
Diversity in Morphometric, Pomological, and Fruit-Quality Traits of Apricot (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
Data Analysis
3. Results
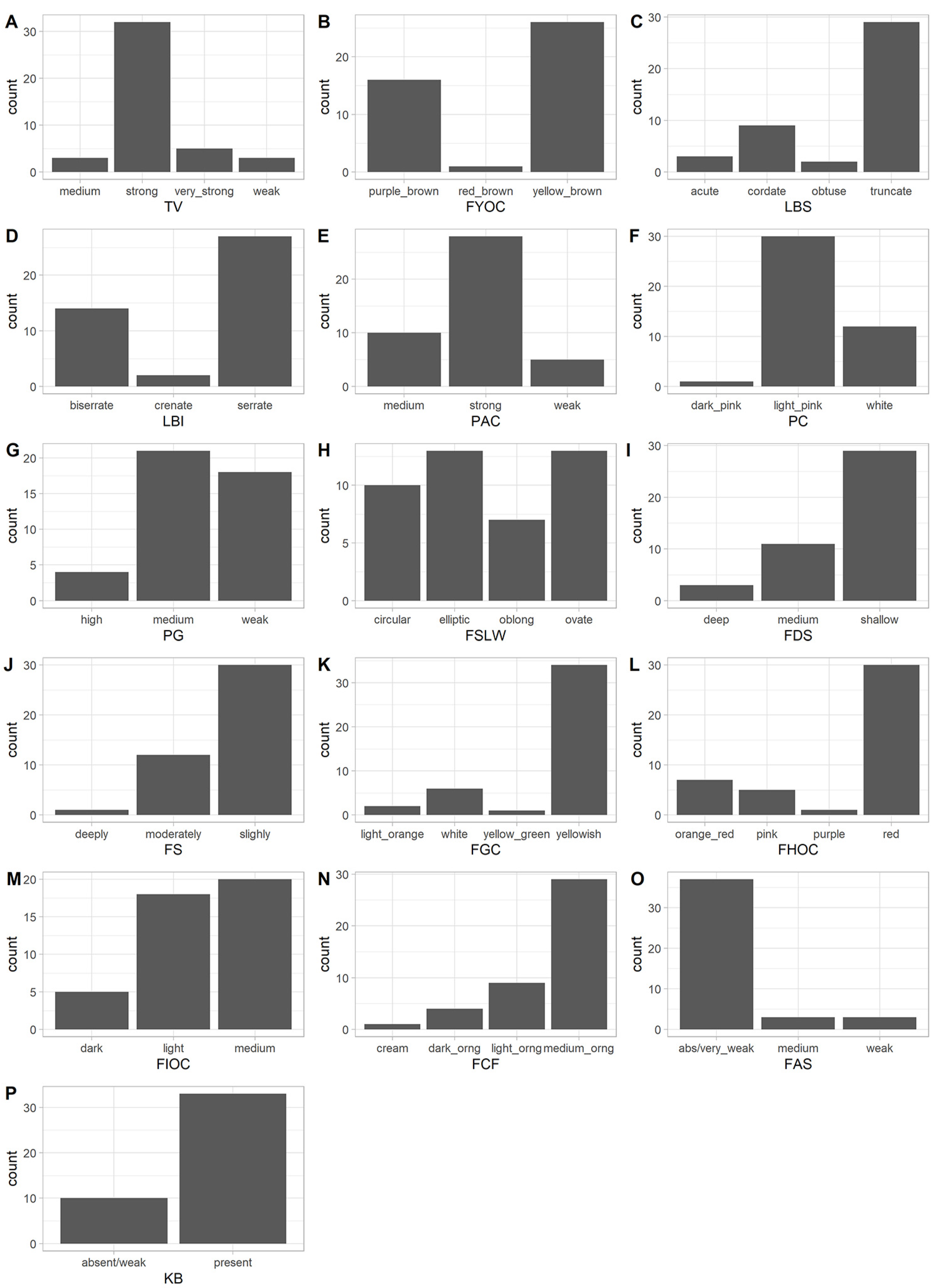
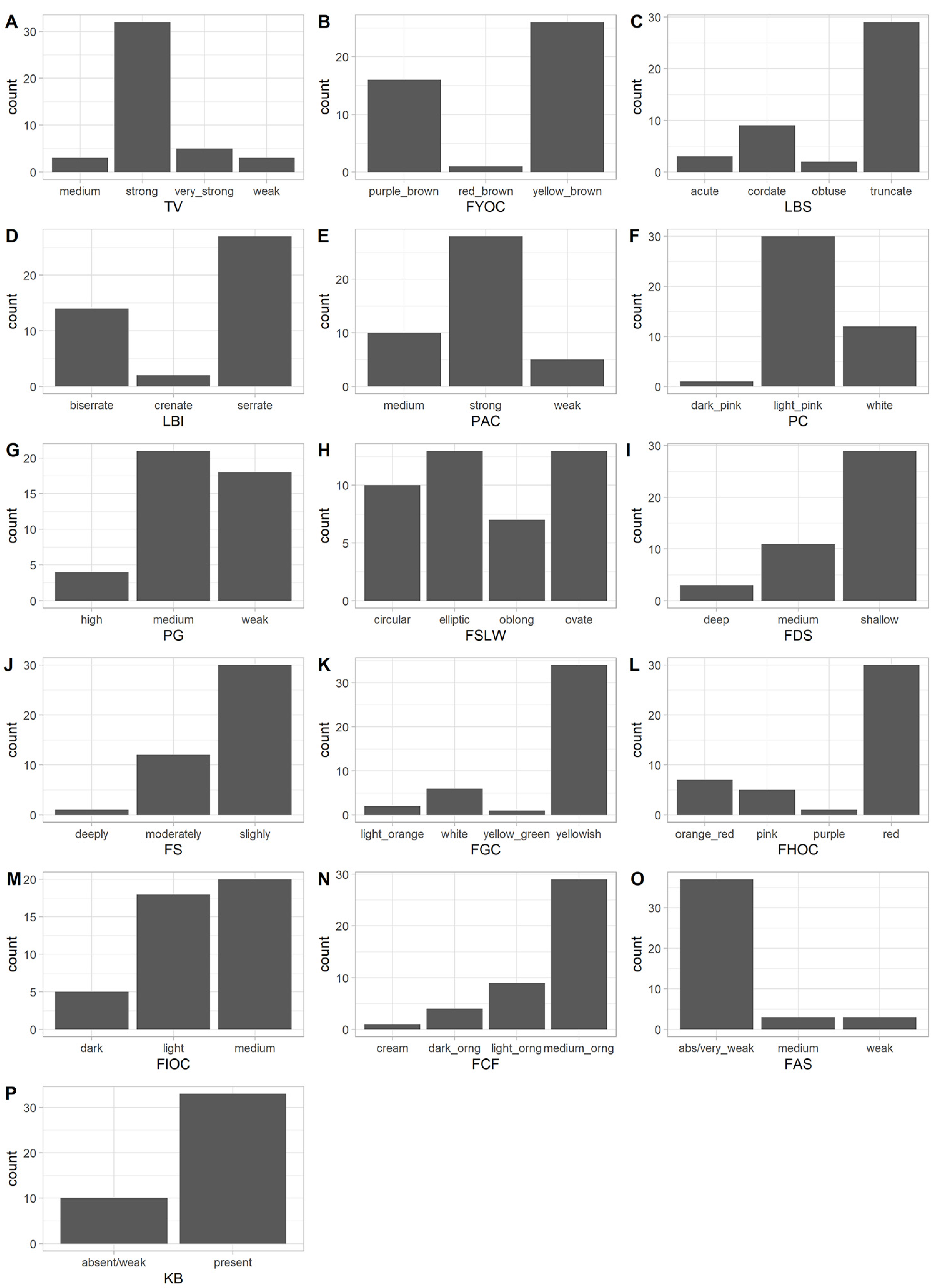
3.1. Analysis of Qualitative Traits
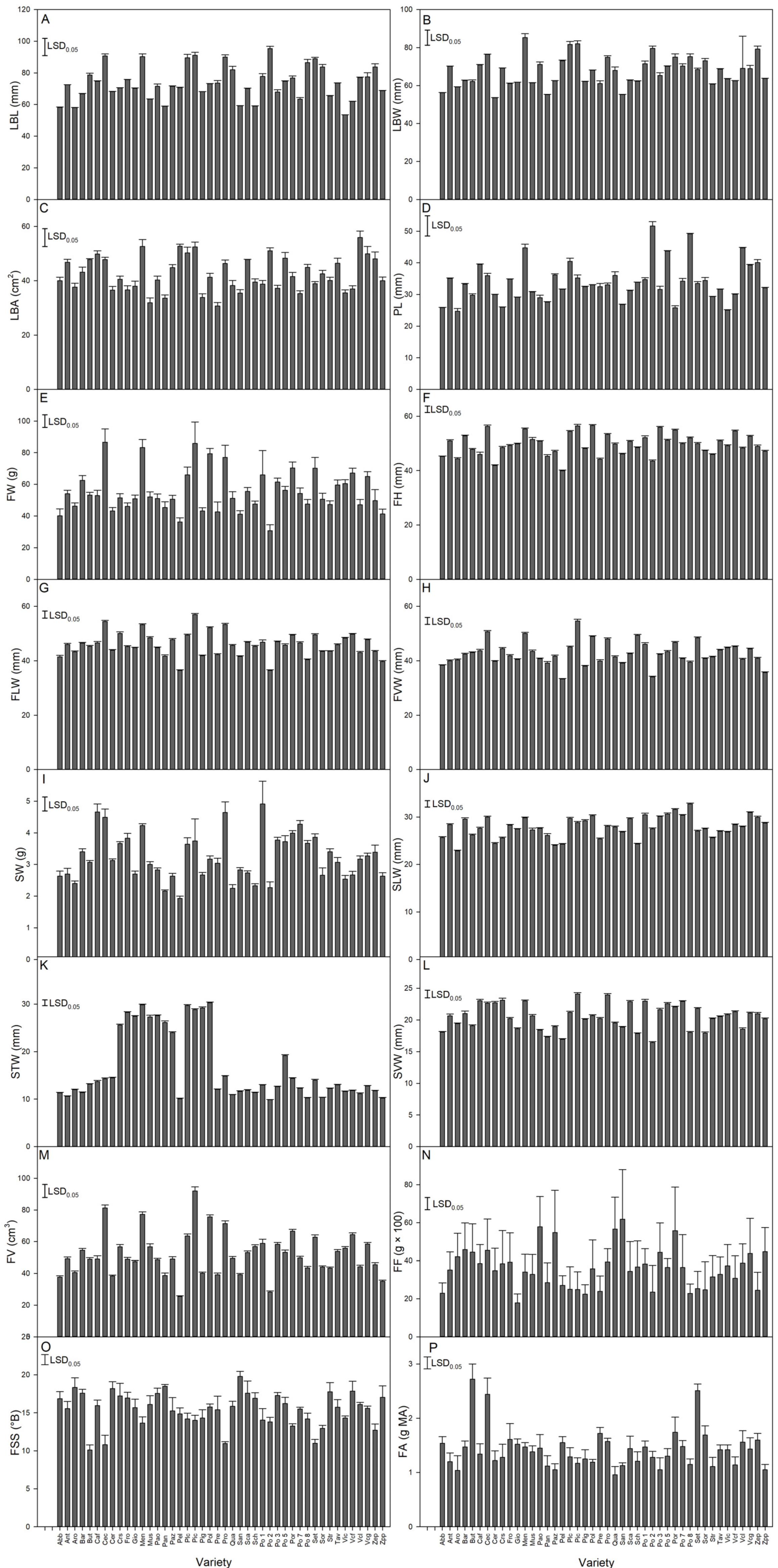
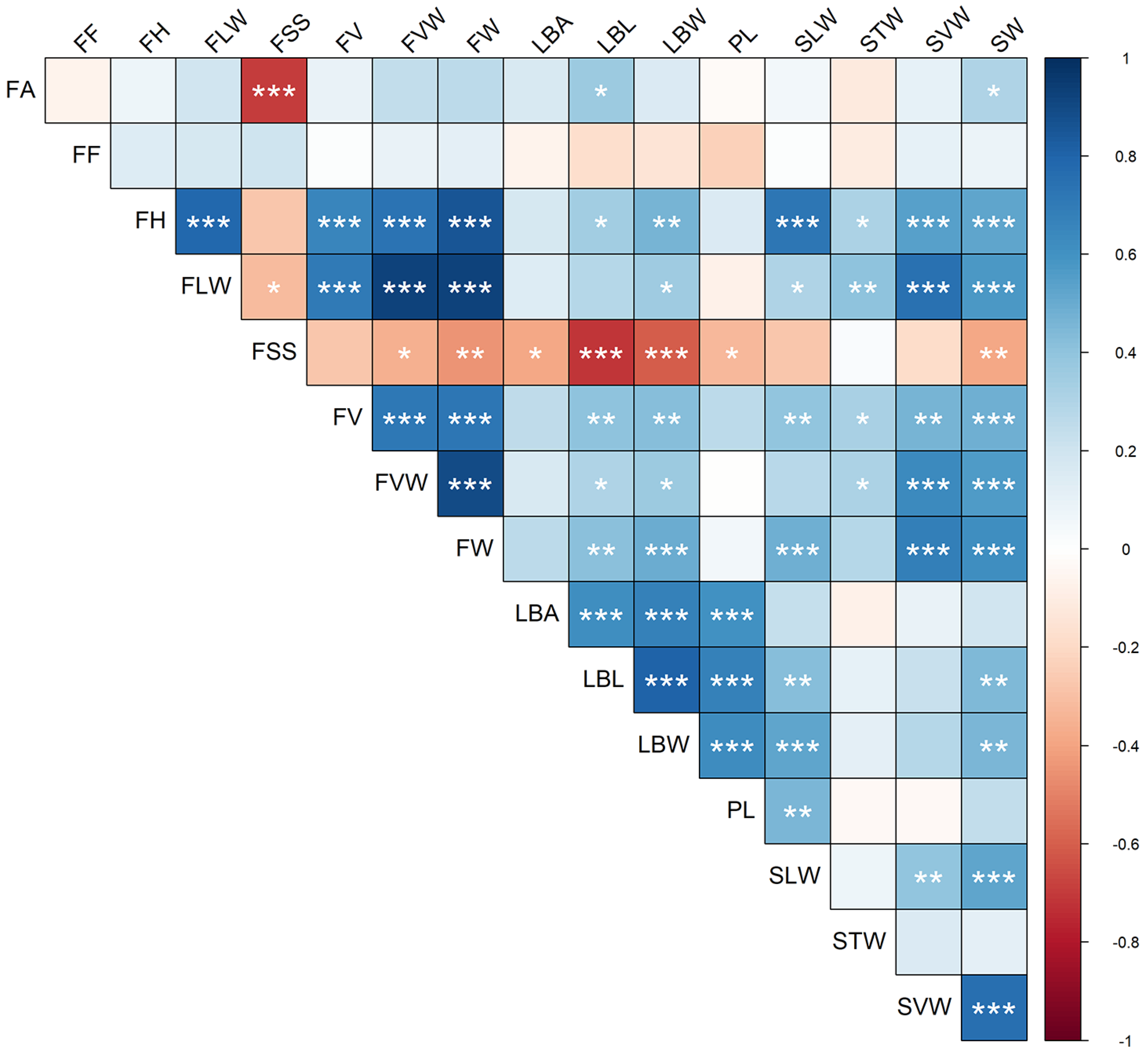
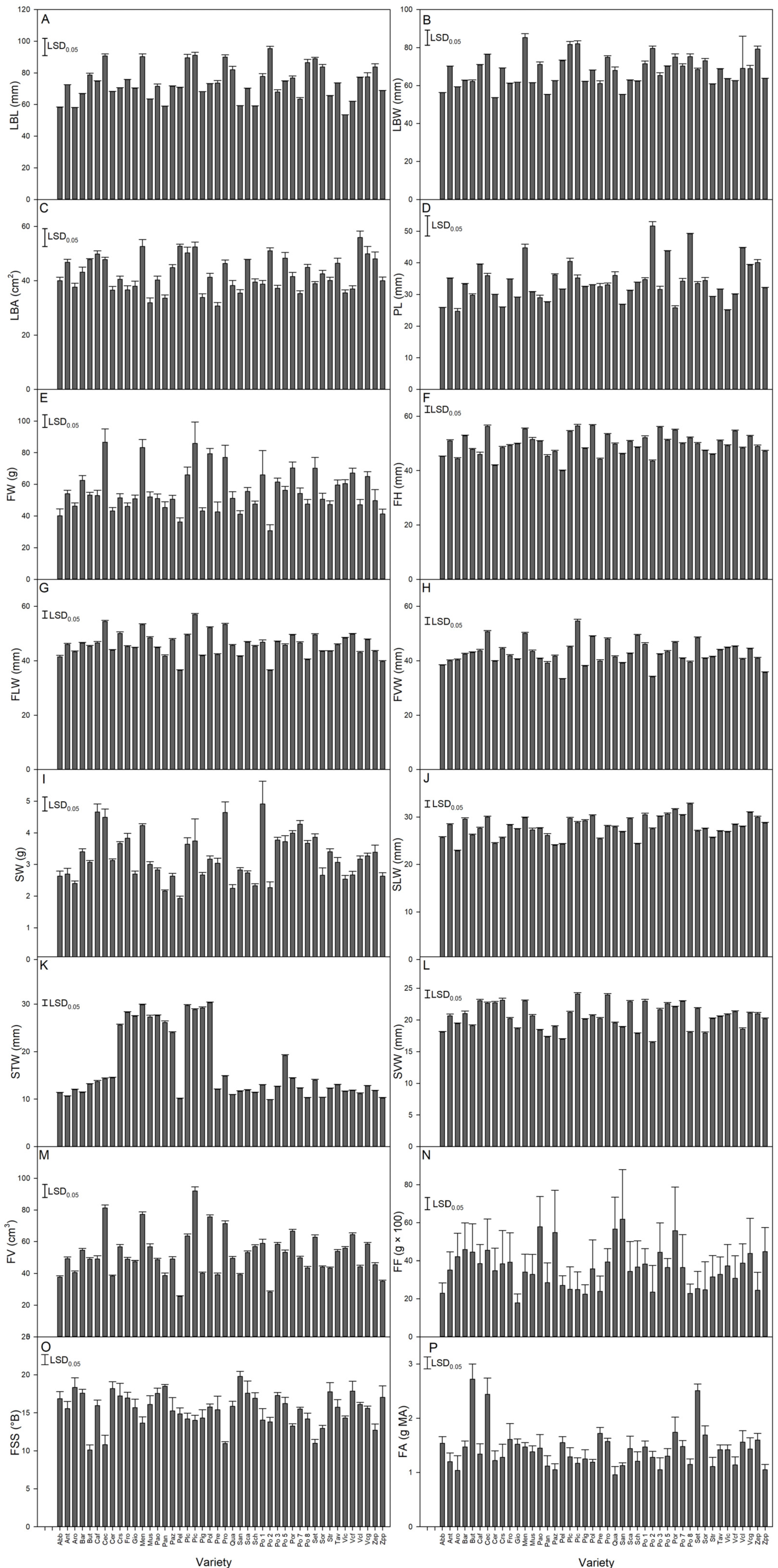
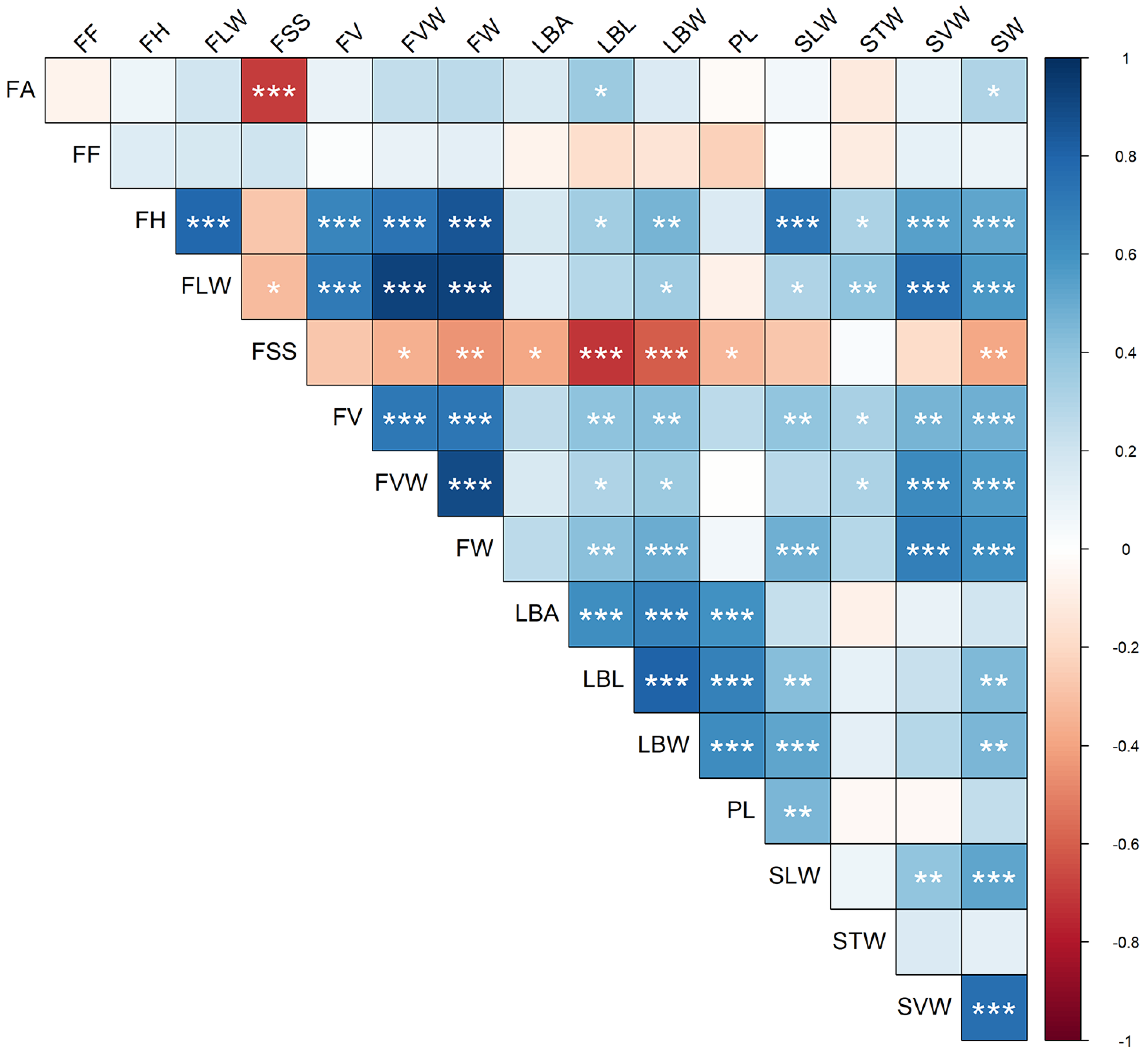
3.2. Analysis of Quantitative Traits
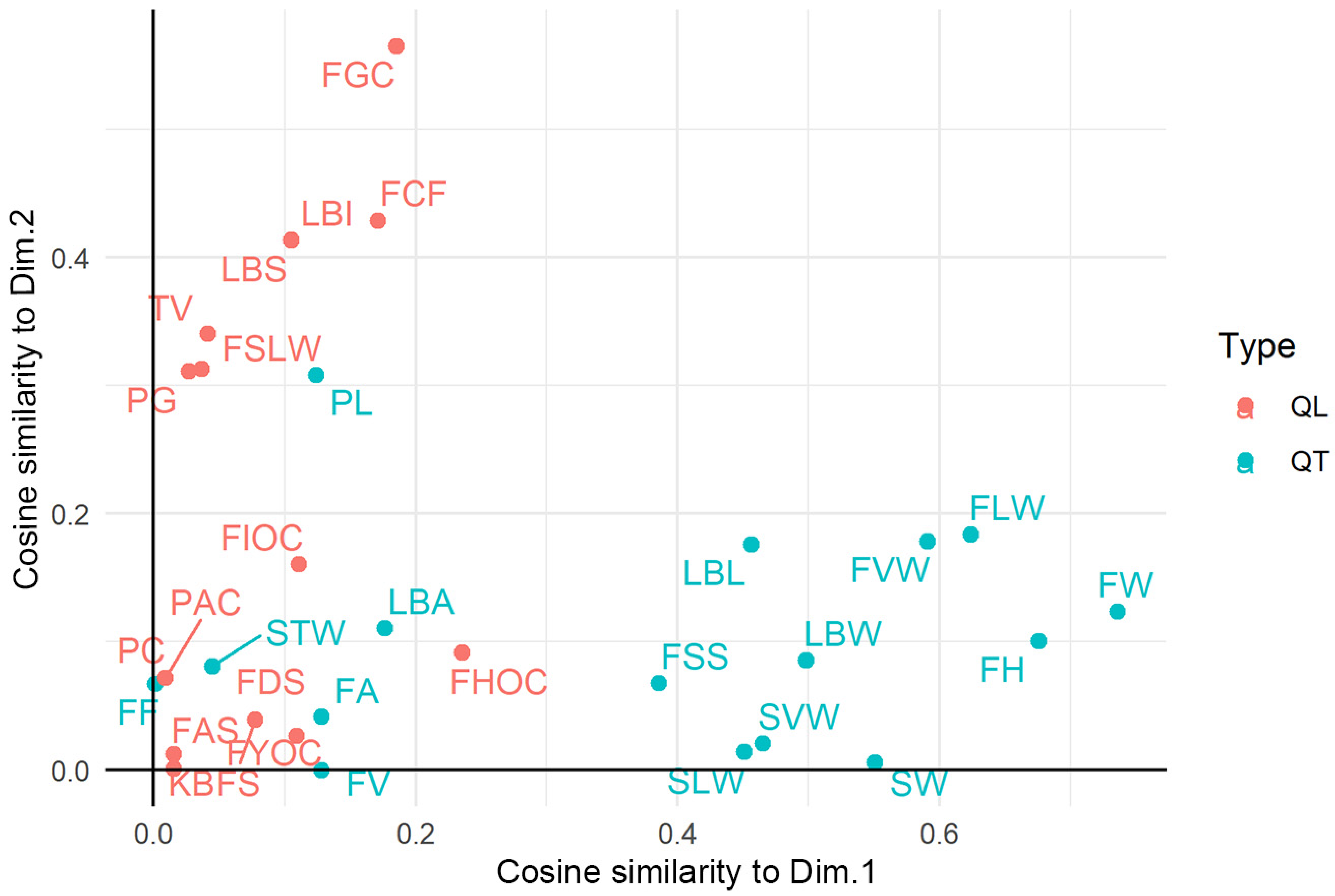
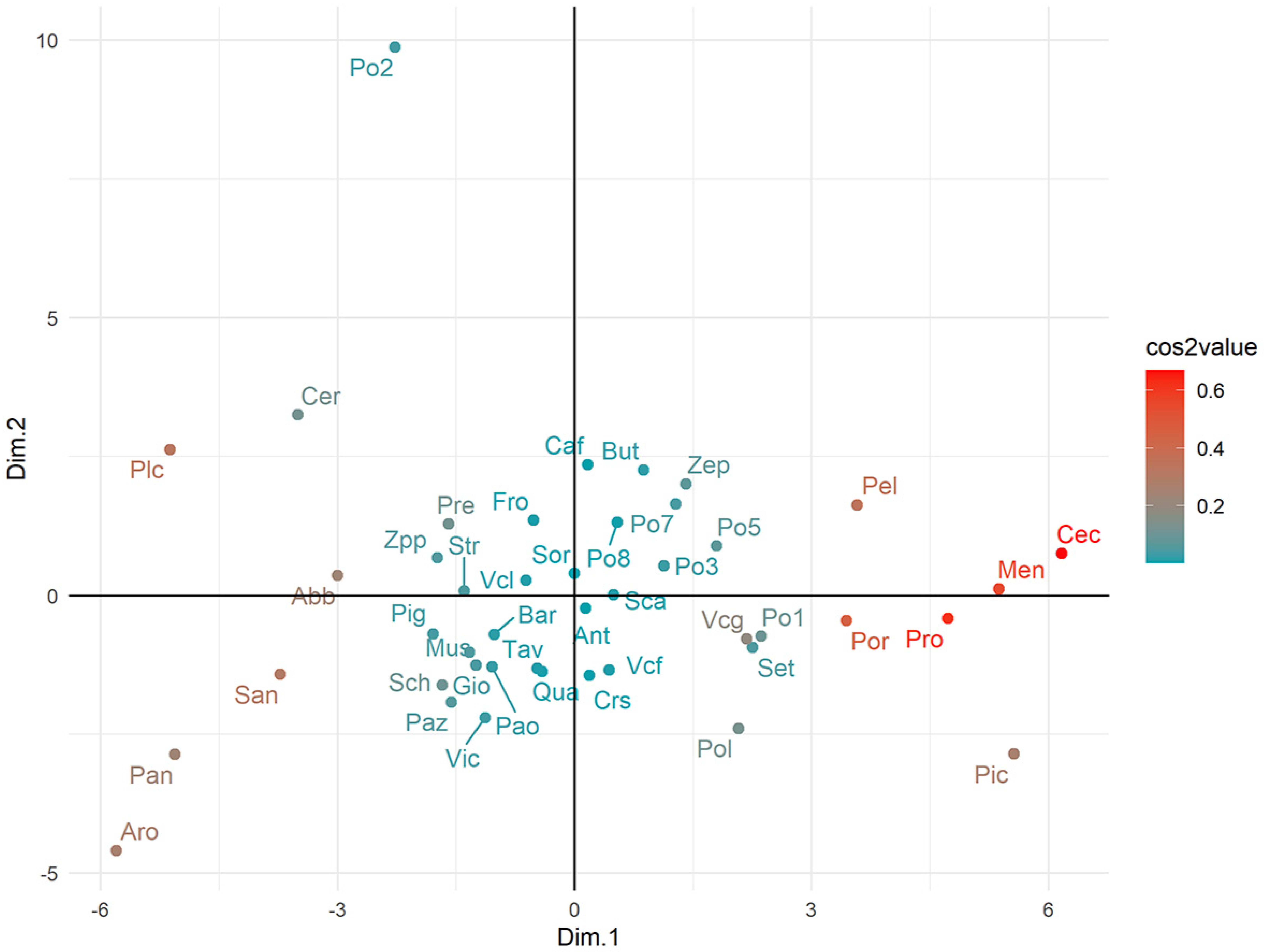
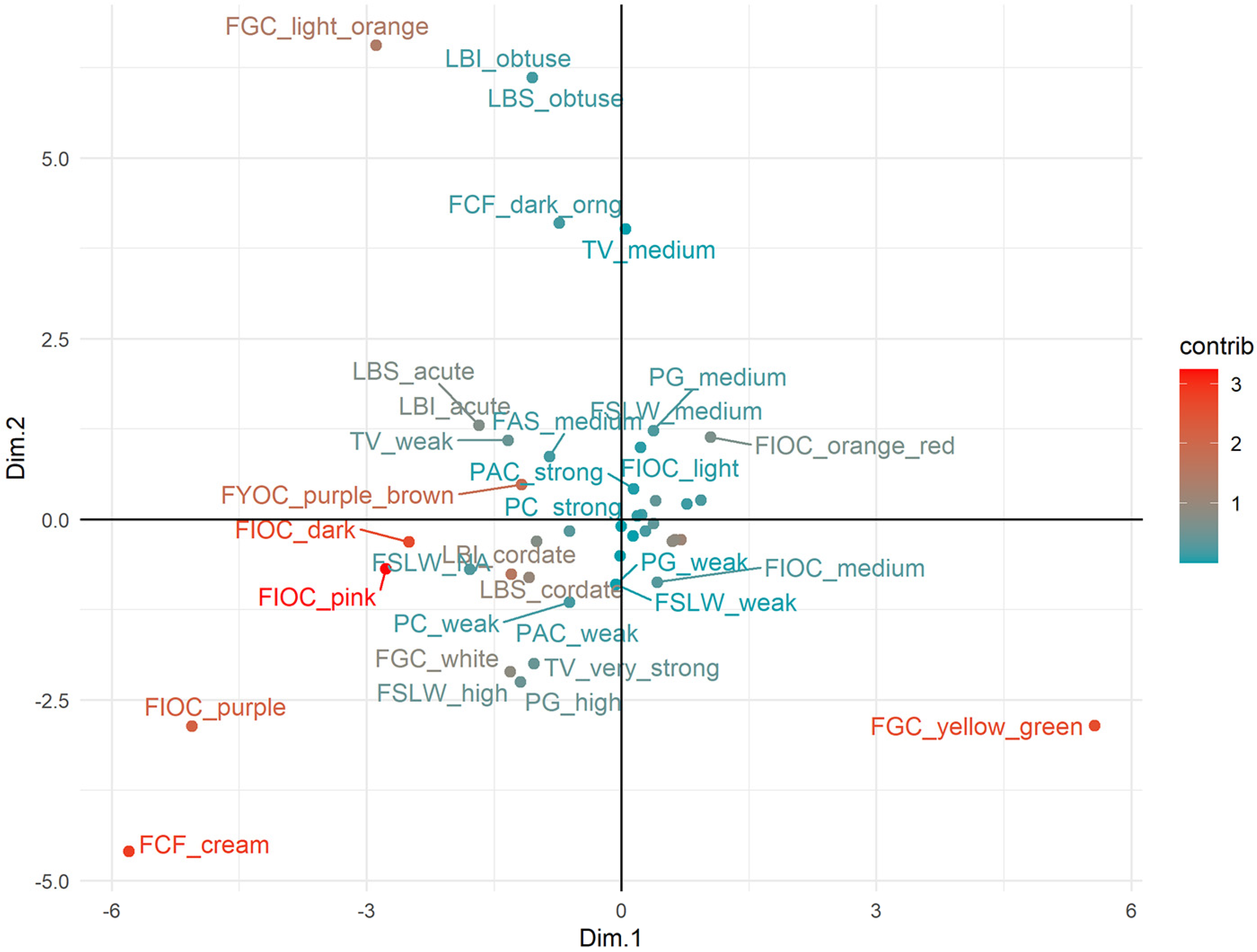
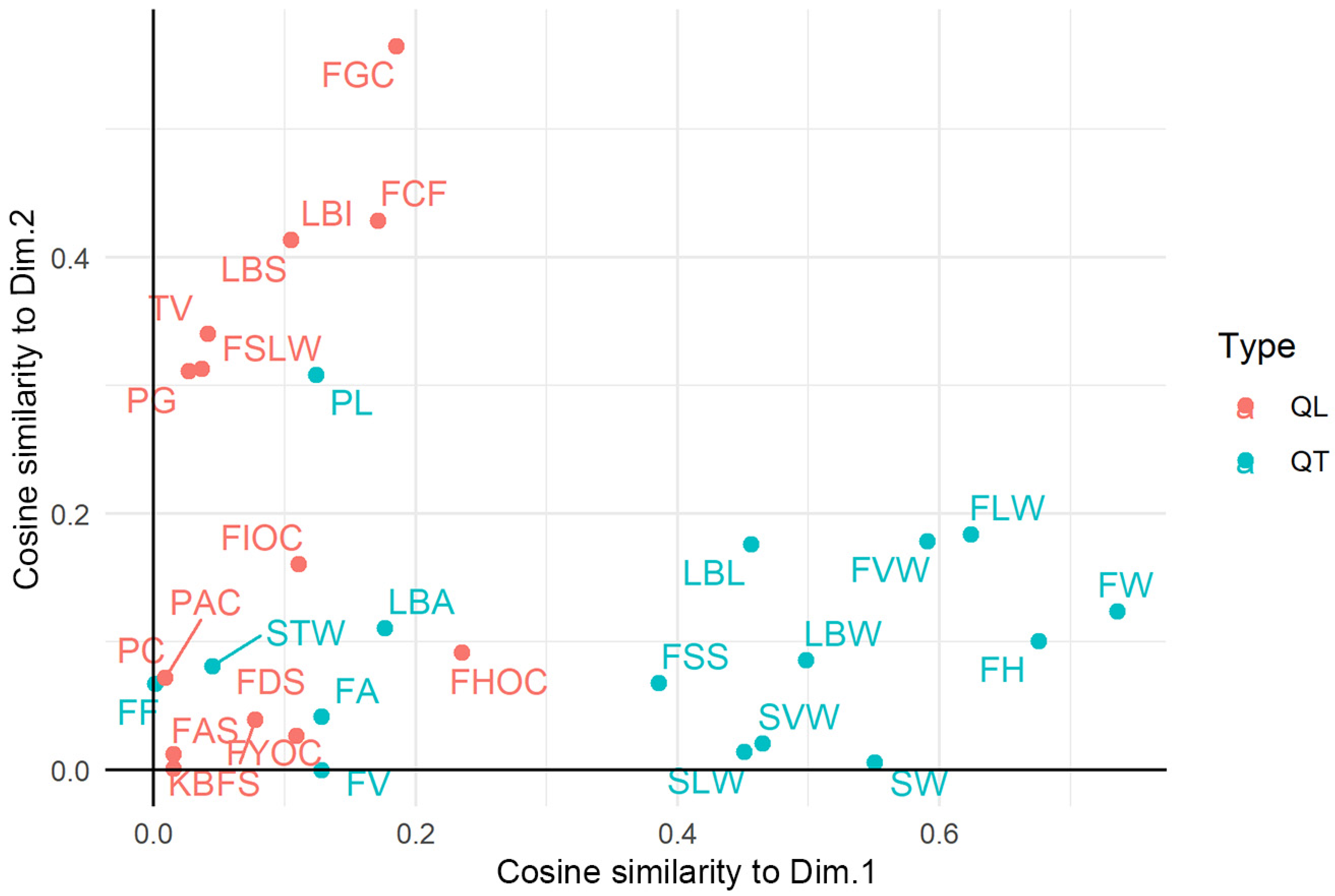
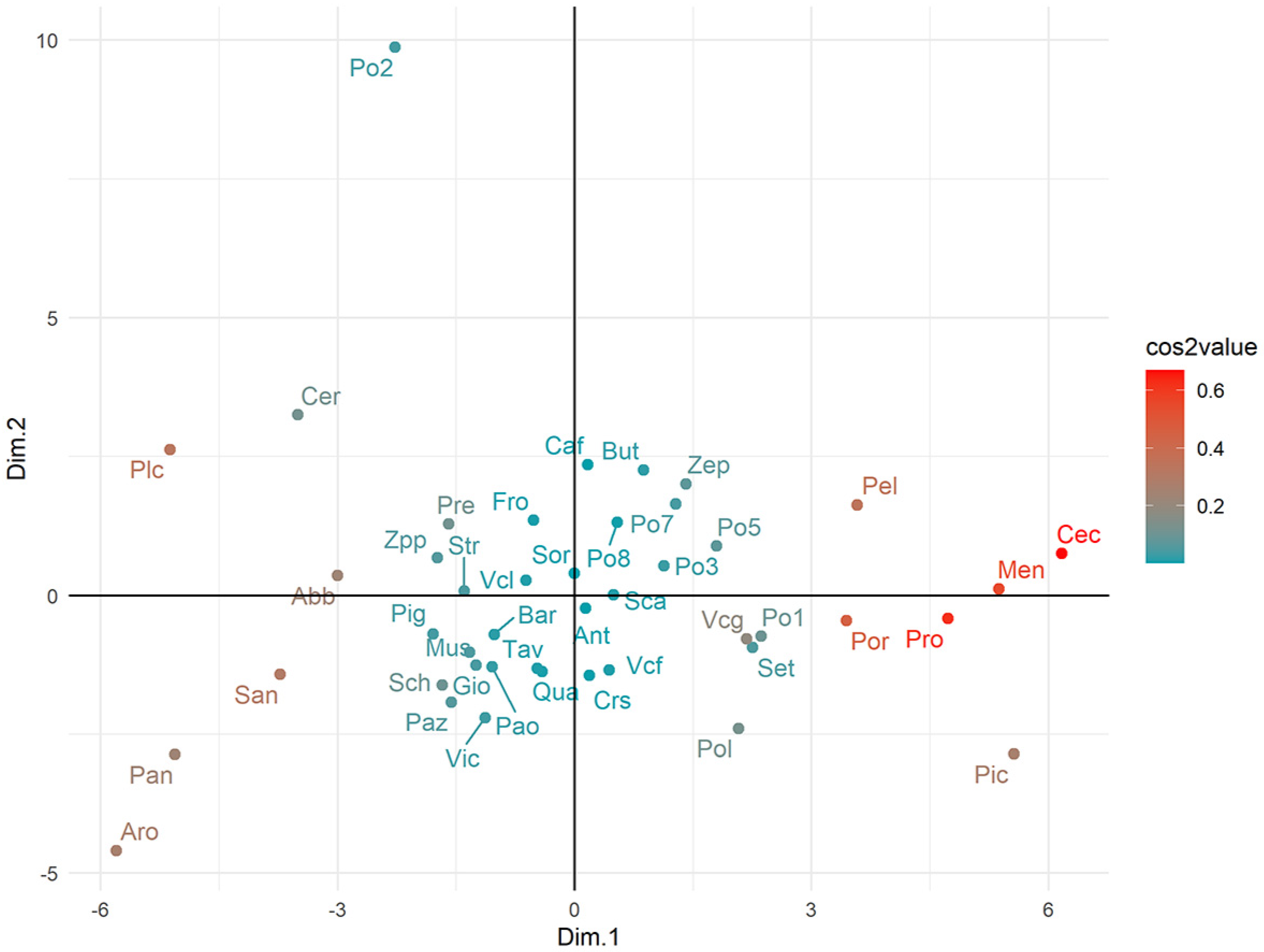
3.3. Factorial Analysis of Mixed Data
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ledbetter, C.A. Apricots. In Temperate Fruit Crop Breeding: Germplasm to Genomics; Hancock, J.F., Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 39–82. [Google Scholar]
- Jashemski, W.F. Ancient Roman gardens in Campania and Tunisia: A comparison of the evidence. J. Gard. Hist. 1996, 16, 231–243. [Google Scholar] [CrossRef]
- Guerriero, R. Albicocco. In Atlante dei Fruttiferi Autoctoni Italiani; Fideghelli, C., Ed.; MIPAF-CREA: Rome, Italy, 2016; Volume 2, pp. 477–552. [Google Scholar]
- Zhebentyayeva, T.; Ledbetter, C.; Burgos, L.; Llácer, G. Apricot. In Fruit Breeding; Springer: New York, NY, USA, 2012; pp. 415–458. [Google Scholar]
- Zeven, A.C. Landraces: A review of definitions and classifications. Euphytica 1998, 104, 127–139. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Warschefsky, E.; Klein, L.L.; Miller, A.J. Using living germplasm collections to characterize, improve, and conserve woody perennials. Crop Sci. 2019, 59, 2365–2380. [Google Scholar]
- Furman, B.; Noorani, A.; Mba, C. On-Farm Crop Diversity for Advancing Food Security and Nutrition. In Landraces-Traditional Variety and Natural Breed; IntechOpen: London, UK, 2021. [Google Scholar]
- Corrado, G.; Rao, R. Towards the genomic basis of local adaptation in landraces. Diversity 2017, 9, 51. [Google Scholar] [CrossRef] [Green Version]
- Van Wallendael, A.; Soltani, A.; Emery, N.C.; Peixoto, M.M.; Olsen, J.; Lowry, D.B. A molecular view of plant local adaptation: Incorporating stress-response networks. Annu. Rev. Plant Biol. 2019, 70, 559–583. [Google Scholar]
- Krška, B. Genetic apricot resources and their utilisation in breeding. In Breeding and Health Benefits of Fruit and Nut Crops; IntechOpen: London, UK, 2018. [Google Scholar]
- Fratianni, F.; Cozzolino, R.; d’Acierno, A.; Ombra, M.N.; Spigno, P.; Riccardi, R.; Malorni, L.; Stocchero, M.; Nazzaro, F. Biochemical Characterization of Some Varieties of Apricot Present in the Vesuvius Area, Southern Italy. Front. Nutr. 2022, 9, 4868. [Google Scholar] [CrossRef] [PubMed]
- Cirillo, C.; Basile, B.; Hernandez, G.; Pannico, A.; Giaccone, M.; Forlani, M. Influence of fruiting shoot on flowering pattern and fruit quality of Vesuvian apricot cultivars. Acta Hortic. 2010, 862, 557–564. [Google Scholar] [CrossRef]
- Li, H.; Rasheed, A.; Hickey, L.T.; He, Z. Fast-forwarding genetic gain. Trends Plant Sci. 2018, 23, 184–186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarvis, D.I.; Brown, A.H.; Cuong, P.H.; Collado-Panduro, L.; Latournerie-Moreno, L.; Gyawali, S.; Tanto, T.; Sawadogo, M.; Mar, I.; Sadiki, M. A global perspective of the richness and evenness of traditional crop-variety diversity maintained by farming communities. Proc. Natl. Acad. Sci.USA 2008, 105, 5326–5331. [Google Scholar]
- Loko, L.E.Y.; Toffa, J.; Adjatin, A.; Akpo, A.J.; Orobiyi, A.; Dansi, A. Folk taxonomy and traditional uses of common bean (Phaseolus vulgaris L.) landraces by the sociolinguistic groups in the central region of the Republic of Benin. J. Ethnobiol. Ethnomedicine 2018, 14, 1–15. [Google Scholar] [CrossRef]
- Martínez Mora, C.; Rodríguez, J.; Cenis, J.L.; Ruiz García, L. Genetic variability among local apricots (Prunus armeniaca L.) from the Southeast of Spain. Span. J. Agric. Res. 2009, 7, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.; Huang, H. SSR fingerprinting Chinese peach cultivars and landraces (Prunus persica) and analysis of their genetic relationships. Sci. Hortic. 2009, 120, 188–193. [Google Scholar] [CrossRef]
- Bourguiba, H.; Khadari, B.; Krichen, L.; Trifi-Farah, N.; Mamouni, A.; Trabelsi, S.; Audergon, J.-M. Genetic relationships between local North African apricot (Prunus armeniaca L.) germplasm and recently introduced varieties. Sci. Hortic. 2013, 152, 61–69. [Google Scholar] [CrossRef]
- Rao, R.; Bencivenni, M.; Mura, L.; Araujo-Burgos, T.; Corrado, G. Molecular characterisation of Vesuvian apricot cultivars: Implications for the certification and authentication of protected plant material. J. Hortic. Sci. Biotechnol. 2010, 85, 42–47. [Google Scholar] [CrossRef]
- Sheikh, Z.N.; Sharma, V.; Shah, R.A.; Raina, S.; Aljabri, M.; Mir, J.I.; AlKenani, N.; Hakeem, K.R. Elucidating Genetic Diversity in Apricot (Prunus armeniaca L.) Cultivated in the North-Western Himalayan Provinces of India Using SSR Markers. Plants 2021, 10, 2668. [Google Scholar] [CrossRef]
- Herrera, S.; Hormaza, J.I.; Lora, J.; Ylla, G.; Rodrigo, J. Molecular Characterization of Genetic Diversity in Apricot Cultivars: Current Situation and Future Perspectives. Agronomy 2021, 11, 1714. [Google Scholar]
- Hagen, L.; Khadari, B.; Lambert, P.; Audergon, J.-M. Genetic diversity in apricot revealed by AFLP markers: Species and cultivar comparisons. Theor. Appl. Genet. 2002, 105, 298–305. [Google Scholar] [CrossRef]
- Sanzol, J.; Herrero, M. The “effective pollination period” in fruit trees. Sci. Hortic. 2001, 90, 1–17. [Google Scholar] [CrossRef]
- Lachkar, A.; Chaar, H.; Mars, M. Reproductive behavior of Tunisian apricot cultivars as related to floral morphometric characteristics and pollen quality. Int. J. Fruit Sci. 2014, 14, 188–204. [Google Scholar] [CrossRef]
- Basile, B.; Cirillo, C.; Santin, A.; Forlani, M. Fruit quality of Vesuvian apricots harvested at different ripening stages after a cold-storage period. Acta Hortic. 2005, 682, 1443–1450. [Google Scholar] [CrossRef]
- Crisosto, C.; Crisosto, G.; Neri, F. Understanding tree fruit quality based on consumer acceptance. In Proceedings of the IV International Conference on Managing Quality in Chains-The Integrated View on Fruits and Vegetables Quality 712, Bangkok, Thailand, 26–30 June 2006; pp. 183–190. [Google Scholar]
- Gurrieri, F.; Audergon, J.-M.; Albagnac, G.; Reich, M. Soluble sugars and carboxylic acids in ripe apricot fruit as parameters for distinguishing different cultivars. Euphytica 2001, 117, 183–189. [Google Scholar] [CrossRef]
- Dray, S.; Dufour, A.-B. The ade4 package: Implementing the duality diagram for ecologists. J. Stat. Softw. 2007, 22, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Baddeley, A.; Turner, R. Spatstat: An R package for analyzing spatial point patterns. J. Stat. Softw. 2005, 12, 1–42. [Google Scholar] [CrossRef] [Green Version]
- Wei, T.; Simko, V.; Levy, M.; Xie, Y.; Jin, Y.; Zemla, J. Package ‘corrplot’. Statistician 2017, 56, 24. [Google Scholar]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Lê, S.; Josse, J.; Husson, F. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kassambara, A. Practical Guide to Principal Component Methods in R: PCA, M (CA), FAMD, MFA, HCPC, Factoextra; Sthda: Montpellier, France, 2017; Volume 2. [Google Scholar]
- Engels, J. A Guide to Effective Management of Germplasm Collections; Bioversity International: Rome, Italy, 2003. [Google Scholar]
- Pennone, F. Costituzione di cultivar di albicocco idonee alla trasformazione industriale. Agric. Ric. 1991, 119, 79–80. [Google Scholar]
- Pugliano, G.; Cirillo, A. Cultivar di albicocche vesuviane che meglio si prestano alla produzione di sciroppati. Inf. Agrar. 1975, 31, 20055–20057. [Google Scholar]
- Corrado, G.; Forlani, M.; Rao, R.; Basile, B. Diversity and Relationships among Neglected Apricot (Prunus armeniaca L.) Landraces Using Morphological Traits and SSR Markers: Implications for Agro-Biodiversity Conservation. Plants 2021, 10, 1341. [Google Scholar] [CrossRef]
- Wani, A.A.; Zargar, S.A.; Malik, A.H.; Kashtwari, M.; Nazir, M.; Khuroo, A.A.; Ahmad, F.; Dar, T.A. Assessment of variability in morphological characters of apricot germplasm of Kashmir, India. Sci. Hortic. 2017, 225, 630–637. [Google Scholar] [CrossRef]
- Salas-Salvadó, J.; Casas-Agustench, P.; Salas-Huetos, A. Cultural and historical aspects of Mediterranean nuts with emphasis on their attributed healthy and nutritional properties. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 1–6. [Google Scholar] [CrossRef]
- Corrado, G.; La Mura, M.; Ambrosino, O.; Pugliano, G.; Varricchio, P.; Rao, R. Relationships of Campanian olive cultivars: Comparative analysis of molecular and phenotypic data. Genome 2009, 52, 692–700. [Google Scholar] [CrossRef] [Green Version]
- Pop, I.F.; Vicol, A.C.; Botu, M.; Raica, P.A.; Vahdati, K.; Pamfil, D. Relationships of walnut cultivars in a germplasm collection: Comparative analysis of phenotypic and molecular data. Sci. Hortic. 2013, 153, 124–135. [Google Scholar] [CrossRef]
- Manco, R.; Basile, B.; Capuozzo, C.; Scognamiglio, P.; Forlani, M.; Rao, R.; Corrado, G. Molecular and Phenotypic Diversity of Traditional European Plum (Prunus domestica L.) Germplasm of Southern Italy. Sustainability 2019, 11, 4112. [Google Scholar] [CrossRef] [Green Version]
- Hormaza, J.; Yamane, H.; Rodrigo, J. Apricot. In Fruits and Nuts; Springer: New York, NY, USA, 2007; pp. 171–187. [Google Scholar]
- Mencarelli, F.; Bellincontro, A.; Forniti, R.; Vizovitis, K.; Botondi, R.; Valentini, M.; Sequi, P.; DiNatale, C.; Basile, B.; Romano, R. Factors affecting the apricot quality for the consumer with special attention to the use of 1-MCP and of NDT for detection of bruising. Acta Hortcult. 2006, 717, 315–320. [Google Scholar] [CrossRef]
- Cirillo, C.; Pannico, A.; Basile, B.; Rivera, C.; Giaccone, M.; Colla, G.; De Pascale, S.; Rouphael, Y. A simple and accurate allometric model to predict single leaf area of twenty-one European apricot cultivars. Eur. J. Hortic. Sci 2017, 82, 65–71. [Google Scholar] [CrossRef]
- Ruiz, D.; Egea, J. Phenotypic diversity and relationships of fruit quality traits in apricot (Prunus armeniaca L.) germplasm. Euphytica 2008, 163, 143–158. [Google Scholar] [CrossRef]
- Bae, H.; Yun, S.K.; Yoon, I.K.; Nam, E.Y.; Kwon, J.H.; Jun, J.H. Assessment of organic acid and sugar composition in apricot, plumcot, plum, and peach during fruit development. J. Appl. Bot. Food Qual. 2014, 87, 24–29. [Google Scholar]
- Zhang, C.; Xie, D.; Bai, T.; Luo, X.; Zhang, F.; Ni, Z.; Chen, Y. Diversity of a large collection of natural populations of mango (Mangifera indica Linn.) revealed by agro-morphological and quality traits. Diversity 2020, 12, 27. [Google Scholar] [CrossRef] [Green Version]
- Krima, S.B.; Slim, A.; Gélisse, S.; Kouki, H.; Nadaud, I.; Sourdille, P.; Yahyaoui, A.; M’barek, S.B.; Suffert, F.; Marcel, T.C. Life story of Tunisian durum wheat landraces revealed by their genetic and phenotypic diversity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Dwivedi, S.; Goldman, I.; Ortiz, R. Pursuing the potential of heirloom cultivars to improve adaptation, nutritional, and culinary features of food crops. Agronomy 2019, 9, 441. [Google Scholar] [CrossRef] [Green Version]
- Testa, R.; Migliore, G.; Schifani, G.; Tinebra, I.; Farina, V. Chemical–physical, sensory analyses and consumers’ quality perception of local vs. imported Loquat fruits: A sustainable development perspective. Agronomy 2020, 10, 870. [Google Scholar] [CrossRef]
- Zeven, A.C. Traditional maintenance breeding of landraces: 2. Practical and theoretical considerations on maintenance of variation of landraces by farmers and gardeners. Euphytica 2002, 123, 147–158. [Google Scholar] [CrossRef]
- Altieri, M.A.; Merrick, L. In situ conservation of crop genetic resources through maintenance of traditional farming systems. Econ. Bot. 1987, 41, 86–96. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basile, B.; Mataffo, A.; Forlani, M.; Corrado, G. Diversity in Morphometric, Pomological, and Fruit-Quality Traits of Apricot (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale. Diversity 2022, 14, 608. https://doi.org/10.3390/d14080608
Basile B, Mataffo A, Forlani M, Corrado G. Diversity in Morphometric, Pomological, and Fruit-Quality Traits of Apricot (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale. Diversity. 2022; 14(8):608. https://doi.org/10.3390/d14080608
Chicago/Turabian StyleBasile, Boris, Alessandro Mataffo, Marcello Forlani, and Giandomenico Corrado. 2022. "Diversity in Morphometric, Pomological, and Fruit-Quality Traits of Apricot (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale" Diversity 14, no. 8: 608. https://doi.org/10.3390/d14080608
APA StyleBasile, B., Mataffo, A., Forlani, M., & Corrado, G. (2022). Diversity in Morphometric, Pomological, and Fruit-Quality Traits of Apricot (Prunus armeniaca) Traditional Varieties: Implications for Landrace Differentiation at Regional Scale. Diversity, 14(8), 608. https://doi.org/10.3390/d14080608