Available online: http://hdl.handle.net/1834/17235 (accessed on 10 November 2020).
- Rockhopper Exploration PLC. Sea Lion Field Development Environmental Baseline Survey Report; Rockhopper Exploration PLC: Salisbury, UK, 2012. [Google Scholar]
- Rockhopper Exploration PLC. Sea Lion Post Drill Environmental Survey Environmental Report; Rockhopper Exploration PLC: Salisbury, UK, 2012. [Google Scholar]
- Auscavitch, S.R.; Waller, R.G. Biogeographical Patterns among Deep Sea Megabenthic Communities across the Drake Passage. Antarct. Sci. 2017, 29, 531–543. [Google Scholar] [CrossRef]
- Costello, M.J. Distinguishing Marine Habitat Classification Concepts for Ecological Data Management. Mar. Ecol. Prog. Ser. 2009, 397, 253–268. [Google Scholar] [CrossRef] [Green Version]
- Howell, K.L.; Davies, J.S.; Narayanaswamy, B.E. Identifying Deep-Sea Megafaunal Epibenthic Assemblages for Use in Habitat Mapping and Marine Protected Area Network Design. J. Mar. Biol. Assoc. U. K. 2010, 90, 33–68. [Google Scholar] [CrossRef]
- Danovaro, R.; Fanelli, E.; Aguzzi, J.; Billett, D.; Carugati, L.; Corinaldesi, C.; Anno, A.D.; Gjerde, K.; Jamieson, A.J.; Kark, S.; et al. Ecological Variables for Developing a Global Deep-Ocean Monitoring and Conservation Strategy. Nat. Ecol. Evol. 2020, 4, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Fallon, S.; Thresher, R.; Adkins, J. Age and Growth of the Cold-Water Scleractinian Solenosmilia Variabilis and Its Reef on SW Pacific Seamounts. Coral Reefs 2014, 33, 31–38. [Google Scholar] [CrossRef]
- Risk, M.J.; Heikoop, J.M.; Snow, N.; Beukens, R. Lifespans and Growth Patterns of Two Deep-Sea Corals: Primnoa resedaeformis and Desmophyllum cristagalli. Hydrobiologia 2002, 471, 125–131. [Google Scholar] [CrossRef]
- Adkins, J.; Henderson, G.; Wang, S.-L.; O’Shea, S.; Mokadem, F. Growth Rates of the Deep-Sea Scleractinia Desmophyllum cristagalli and Enallopsammia rostrata. Earth Planet. Sci. Lett. 2004, 227, 481–490. [Google Scholar] [CrossRef]
- Althaus, F.; Williams, A.; Schlacher, T.A.; Kloser, R.J.; Green, M.A.; Barker, B.A.; Bax, N.J.; Brodie, P. Impacts of Bottom Trawling on Deep-Coral Ecosystems of Seamounts Are Long-Lasting. Mar. Ecol. Prog. Ser. 2009, 397, 279–294. [Google Scholar] [CrossRef]
- Parker, S.J.; Bowden, D.A. Identifying Taxonomic Groups Vulnerable to Bottom Longline Fishing Gear in the Ross Sea Region. CCAMLR Sci. 2010, 17, 105–127. [Google Scholar]
- Williams, A.; Schlacher, T.A.; Rowden, A.A.; Althaus, F.; Clark, M.R.; Bowden, D.A.; Stewart, R.; Bax, N.J.; Consalvey, M.; Kloser, R.J. Seamount Megabenthic Assemblages Fail to Recover from Trawling Impacts. Mar. Ecol. 2010, 31, 183–199. [Google Scholar] [CrossRef]
- Leese, F.; Kop, A.; Wägele, J.; Held, C. Cryptic Speciation in a Benthic Isopod from Patagonian and Falkland Island Waters and the Impact of Glaciations on Its Population Structure. Front. Zool. 2008, 15, 19. [Google Scholar] [CrossRef] [Green Version]
- Hedgpeth, J.W. Introduction to Antarctic Zoogeography. Antarct. Map Folio Ser. 1969, 11, 1–9. [Google Scholar]
- De Broyer, C.; Koubbi, P. The Biogeography of the Southern Ocean. In Biogeographic Atlas of the Southern Ocean; Scientific Committee on Antarctic Research: Cambridge, UK, 2014; pp. 2–9. [Google Scholar]
- Briggs, J.; Bowen, B. A Realignment of Marine Biogeographic Provinces with Particular Reference to Fish Distributions. J. Biogeogr. 2012, 39, 12–30. [Google Scholar] [CrossRef]
- Lagger, A.T.C.; Reyna, T.M.P.; Tatián, G.L.M. Ascidian Distribution Provides New Insights to Help Define the Biogeographic Provinces in the South American Region. Polar Biol. 2018, 41, 1123–1131. [Google Scholar] [CrossRef] [Green Version]
- Combes, V.; Matano, R.P. Progress in Oceanography the Patagonian Shelf Circulation: Drivers and Variability. Prog. Oceanogr. 2018, 167, 24–43. [Google Scholar] [CrossRef]
- Bastida, R.; Roux, A.; Martinez, D.E.; Brook, S. Benthic Communities of the Argentine Continental Shelf. Oceanol. Acta 1992, 15, 687–698. [Google Scholar]
- Ewing, M.; Lonardi, A.G. Sediment Transport and Distribution in the Argentine Basin. 5. Sediment Structure of the Argentina Margin, Basin, and Related Provinces. In Physics and Chemistry of the Earth 8; Pergamon Press: New York, NY, USA, 1971; pp. 123–251. [Google Scholar]
- Muñoz, A.; Acosta, J.; Cristobo, J.; Druet, M.; Uchupi, E.; Group, A. Geomorphology and Shallow Structure of a Segment of the Atlantic Patagonian Margin Earth-Science Reviews Geomorphology and Shallow Structure of a Segment of the Atlantic Patagonian Margin. Earth Sci. Rev. 2013, 121, 73–95. [Google Scholar] [CrossRef]
- Brown, C.S.; Newton, A.M.W.; Huuse, M.; Buckley, F. Iceberg Scours, Pits, and Pockmarks in the North Falkland Basin. Mar. Geol. 2017, 386, 140–152. [Google Scholar] [CrossRef]
- Langenkämper, D.; Zurowietz, M.; Schoening, T.; Nattkemper, T.W. BIIGLE 2.0—Browsing and Annotating Large Marine Image Collections. Front. Mar. Sci. 2017, 4, 83. [Google Scholar] [CrossRef] [Green Version]
- Howell, K.; Davies, J.; Allcock, A.L.; Braga-Henriques, A.; Buhl-Mortensen, P.; Carreiro-Silva, M.; Dominguez-Carrió, C.; Durden, J.; Foster, N.; Game, C.; et al. A Framework for the Development of a Global Standardised Marine Taxon Reference Image Database (SMarTaR-ID) to Support Image-Based Analyses. PLoS ONE 2019, 14, e0218904. [Google Scholar] [CrossRef] [Green Version]
- Beazley, L.; Kenchington, E.L.; Murillo, F.J. Deep-Sea Sponge Grounds Enhance Diversity and Abundance of Epibenthic Megafauna in the Northwest Atlantic. ICES J. Mar. Sci. 2013, 70, 1471–1490. [Google Scholar] [CrossRef]
- Turner, J.A.; Hitchin, R.; Verling, E.; Van Rein, H. Epibiota Remote Monitoring from Digital Imagery: Interpretation Guidelines; JNCC Report; Joint Nature Conservation Committee (JNCC)/NMBAQCS: Peterborough, UK, 2006; 42p. [Google Scholar]
- EUNIS. EUNIS Habitat Classifications. Available online: https://www.eea.europa.eu/data-and-maps/data/eunis-habitat-classification-1 (accessed on 12 July 2021).
- Wright, D.J. Survey Data Analysis for Hemptons Turbot Bank: An Investigation into the Categorization of Survey Data Sets for Mapping a Sand Wave Seafloor System; MESH Report; MESH: Berkhamsted, UK, 2005. [Google Scholar]
- Wilson, M.F.J.; O’Connel, B.; Brown, C.; Guinan, J.C.; Grehan, A.J. Multiscale Terrain Analysis of Multibeam Bathymetry Data for Habitat Mapping on the Continental Slope. Mar. Geol. 2007, 30, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Lauvset, S.; Key, R.; Olsen, A.; van Heuven, S.; Velo, A.; Lin, X.; Schirnick, C. A New Global Interior Ocean Mapped Climatology: The 1 × 1 GLODAP Version 2. Earth Syst. Sci. Data 2016, 8, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Roberts, M.J.; Wheeler, A.J.; Freiwald, A.; Cairns, S.D. Cold-Water Corals: The Biology and Geology of Deep-Sea Coral Habitats; Cambridge University Press: Cambridge, UK, 2009; pp. 1–350. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E. Ecologically Meaningful Transformations for Ordination of Species Data. Oecologia 2011, 129, 271–280. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Clarke, R. Non Parametrc Multivariate Analysis of Change. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Bocard, D.; Gillet, F.; Legendre, P. Numerical Ecology with R; Springer: New York, NY, USA, 2011. [Google Scholar]
- Pearman, T.; Robert, K.; Callaway, A.; Hall, R.; Lo Iacono, C.; Huvenne, V.A.I. Improving the Predictive Capability of Benthic Species Distribution Models by Incorporating Oceanographic Data—Towards Holistic Ecological modelling of a Submarine Canyon. Prog. Oceanogr. 2020, 184, 102338. [Google Scholar] [CrossRef]
- Levin, L.A.; Jetter, R.; Rex, M.; Goodday, R.; Smith, C.; Pineda, J.; Stuart, C.; Robert, R.; Hessler, R.; Pawson, D. Enviornemntal Influences on Regional Deep-Sea Species Diversity. Annu. Rev. Ecol. Syst. 2001, 32, 51–93. [Google Scholar] [CrossRef] [Green Version]
- Robert, K.; Jones, D.O.B.B.; Tyler, P.A.; Van Rooij, D.; Huvenne, V.A.I.; Van Tooij, D.; Huvenne, V.A.I. Finding the Hotspots within a Biodiversity Hotspot: Fine-Scale Biological Predictions within a Submarine Canyon Using High-Resolution Acoustic Mapping Techniques. Mar. Ecol. 2015, 36, 1256–1276. [Google Scholar] [CrossRef]
- R_Core_Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- Auster, P.J.; Gjerde, K.; Heupel, E.; Watling, L.; Grehan, A.; Rogers, A.D. Definition and Detection of Vulnerable Marine Ecosystems on the High Seas: Problems with the ‘Move-on’ Rule. ICES J. Mar. Sci. 2011, 68, 254–264. [Google Scholar] [CrossRef] [Green Version]
- Addamo, A.M.; Vertino, A.; Stolarski, J.; García-Jiménez, R.; Taviani, M.; Machordom, A. Merging Scleractinian Genera: The Overwhelming Genetic Similarity between Solitary Desmophyllum and Colonial Lophelia. BMC Evol. Biol. 2016, 16, 108. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.J.; Guinotte, J.M. Global Habitat Suitability for Framework-Forming Cold-Water Corals. PLoS ONE 2011, 6, e18483. [Google Scholar] [CrossRef]
- Rowden, A.; Pearman, T.; Bowden, D.; Anderson, O.; Clark, M. Determining Coral Density Thresholds for Identifying Structurally Complex Vulnerable Marine Ecosystems in the Deep Sea. Front. Mar. Sci. 2020, 7, 95. [Google Scholar] [CrossRef] [Green Version]
- Buhl-mortensen, L.; Vanreusel, A.; Gooday, A.J.; Levin, L.A.; Priede, I.G.; Gheerardyn, H.; King, N.J.; Raes, M. Biological Structures as a Source of Habitat Heterogeneity and Biodiversity on the Deep Ocean Margins. Mar. Ecol. 2010, 31, 21–50. [Google Scholar] [CrossRef]
- Vad, J.; Orejas, C.; Moreno-navas, J.; Findlay, H.S.; Roberts, J.M. Assessing the Living and Dead Proportions of Cold-Water Coral Colonies: Implications for Deep-Water Marine Protected Area Monitoring in a Changing Ocean. PeerJ 2017, 5, e3705. [Google Scholar] [CrossRef] [PubMed]
- Auster, P.J. Are Deep-Water Corals Important Habitats for Fishes. In Cold-Water Corals and Ecosystems; Erlangen Earth Conference Series; Springer: Berlin/Heidelberg, Germany, 2014; pp. 747–760. [Google Scholar] [CrossRef]
- Miller, R.J.; Hocevar, J.; Stone, R.P.; Fedorov, D.V. Structure-Forming Corals and Sponges and Their Use as Fish Habitat in Bering Sea Submarine Canyons. PLoS ONE 2012, 7, e33885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kutti, T.; Bergstad, O.A.; Fosså, J.H.; Helle, K. Cold-Water Coral Mounds and Sponge-Beds as Habitats for Demersal Fish on the Norwegian Shelf. Deep. Res. Part II Top. Stud. Oceanogr. 2014, 99, 122–133. [Google Scholar] [CrossRef]
- D’Onghia, G.; Calculli, C.; Capezzuto, F.; Carlucci, R.; Carluccio, A.; Maiorano, P.; Pollice, A.; Ricci, P.; Sion, L.; Tursi, A. New Records of Cold-Water Coral Sites and Fish Fauna Characterization of a Potential Network Existing in the Mediterranean Sea. Mar. Ecol. 2016, 37, 1398–1422. [Google Scholar] [CrossRef]
- D’Onghia, G. Cold-Water Corals as Shelter, Feeding and Life-History Critical Habitats for Fish Species: Ecological Interactions and Fishing Impact. In Mediterranean Cold-Water Corals: Past, Present and Future; Springer: Cham, Switzerland, 2019; pp. 335–356. [Google Scholar]
- Robert, K.; Huvenne, V.A.I.; Georgiopoulou, A.; Jones, D.O.B.; Marsh, L.; Carter, D.O.G.; Chaumillon, L. New Approaches to High-Resolution Mapping of Marine Vertical Structures. Sci. Rep. 2017, 7, 9005. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, P.; Hovland, M.; Brattegard, T.; Farestveit, R. Deep Water Bioherms of the Scleractinian Coral Lophelia pertusa (L.) at 641N on Thet Heegian Shelf: Structure and Associated Megafauna. Sarsia 1995, 80, 145–158. [Google Scholar] [CrossRef]
- Forraz Corrêa, P.; Jovane, L.; Murton, B.J.; Sumida, P.Y. Benthic Megafauna Habitats, Community Structure and Environmental Drivers at Rio Grande Rise (SW Atlantic). Deep. Res. Part I Oceanogr. Res. Pap. 2022, 186, 103811. [Google Scholar] [CrossRef]
- Ward, T.; Sorokin, S.J.; Currie, D.; Rogers, P.; Mcleay, L. Epifaunal Assemblages of the Eastern Great Australian Bight: Effectiveness of a Benthic Protection Zone in Representing Regional Biodiversity. Cont. Shelf Res. 2006, 26, 25–40. [Google Scholar] [CrossRef]
- Burgos, J.M.; Buhl-mortensen, L.; Buhl-mortensen, P.; Ólafsdóttir, S.H.; Tuck, I.D. Predicting the Distribution of Indicator Taxa of Vulnerable Marine Ecosystems in the Arctic and Sub-Arctic Waters of the Nordic Seas. Front. Mar. Sci. 2020, 7, 131. [Google Scholar] [CrossRef] [Green Version]
- Hanz, U.; Riekenberg, P.; De Kluijver, A.; Van Der Meer, M.; Middelburg, J.J.; De Goeij, J.M.; Bart, M.C.; Wurz, E.; Colaço, A.; Duineveld, G.C.A.; et al. The Important Role of Sponges in Carbon and Nitrogen Cycling in a Deep-Sea Biological Hotspot. Funct. Ecol. 2022. [Google Scholar] [CrossRef]
- Bart, M.C.; Hudspith, M.; Rapp, H.T.; Verdonschot, P.F.M.; De Goeij, J.M.; Xavier, J.R. A Deep-Sea Sponge Loop? Sponges Transfer Dissolved and Particulate Organic Carbon and Nitrogen to Associated Fauna. Front. Mar. Sci. 2021, 8, 604879. [Google Scholar] [CrossRef]
- Beaulieu, S.E. Life on Glass Houses: Sponge Stalk Communities in the Deep Sea. Mar. Biol. 2001, 138, 803–817. [Google Scholar] [CrossRef]
- Tissot, B.N.; Yoklavich, M.M.; Love, M.S.; York, K.; Amend, M. Benthic Invertebrates That Form Habitat on Deep Banks off Southern California, with Species Reference to Deep Sea Coral. Fish. Bull. 2006, 104, 167–181. [Google Scholar]
- Hawkes, N.; Korabik, M.; Beazley, L.; Rapp, H.T.; Xavier, J.R.; Kenchington, E.; Pen, O.; Hawkes, N.; Korabik, M.; Beazley, L.; et al. Glass Sponge Grounds on the Scotian Shelf and Their Associated Biodiversity. Mar. Ecol. Prog. Ser. 2019, 614, 91–109. [Google Scholar] [CrossRef] [Green Version]
- Pirtle, J. Habitat-Based Assessment of Structure-Forming Megafaubnal Invertebrates and Fishes on Cordell Bank, California. Master’s Thesis, Washington State University, Pullman, WA, USA, 2005. [Google Scholar]
- Bo, M.; Bertolino, M.; Canese, S.; Giusti, M.; Angiolillo, M.; Pansini, M.; Taviani, M. Role of Deep Sponge Grounds in the Mediterranean Sea: A Case Study in Southern Italy. Hydrobiologia 2011, 687, 163–177. [Google Scholar] [CrossRef] [Green Version]
- Maldonado, M.; Aguilar, R.; Bannister, R.J.; James, J.; Conway, K.W.; Dayton, P.K.; Cristina, D.; Gutt, J.; Kelly, M.; Kenchington, E.L.R.; et al. Sponge Grounds as Key Marine Habitats: A Synthetic Review of Types, Structure , Functional Roles , and Conservation Concerns. In Marine Animal Forests; Rossi, S., Bramanti, L., Gori, A., Orejas Saco del Valle, C., Eds.; Springer: Cham, Switzerland, 2016; ISBN 9783319170015. [Google Scholar] [CrossRef]
- Leys, S. Hexactinellid Sponge Ecology: Growth Rates and Seasonality in Deep Water Sponges. J. Exp. Mar. Biol. Ecol. 1998, 230, 111–129. [Google Scholar] [CrossRef]
- Peter, K.; Wang, X.; Vennemann, T.W.; Sinha, B.; Müller, W.E.G. Siliceous Deep-Sea Sponge Monorhaphis Chuni : A Potential Paleoclimate Archive in Ancient Animals. Chem. Geol. 2012, 301, 143–151. [Google Scholar] [CrossRef]
- Klitgaard, A.B. The Fauna Associated with Outer Shelf and Upper Slope Sponges (Porifera, Demospongiae) at the Faroe Islands, Northeastern Atlantic. Sarsia 1995, 80, 1–22. [Google Scholar] [CrossRef] [Green Version]
- OSPAR. Background Document for Deep-Sea Sponge Aggregations; OSPAR Commission: London UK, 2010; ISBN 978-1-907390-26-5. [Google Scholar]
- Henry, L.A.; Roberts, J.M. Applying the OSPAR Habitat Definition of Deep-Sea Sponge Aggregations to Verify Suspected Records of the Habitat in UK Waters. JNCC Rep. 2014, 5, 508. [Google Scholar]
- Parry, M.; Howell, K.L.; Narayanaswamy, B.; Bett, B.J.; Jones, D.O.B.; Hughes, J.; Piechaud, N.; Nickell, T.; Ellwood, H.; Askew, N.; et al. A Deep-Sea Section for the Marine Habitat Classification of Britain and Ireland (v 15.03); JNCC Rep 530; JNCC: Peterborough, UK, 2015; ISSN 0963-8091. [Google Scholar]
- Baillon, S. Diversity, Distribution and Nature of Faunal Associations with Deep-Sea Pennatulacean Corals in the Northwest Atlantic. PLoS ONE 2014, 9, 14–16. [Google Scholar] [CrossRef] [Green Version]
- Baillon, S.; Sciences, O. Characterization and Role of Major Deep-Sea Pennatulacean Corals in the Bathyal Zone of Eastern Canada. Ph.D. Thesis, Memorial University of Newfoundland, St. John’s, NL, Canada, 2014. [Google Scholar]
- Williams, G.C. Living Genera of Sea Pens (Coelenterata: Octocorallia: Pennatulacea): Illustrated Key and Synopses. Zool. J. Linn. Soc. 1995, 113, 93–140. [Google Scholar] [CrossRef]
- Baillon, S.; Hamel, J.; Wareham, V.E.; Mercier, A. Deep Cold-Water Corals as Nurseries for Fish Larvae. Front. Ecol. Environ. 2012, 10, 351–356. [Google Scholar] [CrossRef] [Green Version]
- Levin, L.; Sibeut, M.; Smith, A.; Vanreaisel, A. The Roles of Habitat Heterogeneity in Generating and Maintaining Biodiversity on Continental Margins: An Introduction. Mar. Ecol. 2010, 31, 1–5. [Google Scholar] [CrossRef]
- Rex, M.; Etter, J. Deep-Sea Biodiversity: Pattern and Scale; Harvard University Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Puerta, P.; Johnson, C.; Carreiro-silva, M.; Henry, L.; Kenchington, E.; Morato, T.; Kazanidis, G.; Rueda, J.L.; Urra, J.; Ross, S.; et al. Influence of Water Masses on the Biodiversity and Biogeography of Deep-Sea Benthic Ecosystems in the North Atlantic. Front. Mar. Sci. 2020, 7, 239. [Google Scholar] [CrossRef] [Green Version]
- Buhl-Mortensen, L.; Dolan, M.F.J.; Holte, B.; Dannheim, J.; Buhl-mortensen, P.; Bellec, V. Habitat Complexity and Bottom Fauna Composition at Different Scales on the Continental Shelf and Slope of Northern Norway. Hydrobiologia 2012, 685, 191–219. [Google Scholar] [CrossRef] [Green Version]
- Baker, K.D.; Wareham, V.; Snelgrove, P.V.R.; Haedrich, R.; Fifield, D.; Edinger, E.; Gilkinson, K. Distributional Patterns of Deep-Sea Coral Assemblages in Three Submarine Canyons off Newfoundland, Canada. Mar. Ecol. Prog. Ser. 2012, 445, 235–249. [Google Scholar] [CrossRef] [Green Version]
- Lacharité, M.; Metaxas, A. Hard Substrate in the Deep Ocean: How Sediment Features Influence Epibenthic Megafauna on the Eastern Canadian Margin. Deep Sea Res. Part I Oceanogr. Res. Pap. 2017, 126, 50–61. [Google Scholar] [CrossRef]
- Isola, J.I.; Bravo, M.E.; Bozzano, G.; Palma, F.I.; Ormazabal, J.P.; Principi, S. The Late-Quaternary Deposits of the Piedra Buena Terrace (Patagonian Continental Slope, SW Atlantic): An Example of Interaction between Bottom Currents and Seafloor Morphology. Mar. Geol. 2021, 435, 106459. [Google Scholar] [CrossRef]
- Hernández-molina, F.J.; Paterlini, M.; Violante, R.; Marshall, P.; De Isasi, M.; Somoza, L. Contourite Depositional System on the Argentine Slope: An Exceptional Record Contourite Depositional System on the Argentine Slope: An Exceptional Record of the Influence of Antarctic Water Masses. Geology 2009, 37, 507–510. [Google Scholar] [CrossRef]
- Hebbeln, D.; Van Rooij, D.; Wienberg, C. Good Neighbours Shaped by Vigorous Currents: Cold-Water Coral Mounds and Contourites in the North Atlantic. Mar. Geol. 2016, 378, 171–185. [Google Scholar] [CrossRef]
- De Mol, L.; Van Rooij, D.; Pirlet, H.; Greinert, J.; Frank, N.; Quemmerais, F.; Henriet, J. Cold-Water Coral Habitats in the Penmarc’h and Guilvinec Canyons (Bay of Biscay): Deep-Water versus Shallow-Water Settings. Mar. Geol. 2011, 282, 40–52. [Google Scholar] [CrossRef] [Green Version]
- Howell, K.L.; Holt, R.; Pulido, I.; Stewart, H.; Endrino, I.P.; Stewart, H. When the Species Is Also a Habitat: Comparing the Predictively Modelled Distributions of Lophelia Pertusa and the Reef Habitat It Forms. Biol. Conserv. 2011, 144, 2656–2665. [Google Scholar] [CrossRef] [Green Version]
- Huvenne, V.; Tyler, P.; Fisher, D.; Hauton, C.; Huhnerbach, V.; Le Bas, T.; Wolff, G. A Picture on the Wall: Innovative Mapping Reveals Cold-Water Coral Refuge in Submarine Canyon. PLoS ONE 2011, 6, e28755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gori, A.; Orejas, C.; Madurell, T.; Bramanti, L.; Martins, M.; Quintanilla, E.; Marti-Puig, P.; Lo Iacono, C.; Puig, P.; Requena, S.; et al. Bathymetrical Distribution and Size Structure of Cold-Water Coral Populations in the Cap de Creus and Lacaze-Duthiers Canyons (Northwestern Mediterranean). Biogeosciences 2013, 10, 2049–2060. [Google Scholar] [CrossRef] [Green Version]
- Rengstorf, A.; Yesson, C.; Brown, C.; Grehan, A.J.; Crame, A. High-Resolution Habitat Suitability Modelling Can Improve Conservation of Vulnerable Marine Ecosystems in the Deep Sea. J. Biogeogr. 2013, 40, 1702–1714. [Google Scholar] [CrossRef]
- Fabri, M.; Bargain, A.; Pairaud, I.; Pedel, L.; Taupier-Letage, I. Cold-Water Coral Ecosystems in Cassidaigne Canyon: An Assessment of Their Environmental Living Conditions. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 137, 436–453. [Google Scholar] [CrossRef] [Green Version]
- Mohn, C.; Rengstorf, A.; White, M.; Duineveld, G.; Mienis, F.; Soetaert, K.; Grehan, A. Linking Benthic Hydrodynamics and Cold-Water Coral Occurrences: A High-Resolution Model Study at Three Cold-Water Coral Provinces in the NE Atlantic. Prog. Oceanogr. 2014, 122, 92–101. [Google Scholar] [CrossRef]
- Post, A.; Lavoie, C.; Domack, E.; Leventer, A.; Shevenell, A.; Fraser, A. Environmental Drivers of Benthic Communities and Habitat Heterogeneity on an East Antarctic Shelf. Antarct. Sci. 2016, 29, 17–32. [Google Scholar] [CrossRef]
- Pile, A.J.; Young, C.M. The Natural Diet of a Hexactinellid Sponge: Benthic—Pelagic Coupling in a Deep-Sea Microbial Food Web. Deep Sea Res. Part I Oceanogr. Res. Pap. 2006, 53, 1148–1156. [Google Scholar] [CrossRef]
- Gori, A.; Rossi, S.; Linares, C. Reproductive Cycle and Trophic Ecology in Deep versus Shallow Populations of the Mediterranean Gorgonian Eunicella singularis (Cap de Creus, Size and Spatial Structure in Deep versus Shallow Populations of the Mediterranean Gorgonian Eunicella singulari). Coral Reefs 2012, 31, 823–837. [Google Scholar] [CrossRef]
- Maier, S.R.; Kutti, T.; Bannister, R.J.; Fang, J.K.; Van, P.; Van Rijswijk, P.; Oevelen, D. Van Recycling Pathways in Cold- Water Coral Reefs: Use of Dissolved Organic Matter and Bacteria by Key Suspension Feeding Taxa. Sci. Rep. 2020, 10, 9942. [Google Scholar] [CrossRef]
- Cunha, M.; Paterson, G.; Amaro, T.; Blackbird, S.; De Stiger, H.; Ferreira, C.; Glover, A.; Hilario, A.; Kiriakoulakis, K.; Neal, L.; et al. Biodiversity of Macrofaunal Assemblages from Three Portuguese Submarine Canyons (NE Atlantic). Deep Sea Res. Part II Top. Stud. Oceanogr. 2011, 58, 2433–2447. [Google Scholar] [CrossRef] [Green Version]
- Glorioso, P.D.; Piola, A.R.; Leben, R.R. Mesoscale Eddies in the Subantarctic Front—Southwest Atlantic. Sci. Mar. 2005, 69, 7–15. [Google Scholar] [CrossRef] [Green Version]
- Tomczak, M. A Multiparameter Extension of Temperature/Salinity Diagram Techniques for the Analysis of Non-Isopycnal Mixing. Prog. Oceanogr. 1981, 10, 147–171. [Google Scholar] [CrossRef]
- Roberts, J.M.; Davies, A.J.; Henry, L.A.; Dodds, L.A.; Duineveld, G.C.A.; Lavaleye, M.S.S.; Maier, C.; Van Soest, R.W.M.; Bergman, M.J.N.; Hühnerbach, V.; et al. Mingulay Reef Complex: An Interdisciplinary Study of Cold-Water Coral Habitat, Hydrography and Biodiversity. Mar. Ecol. Prog. Ser. 2009, 397, 139–151. [Google Scholar] [CrossRef] [Green Version]
- Davies, A.J.; Duienveld, G.; Lavaleye, M.S.S.; Bergman, M.J.N.; Van Haren, H.; Roberts, J.M. Downwelling and Deep-Water Currents as Food Supply Mechanisms to the Cold-Water Coral Lophelia pertusa (Scleractinia) at the Mingulay Reef Complex. Limnol. Oceanogr. 2009, 54, 620–629. [Google Scholar] [CrossRef] [Green Version]
- Dullo, W.; Juva, K.; Flögel, S.; Karstensen, J.; Linke, P.; Dullo, W. Tidal Dynamics Control on Cold-Water Coral Growth: A High-Resolution Multivariable Study on Eastern Atlantic Cold-Water Coral Sites. Front. Mar. Sci. 2020, 7, 132. [Google Scholar] [CrossRef]
- Bett, B. UK Atlantic Margin Environmental Survey: Introduction and Overview of Bathyal Benthic Ecology. Cont. Shelf Res. 2001, 21, 917–956. [Google Scholar] [CrossRef]
- Howell, K.; Billett, D.; Tyler, P. Depth-Related Distribution and Abundance of Seastars (Echinodermata:Asteroidea) in the Porcupine Seabight and Porcupine Abyssal Plain, N.E. Atlantic. Deep. Res. Part II Top. Stud. Oceanogr. 2002, 49, 1901–1902. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Description | Unit | Native Resolution |
|---|---|---|---|
| Seabed terrain | |||
| Depth † | Bathymetry extracted from GEBCO. https://www.gebco.net/ (accessed on 10 November 2020) | m | 0.004° |
| Terrain derivatives | |||
| Slope † | A first derivative of bathymetry measuring the change in elevation from one pixel to its neighbour derived from a neighbourhood size of 3 × 3 | ° | 0.004° |
| Eastness † | A first derivative of bathymetry measuring the easterly orientation of maximum change along the slope on a continuous scale (−1 to +1) | - | 0.004° |
| Northness † | A first derivative of bathymetry measuring the northerly orientation of maximum change along the slope on a continuous scale (−1 to +1) | - | 0.004° |
| Curvature | A second derivative of bathymetry measuring the shape of the slope, with values indicating whether a slope is convex or concave | - | 0.004° |
| Fine bathymetric position index (FBPI) † | A derived metric of a cell’s position and elevation relative to its surrounding landscape/cells within a user defined area [94] | - | |
| Broad bathymetric position index (BBPI) | A derived metric of a cell’s position and elevation relative to its surrounding landscape/cells within a user defined area [94] | - | |
| Rugosity | A measure of the ratio of the surface area to the planar area calculated with a neighbourhood size of 3 × 3 pixels [95] | - | |
| Oceanography variables | |||
| Surface chlorophyll | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | mg m−3 | 0.08° |
| Seabed temperature | Extracted from GLOBAL_ANALYSIS_FORECAST_PHY_001_024 http://marine.copernicus.eu (accessed on 10 November 2020) | °C | 0.08° |
| Mean seabed current velocity U | Extracted from GLOBAL_ANALYSIS_FORECAST_PHY_001_024 http://marine.copernicus.eu (accessed on 10 November 2020) | m/s | 0.08° |
| Seabed Ph | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | - | 0.08° |
| Surface Ph | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | - | 0.08° |
| Seabed phosphate | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | μmol kg−1 | 0.08° |
| Surface phosphate | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | μmol kg−1 | 0.08° |
| Surface dissolved oxygen | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | μmol kg−1 | 0.08° |
| Seabed silicate | Extracted from GLOBAL_REANALYSIS_BIO_001_029 http://marine.copernicus.eu (accessed on 10 November 2020) | μmol kg−1 | 0.08° |
| Aragonite saturation state | Extracted from GLODAPv.2.2016b [96] (accessed on 22 March 2018) | μmol kg−1 | 1° |
| Dissolved inorganic carbon | Extracted from GLODAPv.2.2016b [96] (accessed on 22 March 2018) | μmol kg−1 | 1° |
| Calcite saturation state | Extracted from GLODAPv.2.2016b [96] (accessed on 22 March 2018) | μmol kg−1 | 1° |
| Nitrate | Extracted from GLODAPv.2.2016b [96] (accessed on 22 March 2018) | μmol kg−1 | 1° |
| Total alkalinity | Extracted from GLODAPv.2.2016b [96] (accessed on 22 March 2018) | μmol kg−1 | 1° |
| Substrata variables | |||
| Substrata † | Substrate type annotated from imagery based upon EUNIS 2022 classifications [93] |
| Cluster | Characterising Morphospecies | Water Depth (m) | Substratum | N |
|---|---|---|---|---|
| 1 | Stylasteridae sp., Massive Ball Porifera | 1070–1840 | Coarse, biogenic gravel, reef, mixed | 197 |
| 2 | Scleractinia sp. 5, Anthoptilum grandiflorum | 1070–1880 | Coarse, mud, sandy mud Sand, muddy sand | 78 |
| 3 | Flabellum sp., Pennatulacea sp. | 1120–1327 | Mud, sandy mud | 9 |
| 4 | Alcyonacea sp. 19, Hormathiidae sp. 2 | 1840 | Hard | 2 |
| 5 | Cold-water whip coral | 1390–1540 | Mud, sandy mud, coarse | 2 |
| Environmental Variables—Significance of Individual Terms by ANOVA | Adjusted R2 | Significance of RDA Plot by ANOVA | |
|---|---|---|---|
| F-Value | p-Value | ||
| Depth ***, Slope ***, FBPI ***, Substrate ***, Eastness ***, Northness ***, Current Velocity *** | 13 | 4.51, df = 12,275 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pearman, T.R.R.; Brewin, P.E.; Baylis, A.M.M.; Brickle, P. Deep-Sea Epibenthic Megafaunal Assemblages of the Falkland Islands, Southwest Atlantic. Diversity 2022, 14, 637. https://doi.org/10.3390/d14080637
Pearman TRR, Brewin PE, Baylis AMM, Brickle P. Deep-Sea Epibenthic Megafaunal Assemblages of the Falkland Islands, Southwest Atlantic. Diversity. 2022; 14(8):637. https://doi.org/10.3390/d14080637
Chicago/Turabian StylePearman, T. R. R., Paul E. Brewin, Alastair M. M. Baylis, and Paul Brickle. 2022. "Deep-Sea Epibenthic Megafaunal Assemblages of the Falkland Islands, Southwest Atlantic" Diversity 14, no. 8: 637. https://doi.org/10.3390/d14080637
APA StylePearman, T. R. R., Brewin, P. E., Baylis, A. M. M., & Brickle, P. (2022). Deep-Sea Epibenthic Megafaunal Assemblages of the Falkland Islands, Southwest Atlantic. Diversity, 14(8), 637. https://doi.org/10.3390/d14080637