
Integrative Redescription of the Minibiotus intermedius (Plate, 1888)—The Type Species of the Genus Minibiotus R.O. Schuster, 1980

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Microscopy and Imaging
2.3. Morphometrics and Morphological Nomenclature
2.4. Genotyping
2.5. Comparative Molecular Analysis
3. Results
3.1. Taxonomic Account
3.1.1. Short Diagnosis
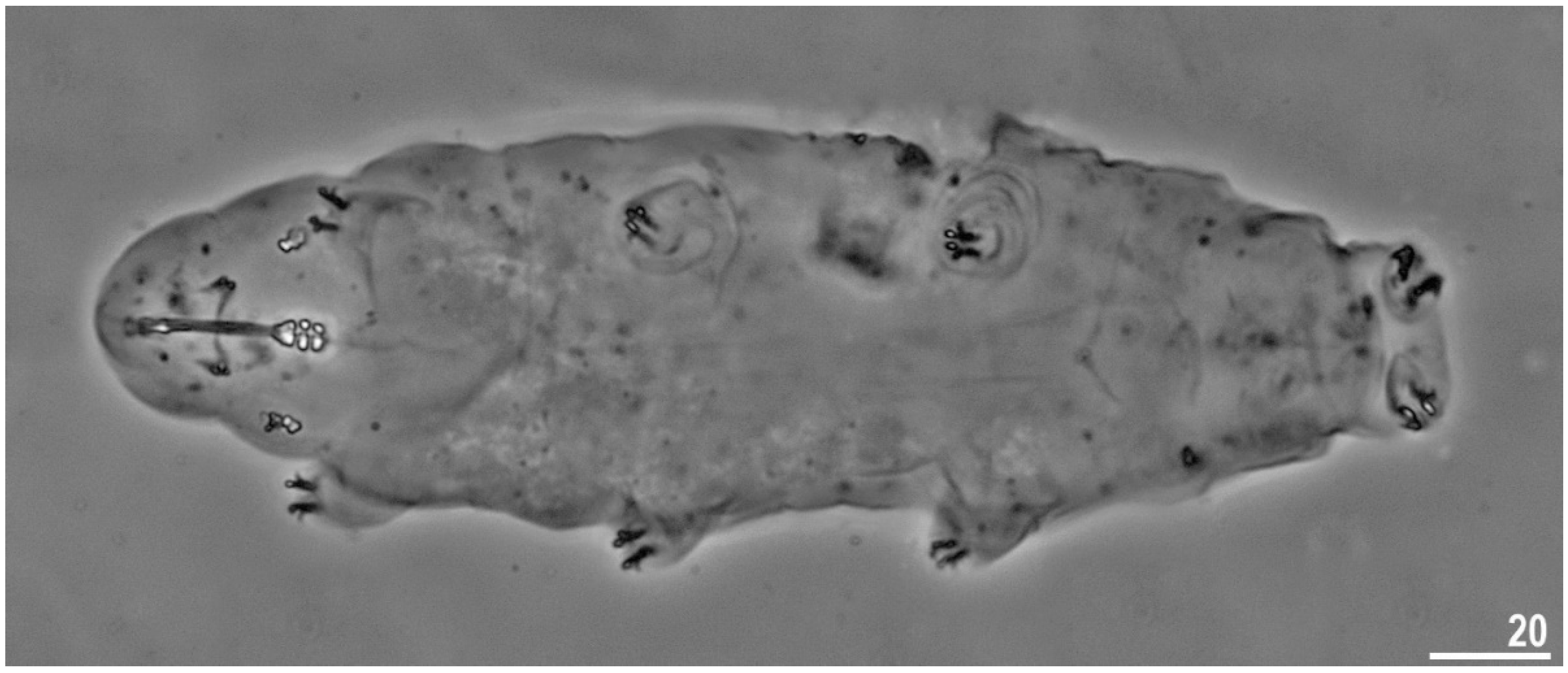
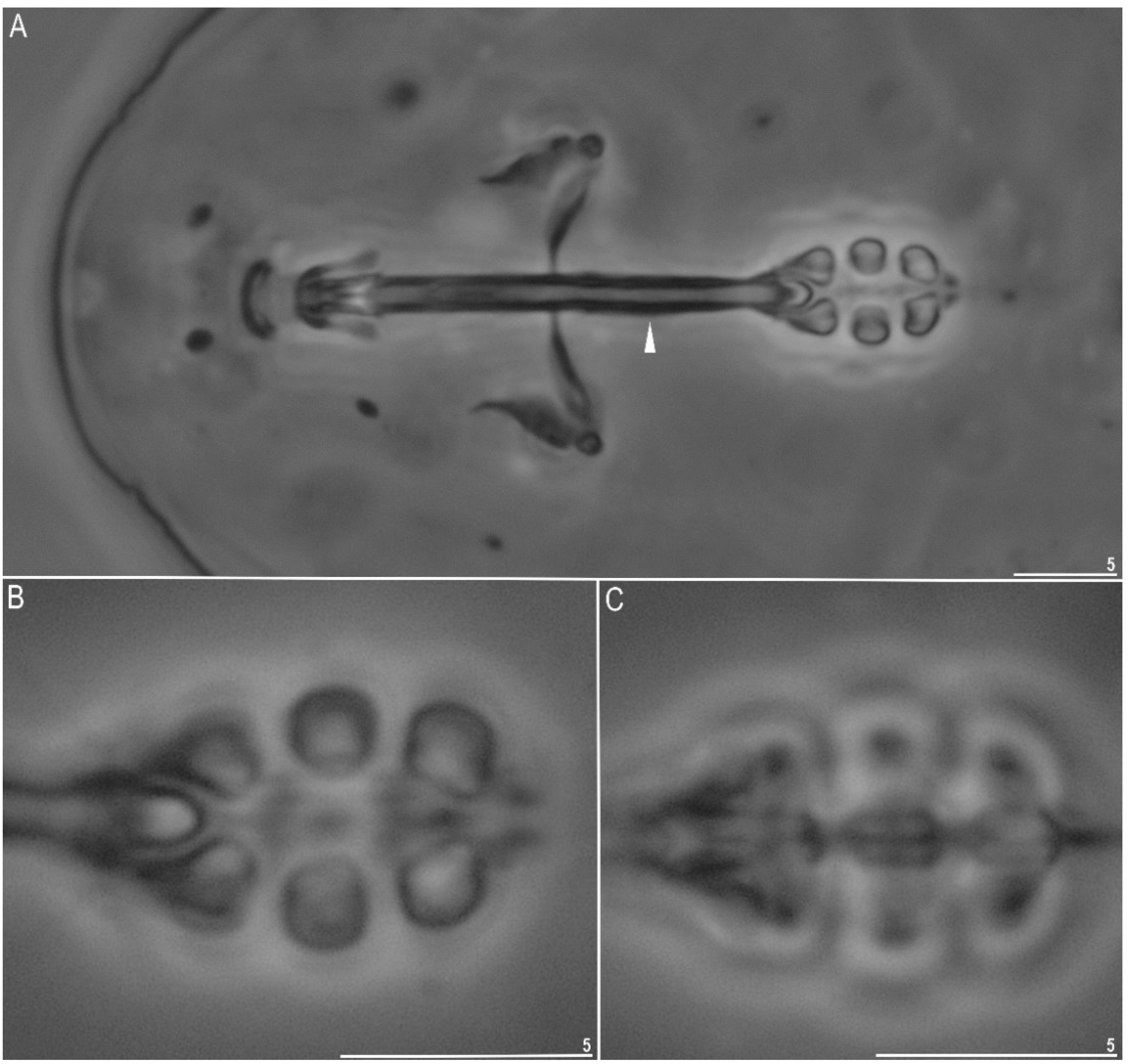
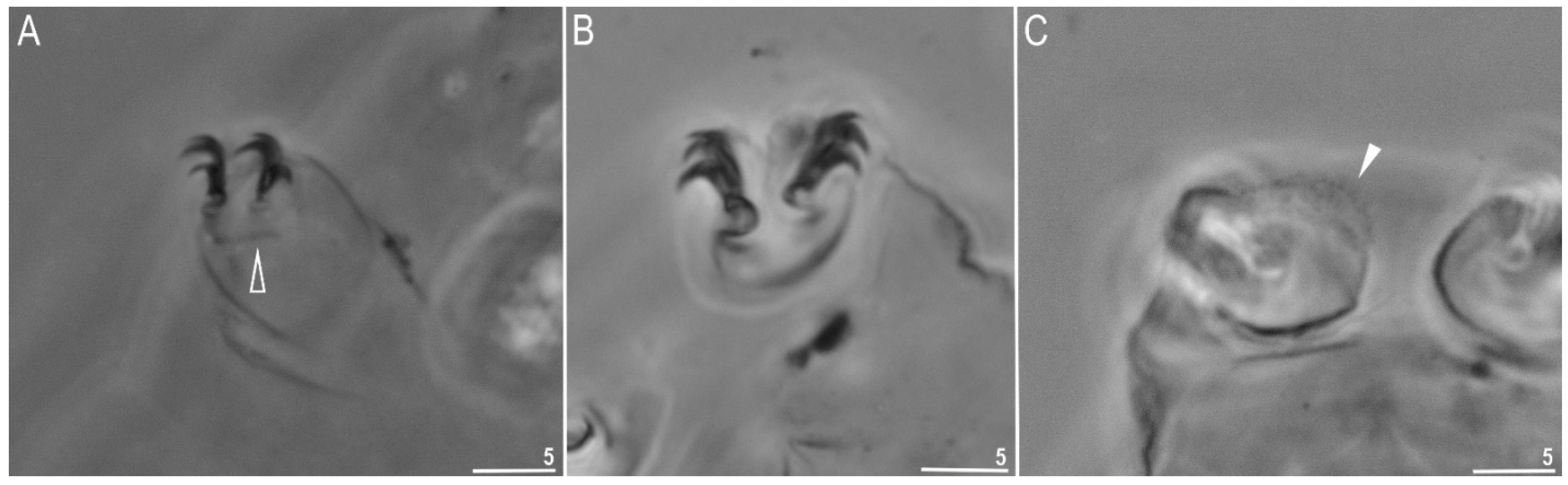
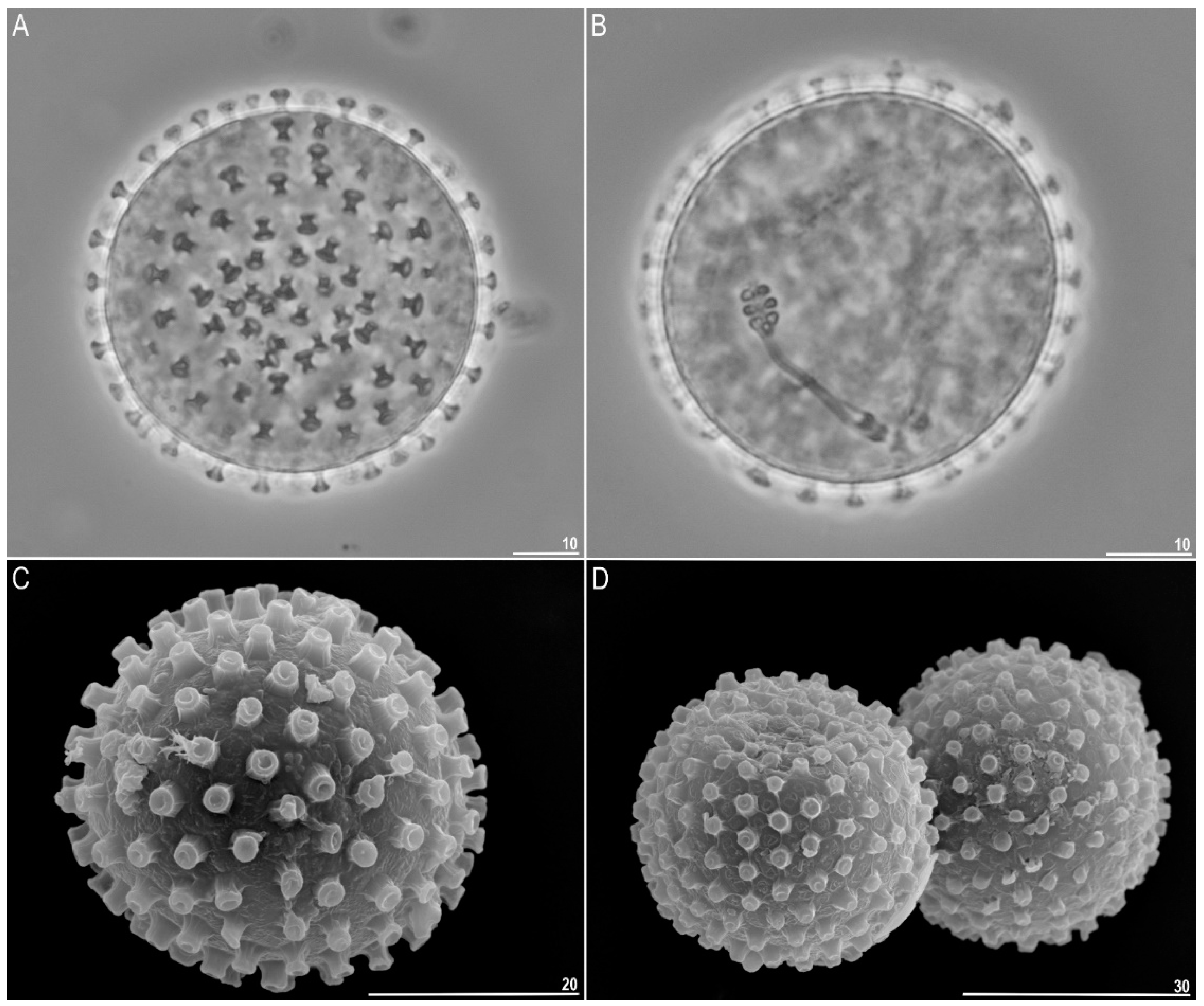
3.1.2. Description of the Neotypic Population
- COI—GenBank: ON005160, 634 bp long;
- 18S rRNA—GenBank: ON005188-89, 1182 bp long;
- 28S rRNA—GenBank: ON005193-95, 753 bp long.
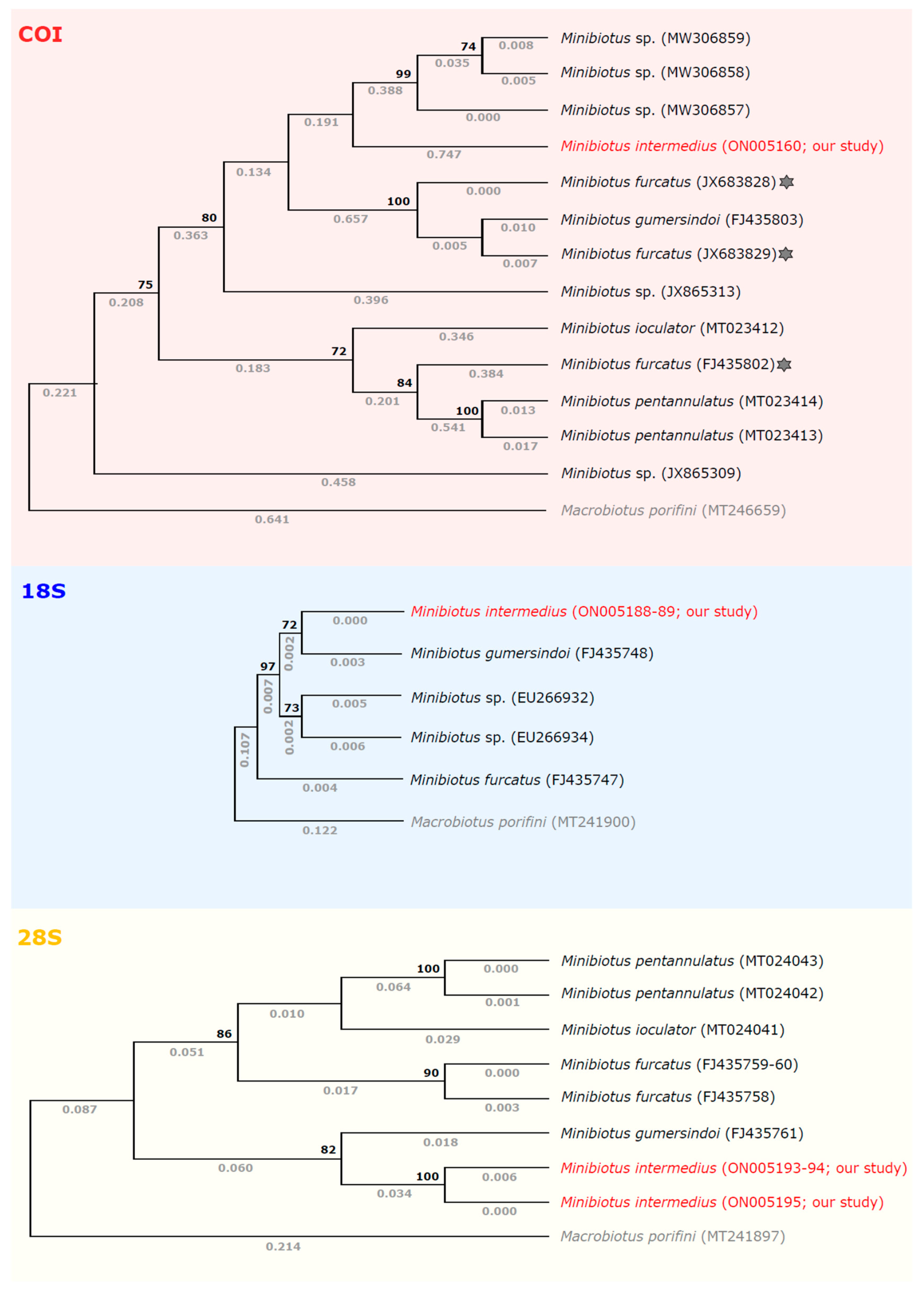
3.2. Comparisons with Other Genetic Sequences of Minibiotus Taxa
- (a)
- (b)
- (c)
3.3. Morphological Differential Diagnosis
- M. continuus by: the presence of eyes, a different macroplacoid length sequence (2 ≤ 3 < 1 in M. intermedius vs. 1 = 2 = 3 in M. continuus), and a higher number of processes on the egg circumference (28–32 in M. intermedius vs. 21–22 in M. continuus).
- M. floriparus by: a different macroplacoid length sequence (2 ≤ 3 < 1 in M. intermedius vs. 2 < 1 = 3 in M. floriparus), a lower pt of the stylet support insertion point (pt: 53.8–56.3 in M. intermedius vs. pt: ca. 64.4 in M. floriparus), the absence of granulation on legs I–III, the lack of pores on the distal tops of egg processes, larger eggs (egg bare diameter: 46.3–54.5 μm and full diameter: 52.1–61.4 μm in M. intermedius vs. ca. 62.0 μm and ca. 70.0 μm, respectively, in M. floriparus), lower egg processes (3.0–3.9 μm in M. intermedius vs. 5.5–6.0 μm in M. floriparus), narrower tops of processes (2.4–3.5 μm in M. intermedius vs. 6.0–7.0 μm in M. floriparus), and a higher number of processes on the egg circumference (28–32 in M. intermedius vs. 20–22 in M. floriparus).
- M. taiti by: lower pt of stylet support insertion point (pt: 53.8–56.3 in M. intermedius vs. pt: ca. 60.3 in M. taiti), the absence of granulation on legs I–III, and the lack of rings of small circles around the central pore on the top of egg processes.
3.4. Establishing of the New Neotype and Neoparatypes of M. intermedius
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, D.R.; Guidetti, R.; Rebecchi, L.; Kaczmarek, Ł.; McInnes, S.J. Phylum Tardigrada. In Thorp and Covich’s Freswater Invertebrates. Keys to Neotropical and Antarctic Fauna; Damborenea, C., Rogers, D.C., Thorp, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; Volume 5, pp. 505–522. [Google Scholar]
- Guidetti, R.G.; Bertolani, R.B. Tardigrade taxonomy: An updated check list of the taxa and a list of characters for their identification. Zootaxa 2005, 845, 1. [Google Scholar] [CrossRef]
- Degma, P.; Guidetti, R. Notes to the current checklist of Tardigrada. Zootaxa 2007, 1579, 41–53. [Google Scholar] [CrossRef]
- Vicente, F.; Bertolani, R. Considerations on the taxonomy of the Phylum Tardigrada. Zootaxa 2013, 3626, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Degma, P.; Bertolani, R.; Guidetti, R. Actual Checklist of Tardigrada Species. (2009–2021, 40th Edition: 19 July 2021). Available online: https://iris.unimore.it/retrieve/358743/Actual%20checklist%20of%20Tardigrada%2040th%20Edition%2019-07-21.pdf (accessed on 25 April 2022).
- Schuster, R.O.; Nelson, D.R.; Grigarick, A.A.; Christenberry, D. Systematic criteria of Eutardigrada. Trans. Am. Microsc. Soc. 1980, 99, 284–303. [Google Scholar] [CrossRef]
- Pilato, G. The systematics of Eutardigrada. J. Zool. Syst. Evol. Res. 1982, 20, 271–284. [Google Scholar] [CrossRef]
- Ramazzotti, G.; Maucci, W. Il Phylum Tardigrada. Mem. Ist. Ital. Idrobiol. 1983, 41, 1–1012. [Google Scholar]
- Claxton, S.K. A revision of the genus Minibiotus (Tardigrada: Macrobiotidae) with descriptions of eleven new species from Australia. Rec. Aust. Mus. 1998, 50, 125–160. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Kristensen, R.M.; Michalczyk, Ł. An integrative description of Minibiotus ioculator sp. nov. from the Republic of South Africa with notes on Minibiotus pentannulatus Londoño et al., 2017 (Tardigrada: Macrobiotidae). Zool. Anz. A J. Comp. Zool. 2020, 286, 117–134. [Google Scholar] [CrossRef]
- Guidetti, R.; Bertolani, R.; Degma, P. New taxonomic position of several Macrobiotus species (Eutardigrada: Macrobiotidae). Zootaxa 2007, 1471, 61–68. [Google Scholar] [CrossRef]
- Ehrenberg, C.G. Beitrag zur Bestimmung des stationären mikroscopischen Lebens in bis 20,000 Fuss Alpenhöhe. Abhand. K. Akad. Wiss. 1859, 429–456. [Google Scholar]
- Guil, N.; Guidetti, R. A new species of Tardigrada (Eutardigrada: Macrobiotidae) from Iberian Peninsula and Canary Islands (Spain). Zootaxa 2005, 889, 1–11. [Google Scholar] [CrossRef]
- Londoño, R.; Daza, A.; Lisi, O.; Quiroga, S. New species of waterbear Minibiotus pentannulatus (Tardigrada: Macrobiotidae) from Colombia. Rev. Mex. Biodivers. 2017, 88, 807–814. [Google Scholar] [CrossRef]
- Guil, N.; Giribet, G. A comprehensive molecular phylogeny of tardigrades-adding genes and taxa to a poorly resolved phylum-level phylogeny. Cladistics 2012, 28, 21–49. [Google Scholar] [CrossRef] [PubMed]
- Bertolani, R.; Guidetti, R.; Marchioro, T.; Altiero, T.; Rebecchi, L.; Cesari, M. Phylogeny of Eutardigrada: New molecular data and their morphological support lead to the identification of new evolutionary lineages. Mol. Phylogenet. Evol. 2014, 76, 110–126. [Google Scholar] [CrossRef]
- Guidetti, R.; Schill, R.O.; Bertolani, R.; Dandekar, T.; Wolf, M. New molecular data for tardigrade phylogeny, with the erection of Paramacrobiotus gen. nov. J. Zool. Syst. Evol. Res. 2009, 47, 315–321. [Google Scholar] [CrossRef]
- Stec, D.; Vecchi, M.; Calhim, S.; Michalczyk, Ł. New multilocus phylogeny reorganises the family Macrobiotidae (Eutardigrada) and unveils complex morphological evolution of the Macrobiotus hufelandi group. Mol. Phylogenet. Evol. 2021, 160, 106987. [Google Scholar] [CrossRef]
- Stec, D.; Smolak, R.; Kaczmarek, Ł.; Michalczyk, Ł. An integrative description of Macrobiotus paulinae sp. nov. (Tardigrada: Eutardigrada: Macrobiotidae: Hufelandi group) from Kenya. Zootaxa 2015, 4052, 501–526. [Google Scholar] [CrossRef] [Green Version]
- Stec, D.; Morek, W. Reaching the Monophyly: Re-Evaluation of the Enigmatic Species Tenuibiotus hyperonyx (Maucci, 1983) and the Genus Tenuibiotus (Eutardigrada). Animals 2022, 12, 404. [Google Scholar] [CrossRef]
- Pilato, G.; Lisi, O.P.V. Tenuibiotus, a new genus of Macrobiotidae (Eutardigrada). Zootaxa 2011, 2761, 34–40. [Google Scholar] [CrossRef]
- Plate, L.H. Beiträge zur Naturgeschichte der Tardigraden. Zool. Jahrb. Anat. 1889, 3, 487–550. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Zawierucha, K.; Buda, J.; Stec, D.; Gawlak, M.; Michalczyk, Ł.; Roszkowska, M. An integrative redescription of the nominal taxon for the Mesobiotus harmsworthi group (Tardigrada: Macrobiotidae) leads to descriptions of two new Mesobiotus species from Arctic. PLoS ONE 2018, 13, e0204756. [Google Scholar] [CrossRef]
- Stec, D.; Morek, W.; Gąsiorek, P.; Michalczyk, Ł. Unmasking hidden species diversity within the Ramazzottius oberhaeuseri complex, with an integrative redescription of the nominal species for the family Ramazzottiidae (Tardigrada: Eutardigrada: Parachela). Syst. Biodivers. 2018, 16, 357–376. [Google Scholar] [CrossRef]
- Guidetti, R.; Cesari, M.; Bertolani, R.; Altiero, T.; Rebecchi, L. High diversity in species, reproductive modes and distribution within the Paramacrobiotus richtersi complex (Eutardigrada, Macrobiotidae). Zool. Lett. 2019, 5, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.; Krzywański, Ł.; Arakawa, K.; Michalczyk, Ł. A new redescription of Richtersius coronifer, supported by transcriptome, provides resources for describing concealed species diversity within the monotypic genus Richtersius (Eutardigrada). Zool. Lett. 2020, 6, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Stec, D.; Krzywański, Ł.; Zawierucha, K.; Michalczyk, Ł. Untangling systematics of the Paramacrobiotus areolatus species complex by an integrative redescription of the nominal species for the group, with multilocus phylogeny and species delineation in the genus Paramacrobiotus. Zool. J. Linn. Soc. 2020, 188, 694–716. [Google Scholar] [CrossRef]
- Stec, D.; Vecchi, M.; Maciejowski, W.; Michalczyk, Ł. Resolving the systematics of Richtersiidae by multilocus phylogeny and an integrative redescription of the nominal species for the genus Crenubiotus (Tardigrada). Sci. Rep. 2020, 10, 19418. [Google Scholar] [CrossRef]
- Roszkowska, M.; Grobys, D.; Bartylak, T.; Gawlak, M.; Kmita, H.; Kepel, A.; Kepel, M.; Parnikoza, I.; Kaczmarek, Ł. Integrative description of five Pseudechiniscus species (Heterotardigrada: Echiniscidae: The suillus-facettalis complex). Zootaxa 2020, 4763, 451–484. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Kayastha, P.; Gawlak, M.; Mioduchowska, M.; Roszkowska, M. An integrative description of Diploechiniscus oihonnae (Richters, 1903) population from near the original type locality in Merok (Norway). Zootaxa 2021, 4964, 83–102. [Google Scholar] [CrossRef]
- Stec, D.; Vecchi, M.; Dudziak, M.; Bartels, P.J.; Calhim, S.; Michalczyk, Ł. Integrative taxonomy resolves species identities within the Macrobiotus pallarii complex (Eutardigrada: Macrobiotidae). Zool. Lett. 2021, 7, 9. [Google Scholar] [CrossRef]
- Stec, D.; Vončina, K.; Kristensen, R.M.; Michalczyk, Ł. The Macrobiotus ariekammensis species complex provides evidence for parallel evolution of claw elongation in macrobiotid tardigrades. Zool. J. Linn. Soc. 2021; in press. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Kayastha, P.; Gawlak, M.; Mioduchowska, M.; Roszkowska, M. An integrative redescription of Echiniscus quadrispinosus quadrispinosus Richters, 1902 (Heterotardigrada; Echiniscidae) from the terra typica in Taunus Mountain Range (Europe; Germany). Eur. Zool. J. 2022; in press. [Google Scholar]
- Dastych, H. Niesporczaki (Tardigrada) Tatrzańskiego Parku Narodowego; Monografie Fauny Polski; PWN: Warszawa, Poland, 1980; pp. 1–232. [Google Scholar]
- Roszkowska, M.; Stec, D.; Gawlak, M.; Kaczmarek, Ł. An integrative description of a new tardigrade species Mesobiotus romani sp. nov. (Macrobiotidae: Harmsworthi group) from the Ecuadorian Pacific coast. Zootaxa 2018, 4450, 550–564. [Google Scholar] [CrossRef]
- Stec, D.; Gąsiorek, P.; Morek, W.; Kosztyła, P.; Zawierucha, K.; Michno, K.; Kaczmarek, Ł.; Prokop, Z.M.; Michalczyk, Ł. Estimating optimal sample size for tardigrade morphometry. Zool. J. Linn. Soc. 2016, 178, 776–784. [Google Scholar] [CrossRef] [Green Version]
- Pilato, G.; Binda, M.G. Definition of families, subfamilies, genera and subgenera of the Eutardigrada, and keys to their identification. Zootaxa 2010, 2404, 1–54. [Google Scholar] [CrossRef] [Green Version]
- Michalczyk, Ł.; Kaczmarek, Ł. A description of the new tardigrade Macrobiotus reinhardti (Eutardigrada: Macrobiotidae, harmsworthi group) with some remarks on the oral cavity armature within the genus Macrobiotus Schultze. Zootaxa 2003, 331, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł. The Macrobiotus hufelandi group (Tardigrada) revisited. Zootaxa 2017, 4363, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, Ł.; Cytan, J.; Zawierucha, K.; Diduszko, D.; Michalczyk, Ł. Tardigrades from Peru (South America), with descriptions of three new species of Parachela. Zootaxa 2014, 3790, 357–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pilato, G. Analisi di nuovi caratteri nello studio degli Eutardigradi. Animalia 1981, 8, 51–57. [Google Scholar]
- Kiosya, Y.; Pogwizd, J.; Matsko, Y.; Vecchi, M.; Stec, D. Phylogenetic position of two Macrobiotus species with a revisional note on Macrobiotus sottilei Pilato, Kiosya, Lisi & Sabella, 2012 (Tardigrada: Eutardigrada: Macrobiotidae). Zootaxa 2021, 4933, 113–135. [Google Scholar] [CrossRef]
- Michalczyk, Ł.; Kaczmarek, Ł. The Tardigrada Register: A comprehensive online data repository for tardigrade taxonomy. J. Limnol. 2013, 72, e22. [Google Scholar] [CrossRef]
- Casquet, J.; Thébaud, C.; Gillespie, R.G. Chelex without boiling, a rapid and easy technique to obtain stable amplifiable DNA from small amounts of ethanol-stored spiders. Mol. Ecol. Resour. 2011, 12, 136–141. [Google Scholar] [CrossRef]
- Kaczmarek, Ł.; Grobys, D.; Kulpa, A.; Bartylak, T.; Kmita, H.; Kepel, M.; Kepel, A.; Roszkowska, M. Two new species of the genus Milnesium Doyère, 1840 (Tardigrada, Apochela, Milnesiidae) from Madagascar. ZooKeys 2019, 884, 1–22. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxi-dase subunit I from diverse metazoan invertebrates. Mol. Marine Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Sands, C.J.; McInnes, S.J.; Marley, N.J.; Goodall-Copestake, W.P.; Convey, P.; Linse, K. Phylum Tardigrada: An “individual” approach. Cladistics 2008, 24, 861–871. [Google Scholar] [CrossRef]
- Mironov, S.V.; Dabert, J.; Dabert, M. A new feather mite species of the genus Proctophyllodes Robin, 1877 (Astigmata: Proctophyllodidae) from the Long-tailed Tit Aegithalos caudatus (Passeriformes: Aegithalidae)—morphological description with DNA barcode data. Zootaxa 2012, 3253, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Mioduchowska, M.; Kačarević, U.; Miamin, V.; Giginiak, Y.; Parnikoza, I.; Roszkowska, M.; Kaczmarek, Ł. Redescription of Antarctic eutardigrade Dastychius improvisus (Dastych, 1984) and some remarks on phylogenetic relationships within Isohypsibioidea. Eur. Zool. J. 2021, 88, 117–131. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; López, R. A new bioinformatics analysis tools framework at EMBL-EBI. Nucleic Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D.; Buckley, T.R. Model Selection and Model Averaging in Phylogenetics: Advantages of Akaike Information Criterion and Bayesian Approaches Over Likelihood Ratio Tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Yano, T.-A. Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Kuzdrowska, K.A.; Mioduchowska, M.; Gawlak, M.; Bartylak, T.; Kepel, A.; Kepel, M.; Kaczmarek, Ł. Integrative description of Macrobiotus porifini sp. nov. (Macrobiotidae) from Madagascar and its phylogenetic position within the hufelandi group. Eur. Zool. J. 2021, 88, 375–389. [Google Scholar] [CrossRef]
- Doyère, P.L.N. Memoire sur les Tardigrades. Ann. Sci. Nat. Ser. Zool. 1840, 14, 269–362. [Google Scholar]
- Richters, F. Tardigrada. In Handbuch der Zoologie; Kükenthal, W., Krumbach, T., Eds.; Walter de Gruyter & Co.: Berlin, Germany; Leipzig, Germany, 1926; Volume 3, pp. 58–61. [Google Scholar]
- Thulin, G. Über die phylogenie und das system der tardigraden. Hereditas 2010, 11, 207–266. [Google Scholar] [CrossRef]
- Marley, N.; McInnes, S.J.; Sands, C. Phylum Tardigrada: A re-evaluation of the Parachela. Zootaxa 2011, 2819, 51–64. [Google Scholar] [CrossRef]
- Vicente, F.; Cesari, M.; Serrano, A.; Bertolani, R. The impact of fire on terrestrial tardigrade biodiversity: A first case-study from Portugal. J. Limnol. 2013, 72, e19. [Google Scholar] [CrossRef] [Green Version]
- Vecchi, M.; Ferrari, C.; Stec, D.; Calhim, S. Desiccation risk favours prevalence and diversity of tardigrade communities and influences their trophic structure in alpine ephemeral rock pools. Hydrobiologia, 2022; in press. [Google Scholar] [CrossRef]
- Pilato, G.; Lisi, O.P.V. Notes on some tardigrades from southern Mexico with description of three new species. Zootaxa 2006, 1236, 53–68. [Google Scholar] [CrossRef]
- McInnes, S.J. Zoogeographic distribution of terrestrial/freshwater tardigrades from current literature. Ann. Mag. Nat. Hist. 1994, 28, 257–352. [Google Scholar] [CrossRef]
- Meyer, H.A. Terrestrial and freshwater Tardigrada of the Americas. Zootaxa 2013, 3747, 1–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł.; McInnes, S.J. Annotated zoogeography of non-marine Tardigrada. Part I: Central America. Zootaxa 2014, 3763, 1–62. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł.; Mcinnes, S.J. Annotated zoogeography of non-marine Tardigrada. Part II: South America. Zootaxa 2015, 3923, 1–107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, Ł.; Michalczyk, Ł.; McInnes, S.J. Annotated zoogeography of non-marine Tardigrada. Part III: North America and Greenland. Zootaxa 2016, 4203, 1–249. [Google Scholar] [CrossRef]
- McInnes, S.J.; Michalczyk, Ł.; Kaczmarek, Ł. Annotated zoogeography of non-marine Tardigrada. Part IV: Africa. Zootaxa 2017, 4284, 1–74. [Google Scholar] [CrossRef] [Green Version]
- Michalczyk, Ł.; Kaczmarek, Ł.; Mcinnes, S.J. Annotated zoogeography of non-marine Tardigrada. Part V: Australasia. Zootaxa 2022, 5107, 1–119. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CHARACTER | N | RANGE | MEAN | SD | Neotype | ||||
|---|---|---|---|---|---|---|---|---|---|
| µm | pt | µm | pt | µm | pt | µm | pt | ||
| Body length | 20 | 149–245 | – | 204 | – | 29 | – | 210 | – |
| Buccopharyngeal tube | |||||||||
| Buccal tube length | 20 | 17.5–24.3 | – | 21.7 | – | 2.0 | – | 22.4 | – |
| Stylet support insertion point | 20 | 9.8–13.3 | 53.8–56.3 | 11.9 | 54.8 | 1.1 | 0.7 | 12.2 | 54.6 |
| Buccal tube external width | 20 | 1.5–2.1 | 7.6–9.2 | 1.9 | 8.5 | 0.2 | 0.4 | 1.9 | 8.5 |
| Buccal tube internal width | 20 | 0.4–0.6 | 2.1–2.9 | 0.6 | 2.5 | 0.1 | 0.2 | 0.6 | 2.7 |
| Ventral lamina length | 19 | 6.2–9.2 | 34.8–38.9 | 8.0 | 37.0 | 0.9 | 1.3 | 7.9 | 35.3 |
| Placoid lengths | |||||||||
| Macroplacoid 1 | 20 | 1.6–2.0 | 8.1–9.6 | 1.9 | 8.8 | 0.1 | 0.5 | 1.9 | 8.5 |
| Macroplacoid 2 | 20 | 1.3–1.9 | 6.9–8.2 | 1.6 | 7.5 | 0.2 | 0.3 | 1.7 | 7.6 |
| Macroplacoid 3 | 20 | 1.4–1.9 | 6.8–8.2 | 1.7 | 7.8 | 0.2 | 0.3 | 1.8 | 8.0 |
| Microplacoid | 20 | 0.4–0.7 | 2.1–3.1 | 0.6 | 2.6 | 0.1 | 0.3 | 0.6 | 2.7 |
| Macroplacoid row | 20 | 5.2–7.2 | 27.2–31.9 | 6.3 | 29.1 | 0.5 | 1.3 | 6.5 | 29.0 |
| Placoid row | 20 | 5.9–8.1 | 30.2–35.4 | 7.1 | 32.8 | 0.6 | 1.4 | 7.3 | 32.6 |
| Claw I heights | |||||||||
| External primary branch | 18 | 4.3–5.8 | 22.1–25.4 | 5.2 | 23.9 | 0.5 | 0.9 | 5.3 | 23.7 |
| External secondary branch | 16 | 2.8–4.8 | 16.0–20.5 | 4.0 | 18.2 | 0.5 | 1.4 | 3.9 | 17.4 |
| Internal primary branch | 18 | 4.2–5.6 | 21.0–24.8 | 5.0 | 22.8 | 0.4 | 1.1 | 5.1 | 22.8 |
| Internal secondary branch | 15 | 2.7–4.5 | 15.3–19.4 | 3.8 | 17.2 | 0.5 | 1.4 | 4.1 | 18.3 |
| Claw II heights | |||||||||
| External primary branch | 17 | 4.5–6.4 | 23.9–27.7 | 5.5 | 25.8 | 0.5 | 1.1 | 5.7 | 25.4 |
| External secondary branch | 17 | 3.0–5.2 | 17.1–22.2 | 4.2 | 19.6 | 0.6 | 1.6 | 4.8 | 21.4 |
| Internal primary branch | 17 | 4.1–5.9 | 22.6–25.2 | 5.2 | 24.0 | 0.5 | 0.8 | 5.5 | 24.6 |
| Internal secondary branch | 17 | 2.9–4.7 | 16.6–20.5 | 4.0 | 18.5 | 0.5 | 1.0 | 4.3 | 19.2 |
| Claw III heights | |||||||||
| External primary branch | 18 | 4.5–6.4 | 23.9–27.7 | 5.5 | 25.7 | 0.5 | 1.1 | 5.6 | 25.0 |
| External secondary branch | 16 | 3.0–5.5 | 16.9–23.5 | 4.3 | 19.8 | 0.6 | 1.7 | 4.6 | 20.5 |
| Internal primary branch | 18 | 4.1–6.2 | 22.3–26.5 | 5.3 | 24.1 | 0.6 | 1.2 | 5.0 | 22.3 |
| Internal secondary branch | 16 | 3.1–5.4 | 17.0–23.1 | 4.1 | 18.7 | 0.6 | 1.5 | 4.0 | 17.9 |
| Claw IV heights | |||||||||
| Anterior primary branch | 13 | 5.3–6.7 | 24.3–28.2 | 6.0 | 26.9 | 0.4 | 1.2 | 6.3 | 28.1 |
| Anterior secondary branch | 13 | 4.0–5.5 | 18.9–23.5 | 4.8 | 21.3 | 0.4 | 1.4 | 4.4 | 19.6 |
| Posterior primary branch | 14 | 5.8–6.8 | 25.5–30.8 | 6.3 | 28.1 | 0.3 | 1.4 | 6.5 | 29.0 |
| Posterior secondary branch | 14 | 4.3–6.0 | 18.6–25.6 | 5.0 | 22.2 | 0.5 | 1.7 | 4.9 | 21.9 |
| CHARACTER | N | RANGE | MEAN | SD |
|---|---|---|---|---|
| Egg bare diameter | 8 | 46.3–54.5 | 51.4 | 2.5 |
| Egg full diameter | 8 | 52.1–61.4 | 58.0 | 2.8 |
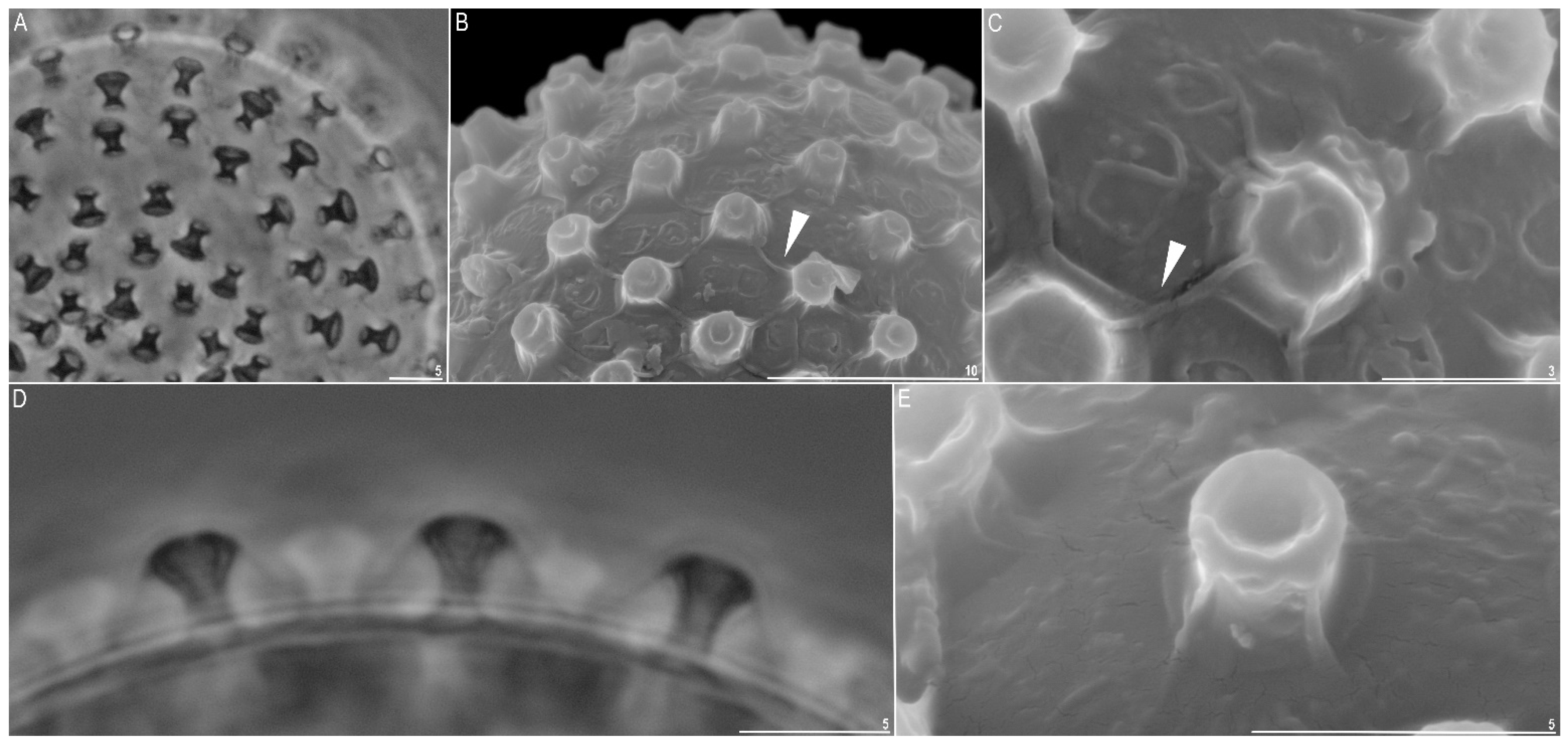
| Process height | 24 | 3.0–3.9 | 3.4 | 0.2 |
| Process base width | 24 | 1.3–2.0 | 1.6 | 0.2 |
| Process base/height ratio | 24 | 41–61% | 47% | 5% |
| Top of processes width | 24 | 2.4–3.5 | 2.9 | 0.3 |
| Inter-process distance | 24 | 2.4–4.0 | 3.3 | 0.5 |
| Number of processes on the egg circumference | 8 | 28–32 | 30.0 | 1.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczmarek, Ł.; Kayastha, P.; Roszkowska, M.; Gawlak, M.; Mioduchowska, M. Integrative Redescription of the Minibiotus intermedius (Plate, 1888)—The Type Species of the Genus Minibiotus R.O. Schuster, 1980. Diversity 2022, 14, 356. https://doi.org/10.3390/d14050356
Kaczmarek Ł, Kayastha P, Roszkowska M, Gawlak M, Mioduchowska M. Integrative Redescription of the Minibiotus intermedius (Plate, 1888)—The Type Species of the Genus Minibiotus R.O. Schuster, 1980. Diversity. 2022; 14(5):356. https://doi.org/10.3390/d14050356
Chicago/Turabian StyleKaczmarek, Łukasz, Pushpalata Kayastha, Milena Roszkowska, Magdalena Gawlak, and Monika Mioduchowska. 2022. "Integrative Redescription of the Minibiotus intermedius (Plate, 1888)—The Type Species of the Genus Minibiotus R.O. Schuster, 1980" Diversity 14, no. 5: 356. https://doi.org/10.3390/d14050356
APA StyleKaczmarek, Ł., Kayastha, P., Roszkowska, M., Gawlak, M., & Mioduchowska, M. (2022). Integrative Redescription of the Minibiotus intermedius (Plate, 1888)—The Type Species of the Genus Minibiotus R.O. Schuster, 1980. Diversity, 14(5), 356. https://doi.org/10.3390/d14050356