
Gene Transfer Agent g5 Gene Reveals Bipolar and Endemic Distribution of Roseobacter Clade Members in Polar Coastal Seawater



Abstract
1. Introduction
2. Materials and Methods
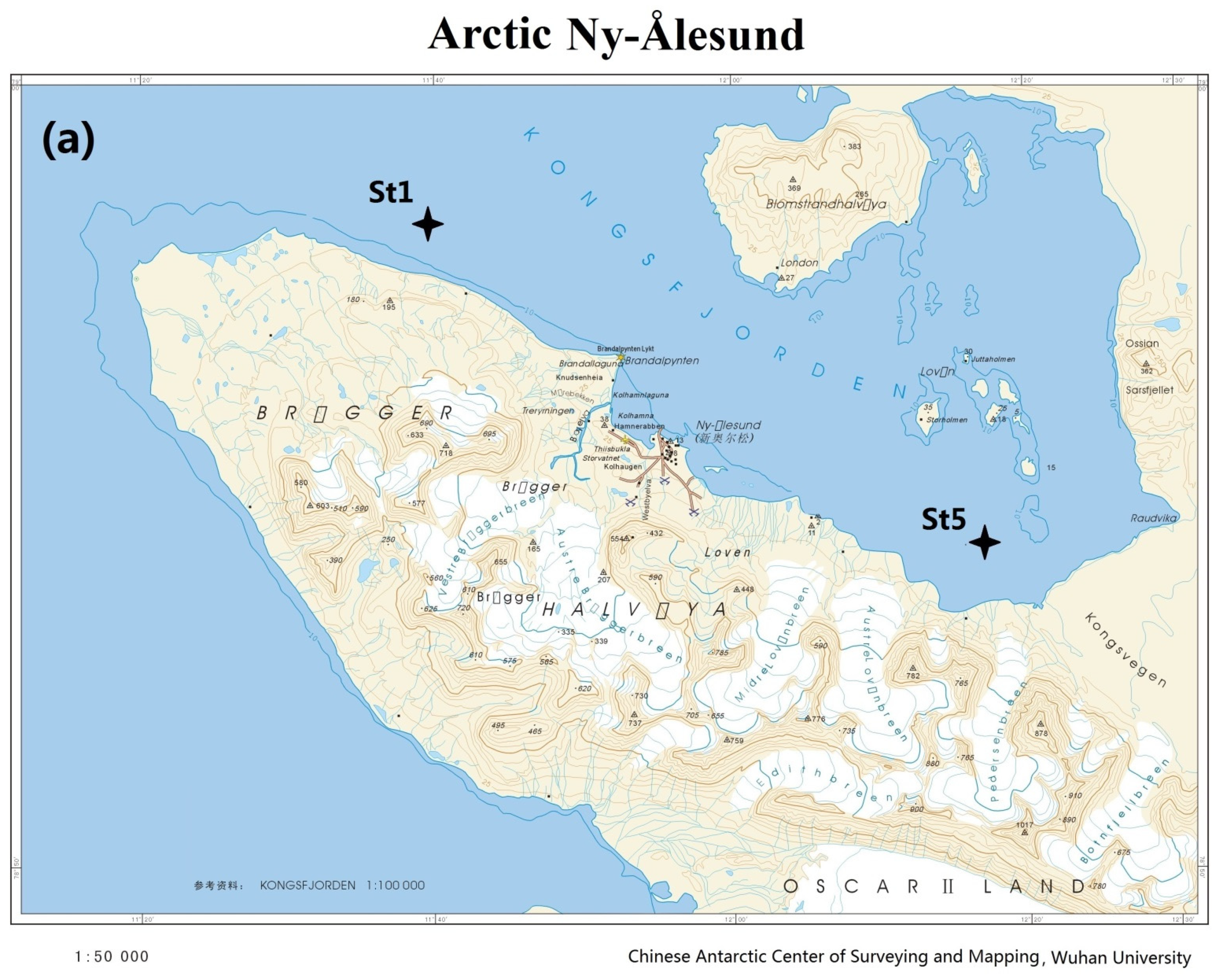
2.1. Sample Collection and DNA Extraction
2.2. Clone Library Construction and Sequencing of the GTA g5 Gene
2.3. Data Analysis
2.4. Nucleotide Sequence Accession Numbers
3. Results and Discussion
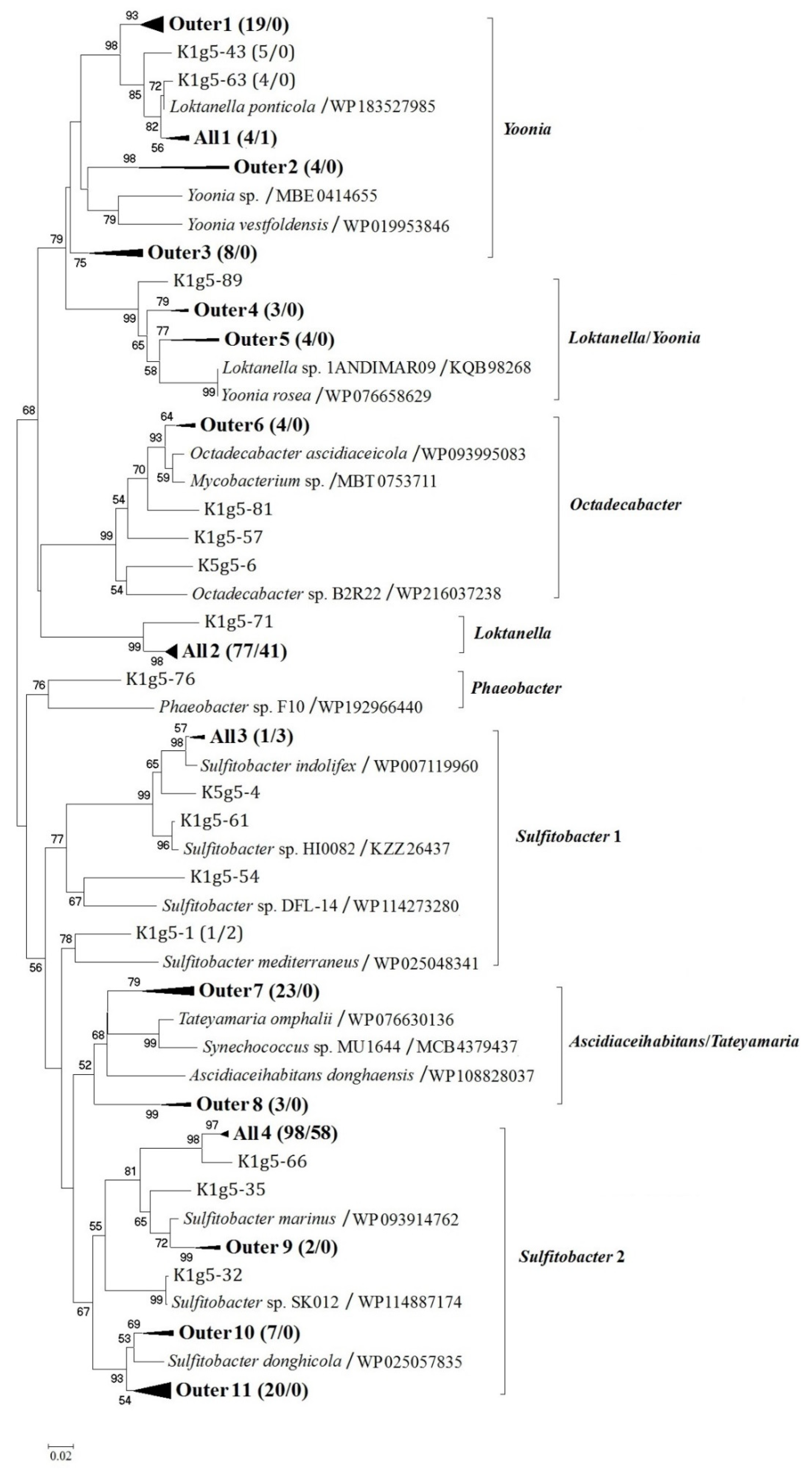
3.1. Statistical Analysis of the g5 Gene Library
3.2. Diversity and Distribution of Arctic g5 Genes
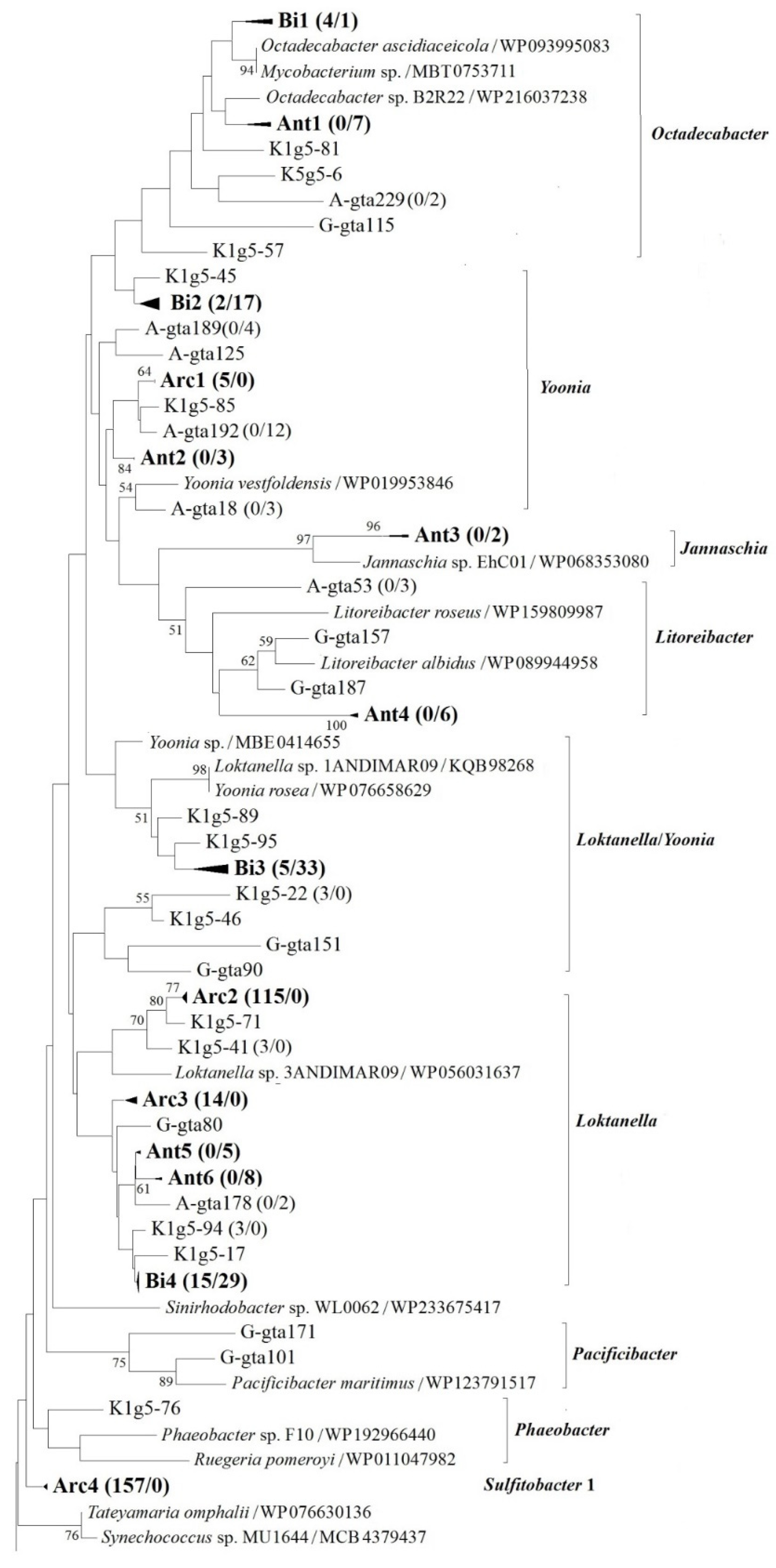
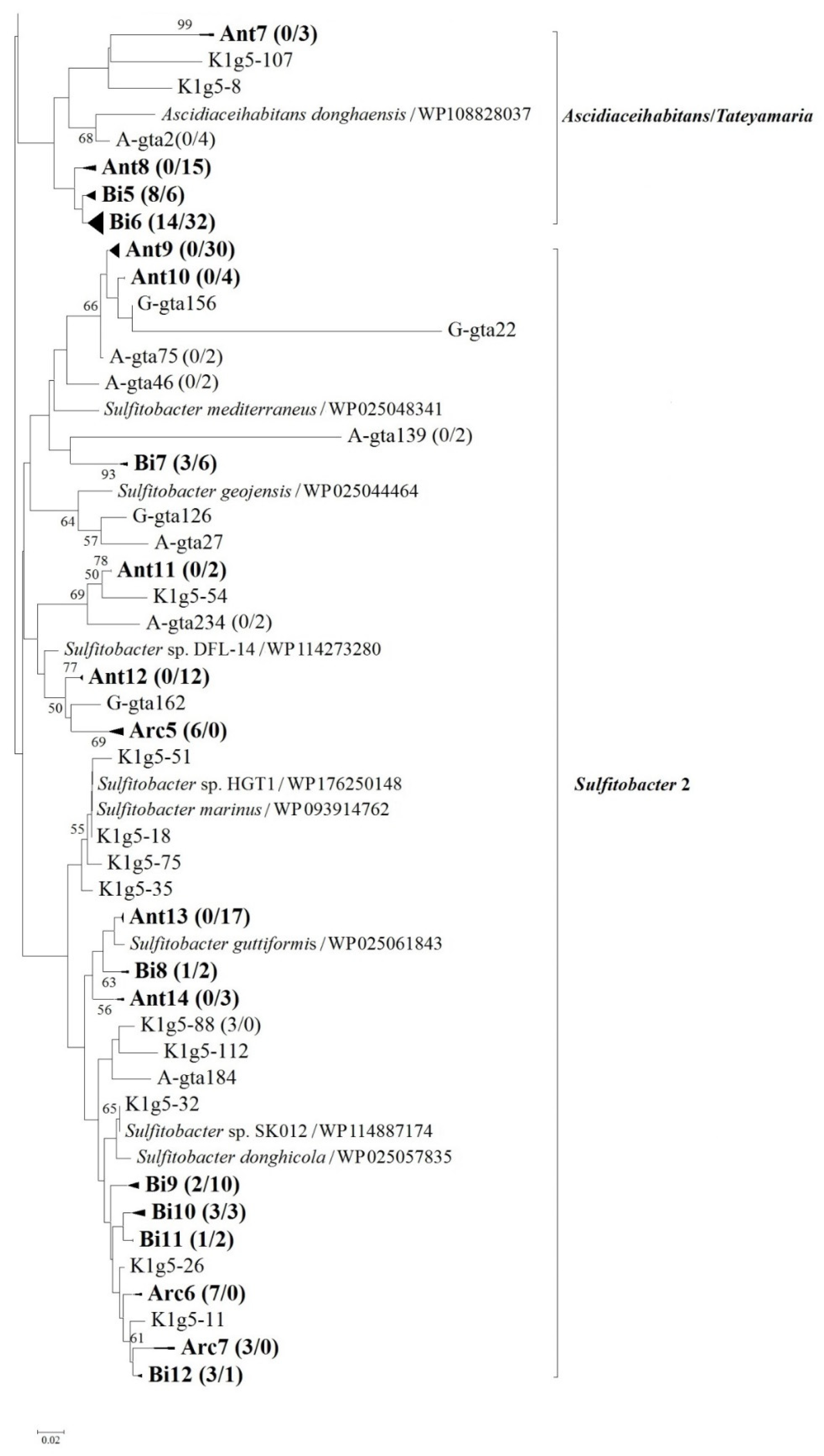
3.3. Diversity and Distribution of Antarctic g5 Genes
3.4. Relationships between Arctic and Antarctic g5 Genes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buchan, A.; Gonzaìlez, J.M.; Moran, M.A. Overview of the marine roseobacter lineage. Appl. Environ. Microbiol. 2005, 71, 5665–5677. [Google Scholar] [CrossRef]
- Garrity, G.M.; Bell, J.A.; Lilburn, T.; Family, I. Rhodobacteraceae fam. nov. In Bergey’s Manual of Systematic Bacteriology, 2nd ed.; Volume 2 (The Proteobacteria), Part C (The Alpha-, Beta-, Delta-, and Epsilonproteobacteria); Brenner, D.J., Krieg, N.R., Staley, J.T., Garrity, G.M., Eds.; Springer: New York, NY, USA, 2005; p. 161. [Google Scholar]
- Liang, K.Y.H.; Orata, F.D.; Boucher, Y.F.; Case, R.J. Roseobacters in a sea of poly- and paraphyly: Whole genome-based taxonomy of the family Rhodobacteraceae and the proposal for the split of the“Roseobacter clade” into a novel family, Roseobacteraceae fam. nov. Front. Microbiol. 2021, 12, 683109. [Google Scholar] [CrossRef]
- Moran, M.A.; Belas, R.; Schell, M.A.; Gonzaìlez, J.M.; Sun, F.; Sun, S.; Binder, B.J.; Edmonds, J.; Ye, W.; Orcutt, B.; et al. Ecological genomics of marine roseobacters. Appl. Environ. Microbiol. 2007, 73, 4559–4569. [Google Scholar] [CrossRef]
- Luo, H.; Moran, M.A. Evolutionary ecology of the marine Roseobacter clade. Microbiol. Mol. Biol. Rev. 2014, 78, 573–587. [Google Scholar] [CrossRef]
- Tolli, J.D.; Sievert, S.M.; Taylor, C.D. Unexpected diversity of bacteria capable of carbon monoxide oxidation in a coastal marine environment, and contribution of the Roseobacter-associated clade to total CO oxidation. Appl. Environ. Microbiol. 2006, 72, 1966–1973. [Google Scholar] [CrossRef]
- Imhoff, J.F.; Rahn, T.; Künzel, S.; Neulinger, S.C. Phylogeny of anoxygenic photosynthesis based on sequences of photosynthetic reaction center proteins and a key enzyme in bacteriochlorophyll biosynthesis, the chlorophyllide reductase. Microorganisms 2019, 7, 576. [Google Scholar] [CrossRef]
- Cude, W.N.; Buchan, A. Acyl-homoserine lactone-based quorum sensing in the roseobacter clade: Complex cell-to-cell communication controls multiple physiologies. Front. Microbiol. 2013, 4, 336. [Google Scholar] [CrossRef]
- Moran, M.A.; González, J.M.; Kiene, R.P. Linking a bacterial taxon to sulfur cycling in the sea: Studies of the marine roseobacter group. Geomicrobiol. J. 2003, 20, 375–388. [Google Scholar] [CrossRef]
- Reisch, C.R.; Moran, M.A.; Whitman, W.B. Bacterial catabolism of dimethylsulfoniopropionate (DMSP). Front. Microbiol. 2011, 2, 172. [Google Scholar] [CrossRef]
- Brinkmeyer, R.; Knittel, K.; Jürgens, J.; Weyland, H.; Amann, R.; Helmke, E. Diversity and structure of bacterial communities in Arctic versus Antarctic pack ice. Appl. Environ. Microbiol. 2003, 69, 6610–6619. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Yu, Y.; Qiao, Z.Y.; Jin, H.Y.; Li, H.R. Diversity of bacterioplankton in coastal seawaters of Fildes Peninsula, King George Island, Antarctica. Arch. Microbiol. 2014, 196, 137–147. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Luo, W.; Li, H.R.; Yu, Y. High diversity of planktonic prokaryotes in Arctic Kongsfjorden seawaters in summer 2015. Polar Biol. 2021, 44, 195–208. [Google Scholar] [CrossRef]
- Esterman, E.S.; Wolf, Y.I.; Kogay, R.; Koonin, E.V.; Zhaxybayeva, O. Evolution DNA packaging in gene transfer agents. Virus Evol. 2021, 7, veab015. [Google Scholar] [CrossRef]
- Hynes, A.P.; Mercer, R.G.; Watton, D.E.; Buckley, C.B.; Lang, A.S. DNA packaging bias and differential expression of gene transfer agent genes within a population during production and release of the Rhodobacter capsulatus gene transfer agent, RcGTA. Mol. Microbiol. 2012, 85, 314–325. [Google Scholar] [CrossRef]
- Brinkhoff, T.; Giebel, H.A.; Simon, M. Diversity, ecology, and genomics of the Roseobacter clade: A short overview. Arch. Microbiol. 2008, 189, 531–539. [Google Scholar] [CrossRef]
- Lang, A.S.; Beatty, J.T. Importance of widespread gene transfer agent genes in alpha-proteobacteria. Trends Microbiol. 2007, 15, 54–62. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, K.; Budinoff, C.; Buchan, A.; Lang, A.; Jiao, N.; Chen, F. Gene transfer agent (GTA) genes reveal diverse and dynamic Roseobacter and Rhodobacter populations in the Chesapeake Bay. ISME J. 2009, 3, 364–373. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Dong, P.Y.; Qiao, Z.Y.; Zheng, T.L. Diversity of the aerobic anoxygenic phototrophy gene pufM in Arctic and Antarctic coastal seawaters. Acta Oceanol. Sin. 2016, 35, 68–77. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Qiao, Z.Y. Diversity of dimethylsulfoniopropionate degradation genes reveals the significance of marine Roseobacter clade in sulfur metabolism in coastal areas of Antarctic Maxwell Bay. Curr. Microbiol. 2019, 76, 967–974. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Zhang, F.; He, J.F.; Lee, S.H.; Qiao, Z.Y.; Yu, Y.; Li, H.R. Bacterioplankton community structure in the Arctic waters as revealed by pyrosequencing of 16S rRNA genes. Antonie Leeuwenhoek 2013, 103, 1309–1319. [Google Scholar] [CrossRef]
- Bosshard, P.P.; Santini, Y.; Grüter, D.G.; Stettler, R.; Bachofen, R. Bacterial diversity and community composition in the chemocline of the meromictic alpine Lake Cadagno as revealed by 16S rRNA gene analysis. FEMS Microbiol. Ecol. 2000, 31, 173–182. [Google Scholar] [CrossRef]
- Bano, N.; Hollibaugh, J.T. Diversity and distribution of DNA sequences with affinity to ammonia-oxidizing bacteria of the β subdivision of the class Proteobacteria in the Arctic Ocean. Appl. Environ. Microbiol. 2000, 66, 1960–1969. [Google Scholar] [CrossRef]
- Zeng, Y.; Zheng, T.; Li, H. Community composition of the marine bacterioplankton in Kongsfjorden (Spitsbergen) as revealed by 16S rRNA gene analysis. Polar Biol. 2009, 32, 1447–1460. [Google Scholar] [CrossRef][Green Version]
- Huber, T.; Faulkner, G.; Hugenholtz, P. Bellerophon: A program to detect chimeric sequences in multiple sequence alignments. Bioinformatics 2004, 20, 2317–2319. [Google Scholar] [CrossRef]
- Kurahashi, M.; Yokota, A. Tateyamaria omphalii gen. nov., sp. nov., an α-Proteobacterium isolated from a top shell Omphalius pfeifferi pfeifferi. Syst. Appl. Microbiol. 2007, 30, 371–375. [Google Scholar] [CrossRef]
- Kim, Y.O.; Park, S.; Nam, B.H.; Lee, C.; Park, J.M.; Kim, D.G.; Yoon, J.H. Ascidiaceihabitans donghaensis gen. nov., sp. nov., isolated from the golden sea squirt Halocynthia aurantium. Int. J. Syst. Evol. Microbiol. 2014, 64, 3970–3975. [Google Scholar] [CrossRef]
- Sinha, R.K.; Krishnan, K.P.; Kerkar, S.; Thresyamma, D.D. Influence of glacial melt and Atlantic water on bacterioplankton community of Kongsfjorden, an Arctic fjord. Ecol. Indic. 2017, 82, 143–151. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Zhang, Y.H.; Li, H.R.; Luo, W. Complete genome of Sulfitobacter sp. BSw21498 isolated from seawater of Arctic Kongsfjorden. Mar. Genom. 2020, 53, 100769. [Google Scholar] [CrossRef]
- Barak-Gavish, N.; Frada, M.J.; Ku, C.; Lee, P.A.; DiTullio, G.R.; Malitsky, S.; Aharoni, A.; Green, S.J.; Rotkopf, R.; Kartvelishvily, E.; et al. Bacterial virulence against an oceanic bloom-forming phytoplankter is mediated by algal DMSP. Sci. Adv. 2018, 4, eaau5716. [Google Scholar] [CrossRef]
- Amin, S.A.; Hmelo, L.R.; van Tol, H.M.; Durham, B.P.; Carlson, L.T.; Heal, K.R.; Morales, R.L.; Berthiaume, C.T.; Parker, M.S.; Djunaedi, B.; et al. Interaction and signalling between a cosmopolitan phytoplankton and associated bacteria. Nature 2015, 522, 98–101. [Google Scholar] [CrossRef]
- Zeng, Y.X.; Qiao, Z.Y.; Yu, Y.; Li, H.R.; Luo, W. Diversity of bacterial dimethylsulfoniopropionate degradation genes in surface seawater of Arctic Kongsfjorden. Sci. Rep. 2016, 6, 33031. [Google Scholar] [CrossRef] [PubMed]
- Wirth, J.S.; Whitman, W.B. Phylogenomic analyses of a clade within the roseobacter group suggest taxonomic reassignments of species of the genera Aestuariivita, Citreicella, Loktanella, Nautella, Pelagibaca, Ruegeria, Thalassobius, Thiobacimonas and Tropicibacter, and the proposal of six novel genera. Int. J. Syst. Evol. Microbiol. 2018, 68, 2393–2411. [Google Scholar]
- Chen, Y.H.; Kuo, J.; Sung, P.J.; Chang, Y.C.; Lu, M.C.; Wong, T.Y.; Liu, J.K.; Weng, C.F.; Twan, W.H.; Kuo, F.W. Isolation of marine bacteria with antimicrobial activities from cultured and field-collected soft corals. World J. Microbiol. Biotechnol. 2012, 28, 3269–3279. [Google Scholar] [CrossRef] [PubMed]
- Villasante, A.; Catalán, N.; Rojas, R.; Lohrmann, K.B.; Romero, J. Microbiota of the digestive gland of red abalone (Haliotis rufescens) is affected by withering syndrome. Microorganisms 2020, 8, 1411. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, M.; Wang, B.; Jiang, K.; Wang, M.; Wang, L. Response of the Litopenaeus vananmei intestinal bacteria and antioxidant system to rearing density and exposure to Vibrio paraheamolyticus E1. J. Invertebr. Pathol. 2020, 170, 107326. [Google Scholar] [CrossRef] [PubMed]
- Zheng, S.; Zhou, S.; Lukwambe, B.; Nicholaus, R.; Yang, W.; Zheng, Z. Bacterioplankton community assembly in migratory fish habitat: A case study of the southern East China Sea. Environ. Sci. Pollut. Res. Int. 2022, 29, 33725–33736. [Google Scholar] [CrossRef]
- Ivanova, E.P.; Pham, D.K.; Wright, J.P.; Nicolau, D.V. Detection of coccoid forms of Sulfitobacter mediterraneus using atomic force microscopy. FEMS Microbiol Lett. 2002, 214, 177–181. [Google Scholar] [CrossRef][Green Version]
- Ivanova, E.P.; Mitik-Dineva, N.; Wang, J.; Pham, D.K.; Wright, J.P.; Nicolau, D.V.; Mocanasu, R.C.; Crawford, R.J. Staleya guttiformis attachment on poly(tert-butylmethacrylate) polymeric surfaces. Micron 2008, 39, 1197–1204. [Google Scholar] [CrossRef]
- Labrenz, M.; Tindall, B.J.; Lawson, P.A.; Collins, M.D.; Schumann, P.; Hirsch, P. Staleya guttiformis gen. nov., sp. nov. and Sulfitobacter brevis sp. nov., alpha-3-Proteobacteria from hypersaline, heliothermal and meromictic antarctic Ekho Lake. Int. J. Syst. Evol. Microbiol. 2000, 50, 303–313. [Google Scholar] [CrossRef]
- Van Trappen, S.; Mergaert, J.; Swings, J. Loktanella salsilacus gen. nov., sp. nov., Loktanella fryxellensis sp. nov. and Loktanella vestfoldensis sp. nov., new members of the Rhodobacter group, isolated from microbial mats in Antarctic lakes. Int. J. Syst. Evol. Microbiol. 2004, 54, 1263–1269. [Google Scholar] [CrossRef]
- Louca, S. The rates of global bacterial and archaeal dispersal. ISME J. 2022, 16, 159–167. [Google Scholar] [CrossRef]
- Staley, J.T.; Gosink, J.J. Poles apart: Biodiversity and biogeography of sea ice bacteria. Ann. Rev. Microbiol. 1999, 53, 198–215. [Google Scholar] [CrossRef]
- Jung, Y.T.; Park, S.; Park, J.M.; Yoon, J.H. Loktanella ponticola sp. nov., isolated from seawater. Int. J. Syst. Evol. Microbiol. 2014, 64, 3717–3723. [Google Scholar] [CrossRef]
- Sun, F.L.; Wang, Y.S.; Wu, M.L.; Jiang, Z.Y.; Sun, C.C.; Cheng, H. Genetic diversity of bacterial communities and gene transfer agents in Northern South China Sea. PLoS ONE 2014, 9, e111892. [Google Scholar] [CrossRef]
- Dang, H.; Li, T.; Chen, M.; Huang, G. Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 2008, 74, 52–60. [Google Scholar] [CrossRef]
- Roux, S.; Brum, J.R.; Dutilh, B.E.; Sunagawa, S.; Duhaime, M.B.; Loy, A.; Poulos, B.T.; Solonenko, N.; Lara, E.; Poulain, J.; et al. Ecogenomics and potential biogeochemical impacts of globally abundant ocean viruses. Nature 2016, 537, 689–693. [Google Scholar] [CrossRef]
- Bárdy, P.; Füzik, T.; Hrebík, D.; Pantůček, R.; Thomas Beatty, J.; Plevka, P. Structure and mechanism of DNA delivery of a gene transfer agent. Nat. Commun. 2020, 11, 3034. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Value of Arctic Kongsfjorden | Value of Antarctic Maxwell Bay | ||
|---|---|---|---|---|
| Sampling station | St1 | St5 | A5 | G5 |
| Sampling site | 78°59′17″ N, 11°39′34″ E | 78°54′20″ N, 12°17′34″ E | 62°12′42″ S, 58°54′41″ W | 62°13′37″ S, 58°56′37″ W |
| Number of clones sequenced | 297 | 107 | 157 | 154 |
| Number of OTUs | 112 | 28 | 89 | 79 |
| Coverage (%) | 75.3 | 90.7 | 68.8 | 68.8 |
| Shannon–wiener index | 3.984 | 2.752 | 4.329 | 4.107 |
| Simpson index | 0.042 | 0.109 | 0.016 | 0.022 |
| Evenness | 0.844 | 0.826 | 0.964 | 0.940 |
| Species richness (Chao1) | 289 | 35 | 141 | 161 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zeng, Y.-X.; Li, H.-R.; Luo, W. Gene Transfer Agent g5 Gene Reveals Bipolar and Endemic Distribution of Roseobacter Clade Members in Polar Coastal Seawater. Diversity 2022, 14, 392. https://doi.org/10.3390/d14050392
Zeng Y-X, Li H-R, Luo W. Gene Transfer Agent g5 Gene Reveals Bipolar and Endemic Distribution of Roseobacter Clade Members in Polar Coastal Seawater. Diversity. 2022; 14(5):392. https://doi.org/10.3390/d14050392
Chicago/Turabian StyleZeng, Yin-Xin, Hui-Rong Li, and Wei Luo. 2022. "Gene Transfer Agent g5 Gene Reveals Bipolar and Endemic Distribution of Roseobacter Clade Members in Polar Coastal Seawater" Diversity 14, no. 5: 392. https://doi.org/10.3390/d14050392
APA StyleZeng, Y.-X., Li, H.-R., & Luo, W. (2022). Gene Transfer Agent g5 Gene Reveals Bipolar and Endemic Distribution of Roseobacter Clade Members in Polar Coastal Seawater. Diversity, 14(5), 392. https://doi.org/10.3390/d14050392