
Reproductive Behaviours and Potentially Associated Sounds of the Mottled Grouper Mycteroperca rubra: Implications for Conservation

 ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.2. Underwater Visual Census (UVC)
2.3. Observations of Reproductive Behaviours
2.4. Acoustic Recordings
2.4.1. Data Collection
2.4.2. Data Analysis
2.4.3. Temporal Patterns of Sound Production
2.4.4. Relationship between Visual Observations and Presumed Acoustic Behaviour
3. Results
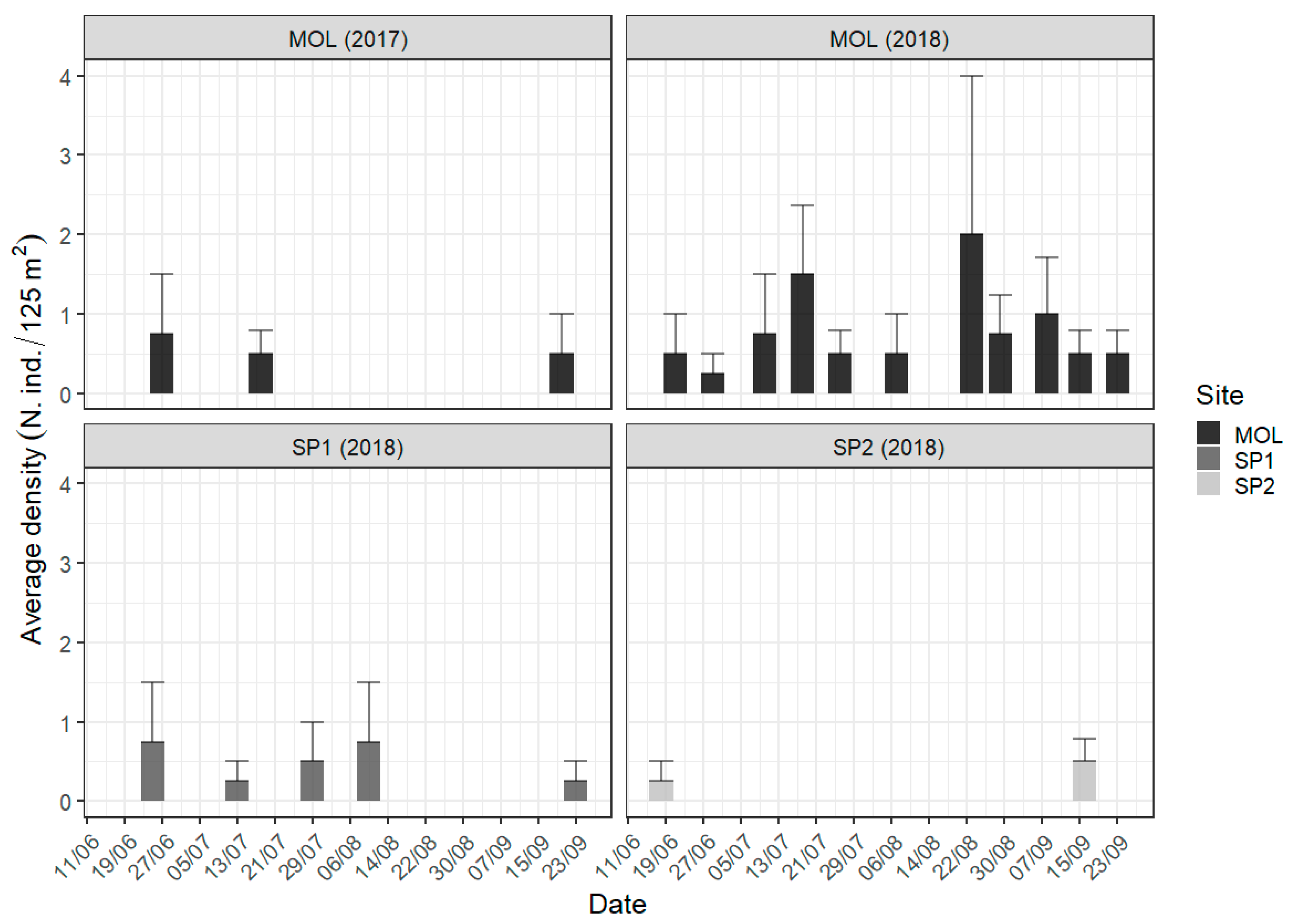
3.1. Underwater Visual Census (UVC)
3.2. Behavioural Observations
3.3. Acoustic Recordings
3.3.1. Temporal Patterns in Sound Production
3.3.2. Relationship between Visual Observations and Presumed Acoustic Behaviour
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jackson, J.B.C. Historical Overfishing and the Recent Collapse of Coastal Ecosystems. Science 2001, 293, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Lotze, H.K.; Tittensor, D.P.; Bryndum-Buchholz, A.; Eddy, T.D.; Cheung, W.W.L.; Galbraith, E.D.; Barange, M.; Barrier, N.; Bianchi, D.; Blanchard, J.L.; et al. Global Ensemble Projections Reveal Trophic Amplification of Ocean Biomass Declines with Climate Change. Proc. Natl. Acad. Sci. USA 2019, 116, 12907–12912. [Google Scholar] [CrossRef] [PubMed]
- Halpern, B.S.; Frazier, M.; Afflerbach, J.; Lowndes, J.S.; Micheli, F.; O’Hara, C.; Scarborough, C.; Selkoe, K.A. Recent Pace of Change in Human Impact on the World’s Ocean. Sci. Rep. 2019, 9, 11609. [Google Scholar] [CrossRef] [PubMed]
- Grorud-Colvert, K.; Sullivan-Stack, J.; Roberts, C.; Constant, V.; Horta, E.; Costa, B.; Pike, E.P.; Kingston, N.; Laffoley, D.; Sala, E.; et al. The MPA Guide: A Framework to Achieve Global Goals for the Ocean. Science 2021, 373, eabf0861. [Google Scholar] [CrossRef] [PubMed]
- Giakoumi, S.; Scianna, C.; Plass-Johnson, J.; Micheli, F.; Grorud-Colvert, K.; Thiriet, P.; Claudet, J.; Di Carlo, G.; Di Franco, A.; Gaines, S.D.; et al. Ecological Effects of Full and Partial Protection in the Crowded Mediterranean Sea: A Regional Meta-Analysis. Sci. Rep. 2017, 7, 8940. [Google Scholar] [CrossRef] [PubMed]
- Rojo, I.; Sánchez-Meca, J.; García-Charton, J.A. Small-Sized and Well-Enforced Marine Protected Areas Provide Ecological Benefits for Piscivorous Fish Populations Worldwide. Mar. Environ. Res. 2019, 149, 100–110. [Google Scholar] [CrossRef]
- Hackradt, C.W.; Garcìa-Charton, J.A.; Harmelin-Vivien, M.; Pérez-Ruzafa, A.; Le Diréach, L.; Bayle-Sempere, J.; Charbonnel, E.; Ody, D.; Renones, O.; Sanchez-Jerez, P.; et al. Response of Rocky Reef Top Predators (Serranidae: Epinephelinae) in and around Marine Protected Areas in the Western Mediterranean Sea. PLoS ONE 2014, 9, e98206. [Google Scholar] [CrossRef]
- Erisman, B.; Heyman, W.; Kobara, S.; Ezer, T.; Pittman, S.; Aburto-Oropeza, O.; Nemeth, R.S. Fish Spawning Aggregations: Where Well-Placed Management Actions Can Yield Big Benefits for Fisheries and Conservation. Fish Fish. 2017, 18, 128–144. [Google Scholar] [CrossRef]
- Grüss, A.; Robinson, J.; Heppell, S.S.; Heppell, S.A.; Semmens, B.X. Conservation and Fisheries Effects of Spawning Aggregation Marine Protected Areas: What We Know, Where We Should Go, and What We Need to Get There. ICES J. Mar. Sci. 2014, 71, 1515–1534. [Google Scholar] [CrossRef]
- Di Franco, A.; Di Lorenzo, M.; Guidetti, P. Spatial Patterns of Density at Multiple Life Stages in Protected and Fished Conditions: An Example from a Mediterranean Coastal Fish. J. Sea Res. 2013, 76, 73–81. [Google Scholar] [CrossRef]
- Harrison, H.B.; Williamson, D.H.; Evans, R.D.; Almany, G.R.; Thorrold, S.R.; Russ, G.R.; Feldheim, K.A.; van Herwerden, L.; Planes, S.; Srinivasan, M.; et al. Larval Export from Marine Reserves and the Recruitment Benefit for Fish and Fisheries. Curr. Biol. 2012, 22, 1023–1028. [Google Scholar] [CrossRef] [PubMed]
- Harrison, H.B.; Bode, M.; Williamson, D.H.; Berumen, M.L.; Jones, G.P. A Connectivity Portfolio Effect Stabilizes Marine Reserve Performance. Proc. Natl. Acad. Sci. USA 2020, 117, 25595–25600. [Google Scholar] [CrossRef] [PubMed]
- Marshall, D.J.; Gaines, S.; Warner, R.; Barneche, D.R.; Bode, M. Underestimating the Benefits of Marine Protected Areas for the Replenishment of Fished Populations. Front. Ecol. Environ. 2019, 17, 407–413. [Google Scholar] [CrossRef]
- European Commission. Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. EU Biodiversity Strategy for 2030. Bringing Nature Back into Our Lives. COM/2020/380 final, Brussels. 2020. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:52020DC0380 (accessed on 10 February 2022).
- Hermoso, V.; Carvalho, S.B.; Giakoumi, S.; Goldsborough, D.; Katsanevakis, S.; Leontiou, S.; Markantonatou, V.; Rumes, B.; Vogiatzakis, I.N.; Yates, K.L. The EU Biodiversity Strategy for 2030: Opportunities and Challenges on the Path towards Biodiversity Recovery. Environ. Sci. Policy 2022, 127, 263–271. [Google Scholar] [CrossRef]
- Biggs, C.R.; Heyman, W.D.; Farmer, N.A.; Kobara, S.; Bolser, D.G.; Robinson, J.; Lowerre-Barbieri, S.K.; Erisman, B.E. The Importance of Spawning Behavior in Understanding the Vulnerability of Exploited Marine Fishes in the U.S. Gulf of Mexico. PeerJ 2021, 9, e11814. [Google Scholar] [CrossRef]
- Sadovy de Mitcheson, Y.; Colin, P.L. Reef Fish Spawning Aggregations: Biology, Research and Management; Sadovy de Mitcheson, Y., Colin, P.L., Eds.; Springer: Dordrecht, The Netherlands, 2012; p. 622. [Google Scholar]
- Sadovy de Mitcheson, Y. Mainstreaming Fish Spawning Aggregations into Fishery Management Calls for a Precautionary Approach. Bioscience 2016, 66, 295–306. [Google Scholar] [CrossRef]
- Domeier, M. Revisting Spawning Aggregations: Definitions and Challenges. In Reef Fish Spawning Aggregations: Biology, Research and Management; Sadovy de Mitcheson, Y., Colin, P.L., Eds.; Springer: Dordrecht, The Netherlands, 2012; pp. 1–20. [Google Scholar]
- Domeier, M.L.; Colin, P.L. Tropical Reef Fish Spawning Aggregations: Defined and Reviewed. Bull. Mar. Sci. 1997, 60, 698–726. [Google Scholar]
- Sadovy, Y.; Domeier, M. Are Aggregation-Fisheries Sustainable? Reef Fish Fisheries as a Case Study. Coral Reefs 2005, 24, 254–262. [Google Scholar] [CrossRef]
- Sadovy de Mitcheson, Y.; Craig, M.T.; Bertoncini, A.A.; Carpenter, K.E.; Cheung, W.W.L.; Choat, J.H.; Cornish, A.S.; Fennessy, S.T.; Ferreira, B.P.; Heemstra, P.C.; et al. Fishing Groupers towards Extinction: A Global Assessment of Threats and Extinction Risks in a Billion Dollar Fishery. Fish Fish. 2013, 14, 119–136. [Google Scholar] [CrossRef]
- Amorim, M.C.P. Diversity of Sound Production in Fish. In Communication in Fishes; Ladich, F., Collin, S.P., Moller, P., Kapoor, B.G., Eds.; Science Publishers: Enfield, NH, USA; Plymouth, UK, 2006; Volume 1, pp. 71–105. [Google Scholar]
- Rountree, R.A.; Gilmore, R.G.; Goudey, C.A.; Hawkins, A.D.; Luczkovich, J.J.; Mann, D.A. Listening to Fish: Applications of Passive Acoustics to Fisheries Science. Fisheries 2006, 31, 433–446. [Google Scholar] [CrossRef]
- Locascio, J.V.; Mann, D.A. Diel Periodicity of Fish Sound Production in Charlotte Harbor, Florida. Trans. Am. Fish. Soc. 2008, 137, 606–615. [Google Scholar] [CrossRef]
- Luczkovich, J.J.; Mann, D.A.; Rountree, R.A. Passive Acoustics as a Tool in Fisheries Science. Trans. Am. Fish. Soc. 2008, 137, 533–541. [Google Scholar] [CrossRef]
- Picciulin, M.; Calcagno, G.; Sebastianutto, L.; Bonacito, C.; Codarin, A.; Costantini, M.; Ferrero, E.A. Diagnostics of Nocturnal Calls of Sciaena Umbra (L., Fam. Sciaenidae) in a Nearshore Mediterranean Marine Reserve. Bioacoustics Int. J. Anim. Sound Rec. 2012, 22, 109–120. [Google Scholar] [CrossRef]
- Locascio, J.V.; Burton, M.L. A Passive Acoustic Survey of Fish Sound Production at Riley’s Hump within Tortugas South Ecological Reserve: Implications Regarding Spawning and Habitat Use. Fish. Bull. 2015, 114, 103–116. [Google Scholar] [CrossRef]
- Erisman, B.E.; Rowell, T.J. A Sound Worth Saving: Acoustic Characteristics of a Massive Fish Spawning Aggregation. Biol. Lett. 2017, 13, 20170656. [Google Scholar] [CrossRef] [PubMed]
- Lindseth, A.; Lobel, P. Underwater Soundscape Monitoring and Fish Bioacoustics: A Review. Fishes 2018, 3, 36. [Google Scholar] [CrossRef]
- Caiger, P.E.; Dean, M.J.; DeAngelis, A.I.; Hatch, L.T.; Rice, A.N.; Stanley, J.A.; Tholke, C.; Zemeckis, D.R.; van Parijs, S.M. A Decade of Monitoring Atlantic Cod Gadus Morhua Spawning Aggregations in Massachusetts Bay Using Passive Acoustics. Mar. Ecol. Prog. Ser. 2020, 635, 89–103. [Google Scholar] [CrossRef]
- Parmentier, E.; Di Iorio, L.; Picciulin, M.; Malavasi, S.; Lagardère, J.-P.; Bertucci, F. Consistency in Spatiotemporal Sound Features Supports the Use of Passive Acoustic for Long-Term Monitoring. Anim. Conserv. 2017, 21, 211–220. [Google Scholar] [CrossRef]
- Mann, D.A.; Locascio, J.V.; Coleman, F.C.; Koenig, C.C. Goliath Grouper Epinephelus Itajara Sound Production and Movement Patterns on Aggregation Sites. Endanger. Species Res. 2009, 7, 229–236. [Google Scholar] [CrossRef]
- Mann, D.; Locascio, J.; Schärer, M.; Nemeth, M.; Appeldoorn, R. Sound Production by Red Hind Epinephelus Guttatus in Spatially Segregated Spawning Aggregations. Aquat. Biol. 2010, 10, 149–154. [Google Scholar] [CrossRef]
- Nelson, M.D.; Koenig, C.C.; Coleman, F.C.; Mann, D.A. Sound Production of Red Grouper Epinephelus Morio on the West Florida Shelf. Aquat. Biol. 2011, 12, 97–108. [Google Scholar] [CrossRef]
- Schärer, M.T.; Nemeth, M.I.; Mann, D.; Locascio, J.; Appeldoorn, R.S.; Rowell, T.J. Sound Production and Reproductive Behavior of Yellowfin Grouper, Mycteroperca venenosa (Serranidae) at a Spawning Aggregation. Copeia 2012, 1, 135–144. [Google Scholar] [CrossRef]
- Schärer, M.T.; Rowell, T.J.; Nemeth, M.I.; Appeldoorn, R.S. Sound Production Associated with Reproductive Behavior of Nassau Grouper Epinephelus striatus at Spawning Aggregations. Endanger. Species Res. 2012, 19, 29–38. [Google Scholar] [CrossRef]
- Schärer, M.T.; Nemeth, M.I.; Rowell, T.J.; Appeldoorn, R.S. Sounds Associated with the Reproductive Behavior of the Black Grouper (Mycteroperca bonaci). Mar. Biol. 2014, 161, 141–147. [Google Scholar] [CrossRef]
- Rowell, T.J.; Aburto-Oropeza, O.; Cota-Nieto, J.J.; Steele, M.A.; Erisman, B.E. Reproductive Behaviour and Concurrent Sound Production of Gulf Grouper Mycteroperca jordani (Epinephelidae) at a Spawning Aggregation Site. J. Fish Biol. 2019, 94, 277–296. [Google Scholar] [CrossRef]
- Ibrahim, A.K.; Chérubin, L.M.; Zhuang, H.; Umpierre, M.T.S.; Dalgleish, F.; Ouyang, B.; Dalgleish, A.; Zhuang, H.; Sch, M.T. An Approach for Automatic Classification of Grouper Vocalizations with Passive Acoustic Monitoring. J. Acoust. Soc. Am. 2018, 143, 666–676. [Google Scholar] [CrossRef]
- Wilson, K.; Semmens, B.; Pattengill-Semmens, C.; McCoy, C.; McCoy, C. Potential for Grouper Acoustic Competition and Partitioning at a Multispecies Spawning Site off Little Cayman, Cayman Islands. Mar. Ecol. Prog. Ser. 2020, 634, 127–146. [Google Scholar] [CrossRef]
- Colin, P.L. Reproduction of the Nassau Grouper, Epinephelus striatus (Pisces: Serranidae) and Its Relationship to Environmental Conditions. Environ. Biol. Fishes 1992, 34, 357–377. [Google Scholar] [CrossRef]
- Luckhurst, B.E. Observations of a Black Grouper (Mycteroperca bonaci) Spawning Aggregation in Bermuda. Gulf Caribb. Res. 2010, 22, 43–49. [Google Scholar] [CrossRef]
- Erisman, B.E.; Buckhorn, M.L.; Hastings, P.A. Spawning Patterns in the Leopard Grouper, Mycteroperca Rosacea, in Comparison with Other Aggregating Groupers. Mar. Biol. 2007, 151, 1849–1861. [Google Scholar] [CrossRef]
- Craig, M.T.; Sadovy de Mitcheson, Y.; Heemstra, P.C. Groupers of the World: A Field and Market Guide; NISC (Pty) Ltd.: Grahamstown, South Africa, 2011. [Google Scholar]
- Bertucci, F.; Lejeune, P.; Payrot, J.; Parmentier, E. Sound Production by Dusky Grouper Epinephelus Marginatus at Spawning Aggregation Sites. J. Fish Biol. 2015, 87, 400–421. [Google Scholar] [CrossRef]
- Hereu, B.; Diaz, D.; Pasqual, J.; Zabala, M.; Sala, E. Temporal Patterns of Spawning of the Dusky Grouper Epinephelus Marginatus in Relation to Environmental Factors. Mar. Ecol. Prog. Ser. 2006, 325, 187–194. [Google Scholar] [CrossRef]
- Zabala, M.; Garcia-Rubies, A.; Louisy, P.; Sala, E. Spawning Behaviour of the Mediterranean Dusky Grouper Epinephelus Marginatus (Lowe, 1834) (Pisces, Serranidae) in the Medes Islands Marine Reserve (NW Mediterranean, Spain). Sci. Mar. 1997, 61, 65–77. [Google Scholar]
- Condini, M.V.; García-Charton, J.A.; Garcia, A.M. A Review of the Biology, Ecology, Behavior and Conservation Status of the Dusky Grouper, Epinephelus marginatus (Lowe 1834). Rev. Fish Biol. Fish. 2017, 28, 301–330. [Google Scholar] [CrossRef]
- Tortonese, E. Fauna d’Italia, “Osteichtyes”: Pesci Ossei.; Calderini: Bologna, Italy, 1975; Volume 11. [Google Scholar]
- La Mesa, G.; Di Muccio, S.; Vacchi, M. Abundance, Size Distribution and Habitat Preferences in the Grouper Assemblage of the Ustica Marine Reserve (SW Mediterranean). Cybium 2006, 30, 365–377. [Google Scholar]
- Aronov, A.; Goren, M. Ecology of the Mottled Grouper (Mycteroperca rubra) in the Eastern Mediterranean. Electron. J. Ichthyol. 2008, 2, 43–55. [Google Scholar]
- Pollard, D.A.; Francour, P. Mycteroperca rubra, Mottled Grouper. The IUCN Red List of Threatened Species, 2018; e.T14054A42691814. [Google Scholar] [CrossRef]
- Glamuzina, B.; Tutman, P.; Kozul, V.; Glavic, N.; Skaramuca, B. The First Recorded Occurrence of the Mottled Grouper, Mycteroperca rubra (Serranidae), in the Southeastern Adriatic Sea. Cybium 2002, 26, 156–158. [Google Scholar]
- Cottalorda, J.-M.; Dominici, J.-M.; Harmelin, J.-G.; Harmelin-Vivien, M.; Louisy, P.; Francour, P. Etude et Synthèse Des Principales Données Disponibles Sur Les Espèces de «mérous» de La Réserve Naturelle de Scandola et de Ses Environs Immédiats; Contrat Parc Naturel Régional de Corse/GIS Posidonie; ECOMERS Publication; Université Nice Sophia Antipolis: Nice, France, 2012. [Google Scholar]
- Psomadakis, P.N.; Giustino, S.; Vacchi, M. Mediterranean Fish Biodiversity: An Updated Inventory with Focus on the Ligurian and Tyrrhenian Seas. Zootaxa 2012, 3263, 1–46. [Google Scholar] [CrossRef]
- Siau, Y.; Bouain, A. Preliminary Indications on Growth and Reproduction in the Protogynous Grouper Mycteroperca rubra (Pisces, Serranidae). J. Afr. Ecol. 1994, 108, 353–359. [Google Scholar]
- Gücü, A.C.; Bingel, F. Trawlable Species Assamblages on the Continental Shelf of the Northeastern Levant Sea with an Emphasis on Lessepsian Migration. Acta Adriat. 1994, 35, 83–100. [Google Scholar]
- Özvarol, Y.; Gökoğlu, M. Length-Weight Relationship of Hyporthodus haifensis and Mycteroperca rubra (Pisces; Serranidae) from the North-Eastern Mediterranean Sea, Turkey. J. Appl. Ichthyol. 2015, 31, 1165–1167. [Google Scholar] [CrossRef]
- Mavruk, S.; Saygu, İ.; Bengil, F.; Alan, V.; Azzurro, E. Grouper Fishery in the Northeastern Mediterranean: An Assessment Based on Interviews on Resource Users. Mar. Policy 2018, 87, 141–148. [Google Scholar] [CrossRef]
- Guidetti, P.; Micheli, F. Ancient Art Serving Marine Conservation. Front. Ecol. Environ. 2011, 9, 374–375. [Google Scholar] [CrossRef]
- Consoli, P.; Martino, A.; Romeo, T.; Sinopoli, M.; Perzia, P.; Canese, S.; Vivona, P.; Andaloro, F. The Effect of Shipwrecks on Associated Fish Assemblages in the Central Mediterranean Sea. J. Mar. Biol. Assoc. United Kingd. 2015, 95, 17–24. [Google Scholar] [CrossRef]
- Irigoyen, A.J.; Rojo, I.; Calò, A.; Trobbiani, G.; Sánchez-Carnero, N.; García-Charton, J.A. The “Tracked Roaming Transect” and Distance Sampling Methods Increase the Efficiency of Underwater Visual Censuses. PLoS ONE 2018, 13, e0190990. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.; García-Charton, J.A.; Bayle-Sempere, J.T.; Charbonnel, E.; Le Diréach, L.; Ody, D.; Pérez-Ruzafa, Á.; Reñones, O.; Sánchez-Jerez, P.; Valle, C. Importance of Marine Reserves for the Population Dynamics of Groupers (Epinephelinae) in the Western Mediterranean. In Proceedings of the Second Symposium on Mediterranean Groupers, Nice, France, 10–13 May 2007; Francour, P., Gratiot, J., Eds.; Université Nice Sophia Antipolis: Nice, France, 2007; pp. 91–93. [Google Scholar]
- Desiderà, E.; Trainito, E.; Navone, A.; Blandin, R.; Magnani, L.; Panzalis, P.; Mazzoldi, C.; Guidetti, P. Using Complementary Visual Approaches to Investigate Residency, Site Fidelity and Movement Patterns of the Dusky Grouper (Epinephelus Marginatus) in a Mediterranean Marine Protected Area. Mar. Biol. 2021, 168, 111. [Google Scholar] [CrossRef]
- Desiderà, E. Reproductive Behaviours of Groupers (Epinephelidae) in the Tavolara-Punta Coda Cavallo Marine Protected Area (NW Mediterranean Sea). Ph.D. Thesis, University of Padova, Padova, Italy, Université Côte d’Azur, Nice, France, 21 February 2020. [Google Scholar]
- Sahyoun, R.; Bussotti, S.; Di Franco, A.; Navone, A.; Panzalis, P.; Guidetti, P. Protection Effects on Mediterranean Fish Assemblages Associated with Different Rocky Habitats. J. Mar. Biol. Assoc. UK 2013, 93, 425–435. [Google Scholar] [CrossRef]
- Di Franco, A.; Bussotti, S.; Navone, A.; Panzalis, P.; Guidetti, P. Evaluating Effects of Total and Partial Restrictions to Fishing on Mediterranean Rocky-Reef Fish Assemblages. Mar. Ecol. Prog. Ser. 2009, 387, 275–285. [Google Scholar] [CrossRef]
- Harmelin-Vivien, M.; Harmelin, J.G.; Chauvet, C.; Duval, C.; Galzin, R.; Lejeune, P.; Barnabé, G.; Blane, F.; Chevalier, R.; Duclere, J.; et al. Evaluation Visuelle Des Peuplements et Populations de Poissons: Méthodes et Problémes. (The Underwater Observation of Fish Communities and Fish Populations. Methods and Problems). Rev. d’Écologie—La Terre La Vie 1985, 40, 467–539. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 10 February 2022).
- Parmentier, E.; Vandewalle, P.; Brié, C.; Dinraths, L.; Lecchini, D. Comparative Study on Sound Production in Different Holocentridae Species. Front. Zool. 2011, 8, 12. [Google Scholar] [CrossRef]
- Hazlett, B.; Winn, H.E. Sound Producing Mechanism of the Nassau Grouper, Epinephalus striatus. Copeia 1962, 1962, 447–449. [Google Scholar] [CrossRef]
- Wall, C.C.; Simard, P.; Lembke, C.; Mann, D.A. Large-Scale Passive Acoustic Monitoring of Fish Sound Production on the West Florida Shelf. Mar. Ecol. Prog. Ser. 2013, 484, 173–188. [Google Scholar] [CrossRef]
- Ladich, F. Sound Communication in Fishes. In Sound Communication in Fishes; Ladich, F., Ed.; Springer: Vienna, Austria, 2015; pp. 127–148. ISBN 978-3-7091-1845-0. [Google Scholar]
- Desiderà, E.; Guidetti, P.; Panzalis, P.; Navone, A.; Valentini-Poirrier, C.-A.; Boissery, P.; Gervaise, C.; Di Iorio, L. Acoustic Fish Communities: Sound Diversity of Rocky Habitats Reflects Fish Species Diversity. Mar. Ecol. Prog. Ser. 2019, 608, 183–197. [Google Scholar] [CrossRef]
- Bazile Kinda, G.; Simard, Y.; Gervaise, C.; Mars, J.I.; Fortier, L. Under-Ice Ambient Noise in Eastern Beaufort Sea, Canadian Arctic, and Its Relation to Environmental Forcing. J. Acoust. Soc. Am. 2013, 134, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Jublier, N.; Bertucci, F.; Kéver, L.; Colleye, O.; Ballesta, L.; Nemeth, R.S.; Lecchini, D.; Rhodes, K.L.; Parmentier, E. Passive Monitoring of Phenological Acoustic Patterns Reveals the Sound of the Camouflage Grouper, Epinephelus Polyphekadion. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 42–52. [Google Scholar] [CrossRef]
- Colin, P.L.; Sadovy, Y.J.; Domeier, M.L. Manual for the Study and Conservation of Reef Fish Spawning Aggregations; Special Publication No. 1 (Version 1.0); Society for the Conservation of Reef Fish Aggregations (SCRFA): Columbia, SC, USA, 2003; pp. 1–98. [Google Scholar]
- Di Iorio, L.; Audax, M.; Deter, J.; Holon, F.; Lossent, J.; Gervaise, C.; Boissery, P. Biogeography of Acoustic Biodiversity of NW Mediterranean Coralligenous Reefs. Sci. Rep. 2021, 11, 16991. [Google Scholar] [CrossRef]
- Bolgan, M.; Di Iorio, L.; Dailianis, T.; Catalan, I.A.; Lejeune, P.; Picciulin, M.; Parmentier, E. Fish Acoustic Community Structure in Neptune Seagrass Meadows across the Mediterranean Basin. Aquat. Conserv. Mar. Freshw. Ecosyst. 2022, 32, 329–347. [Google Scholar] [CrossRef]
- Desiderà, E.; Magnani, L.; Navone, A.; Guidetti, P.; Mazzoldi, C. First Direct Evidence of Reproductive Behaviour of the White Grouper, Epinephelus aeneus (Geoffroy Saint-Hilaire, 1817). Mediterr. Mar. Sci. 2019, 20, 326. [Google Scholar] [CrossRef]
- Bolgan, M.; Soulard, J.; Di Iorio, L.; Gervaise, C.; Lejeune, P.; Gobert, S.; Parmentier, E. The Sea Chordophones Make the Mysterious /Kwa/: Emitter Identification of the Dominating Fish Sound in Mediterranean Seagrass Meadows. J. Exp. Biol. 2019, 222, jeb.196931. [Google Scholar] [CrossRef]
- Kéver, L.; Lejeune, P.; Michel, L.N.; Parmentier, E. Passive Acoustic Recording of Ophidion Rochei Calling Activity in Calvi Bay (France). Mar. Ecol. 2016, 37, 1315–1324. [Google Scholar] [CrossRef]
- Picciulin, M.; Sebastianutto, L.; Costantini, M.; Rocca, M.; Ferrero, E.A. Aggressive Territorial Ethogram of the Red-Mouthed Goby, Gobius Cruentatus (Gmelin, 1789). Electron. J. Ichthyol. 2006, 2, 38–49. [Google Scholar]
- Picciulin, M.; Costantini, M.; Hawkins, A.D.; Ferrero, E.A. Sound Emissions of the Mediterranean Damselfish Chromis Chromis (Pomacentridae). Bioacoustics 2002, 12, 236–238. [Google Scholar] [CrossRef]
- Louisy, P.; Ganteaume, A.; Francour, P. Les Relations Des Espèces de Mérous à Leur Habitat—Epinephelus marginatus, E. costae et Mycteroperca rubra—Dans La Région de Kas, Turquie, Méditerranée Orientale. In Proceedings of the Second Symposium on Mediterranean Groupers, Nice, France, 10–13 May 2007; Francour, P., Gratiot, J., Eds.; Université Nice Sophia Antipolis: Nice, France, 2007; pp. 121–124. [Google Scholar]
- Hackradt, C.W.; Félix-Hackradt, F.C.; Treviño-Otón, J.; Pérez-Ruzafa, Á.; García-Charton, J.A. Density-Driven Habitat Use Differences across Fishing Zones by Predator Fishes (Serranidae) in South-Western Mediterranean Rocky Reefs. Hydrobiologia 2020, 847, 757–770. [Google Scholar] [CrossRef]
- Micheli, F.; Niccolini, F. Achieving Success under Pressure in the Conservation of Intensely Used Coastal Areas. Ecol. Soc. 2013, 18, 19. [Google Scholar] [CrossRef]
- Rojo, I.; Irigoyen, A.J.; Cuadros, A.; Calò, A.; Pereñíguez, J.M.; Hernández-Andreu, R.; Félix-Hackradt, F.C.; Carreño, F.; Hackradt, C.W.; García-Charton, J.A. Detection of Protection Benefits for Predatory Fishes Depends on Census Methodology. Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 1670–1685. [Google Scholar] [CrossRef]
- Sala, E.; Ballesteros, E.; Dendrinos, P.; Di Franco, A.; Ferretti, F.; Foley, D.; Fraschetti, S.; Friedlander, A.; Garrabou, J.; Güçlüsoy, H.; et al. The Structure of Mediterranean Rocky Reef Ecosystems across Environmental and Human Gradients, and Conservation Implications. PLoS ONE 2012, 7, e32742. [Google Scholar] [CrossRef]
- Nemeth, R.S. Ecosystem Aspects of Species That Aggregate to Spawn. In Reef Fish Spawning Aggregations: Biology, Research and Management; Springer: Dordrecht, The Netherlands, 2012; pp. 21–55. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | 2017 | 2018 | Total | |
|---|---|---|---|---|
| Total N. UVC transects | MOL | 32 | 56 | 88 |
| SP1 | 24 | 40 | 64 | |
| SP2 | 24 | 28 | 52 | |
| N. UVC transects where M. rubra was recorded | MOL | 5 | 18 | 23 |
| SP1 | 0 | 5 | 5 | |
| SP2 | 0 | 3 | 3 |
| 2017 | 2018 | ||||||
|---|---|---|---|---|---|---|---|
| Site | After Sunrise | During Daytime | Before Sunset | After Sunrise | During Daytime | Before Sunset | Total |
| MOL | 0 | 6 | 8 | 7 | 4 | 17 | 42 |
| SP1 | 3 | 7 | 5 | 10 | 6 | 15 | 46 |
| SP2 | 3 | 3 | 5 | 9 | 1 | 11 | 32 |
| Total | 6 | 16 | 18 | 26 | 11 | 43 | 120 |
| Sound Type | Variable | Minimum | Mean | ±SD | Maximum | n |
|---|---|---|---|---|---|---|
| Low-frequency Fast | ||||||
| Pulse Train (LFPT) | Duration (ms) | 242.0 | 477.0 | 122.5 | 817.0 | 43 |
| Pulse sound−1 | 4.0 | 9.1 | 2.7 | 15.0 | 43 | |
| Inter-pulse interval (IPI, ms) | 42.0 | 59.1 | 12.5 | 131.0 | 347 | |
| Peak frequency (Hz) | 30.5 | 82.2 | 35.3 | 219.1 | 87 | |
| Frequency 5% (Hz) | 0.0 | 43.7 | 16.0 | 62.6 | 87 | |
| Frequency 95% (Hz) | 62.6 | 180.4 | 45.4 | 313.0 | 87 | |
| Bandwidth 90% (Hz) | 0.0 | 136.8 | 53.2 | 281.7 | 87 | |
| Downsweeping Sound (DS) | ||||||
| Duration (ms) | 120.0 | 337.1 | 164.2 | 1241.0 | 58 | |
| Peak frequency (Hz) | 45.8 | 216.6 | 70.3 | 427.2 | 86 | |
| Frequency 5% (Hz) | 45.8 | 140.0 | 34.8 | 198.4 | 86 | |
| Frequency 95% (Hz) | 244.1 | 319.4 | 59.6 | 442.5 | 86 | |
| Bandwidth 90% (Hz) | 61.0 | 179.4 | 72.3 | 381.4 | 86 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Desiderà, E.; Mazzoldi, C.; Navone, A.; Panzalis, P.; Gervaise, C.; Guidetti, P.; Di Iorio, L. Reproductive Behaviours and Potentially Associated Sounds of the Mottled Grouper Mycteroperca rubra: Implications for Conservation. Diversity 2022, 14, 318. https://doi.org/10.3390/d14050318
Desiderà E, Mazzoldi C, Navone A, Panzalis P, Gervaise C, Guidetti P, Di Iorio L. Reproductive Behaviours and Potentially Associated Sounds of the Mottled Grouper Mycteroperca rubra: Implications for Conservation. Diversity. 2022; 14(5):318. https://doi.org/10.3390/d14050318
Chicago/Turabian StyleDesiderà, Elena, Carlotta Mazzoldi, Augusto Navone, Pieraugusto Panzalis, Cédric Gervaise, Paolo Guidetti, and Lucia Di Iorio. 2022. "Reproductive Behaviours and Potentially Associated Sounds of the Mottled Grouper Mycteroperca rubra: Implications for Conservation" Diversity 14, no. 5: 318. https://doi.org/10.3390/d14050318
APA StyleDesiderà, E., Mazzoldi, C., Navone, A., Panzalis, P., Gervaise, C., Guidetti, P., & Di Iorio, L. (2022). Reproductive Behaviours and Potentially Associated Sounds of the Mottled Grouper Mycteroperca rubra: Implications for Conservation. Diversity, 14(5), 318. https://doi.org/10.3390/d14050318