
The Biodiversity of Calcaxonian Octocorals from the Irish Continental Slope Inferred from Multilocus Mitochondrial Barcoding

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetics
2.1.1. DNA Extraction and Polymerase Chain Reaction (PCR)
2.1.2. DNA Sequence Quality Control, Haplotype Discovery, and Phylogenetic Inference

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequences 5′–3′ | Gene Boundaries Crossed | Thermocycle Profile | Source |
|---|---|---|---|---|
| CO3Bam5657f | GCT GCT AGT TGG TAT TGG CAT | CO3—mtMutS | a 94 °C, 30 s: 55 °C, 30 s: 72 °C, 45 s. 35x or | [44,45] |
| MUT3458R | TSG AGC AAA AGC CAC TCC | a 94 °C, 30 s: 55 °C, 60 s: 72 °C, 120 s. 35x | ||
| ND4L2475F | TAG GYT TAT TTA CTC ATA CWA T | ND4L—mtMutS | b 94 °C, 20 s: 50 °C, 30 s: 72 °C, 50 s. 35x | [44,46] |
| MUT3458R | TSG AGC AAA AGC CAC TCC | |||
| COII8086f | CAT AAC AGG ACT AGC AGC ATC | COII—igr1—COI | b 94 °C, 30 s: 47 °C, 30 s: 72 °C, 30 s. 40x | [34,47] |
| COIOCTr | ATC ATA GCA TAG ACC ATA CC | |||
| 16s647F | ACA CAG CTC GGT TTC TAT CTA CCA | 16S rRNA-ND2 | b 94 °C, 30 s: 47 °C, 30 s: 72 °C, 60 s. 30x | [47,48] |
| ND2_1417R | CAT CGG GGG CCC ACA TAT G | |||
| CytbBam1279f | AGG AGC CAA TCC AGT AGA GGA ACC | COB–ND6 | b 94 °C, 30 s: 55 °C, 30 s: 72 °C, 45 s. 35x | [36] |
| ND6Bam1648r | TAY AGG TAA GAA ATG CGA GTG ATC |
2.1.3. Genetic Species Delimitation
2.2. Morphological Investigation
2.2.1. Light Microscopy Imaging and In Situ Sclerite Visualisation
2.2.2. Scanning Electron Microscopy
2.3. Taxon Identification and Nomenclature Used throughout This Study
3. Results
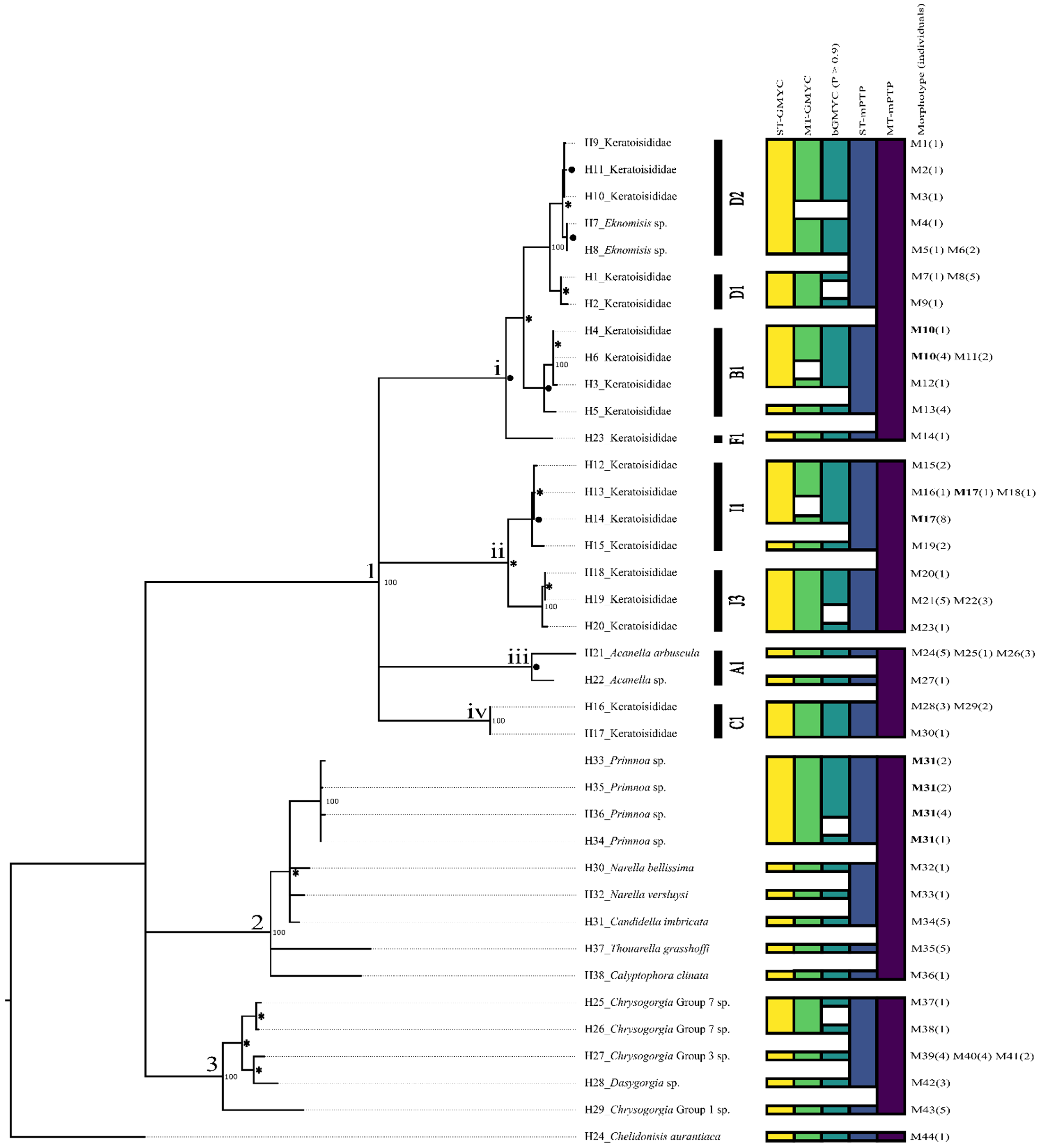
3.1. Clade 1—Keratoisididae
3.2. Clade 2—Primnoidae
3.3. Clade 3—Chrysogorgiidae
3.4. Chelidonisididae
3.5. Uncertainty in Species Estimates
4. Discussion
4.1. Morphological and Genetic Diversity of Calcaxonians
4.2. The Usefulness of Genetic Species Delimitation in Octocorals
4.3. Comparisons of Keratoisidid Diversity with Other Regions
4.4. Interesting New Occurence Records
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ramirez-Llodra, E.; Tyler, P.A.; Baker, M.C.; Bergstad, O.A.; Clark, M.R.; Escobar, E.; Levin, L.A.; Menot, L.; Rowden, A.A.; Smith, C.R.; et al. Man and the last great wilderness: Human impact on the deep sea. PLoS ONE 2011, 6, e22588. [Google Scholar] [CrossRef] [Green Version]
- Fernandez-Arcaya, U.; Ramirez-Llodra, E.; Aguzzi, J.; Allcock, A.L.; Davies, J.S.; Dissanayake, A.; Harris, P.; Howell, K.; Huvenne, V.A.I.; Macmillan-Lawler, M.; et al. Ecological Role of Submarine Canyons and Need for Canyon Conservation: A Review. Front. Mar. Sci. 2017, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Morato, T.; Hoyle, S.D.; Allain, V.; Nicol, S.J. Seamounts are hotspots of pelagic biodiversity in the open ocean. Proc. Natl. Acad. Sci. USA 2010, 107, 9707–9711. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Wheeler, A.J.; Freiwald, A. Reefs of the deep: The biology and geology of cold-water coral ecosystems. Science 2006, 312, 543–547. [Google Scholar] [CrossRef] [Green Version]
- Levin, L.A.; Sibuet, M. Understanding continental margin biodiversity: A new imperative. Ann. Rev. Mar. Sci. 2012, 4, 79–112. [Google Scholar] [CrossRef]
- White, M.; Dorschel, B. The importance of the permanent thermocline to the cold water coral carbonate mound distribution in the NE Atlantic. Earth Planet. Sci. Lett. 2010, 296, 395–402. [Google Scholar] [CrossRef]
- Johnson, M.P.; White, M.; Wilson, A.; Würzberg, L.; Schwabe, E.; Folch, H.; Allcock, A.L. A vertical wall dominated by Acesta excavata and Neopycnodonte zibrowii, part of an undersampled group of deep-sea habitats. PLoS ONE 2013, 8, e79917. [Google Scholar] [CrossRef]
- Pearman, T.R.R.; Robert, K.; Callaway, A.; Hall, R.; Lo Iacono, C.; Huvenne, V.A.I. Improving the predictive capability of benthic species distribution models by incorporating oceanographic data—Towards holistic ecological modelling of a submarine canyon. Prog. Oceanogr. 2020, 184, 102338. [Google Scholar] [CrossRef]
- Watling, L.; France, S.C.; Pante, E.; Simpson, A. Biology of Deep-Water Octocorals. In Advances in Marine Biology; Lesser, M.B.T.-A., In, M.B., Eds.; Academic Press: Cambridge, MA, USA, 2011; Volume 60, pp. 41–122. ISBN 0065-2881. [Google Scholar]
- Etnoyer, P.; Warrenchuk, J. A catshark nursery in a deep gorgonian field in the Mississippi Canyon, Gulf of Mexico. Bull. Mar. Sci. 2007, 81, 553–559. [Google Scholar]
- Shea, E.K.; Ziegler, A.; Faber, C.; Shank, T.M. Dumbo octopod hatchling provides insight into early cirrate life cycle. Curr. Biol. 2018, 28, R144–R145. [Google Scholar] [CrossRef] [Green Version]
- de Neves, B.M.; Wareham Hayes, V.; Herder, E.; Hedges, K.; Grant, C.; Archambault, P. Cold-Water Soft Corals (Cnidaria: Nephtheidae) as Habitat for Juvenile Basket Stars (Echinodermata: Gorgonocephalidae). Front. Mar. Sci. 2020, 7, 547896. [Google Scholar] [CrossRef]
- Prouty, N.G.; Fisher, C.R.; Demopoulos, A.W.J.; Druffel, E.R.M. Growth rates and ages of deep-sea corals impacted by the Deepwater Horizon oil spill. Deep. Res. Part II Top. Stud. Oceanogr. 2016, 129, 196–212. [Google Scholar] [CrossRef]
- France, S.C. Genetic analysis of bamboo corals (Cnidaria: Octocorallia: Isididae): Does lack of colony branching distinguish Lepidisis from Keratoisis? Bull. Mar. Sci. 2007, 81, 323–333. [Google Scholar]
- Dueñas, L.F.; Alderslade, P.; Sánchez, J.A. Molecular systematics of the deep-sea bamboo corals (Octocorallia: Isididae: Keratoisidinae) from New Zealand with descriptions of two new species of Keratoisis. Mol. Phylogenet. Evol. 2014, 74, 15–28. [Google Scholar] [CrossRef]
- Watling, L.; Heestand Saucier, E.; France, S.C. Towards a revision of the bamboo corals (Octocorallia): Part 4, delineating the family Keratoisididae. Zootaxa 2022, 5093, 337–375. [Google Scholar] [CrossRef]
- Cordeiro, R.; McFadden, C.S.; Van Ofwegen, L.; Williams, G. World List of Octocorallia. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=125294 (accessed on 25 May 2022).
- Untiedt, C.B.; Quattrini, A.M.; McFadden, C.S.; Alderslade, P.A.; Pante, E.; Burridge, C.P. Phylogenetic Relationships Within Chrysogorgia (Alcyonacea: Octocorallia), a Morphologically Diverse Genus of Octocoral, Revealed Using a Target Enrichment Approach. Front. Mar. Sci. 2021, 7, 599984. [Google Scholar] [CrossRef]
- Taylor, M.L.; Rogers, A.D. Primnoidae (Cnidaria: Octocorallia) of the SW Indian Ocean: New species, genus revisions and systematics. Zool. J. Linn. Soc. 2017, 181, 70–97. [Google Scholar] [CrossRef] [Green Version]
- Cairns, S.D.; Wirshing, H.H. A phylogenetic analysis of the Primnoidae (Anthozoa: Octocorallia: Calcaxonia) with analyses of character evolution and a key to the genera and subgenera. BMC Evol. Biol. 2018, 18, 66. [Google Scholar] [CrossRef] [Green Version]
- Núñez-Flores, M.; Gomez-Uchida, D.; López-González, P.J. Molecular systematics of Thouarella (Octocorallia: Primnoidae) with the description of three new species from the Southern Ocean based on combined molecular and morphological evidence. Invertebr. Syst. 2021, 35, 655–674. [Google Scholar] [CrossRef]
- Davies, J.S.; Stewart, H.A.; Narayanaswamy, B.E.; Jacobs, C.; Spicer, J.; Golding, N.; Howell, K.L. Benthic assemblages of the Anton Dohrn Seamount (NE Atlantic): Defining deep-sea biotopes to support habitat mapping and management efforts with a focus on vulnerable marine ecosystems. PLoS ONE 2015, 10, e124815. [Google Scholar] [CrossRef] [Green Version]
- Robert, K.; Jones, D.O.B.; Tyler, P.A.; Van Rooij, D.; Huvenne, V.A.I. Finding the hotspots within a biodiversity hotspot: Fine-scale biological predictions within a submarine canyon using high-resolution acoustic mapping techniques. Mar. Ecol. 2015, 36, 1256–1276. [Google Scholar] [CrossRef]
- Pante, E.; Abdelkrim, J.; Viricel, A.; Gey, D.; France, S.C.; Boisselier, M.C.; Samadi, S. Use of RAD sequencing for delimiting species. Heredity 2015, 114, 450–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrera, S.; Shank, T.M. RAD sequencing enables unprecedented phylogenetic resolution and objective species delimitation in recalcitrant divergent taxa. Mol. Phylogenet. Evol. 2016, 100, 70–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson, K.L.; Pentico, A.; Quattrini, A.M.; McFadden, C.S. New approaches to species delimitation and population structure of anthozoans: Two case studies of octocorals using ultraconserved elements and exons. Mol. Ecol. Resour. 2021, 21, 78–92. [Google Scholar] [CrossRef] [PubMed]
- Quattrini, A.M.; Wu, T.; Soong, K.; Jeng, M.S.; Benayahu, Y.; McFadden, C.S. A next generation approach to species delimitation reveals the role of hybridization in a cryptic species complex of corals. BMC Evol. Biol. 2019, 19, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, M.J.; Wiklund, H.; Neal, L.; Jeffreys, R.; Linse, K.; Ruhl, H.; Glover, A.G. DNA barcoding uncovers cryptic diversity in 50% of deep-sea antarctic polychaetes. R. Soc. Open Sci. 2016, 3, 160432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puckridge, M.; Andreakis, N.; Appleyard, S.A.; Ward, R.D. Cryptic diversity in flathead fishes (Scorpaeniformes: Platycephalidae) across the Indo-West Pacific uncovered by DNA barcoding. Mol. Ecol. Resour. 2013, 13, 32–42. [Google Scholar] [CrossRef] [PubMed]
- Muthye, V.; Mackereth, C.D.; Stewart, J.B.; Lavrov, D.V. Large dataset of octocoral mitochondrial genomes provides new insights into mt-mutS evolution and function. DNA Repair 2022, 110, 103273. [Google Scholar] [CrossRef]
- Bilewitch, J.P.; Degnan, S.M. A unique horizontal gene transfer event has provided the octocoral mitochondrial genome with an active mismatch repair gene that has potential for an unusual self-contained function. BMC Evol. Biol. 2011, 11, 228. [Google Scholar] [CrossRef] [Green Version]
- France, S.C.; Hoover, L.L. DNA sequences of the mitochondrial COI gene have low levels of divergence among deep-sea octocorals (Cnidaria: Anthozoa). Hydrobiologia 2002, 471, 149–155. [Google Scholar] [CrossRef]
- France, S.C.; Hoover, L.L. Analysis of variation in mitochondrial DNA sequences (ND3, ND4L, MSH) among Octocorallia (=Alcyonaria) (Cnidaria: Anthozoa). Bull. Biol. Soc. Washingt. 2001, 10, 110–118. [Google Scholar]
- McFadden, C.S.; Benayahu, Y.; Pante, E.; Thoma, J.N.; Nevarez, P.A.; France, S.C. Limitations of mitochondrial gene barcoding in Octocorallia. Mol. Ecol. Resour. 2011, 11, 19–31. [Google Scholar] [CrossRef] [PubMed]
- Benayahu, Y.; van Ofwegen, L.P.; Dai, C.F.; Jeng, M.S.; Soong, K.; Shlagman, A.; Du, S.W.; Hong, P.; Imam, N.H.; Chung, A.; et al. The octocorals of dongsha atoll (South China sea): An iterative approach to species identification using classical taxonomy anmolecular barcodes. Zool. Stud. 2018, 57, e50. [Google Scholar] [CrossRef]
- van der Ham, J.L.; Brugler, M.R.; France, S.C. Exploring the utility of an indel-rich, mitochondrial intergenic region as a molecular barcode for bamboo corals (Octocorallia: Isididae). Mar. Genomics 2009, 2, 183–192. [Google Scholar] [CrossRef]
- Baco, A.R.; Cairns, S.D. Comparing molecular variation to morphological species designations in the deep-sea coral Narella reveals new insights into seamount coral ranges. PLoS ONE 2012, 7, e45555. [Google Scholar] [CrossRef] [Green Version]
- Ross, L.K.; Ross, R.E.; Stewart, H.A.; Howell, K.L. The influence of data resolution on predicted distribution and estimates of extent of current protection of three “listed” deep-sea habitats. PLoS ONE 2015, 10, e140061. [Google Scholar] [CrossRef] [Green Version]
- Esri Inc. ArcGIS Pro; Version 2.8; Esri Inc.: Redlands, CA, USA, 2020. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Brugler, M.R.; France, S.C. The mitochondrial genome of a deep-sea bamboo coral (Cnidaria, Anthozoa, Octocorallia, Isididae): Genome structure and putative origins of replication are not conserved among octocorals. J. Mol. Evol. 2008, 67, 125. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, J.A.; McFadden, C.; France, S.; Lasker, H. Molecular Phylogenetic analyses of shallow-water Caribbean octocorals. Mar. Biol. 2003, 142, 975–987. [Google Scholar] [CrossRef]
- Pante, E.; France, S.C.; Couloux, A.; Cruaud, C.; McFadden, C.S.; Samadi, S.; Watling, L. Deep-sea origin and in-situ diversification of chrysogorgiid octocorals. PLoS ONE 2012, 7, e38357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFadden, C.S.; Tullis, I.D.; Breton Hutchinson, M.; Winner, K.; Sohm, J.A. Variation in Coding (NADH Dehydrogenase Subunits 2, 3, and 6) and Noncoding Intergenic Spacer Regions of the Mitochondrial Genome in Octocorallia (Cnidaria: Anthozoa). Mar. Biotechnol. 2004, 6, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Easton, E.E.; Hicks, D. 161 Primers for PCR-Amplification of Octocorallia Mitochondrial Genome Fragments. Available online: https://figshare.com/articles/dataset/161_primers_for_PCR-amplification_of_Octocorallia_mitochondrial_genome_fragments/19617519 (accessed on 3 May 2022).
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Lanfear, R.; Calcott, B.; Ho, S.Y.W.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef]
- Saucier, E.H.; France, S.C.; Watling, L. Toward a revision of the bamboo corals: Part 3, deconstructing the Family Isididae. Zootaxa 2021, 5047, 247–272. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; DeWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pons, J.; Barraclough, T.G.; Gomez-Zurita, J.; Cardoso, A.; Duran, D.P.; Hazell, S.; Kamoun, S.; Sumlin, W.D.; Vogler, A.P. Sequence-based species delimitation for the DNA taxonomy of undescribed insects. Syst. Biol. 2006, 55, 595–609. [Google Scholar] [CrossRef] [Green Version]
- Reid, N.M.; Carstens, B.C. Phylogenetic estimation error can decrease the accuracy of species delimitation: A Bayesian implementation of the general mixed Yule-coalescent model. BMC Evol. Biol. 2012, 12, 196. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A general species delimitation method with applications to phylogenetic placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software Platform for Bayesian Evolutionary Analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Douglas, J.; Zhang, R.; Bouckaert, R. Adaptive dating and fast proposals: Revisiting the phylogenetic relaxed clock model. PLoS Comput. Biol. 2021, 17, e1008322. [Google Scholar] [CrossRef]
- Fujisawa, T.; Ezard, T.; Barraclough, T.G. Splits: Species’ limits by threshold statistics. R Packag. Version 2013, 1, r29. [Google Scholar]
- Cairns, S.D.; Bayer, F.M. Studies on western Atlantic Octocorallia (Coelenterata: Anthozoa). Part 5: The genera Plumarella Gray, 1870; Acanthoprimnoa, n. gen.; and Candidella Bayer, 1954. Proc. Biol. Soc. Washingt. 2004, 117, 447–487. [Google Scholar]
- Cairns, S.D. Studies on western Atlantic Octocorallia (Coelenterata: Anthozoa). Part 6: The genera Primnoella Gray, 1858; Thouarella Gray, 1870; Dasystenella Versluys, 1906. Proc. Biol. Soc. Washingt. 2006, 119, 161–194. [Google Scholar] [CrossRef]
- Cairns, S.D. Studies on western Atlantic Octocorallia (Gorgonacea: Primnoidae). Part 8: New records of Primnoidae from the New England and Corner Rise Seamounts. Proc. Biol. Soc. Washingt. 2007, 120, 243–263. [Google Scholar] [CrossRef]
- Cairns, S.D.; Bayer, F.M. A review of the genus Primnoa (Octocorallia: Gorgonacea: Primnoidae), with the description of two new species. Bull. Mar. Sci. 2005, 77, 225–256. [Google Scholar]
- Cairns, S.D.; Taylor, M.L. An illustrated key to the species of the genus narella (Cnidaria, octocorallia, primnoidae). Zookeys 2019, 822, 1–15. [Google Scholar] [CrossRef]
- Taylor, M.L.; Cairns, S.D.; Agnew, D.J.; Rogers, A.D. A revision of the genus Thouarella Gray, 1870 (Octocorallia: Primnoidae), including an illustrated dichotomous key, a new species description, and comments on Plumarella Gray, 1870 and Dasystenella, Versluys, 1906. Zootaxa 2013, 3602, 1–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watling, L.; France, S.C. A new genus and species of bamboo coral (Octocorallia: Isididae: Keratoisidinae) from the New England seamounts. Bull. Peabody Museum Nat. Hist. 2011, 52, 209–221. [Google Scholar] [CrossRef]
- Saucier, E.H.; Sajjadi, A.; France, S.C. A taxonomic review of the genus Acanella (Cnidaria: Octocorallia: Isididae) in the North Atlantic Ocean, with descriptions of two new species. Zootaxa 2017, 4323, 359–390. [Google Scholar] [CrossRef]
- Bayer, F.M.; Stefani, J. New and previously known taxa of isidid octocorals (Coelenterata: Gorgonacea), partly from Antarctic waters. Proc. Biol. Soc. Washingt. 1987, 100, 937–991. [Google Scholar]
- Hadley, A. CombineZP—Image Stacking Software. 2010. Available online: https://combinezp.software.informer.com/ (accessed on 3 May 2022).
- Bayer, F.M.; Grasshoff, M.; Verseveldt, J. Illustrated Trilingual Glossary of Morphological and Anatomical Terms Applied to Octocorallia; Brill Archive: Leiden, The Netherlands, 1983; ISBN 9004071393. [Google Scholar]
- Hogan, R.; Hopkins, K.; Wheeler, A.; Allcock, A.; Yesson, C. Novel diversity in mitochondrial genomes of deep-sea Pennatulacea (Cnidaria: Anthozoa: Octocorallia). Mitochondrial DNA Part A 2019, 30, 764–777. [Google Scholar] [CrossRef] [Green Version]
- Watling, L.; France, S.C. Toward a revision of the bamboo corals: Part 2, untangling the genus Lepidisis (Octocorallia: Isididae). Bull. Peabody Museum Nat. Hist. 2021, 62, 97–110. [Google Scholar] [CrossRef]
- Dueñas, L.; Sanchez, J.A. Character lability in deep-sea bamboo corals (Octocorallia, Isididae, Keratoisidinae). Mar. Ecol. Prog. Ser. 2009, 397, 11–23. [Google Scholar] [CrossRef] [Green Version]
- Grasshoff, M. Die Gorgonaria des Expeditionen von “Travailleur” 1880–1882 und “Talisman” 1883 (Cnidaria, Anthozoa). Bull. Du Muséum Natl. D’histoire Nat. Sect. A Zool. Biol. Écologie Anim. 1986, 8, 9–38. [Google Scholar]
- Alderslade, P.; McFadden, C.S. A new genus and species of the family Isididae (Coelenterata: Octocorallia) from a CMAR Biodiversity study, and a discussion on the subfamilial placement of some nominal isidid genera. Zootaxa 2012, 3154, 21–39. [Google Scholar] [CrossRef]
- Radice, V.Z.; Quattrini, A.M.; Wareham, V.E.; Edinger, E.N.; Cordes, E.E. Vertical water mass structure in the North Atlantic influences the bathymetric distribution of species in the deep-sea coral genus Paramuricea. Deep. Res. Part I Oceanogr. Res. Pap. 2016, 116, 253–263. [Google Scholar] [CrossRef]
- Quattrini, A.M.; Herrera, S.; Adams, J.M.; Grinyó, J.; Allcock, A.L.; Shuler, A.; Wirshing, H.H.; Cordes, E.E.; McFadden, C.S. Phylogeography of Paramuricea: The Role of Depth and Water Mass in the Evolution and Distribution of Deep-Sea Corals. Front. Mar. Sci. 2022, 9, 849402. [Google Scholar] [CrossRef]
- Prada, C.; Schizas, N.V.; Yoshioka, P.M. Phenotypic plasticity or speciation? A case from a clonal marine organism. BMC Evol. Biol. 2008, 8, 47. [Google Scholar] [CrossRef] [Green Version]
- West, J.M.; Harvell, C.D.; Walls, A.M. Morphological plasticity in a gorgonian coral (Briareum asbestinum) over a depth cline. Mar. Ecol. Prog. Ser. 1993, 94, 61–69. [Google Scholar] [CrossRef]
- Everett, M.V.; Park, L.K. Exploring deep-water coral communities using environmental DNA. Deep. Res. Part II Top. Stud. Oceanogr. 2018, 150, 229–241. [Google Scholar] [CrossRef]
- McFadden, C.S.; Brown, A.S.; Brayton, C.; Hunt, C.B.; van Ofwegen, L.P. Application of DNA barcoding in biodiversity studies of shallow-water octocorals: Molecular proxies agree with morphological estimates of species richness in Palau. Coral Reefs 2014, 33, 275–286. [Google Scholar] [CrossRef]
- McFadden, C.S.; Haverkort-Yeh, R.; Reynolds, A.M.; Halàsz, A.; Quattrini, A.M.; Forsman, Z.H.; Benayahu, Y.; Toonen, R.J. Species boundaries in the absence of morphological, ecological or geographical differentiation in the Red Sea octocoral genus Ovabunda (Alcyonacea: Xeniidae). Mol. Phylogenet. Evol. 2017, 112, 174–184. [Google Scholar] [CrossRef] [Green Version]
- Calderón, I.; Garrabou, J.; Aurelle, D. Evaluation of the utility of COI and ITS markers as tools for population genetic studies of temperate gorgonians. J. Exp. Mar. Biol. Ecol. 2006, 336, 184–197. [Google Scholar] [CrossRef]
- Dellicour, S.; Flot, J.F. The hitchhiker’s guide to single-locus species delimitation. Mol. Ecol. Resour. 2018, 18, 1234–1246. [Google Scholar] [CrossRef]
- Morris, K.J.; Tyler, P.A.; Masson, D.G.; Huvenne, V.I.A.; Rogers, A.D. Distribution of cold-water corals in the Whittard Canyon, NE Atlantic Ocean. Deep. Res. Part II Top. Stud. Oceanogr. 2013, 92, 136–144. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.M.; Harvey, S.M.; Lamont, P.A.; Gage, J.D.; Humphery, J.D. Seabed photography, environmental assessment and evidence for deep-water trawling on the continental margin west of the Hebrides. Hydrobiologia 2000, 441, 173–183. [Google Scholar] [CrossRef]
- Hughes, D.J.; Gage, J.D. Benthic metazoan biomass, community structure and bioturbation at three contrasting deep-water sites on the northwest European continental margin. Prog. Oceanogr. 2004, 63, 29–55. [Google Scholar] [CrossRef]
- Sayago-Gil, M.; Durán-Muñoz, P.; Javier Murillo, F.; Díaz-del-Río, V.; Serrano, A.; Miguel Fernández-Salas, L. A study of Geomorphological Features of the Seabed and the Relationship to Deep-Sea Communities on the Western Slope of Hatton Bank (Ne Atlantic Ocean). In Seafloor Geomorphology as Benthic Habitat; Harris, P.T., Baker, E.K., Eds.; Elsevier: Amsterdam, The Netherlands, 2012; pp. 751–761. ISBN 9780123851406. [Google Scholar]
- Durán Muñoz, P.; Murillo, F.J.; Sayago-Gil, M.; Serrano, A.; Laporta, M.; Otero, I.; Gómez, C. Effects of deep-sea bottom longlining on the Hatton Bank fish communities and benthic ecosystem, north-east Atlantic. J. Mar. Biol. Assoc. UK 2011, 91, 939–952. [Google Scholar] [CrossRef]
- de Neves, B.M.; Edinger, E.; Hillaire-Marcel, C.; Saucier, E.H.; France, S.C.; Treble, M.A.; Wareham, V.E. Deep-water bamboo coral forests in a muddy Arctic environment. Mar. Biodivers. 2015, 45, 867–871. [Google Scholar] [CrossRef]
- Altuna, Á.; Ríos, P.; De Gijón, C.O.; De Oceanografía, I.E.; Asturias, C.P. De Calcaxonian octocorals (Cnidaria: Anthozoa) from demersales, ecomarg and indemares expeditions to bathyal waters off north and northwest Spain (northeast Atlantic). In Proceedings of the XVIII Iberian Symposium of Marine Biology Studies, Donostia-San Sebastián, Spain, 11–14 September 2012; p. 33212. [Google Scholar]
- Lapointe, A.; Watling, L.; Gontz, A.M. Deep-sea benthic megafaunal communities on the New England and Corner Rise Seamounts, Northwest Atlantic Ocean. In Seafloor Geomorphology as Benthic Habitat; Harris, P.T., Baker, E., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 917–932. ISBN 978-0-12-814960-7. [Google Scholar]
- Lapointe, A.E.; Watling, L.; France, S.C.; Auster, P.J. Megabenthic assemblages in the lower bathyal (700–3000 m) on the New England and Corner Rise Seamounts, Northwest Atlantic. Deep. Res. Part I Oceanogr. Res. Pap. 2020, 165, 103366. [Google Scholar] [CrossRef]
- Cairns, S.D. The marine fauna of New Zealand: New zealand primnoidae (anthozoa: Alcyonacea) part 1. genera narella, narelloides, metanarella, calyptrophora, and helicoprimnoa. NIWA Biodivers. Mem. 2012, 126, 5–72. [Google Scholar]
- Stephens, J. Alcyonarian and Madreporarian Corals of the Irish Coasts; HM Stationery Office: London, UK, 1909. [Google Scholar]
- Bo, M.; Coppari, M.; Betti, F.; Massa, F.; Gay, G.; Cattaneo-Vietti, R.; Bavestrello, G. Unveiling the deep biodiversity of the Janua Seamount (Ligurian Sea): First Mediterranean sighting of the rare Atlantic bamboo coral Chelidonisis aurantiaca Studer, 1890. Deep. Res. Part I Oceanogr. Res. Pap. 2020, 156, 103186. [Google Scholar] [CrossRef]
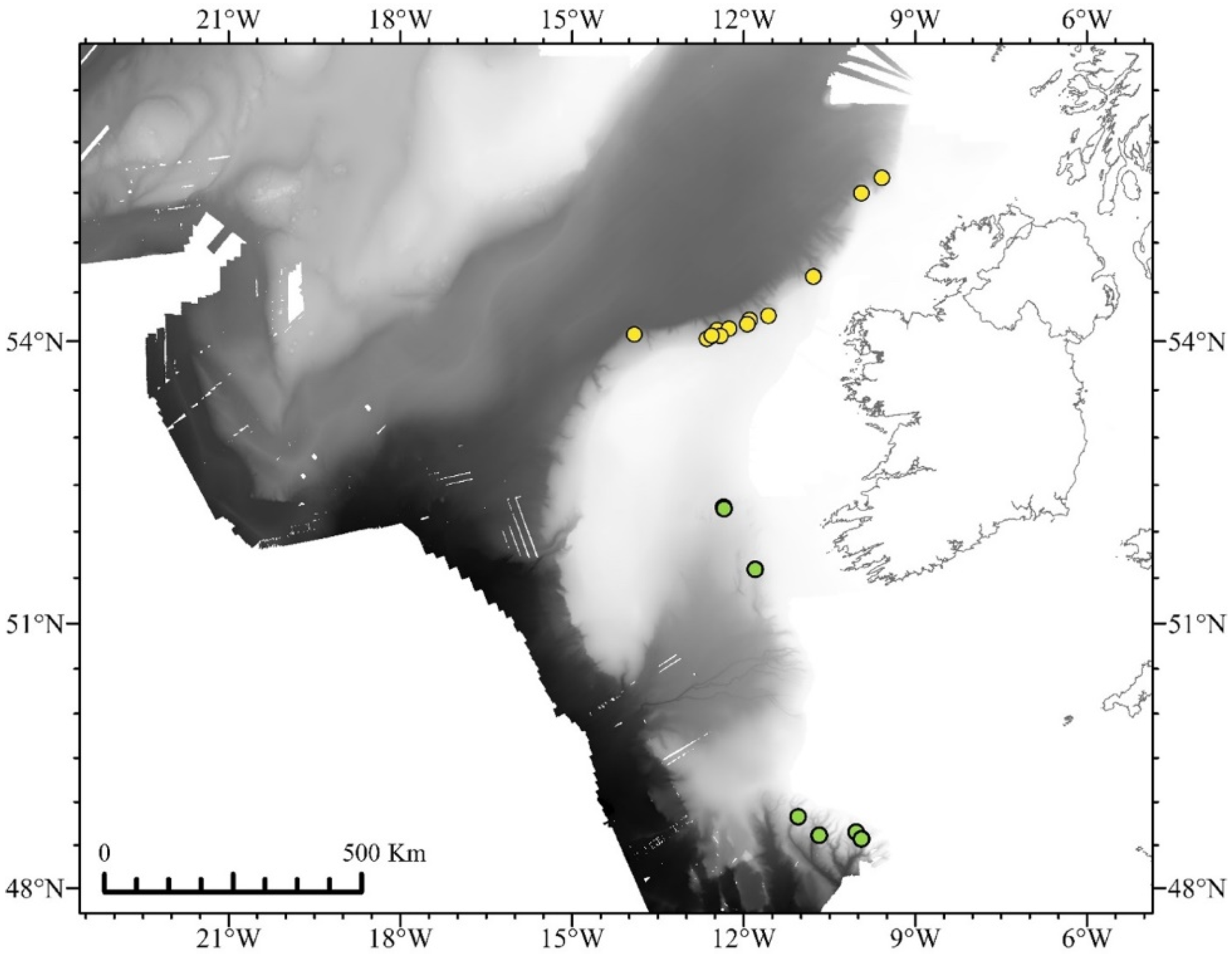




| Specimen Voucher | USNM Number | Specimen ID | Haplotype/Morphotype | mtMutS | COI + igr1 | 16s rRNA-ND2 | igr4 |
|---|---|---|---|---|---|---|---|
| CE-17-266 | 1593468 | Keratoisididae D1 sp. | 1/7 | ON971007 | ON971056 | ON971105 | ON971154 |
| CE-18-646 | 1593473 | Keratoisididae D1 sp. | 1/8 | ON971008 | ON971057 | ON971106 | ON971155 |
| CE-18-508 | 1593474 | Keratoisididae D1 sp. | 2/9 | ON971009 | ON971058 | ON971107 | ON971156 |
| CE-17-269 | 1593475 | Keratoisididae B1 sp. | 3/12 | ON971010 | ON971059 | ON971108 | ON971157 |
| CE-17-174 | 1593476 | Keratoisididae B1 sp. | 4/10 | ON971011 | ON971060 | ON971109 | ON971158 |
| CE-17-206 | 1593477 | Keratoisididae B1 sp. | 5/13 | ON971012 | ON971061 | ON971110 | ON971159 |
| CE-17-146 | 1593481 | Keratoisididae B1 sp. | 6/10 | ON971013 | ON971062 | ON971111 | ON971160 |
| CE-17-424 | 1593483 | Keratoisididae B1 sp. | 6/11 | ON971014 | ON971063 | ON971112 | ON971161 |
| CE-17-201 | 1593489 | Eknomisis sp. | 7/4 | ON971015 | ON971064 | ON971113 | ON971162 |
| CE-18-129 | 1593490 | Eknomisis sp. | 8/5 | ON971016 | ON971065 | ON971114 | ON971163 |
| CE-18-387 | 1593492 | Eknomisis sp. | 8/6 | ON971017 | ON971066 | ON971115 | ON971164 |
| CE-18-288 | 1593493 | Keratoisididae D2 sp. | 9/1 | ON971018 | ON971067 | ON971116 | ON971165 |
| CE-17-222 | 1593498 | Keratoisididae D2 sp. | 10/3 | ON971019 | ON971068 | ON971117 | ON971166 |
| CE-18-039 | 1593495 | Keratoisididae D2 sp. | 11/2 | ON971020 | ON971069 | ON971118 | ON971167 |
| CE-18-203 | 1593497 | Keratoisididae I1 sp. | 12/15 | ON971021 | ON971070 | ON971119 | ON971168 |
| CE-17-234 | 1593498 | Keratoisididae I1 sp. | 13/16 | ON971022 | ON971071 | ON971120 | ON971169 |
| CE-18-063 | 1593499 | Keratoisididae I1 sp. | 13/17 | ON971023 | ON971072 | ON971121 | ON971170 |
| CE-18-415 | 1593500 | Keratoisididae I1 sp. | 13/18 | ON971024 | ON971073 | ON971122 | ON971171 |
| CE-18-110 | 1593503 | Keratoisididae I1 sp. | 14/17 | ON971025 | ON971074 | ON971123 | ON971172 |
| CE-17-125 | 1593509 | Keratoisididae I1 sp. | 15/19 | ON971026 | ON971075 | ON971124 | ON971173 |
| CE-17-287 | 1593512 | Keratoisididae C1 sp. | 16/28 | ON971027 | ON971076 | ON971125 | ON971174 |
| CE-18-242 | 1593514 | Keratoisididae C1 sp. | 16/29 | ON971028 | ON971077 | ON971126 | ON971175 |
| CE-17-356 | 1593516 | Keratoisididae C1 sp. | 17/30 | ON971029 | ON971078 | ON971127 | ON971176 |
| CE-17-276 | 1593517 | Keratoisididae J3 sp. | 18/20 | ON971030 | ON971079 | ON971128 | ON971177 |
| CE-17-274 | 1593518 | Keratoisididae J3 sp. | 19/21 | ON971031 | ON971080 | ON971129 | ON971178 |
| CE-18-538 | 1593524 | Keratoisididae J3 sp. | 19/22 | ON971032 | ON971081 | ON971130 | ON971179 |
| CE-17-367 | 1593526 | Keratoisididae J3 sp. | 20/23 | ON971033 | ON971082 | ON971131 | ON971180 |
| CE-17-304 | 1593531 | Acanella arbuscula | 21/24 | ON971034 | ON971083 | ON971132 | ON971181 |
| CE-17-357 | 1593535 | Acanella arbuscula | 21/25 | ON971035 | ON971084 | ON971133 | ON971182 |
| CE-17-425 | 1593532 | Acanella arbuscula | 21/26 | ON971036 | ON971085 | ON971134 | ON971183 |
| CE-18-339 | 1593536 | Acanella sp. | 22/27 | ON971037 | ON971086 | ON971135 | ON971184 |
| CE-17-216 | 1593537 | Keratoisididae F1 sp. | 23/14 | ON971038 | ON971087 | ON971136 | ON971185 |
| CE-17-181 | 1593580 | Chelidonisis aurantiaca | 24/44 | ON971039 | ON971088 | ON971137 | ON971186 |
| CE-17-364 | 1593538 | Chrysogorgia Group 7 sp. | 25/37 | ON971040 | ON971089 | ON971138 | ON971187 |
| CE-17-271 | 1593539 | Chrysogorgia Group 7 sp. | 26/38 | ON971041 | ON971090 | ON971139 | ON971188 |
| CE-17-129 | 1593543 | Chrysogorgia Group 3 sp. | 27/39 | ON971042 | ON971091 | ON971140 | ON971189 |
| CE-17-283 | 1593545 | Chrysogorgia Group 3 sp. | 27/40 | ON971043 | ON971092 | ON971141 | ON971190 |
| CE-18-292 | 1593548 | Chrysogorgia Group 3 sp. | 27/41 | ON971044 | ON971093 | ON971142 | ON971191 |
| CE-18-418 | 1593551 | Dasygorgia sp. | 28/42 | ON971045 | ON971094 | ON971143 | ON971192 |
| CE-18-237 | 1593554 | Chrysogorgia Group 1 sp. | 29/43 | ON971046 | ON971095 | ON971144 | ON971193 |
| CE-18-434 | 1593558 | Narella bellissima | 30/32 | ON971047 | ON971096 | ON971145 | ON971194 |
| CE-17-290 | 1593559 | Candidella imbricata | 31/34 | ON971048 | ON971097 | ON971146 | ON971195 |
| CE-18-321 | 1593564 | Narella versluysi | 32/33 | ON971049 | ON971098 | ON971147 | ON971196 |
| CE-17-095 | 1593566 | Primnoa sp. | 33/31 | ON971050 | ON971099 | ON971148 | ON971197 |
| CE-17-160 | 1593567 | Primnoa sp. | 34/31 | ON971051 | ON971100 | ON971149 | ON971198 |
| CE-17-082 | 1593568 | Primnoa sp. | 35/31 | ON971052 | ON971101 | ON971150 | ON971199 |
| CE-17-101 | 1593572 | Primnoa sp. | 36/31 | ON971053 | ON971102 | ON971151 | ON971200 |
| CE-18-261 | 1593574 | Thouarella grasshoffi | 37/35 | ON971054 | ON971103 | ON971152 | ON971201 |
| CE-18-568 | 1593579 | Calyptrophora clinata | 38/36 | ON971055 | ON971104 | ON971153 | ON971202 |
| Morphotype (Haplotype, Figures) | Designation | Colony Shape | Branching | Polyp Morphology | Coenenchyme | Tentacles | Polyp Body Sclerites | Tentacle Sclerites | Pharynx Sclerites | Coenenchyme Sclerites |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 (9; Figure 2l and Figure S1) | Keratoisididae D2 sp. | Planar | Sparse dichotomous, internodal | Cylindrical. Originate from all sides of coenenchyme | Thick. Cream to pink | In the oral area | Needles arranged longitudinally | Blunt ended rods Needles | Thorned rodlets | Needles |
| 2 (11, Figure 2n and Figure S2) | Keratoisididae D2 sp. | Unknown | Unknown | Orange | Unknown | Folded over mouth | Rods arranged obliquely and longitudinally | Blunt ended rods and flattened rods | Unknown | Rods |
| 3 (10, Figure 2m and Figure S3) | Keratoisididae D2 sp. | Arborescent | Frequent dichotomous, internodal | Cylindrical. Orange to pink. Originate on all sides of coenenchyme | Thick | In the oral area | Needles and rods arranged longitudinally | Blunt ended rods and flattened rods | Thorned rodlets | Needles and rods |
| 4 (7, Figure 2i and Figure S4) | Eknomisis sp. | Arborescent | Sparse dichotomous, internodal | Distally tapered. Pink Originate in two planes? | Thin | In the oral area | Rods arranged obliquely | Blunt ended rods and flattened rods | Thorned rodlets | Rods |
| 5 (8, Figure 2j and Figure S5) | Eknomisis sp. | Arborescent | Sparse dichotomous, internodal | Distally tapered. Pink Originate on all sides of coenenchyme | Thin | In the oral area | Needles and rods arranged obliquely | Blunt ended rods and flattened rods | Thorned rodlets | Rods |
| 6 (8, Figure 2k and Figure S6) | Eknomisis sp. | Arborescent | Sparse dichotomous, internodal | Barrel shaped. | Thin | Folded over mouth | Needles and rods arranged obliquely | Blunt ended rods and flattened rods | Thorned rodlets | Needles and rods |
| 7 (1, Figure 2a and Figure S7) | Keratoisididae D1 sp. | Planar | Frequent dichotomous, internodal | Pink to orange. Cylindrical. | Thick | In the oral area | Needles and rods arranged obliquely and longitudinally | Blunt ended rods and flattened rods | Thorned rodlets | Unknown |
| 8 (1, Figure 2b and Figure S8) | Keratoisididae D1 sp. | Arborescent | Sparse dichotomous, internodal | Pink to orange. Cylindrical. Originate on all sides of coenenchyme | Thick | In the oral area | Needles and rods arranged obliquely | Blunt ended rods and flattened rods | Thorned rodlets | Unknown |
| 9 (2, Figure 2c and Figure S9) | Keratoisididae D1 sp. | Planar | Sparse dichotomous, internodal | Pink to orange. Cylindrical. Originate on all sides of coenenchyme | Thin | In the oral area | Needles and rods arranged obliquely | Blunt ended rods and flattened rods | Thorned rodlets | Unknown |
| 10 (4, Figure 2e and Figure S10) | Keratoisididae B1 sp. | Whip or planar | Branched and unbranched forms | Trumpet shape. Originate on all sides of coenenchyme | Thick | Folded over mouth | Needles and scales arranged obliquely and longitudinally | Flattened rods | Thorned rodlets | Needles and scales |
| 10 (6, Figure 2g and Figure S11) | Keratoisididae B1 sp. | Whip or planar | Branched and unbranched forms | Trumpet shaped. Originate on all sides of coenenchyme | Thick | Folded over mouth | Needles and scales arranged obliquely and longitudinally | Elongated, narrow flattened rods | Thorned rodlets | Needles and scales |
| 11 (6, Figure 2h and Figure S12) | Keratoisididae B1 sp. | Whip or planar | Branched and unbranched forms | Barrel shaped. Originate on all sides of coenenchyme | Thick | Folded over mouth | Needles and scales arranged obliquely and longitudinally | Flattened rods | Thorned rodlets | Unknown |
| 12 (3, Figure 2d and Figure S13) | Keratoisididae B1 sp. | Whip | Unbranched | Barrel shaped. Originate on all sides of coenenchyme | Thick | Folded over mouth | Needles and scales arranged obliquely and longitudinally | Flattened rods | Thorned rodlets | Unknown |
| 13 (5, Figure 2f and Figure S14) | Keratoisididae B1 sp. | Whip | Unbranched | Hourglass. Originate on all sides of coenenchyme | Thick | Folded over mouth | Needles and scales arranged obliquely and longitudinally | Flattened rods | Thorned rodlets | Needles and scales |
| 14 (23, Figure 4b and Figure S15) | Keratoisididae F1 sp. | Whip | Unbranched | Tall, narrow. Originate on all sides of coenenchyme | Thin | Folded over mouth | Needles and rods | Blunt ended rods and flattened rods | Elongated, thorned rodlets | Needles |
| 15 (12, Figure 2o and Figure S16) | Keratoisididae I1 sp. | Arborescent | Sparse dichotomous | Either barrel shaped, or L-shaped. Originate on all sides of coenenchyme | Thin | Loosely gathered over mouth | Sclerites scarce. Needles occurring in distal end. | Flattened rods | Thorny star | Unknown |
| 16 (13, Figure 3a and Figure S17) | Keratoisididae I1 sp. | Sparse bush | Di– and trichotomous, nodal | Polyps slightly flare distally. Funnel shaped | Thin | Loosely gathered over mouth | Sclerites scarce. Needles occurring in distal end. | Flattened rods | Thorny star | Unknown |
| 17 (13, Figure 3b and Figure S18) | Keratoisididae I1 sp. | Bush | Di– and trichotomous, nodal | Polyps slightly flare distally. Funnel shaped | Thin | Loosely gathered over mouth | Needles arranged longitudinally | Flattened rods | Thorny star | Needles |
| 17 (14, Figure 3d and Figure S19) | Keratoisididae I1 sp. | Bush | Di– and trichotomous, nodal | Polyps slightly flare distally. Funnel shaped | Thin | Loosely gathered over mouth | Needles and rods arranged longitudinally | Flattened rods | Unknown | Needles |
| 18 (13, Figure 3c and Figure S20) | Keratoisididae I1 sp. | Arborescent | Di– and trichotomous, nodal | Funnel shaped | Thin | Loosely gathered over mouth | Rods arranged obliquely | Flattened rods | Thorny star | Rods |
| 19 (15, Figure 3e and Figure S21) | Keratoisididae I1 sp. | Whip | Unbranched | Funnel/cylindrical shaped | Thin | Loosely gathered over mouth | Needles and rods arranged obliquely and longitudinally | Flattened rods | Thorny star | Needles and rods |
| 20 (18, Figure 3i and Figure S22) | Keratoisididae J3 sp. | Planar | Branched | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown | Unknown |
| 21 (19, Figure 3j and Figure S23) | Keratoisididae J3 sp. | Planar | Branched | Trumpet shaped, originating from all sides of the coenenchyme | Thick | Folded over mouth | Spindles and rods arranged longitudinally | Flattened rods | Flattened rods | Spindles and Needles |
| 22 (19, Figure 3k and Figure S24) | Keratoisididae J3 sp. | Planar | Di– and trichotomous, nodal | Taper distally, originating from all sides of the coenenchyme | Thin | In the oral area | Rods arranged obliquely and longitudinally | Flattened rods | Flattened rods | Rods |
| 23 (20, Figure 3l and Figure S25) | Keratoisididae J3 sp. | Planar | Dichotomous, nodal | Barrel shaped. originating from all sides of the coenenchyme | Thick | Folded over mouth | Needles and rods arranged longitudinally | Flattened rods | Sparsely toothed rods | Rods, flattened rods |
| 24 (21, Figure 3m and Figure S26) | Acanella arbuscula | Loose bush | Sparse di– and trichotomous, nodal | Funnel shaped | Thin | Folded over mouth | Rods arranged obliquely | Rods and flattened rods | Elongated thorny rods | Rods |
| 25 (21, Figure 3n and Figure S27) | Acanella arbuscula | Tight bush | Nodal | Slender, same width throughout | Thin | Folded over mouth | Needles arranged obliquely | Rods and flattened rods | Rod | Rods |
| 26 (21, Figure 3o and Figure S28) | Acanella arbuscula | Funnel shaped bush | Sparse di– and trichotomous, nodal | Barrel shaped | Thin, thickening distally | Folded over mouth | Needles arranged obliquely | Rods and flattened rods | Elongated thorny rods | Rods |
| 27 (22, Figure 4a and Figure S29) | Acanella sp. | Fan/bush | dichotomous, nodal | Barrel | Thin, thickening distally | Contracted over mouth | Rods and needles arranged obliquely | Rods and flattened rods | Elongated thorny rods | Needles and rods |
| 28 (16, Figure 3f and Figure S30) | Keratoisididae C1 sp. | Whip | Unbranched | Barrel shaped, originate on all sides | Thick | Folded over mouth | Sparse, needles, arranged obliquely and longitudinally | Flattened rods | Elongated diamond rods | Unknown |
| 29 (16, Figure 3g and Figure S31) | Keratoisididae C1 sp. | Whip | Unbranched | Barrel shaped, originate on all sides | Thick | Folded over mouth | Sparse, needles, arranged obliquely and longitudinally | Flattened rods | Elongated diamond rods | Needles and elongated rods |
| 30 (17, Figure 3h and Figure S32) | Keratoisididae C1 sp. | Whip | Unbranched | Barrel shaped, originate on all sides | Thick | Folded over mouth | Sparse, needles, arranged obliquely | Flattened rods | Elongated diamond rods | Rods and flattened rods |
| Family | Lower Bound of Species Estimate | Upper Bound of Species Estimate |
|---|---|---|
| Keratoisididae | 14 | 27 |
| Primnoidae | 6 | 7 |
| Chrysogorgiidae | 4 | 5 |
| Chelidonisididae | 1 | 1 |
| Total | 25 | 40 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morrissey, D.; Untiedt, C.B.; Croke, K.; Robinson, A.; Turley, E.; Allcock, A.L. The Biodiversity of Calcaxonian Octocorals from the Irish Continental Slope Inferred from Multilocus Mitochondrial Barcoding. Diversity 2022, 14, 576. https://doi.org/10.3390/d14070576
Morrissey D, Untiedt CB, Croke K, Robinson A, Turley E, Allcock AL. The Biodiversity of Calcaxonian Octocorals from the Irish Continental Slope Inferred from Multilocus Mitochondrial Barcoding. Diversity. 2022; 14(7):576. https://doi.org/10.3390/d14070576
Chicago/Turabian StyleMorrissey, Declan, Candice B. Untiedt, Karen Croke, Aisling Robinson, Eva Turley, and A. Louise Allcock. 2022. "The Biodiversity of Calcaxonian Octocorals from the Irish Continental Slope Inferred from Multilocus Mitochondrial Barcoding" Diversity 14, no. 7: 576. https://doi.org/10.3390/d14070576
APA StyleMorrissey, D., Untiedt, C. B., Croke, K., Robinson, A., Turley, E., & Allcock, A. L. (2022). The Biodiversity of Calcaxonian Octocorals from the Irish Continental Slope Inferred from Multilocus Mitochondrial Barcoding. Diversity, 14(7), 576. https://doi.org/10.3390/d14070576