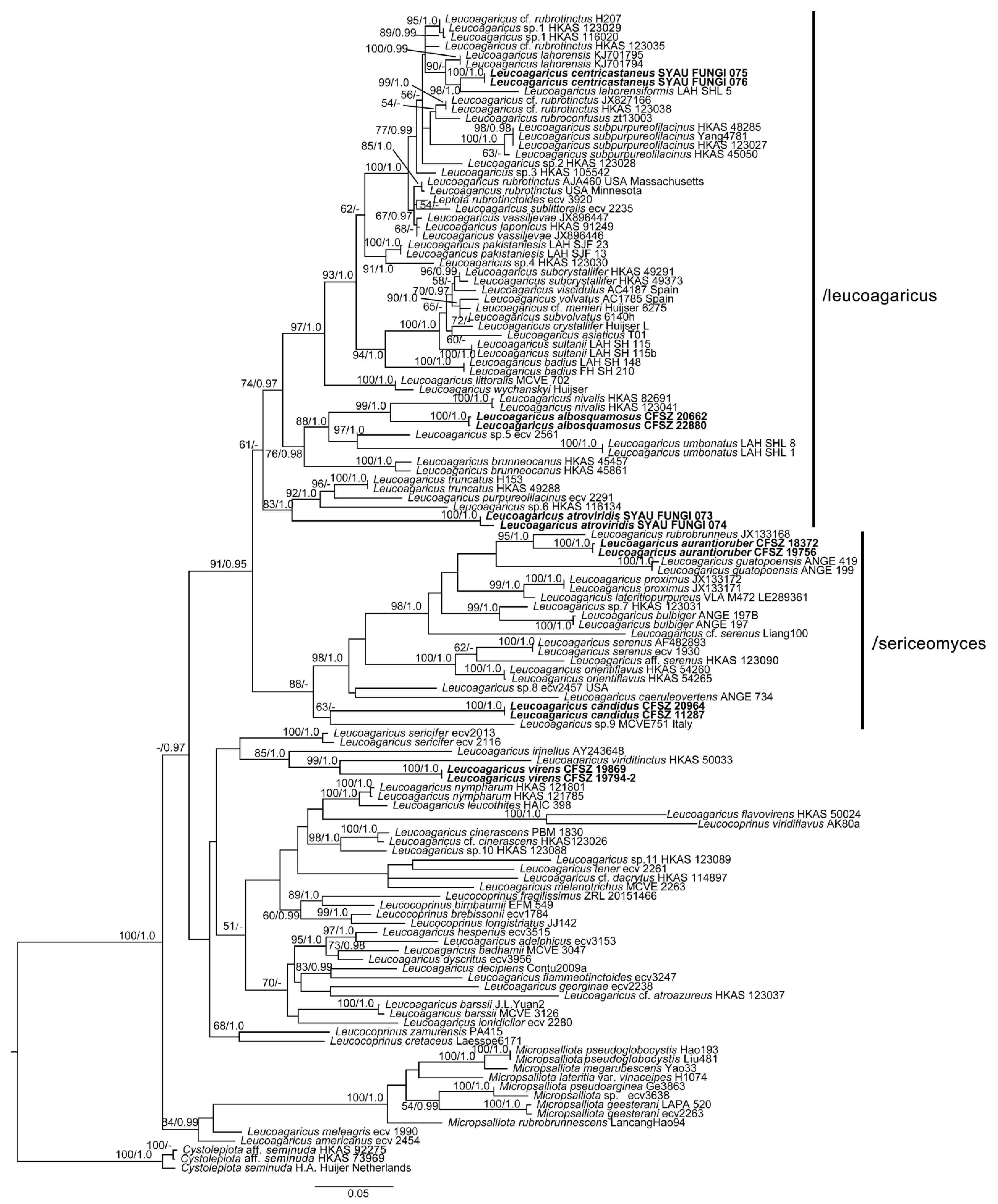
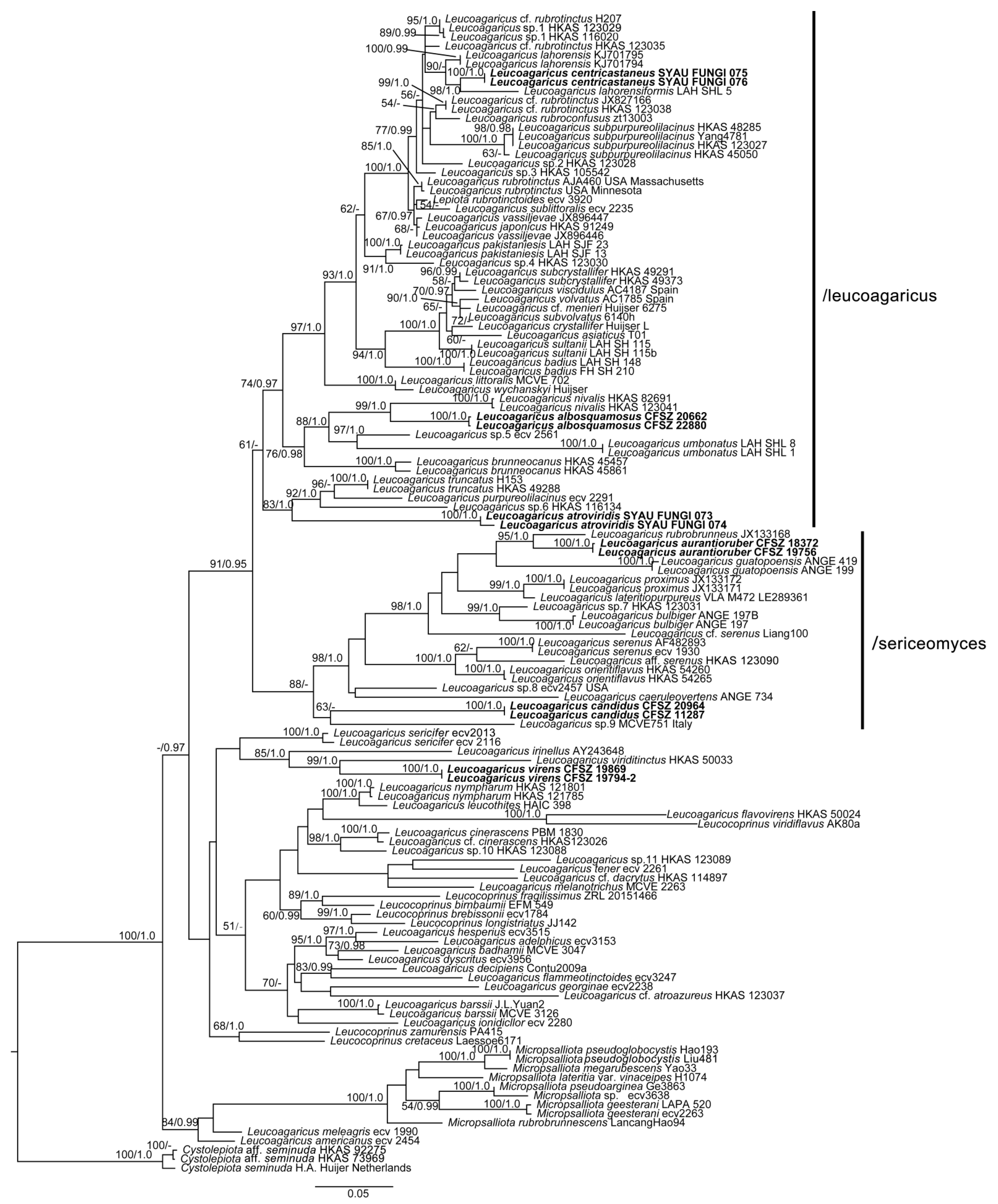
3.2. Taxonomy
Figure 2.
Basidiomata of Leucoagaricus albosquamosus. (A,B): CFSZ 20662; (C): CFSZ 22880; (D): CFSZ 22890). Scale bars: 1 cm.
Figure 2.
Basidiomata of Leucoagaricus albosquamosus. (A,B): CFSZ 20662; (C): CFSZ 22880; (D): CFSZ 22890). Scale bars: 1 cm.
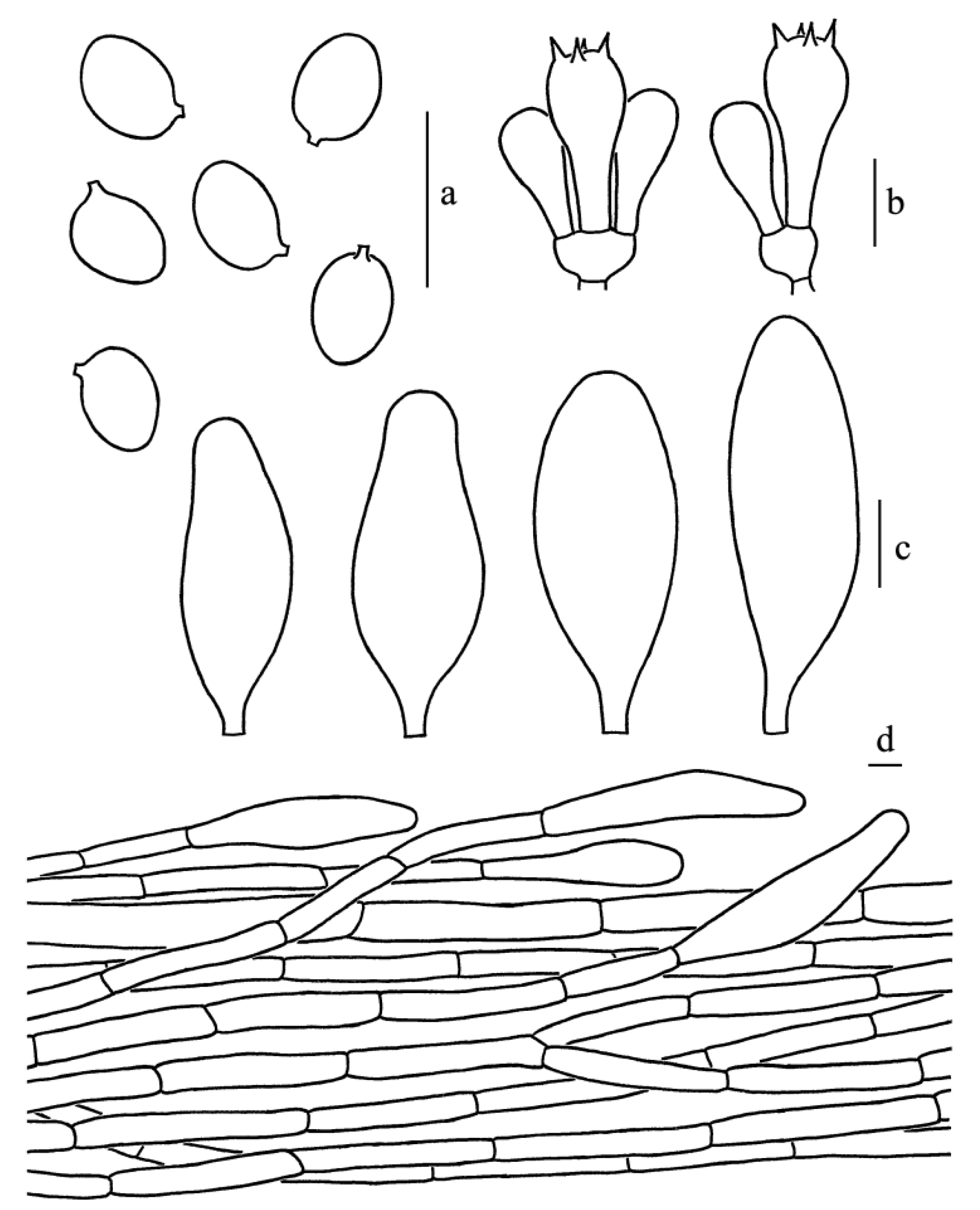
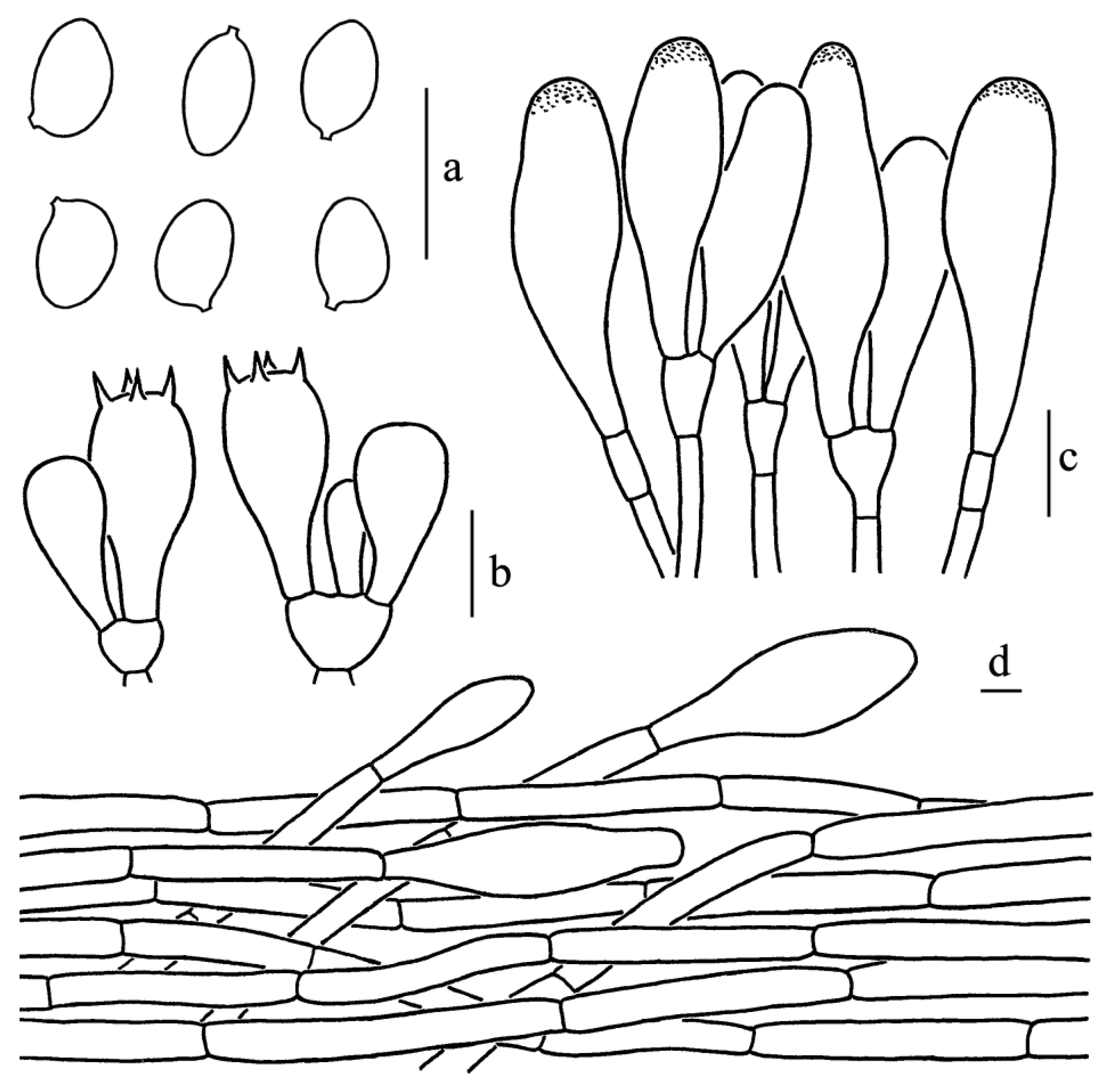
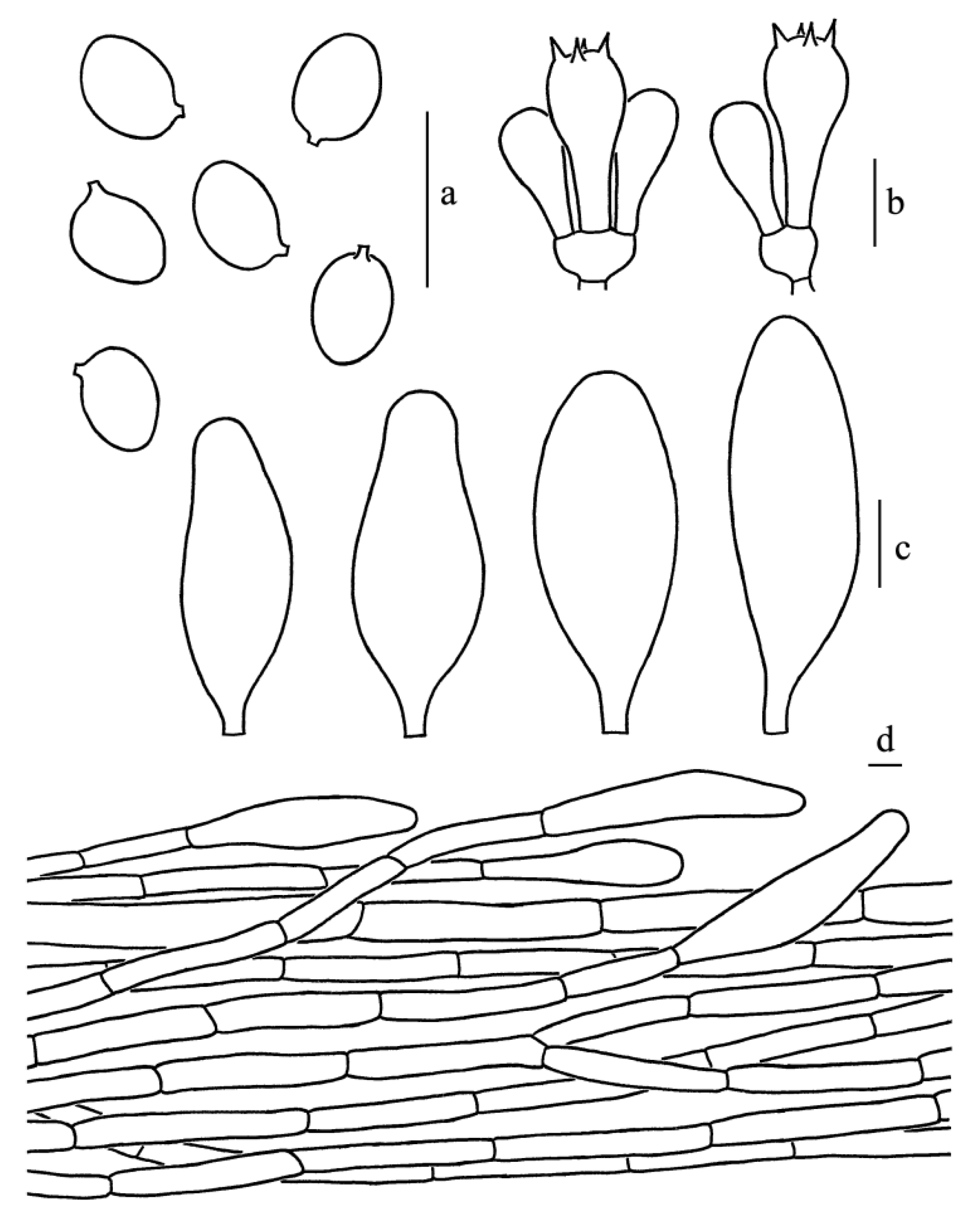
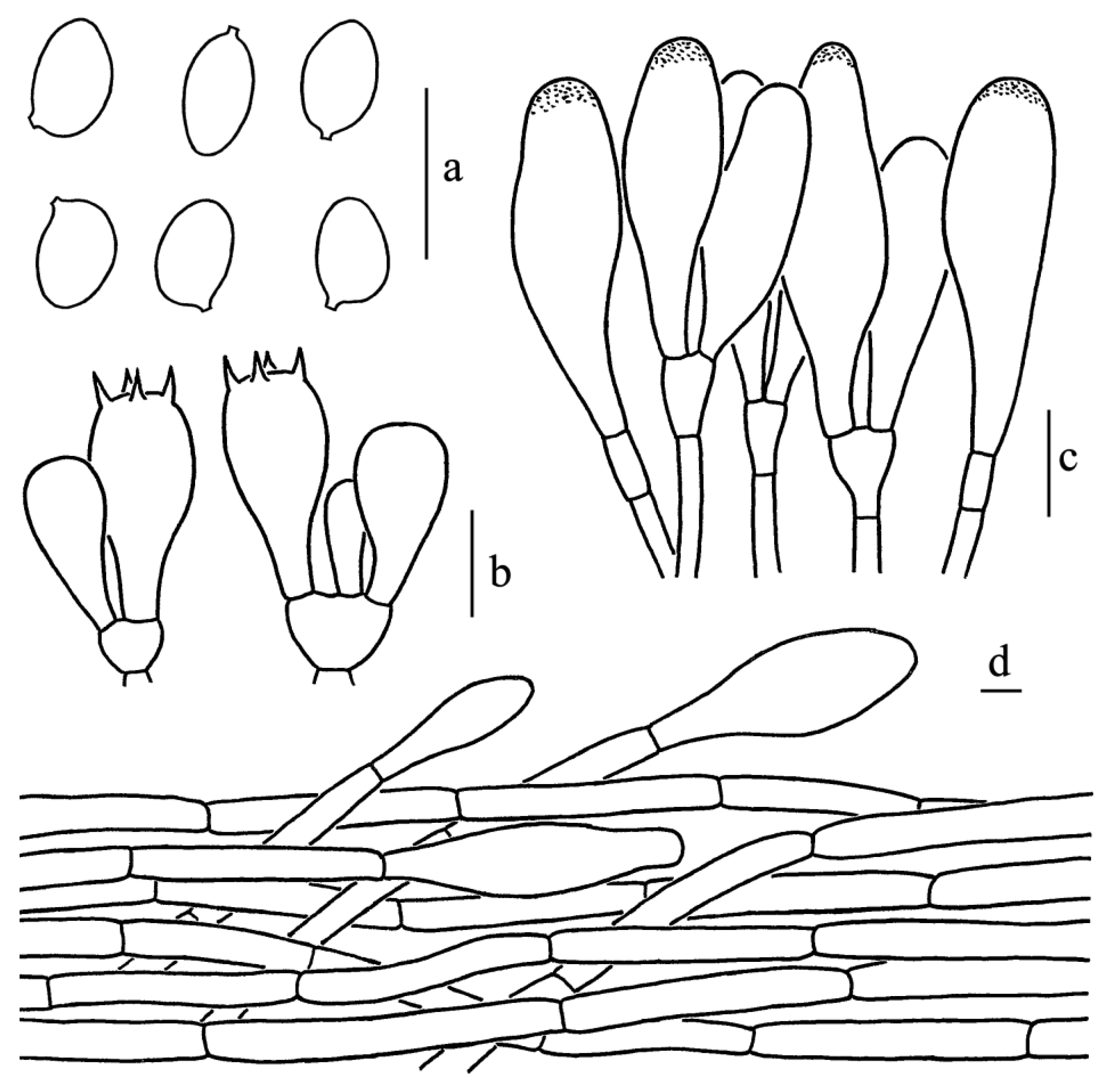
Figure 3.
Leucoagaricus albosquamosus (CFSZ20662). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Figure 3.
Leucoagaricus albosquamosus (CFSZ20662). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Diagnosis: Leucoagaricus albosquamosus is characterized by its white and somewhat plicate pileus with light brownish center, whitish squamules, broadly ellipsoid to ellipsoid basidiospores, subclavate to subfusiform cheilocystidia and squamules on the pileus composed of hyaline, septate cylindrical hyphae measuring 6–13.5 μm in diameter.
Type: CHINA. Inner Mongolia Autonomous Region: Xibo River Green Belt, Songshan District, Chifeng City, China, 12 August 2019, T.Z. Liu (CFSZ 20662, holotype).
GenBank Acc. No.: ITS = OM976879, LSU = OM976865
Description: Pileus 19–36 mm in diameter; center light brownish (3B3–3B4); somewhat plicate; surface, white (1A1); squamules, white (1A1) to whitish, slightly upturned when mature. Context, white (1A1); no change in color upon bruising. Lamellae, white (1A1), free, crowded, 2–4 mm broad, with lamellulae in 1–2 tiers, with entire edges. Stipe 34–46 × 3–5 mm, white (1A1), subcylindric, hollow, nearly smooth. Annulus, white (1A1), simple, membranous, located in the middle to upper part of the stipe. Flavor and odor not recorded. Spore print, white.
Basidiospores [65/14/3], 5.0–6.5(7.0) × 4.0–5.0(5.5) μm, Q = (1.11)1.22–1.44(1.50), Qm = 1.31 ± 0.08, broadly ellipsoid to ellipsoid from side view and front view, hyaline, without germ pore, slightly thick-walled (about 0.5 μm), dextrinoid, metachromatic. Basidia, 19.0–23.0 × 8.5–9.5 μm, clavate, hyaline, thin-walled, four-spored, rarely two-spored; sterigmata, 2.5 μm long. Cheilocystidia, 35.0–47.0 × 12.5–19.0 μm, subclavate to subfusiform, hyaline. Pleurocystidia, absent. Lamellar trama, trabecular. Squamules on the pileus composed of septate cylindrical hyphae 6–13.5 μm in diameter, hyaline. Stipitipellis composed of hyaline, cylindrical hyphae, 5.0–12.0 μm in diameter. Clamp connections absent in all parts of the basidiomata.
Habitat and Habit: scattered on soil in summer. So far, known only known in Chifeng, Inner Mongolia Autonomous Region.
Other specimens examined: CHINA. Inner Mongolia Autonomous Region: Xibo River Green Belt, Songshan District, Chifeng City, China, 18 August 2020, T.Z. Liu (CFSZ 22880). Inner Mongolia Autonomous Region: Stone park, Songshan District, Chifeng City, 19 August 2020, T.Z. Liu (CFSZ 22890).
Comments: The somewhat plicate pileus of
La. albosquamosus may suggest that this species be placed in
Leucocoprinus. Considering
La. albosquamosus is nested within a strongly supported clade that corresponds to section
Leucoagaricus, we placed this species within
Leucoagaricus rather than
Leucocoprinus. According to phylogenetic analysis (
Figure 1),
Leucoagaricus albosquamosus is a sister taxon to
Leucoagaricus nivalis (W.F. Chiu) Z. W. Ge & Zhu L. Yang. Together, these two taxa form a sister clade to a clade jointly formed by
Leucoagaricus umbonatus S. Hussain, H. Ahmad & Afshan and an undescribed species of
Leucoagaricus [
19,
30,
31].
Leucoagaricus nivalis, originally described in Yunnan, southwestern China, resembles
La. albosquamosus in its overall white, small basidiomata and subclavate cheilocystidia. However,
La. nivalis is a white species with narrower amygdaliform basidiospores measuring 6.0–7.5(9.0) × (3.0)3.5–4.5(5.0) μm and glabrous to radially fibrillose white pileus [
31].
Leucoagaricus umbonatus, a white species originally described in Pakistan, is similar to
La. albosquamosus in the similarly bulbous stipe base, similarly shaped basidiospores and similarly sized clavate basidia. However,
La. umbonatus has smaller ventricose to fusiform cheilocystidia (24–29 × 8–12 μm) and cutis of radially arranged white fibrils [
19].
Leucoagaricus sericifer (Locq.) Vellinga is also similar to
La. albosquamosus in its white pileus and the shape of the basidiospores. However,
La. sericifer has oblong amygdaliform to cylindrical amygdaliform basidiospores, as well as more or less lageniform cheilocystidia [
6]. Considering the somewhat plicate pileus with squamules,
La. albosquamosus is somewhat similar to
Leucocoprinus cepistipes (Sowerby) Pat. However,
Leucocoprinus cepistipes has small beige–brown to ochraceous brown squamules and lageniform to clavate cheilocystidia, often with apical excrescence [
6].
Figure 4.
Basidiomata of Leucoagaricus atroviridis. (A,B) SYAU FUNGI 073; (C,D) SYAU FUNGI 074. Scale bars: 1 cm.
Figure 4.
Basidiomata of Leucoagaricus atroviridis. (A,B) SYAU FUNGI 073; (C,D) SYAU FUNGI 074. Scale bars: 1 cm.
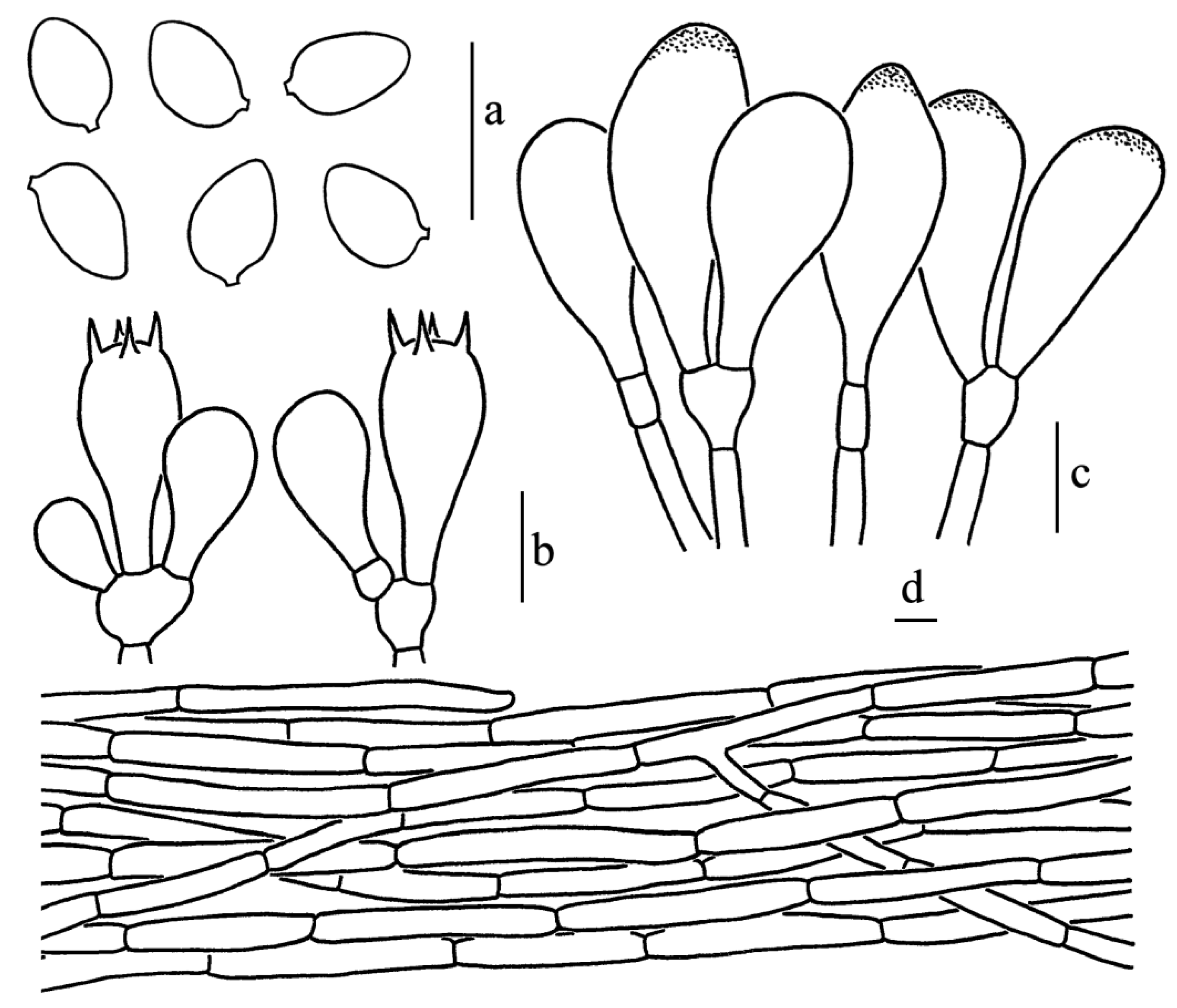
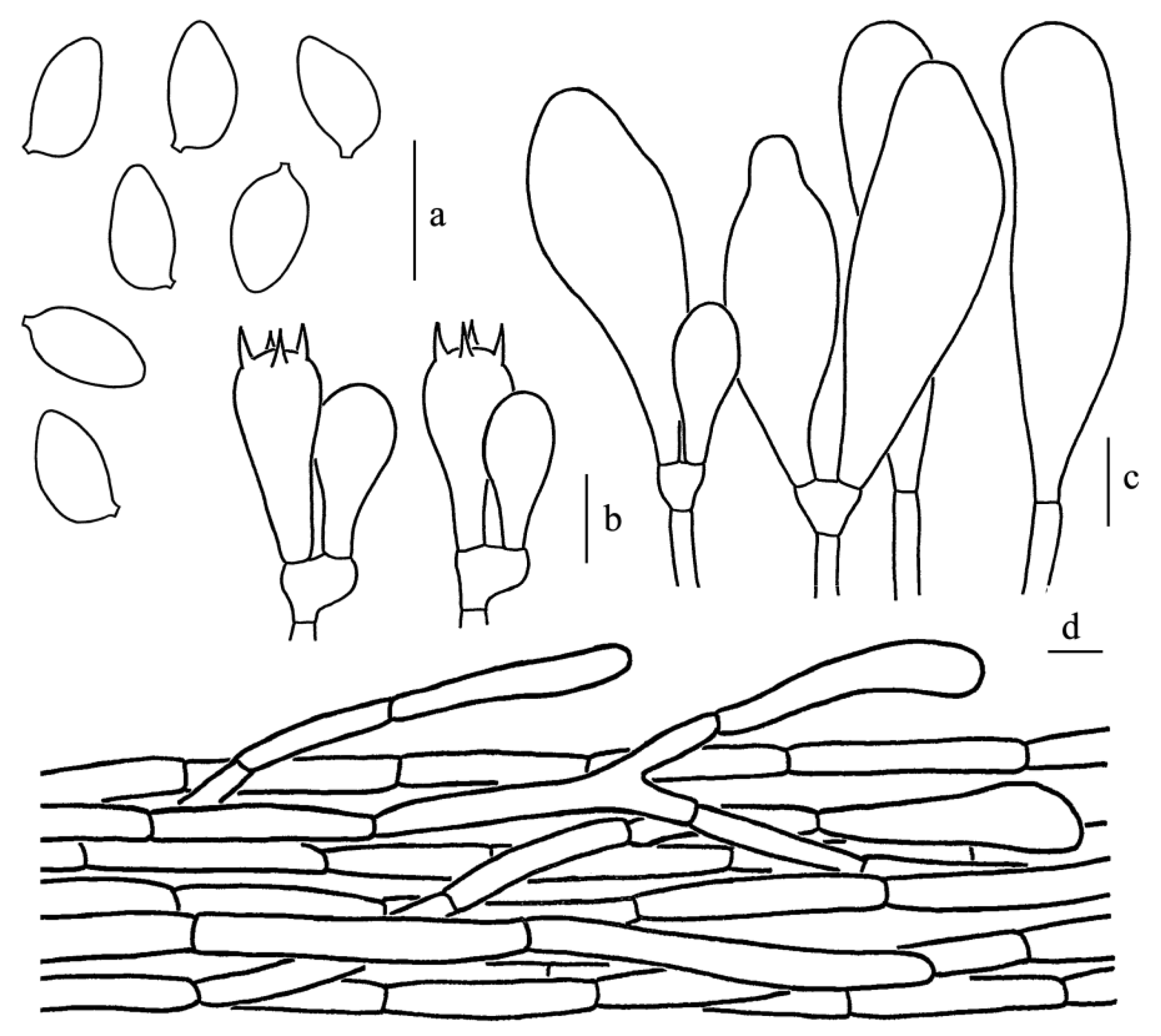
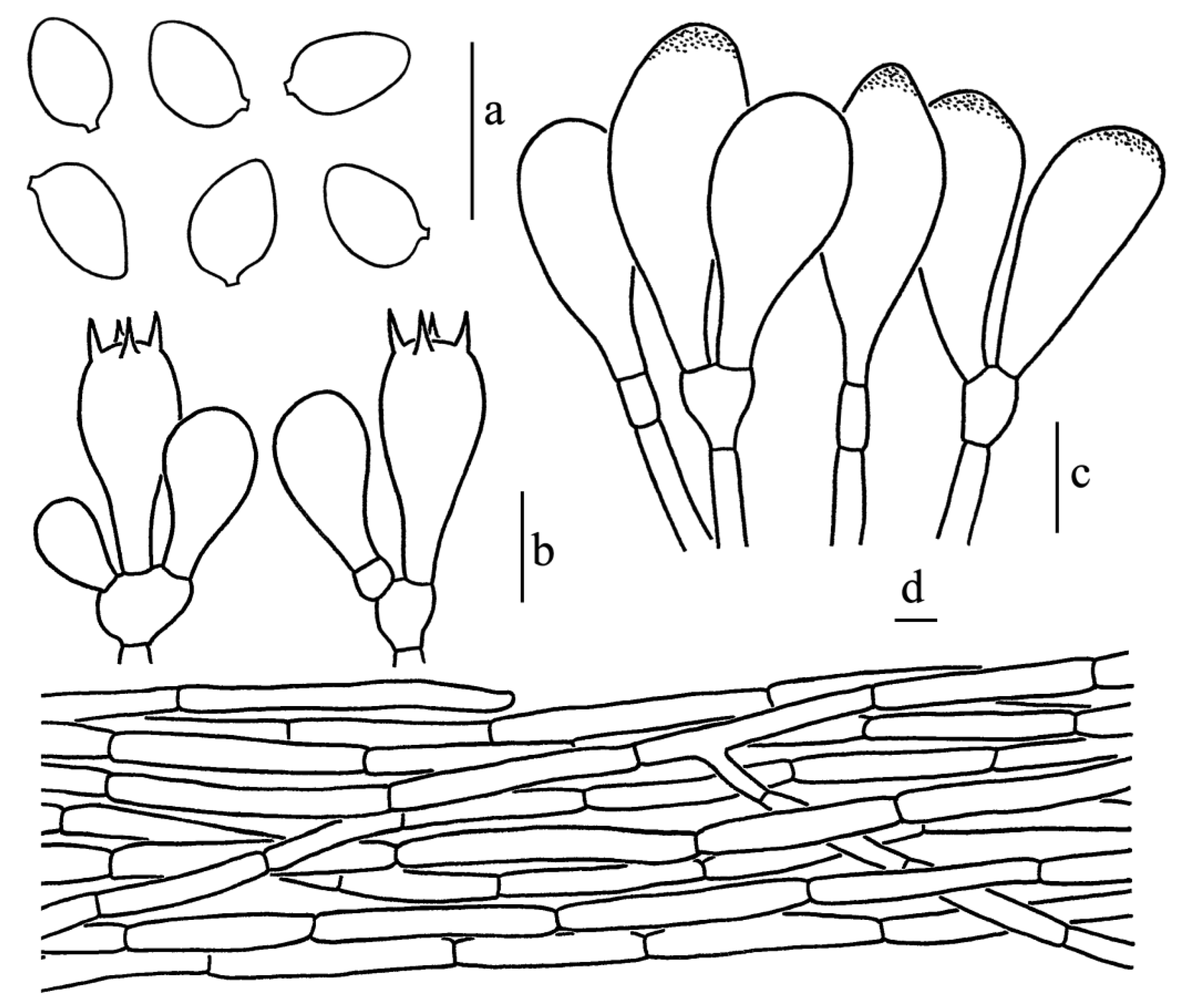
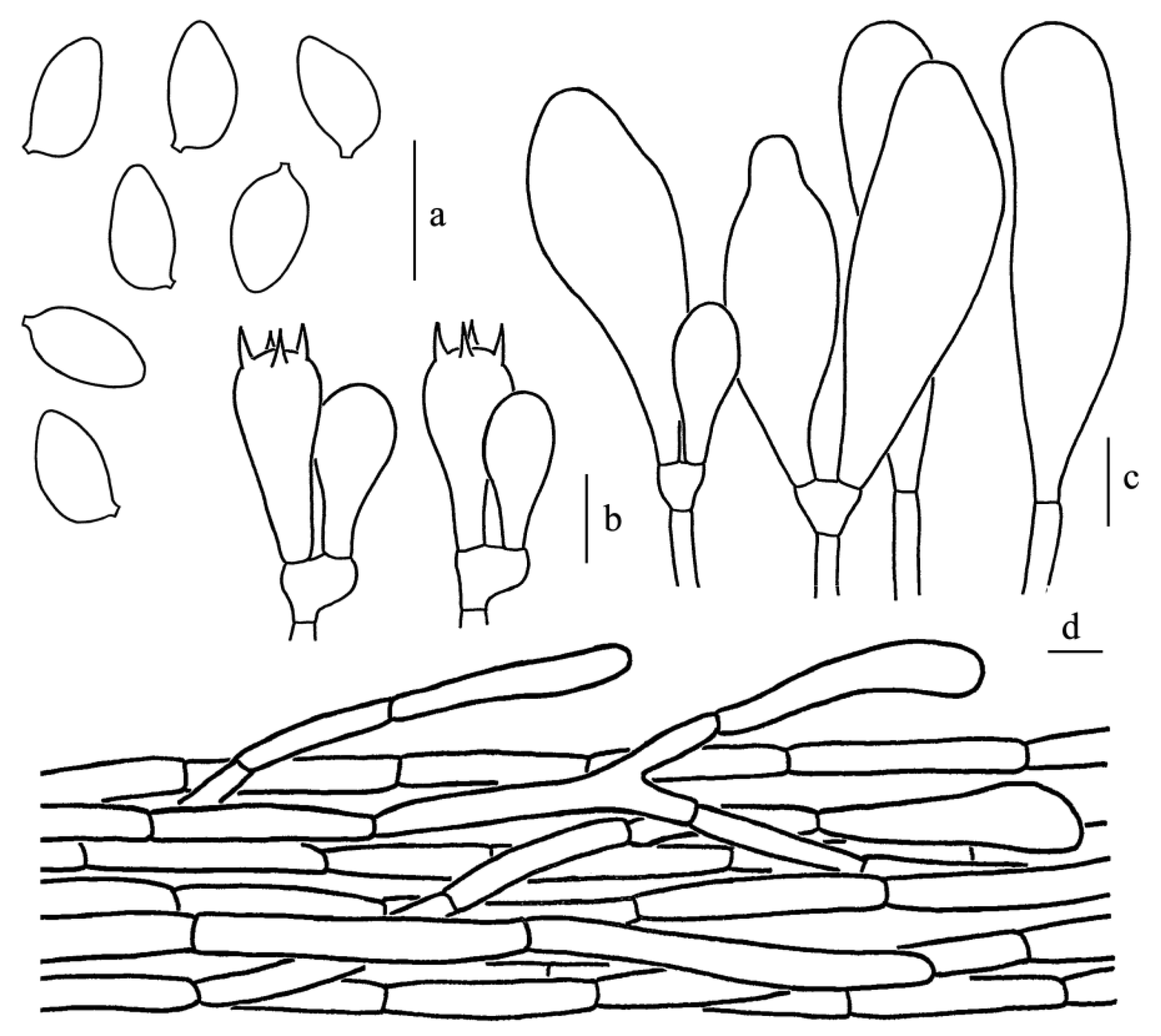
Figure 5.
Leucoagaricus atroviridis (SYAU FUNGI 073). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Figure 5.
Leucoagaricus atroviridis (SYAU FUNGI 073). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Diagnosis: Leucoagaricus atroviridis is characterized by bottle-green to dark brown, fibrillose squamules, and broadly clavate cheilocystidia with obvious refractive contents.
Type: CHINA. Liaoning Province: Shenyang, Shenyang Agricultural University, August 2020, X.D. Yu (SYAU FUNGI 073, holotype).
GenBank Acc. No.: ITS = OM976852, LSU = OM976868
Description: Pileus 48–114 mm in diameter, covered with bottle-green (1E7) to dark brown (2F6) radially fibrillose squamules, umbonate. Context, white (1A1), up to 4 mm thick at center, no change in color when bruised. Lamellae-free, crowded, white (1A1), edge finely scalloped. Stipe, 69–73 × 6–10 mm, white (1A1), smooth, hollow, subcylindrical or attenuate upwards with bulbous base up to 13–16 mm wide. Annulus, white (1A1), located at the middle to upper part of the stipe. Flavor and odor not recorded. Spore print not recorded.
Basidiospores [60/3/2], 5.5–7.0(7.5) × 4.0–5.0 μm, Q = (1.10)1.22–1.50(1.63), Qm = 1.36 ± 0.11, ellipsoid to ovoid from side view and front view, without germ pore, hyaline, smooth, slightly thick-walled (about 0.5 μm); dextrinoid, metachromatic. Basidia, 17.0–28.0 × 7.5–10.0 μm, clavate, hyaline, four-spored, rarely two-spored; sterigmata, 3.5 μm long. Cheilocystidia, 21.5–33.5 × 9.5–15.5 μm, hyaline, broadly-clavate to broadly fusiform, thin-walled. Some cheilocystidia with obvious refractive contents on the apex. Pleurocystidia absent. Lamellar trama, subregular. Pileus squamules composed of 5.0–8.5 μm wide subcylindrical repent hyphae with light greenish (30B5) to light brownish (3B3) intracellular pigments. Stipitipellis composed of cylindrical to slightly inflated hyphae, 5.0–11.0 μm in diameter, hyaline. Clamp connections absent in all parts of the basidiomata.
Habitat and Habit: solitary in grass in summer. So far, known only in Shenyang, Liaoning Province in northeastern China.
Other specimens examined: CHINA. Liaoning Province: Shenyang, Shenyang Agricultural University, August 2020, X.D. Yu (SYAU FUNGI 074).
Comments: Macroscopically, Leucoagaricus atroviridis is characterized by medium to large basidiomata, stipe with a bulbous base and bottle-green to dark brown fibrils.
Phylogenetically,
Leucoagaricus atroviridis is close to
Leucoagaricus truncatus Z. W. Ge & Zhu L. Yang and
Leucoagaricus purpureolilacinus Huijsman (
Figure 1). However,
La. purpureolilacinus, originally described in the Netherlands, has a purplish brown pileus and clavate to subfusiform cheilocystidia [
6].
La. truncatus, originally described in Sichuan, southwestern China, differs in having a truncated pileus with orange–white to gray–orange squamules and larger basidiospores [
11].
Figure 6.
Basidiomata of Leucoagaricus aurantioruber. (A,B) CFSZ 18372; (C–E) CFSZ 19756). Scale bars: 1 cm.
Figure 6.
Basidiomata of Leucoagaricus aurantioruber. (A,B) CFSZ 18372; (C–E) CFSZ 19756). Scale bars: 1 cm.
Figure 7.
Leucoagaricus aurantioruber (CFSZ 19756). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Figure 7.
Leucoagaricus aurantioruber (CFSZ 19756). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Diagnosis: Leucoagaricus aurantioruber is characterized by pileus with minute brown to orange–reddish, radially arranged fibrillose squamules; ellipsoid to ovoid basidiospores; subfusiform cheilocystidia with tiny crystals; and squamules composed of branched, thin-walled hyphae.
Type: CHINA. Inner Mongolia Autonomous Region: Sandaohe, Heilihe National Nature Reserve, Ningcheng County, Chifeng City, 19 August 2018, T.Z. Liu, L. Zhang, Y. Bai (CFSZ 19756, holotype).
GenBank Acc. No.: ITS = OM976875, LSU = OM976863
Description: Pileus 15–24 mm in diameter; at first, nearly hemispherical, umbonate. Surface, orange–reddish (6B7) to orange–brown (6C8), covered with minute brown (6C6) to light orange–reddish (6A4) radially fibrillose squamules; margin, often radially striate; context, white (1A1), up to 3 mm thick at center; no change in color when bruised. Lamellae-free, crowded, white (1A1), thin, 1.5–2 mm wide, with 2–3 tiers of lamellulae, edge finely scalloped. Stipe 33–42 × 2–5 mm, hollow, clavate, attenuate upward, white (1A1), smooth above annulus, slightly fibrillose in lower part. Annulus simple, white (1A1), located in the middle to upper part of the stipe, membranous. Flavor and odor not recorded. Spore print not recorded.
Basidiospores [40/3/2], 6.0–7.5(8.0) × (3.5)4.0–5.0 μm, Q = (1.20)1.30–1.75(2.14), Qm = 1.54 ± 0.15, ellipsoid to ovoid from side view, ellipsoid from the front view, without germ pore, hyaline, smooth, slightly thick-walled, dextrinoid, metachromatic. Basidia, 16.5–22.0 × 8.0–11.0 μm, clavate, hyaline, four-spored; sterigmata, 4 μm long. Cheilocystidia, 34.5–41.0 × 10.0–14.5 μm, subfusiform, often with a terminal appendage, apex with tiny crystals. Pleurocystidia, absent. Lamellar trama, trabecular. Pileus squamules composed of branched, thin-walled hyphae, 5.0–9.5 μm in diameter. Stipitipellis composed of cylindrical hyphae, 5.5–12.5 μm in diameter, hyaline. Clamp connections absent in all parts of the basidiomata.
Habitat and Habit: solitary to scattered on soil in summer.
Other specimens examined: CHINA. Inner Mongolia Autonomous Region: Sandaohe, Heilihe National Nature Reserve, Ningcheng County, Chifeng City, 21 August 2017, T.Z. Liu (CFSZ 18372).
Comments:
Leucoagaricus aurantioruber and
Leucoagaricus rubrotinctus (Peck) Singer are very similar in having orange–reddish pileus. However,
La. aurantioruber has ellipsoid to ovoid basidiospores and subfusiform cheilocystidia with tiny crystals on the apex [
32].
La. rubrobrunneus E.F. Malysheva, Svetash. & Bulakh, originally described in the Russian far east, is similar to
La. aurantioruber in having subfusiform cheilocystidia, forming a sterile lamellae edge. However,
La. aurantioruber is distinctly different from
La. rubrobrunneus in having larger basidiospores and larger pileus (15–24 mm), pileus squamules composed of branched, thin-walled hyphae measuring 5.0–9.5 μm in diameter and cheilocystidia with tiny crystals on the apex [
33].
La. bulbiger Justo, Bizzi & Angelini and
La. proximus are also similar to
La. aurantioruber in having similarly shaped basidiospores and pileus squamules. However,
La. bulbiger, originally described in the Dominican Republic, has a yellow-–orange pileus, a bulbous stipe base and polymorphic cheilocystidia [
21].
La. proximus E.F. Malysheva, Svetash. & Bulakh, originally described from the Russian far east, has smaller-sized cheilocystidia measuring 27.0–40.0 × 6.0–14.0 μm [
33].
Leucoagaricus rubroconfusus Migl. & Coccia resembles La. aurantioruber in having orange fibrillose squamules and ellipsoid to ovoid basidiospores of approximately the same length. However, La. rubroconfusus has smaller cheilocystidia without crystals on the apex.
The new species is also similar to
La. japonicus (Kawam. ex Hongo) Hongo in having ellipsoid or ovoid basidiospores. However,
La. japonicus has testaceous, scaly pileus; longer basidiospores; and clavate to fusiform cheilocystidia without crystals [
5,
32,
34].
Figure 8.
Basidiomata of Leucoagaricus candidus. (CFSZ 20964). Scale bars: 1 cm.
Figure 8.
Basidiomata of Leucoagaricus candidus. (CFSZ 20964). Scale bars: 1 cm.
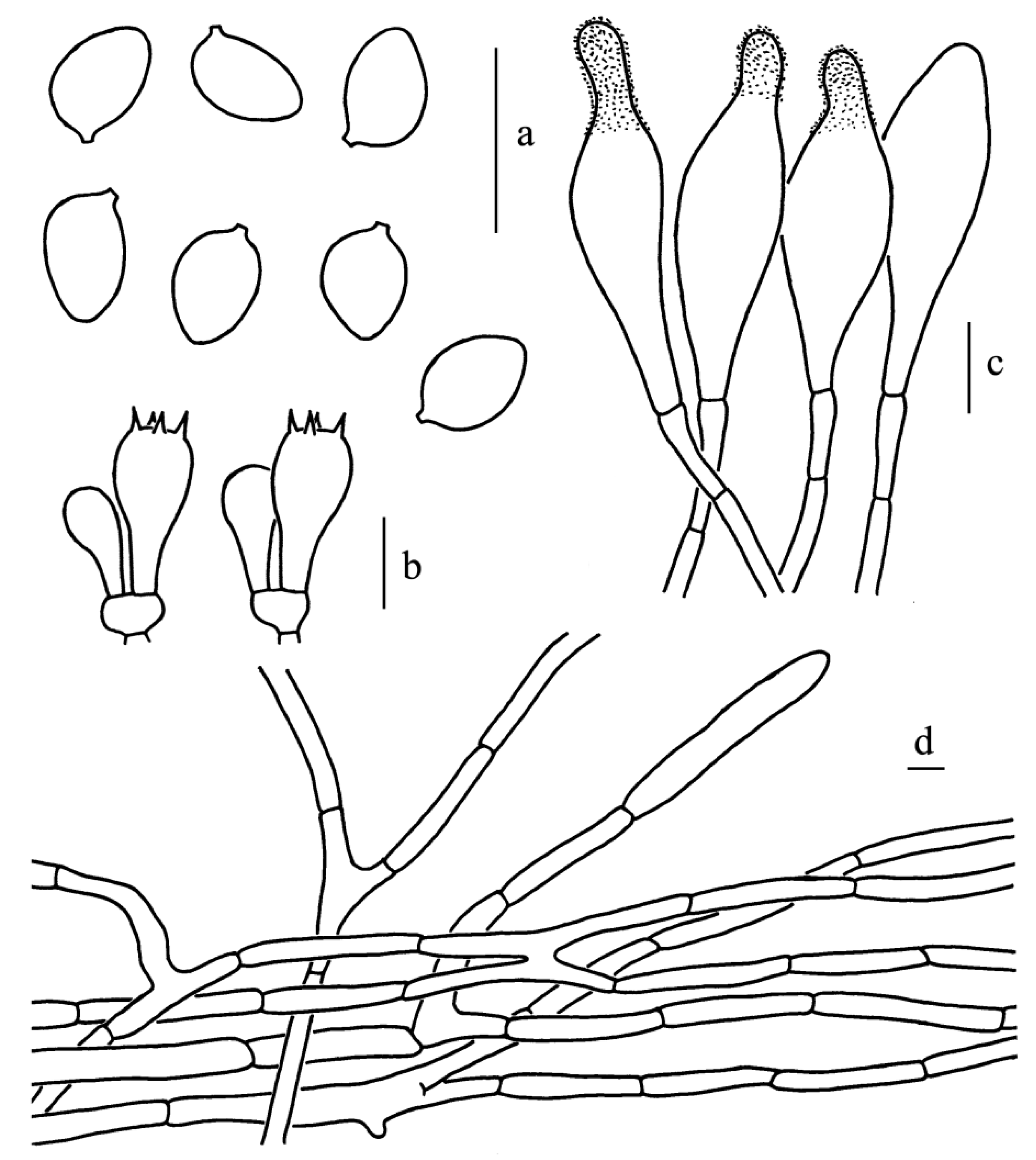
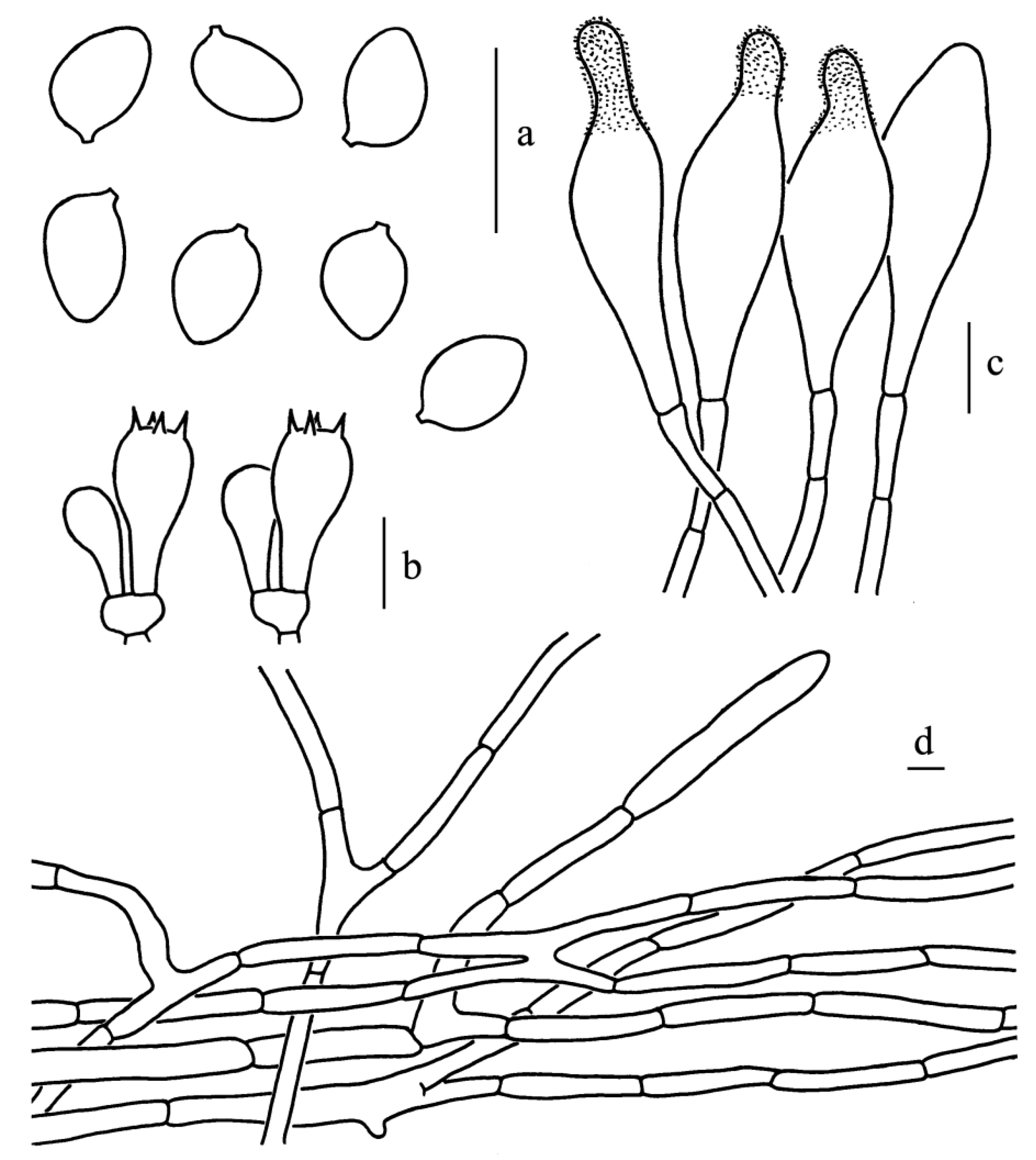
Figure 9.
Leucoagaricus candidus (CFSZ 20964). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus surface. Scale bars: 10 μm.
Figure 9.
Leucoagaricus candidus (CFSZ 20964). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus surface. Scale bars: 10 μm.
Diagnosis: Leucoagaricus candidus is characterized by its white basidiomata, white pileus squamules, a margin often with appendiculate white veil remnants and ellipsoid to broadly ellipsoid basidiospores.
Type: CHINA. Inner Mongolia Autonomous Region: Sidaogou, Heilihe National Nature Reserve, Ningcheng County, Chifeng City, 21 August 2019, T.Z. Liu, K. Kang (CFSZ 20964, holotype).
GenBank Acc. No.: ITS = OM976877, LSU = OM976864
Description: Pileus 15–31 mm in diameter, white (1A1), ovoid when young, becoming convex to applanate with age, glabrous to radially fibrillose, margin often with fine striations when mature, appendiculate with veil remnants. Context: white (1A1), thin, no change in color upon bruising. Lamellae-free, crowded, white (1A1), up to 3 mm in height, edge finely scalloped. Stipe, 27–38 × 3–7 mm, subcylindrical, white (1A1), clavate, hollow, white (1A1) to milk white (1A2), smooth to silky above annulus, with white, minute or fibrillose squamules below annulus. Annulus white (1A1), membranous, located in the middle to upper part of the stipe. Flavor and odor unknown. Spore print not recorded.
Basidiospores [62/5/3], (5.5)6.0–7.5(8.0) × (4.0)4.5–5.0 μm; Q = (1.20)1.33–1.63(1.78), Qm = 1.46 ± 0.13; ellipsoid in side view, occasionally broadly ellipsoid or elongate; ellipsoid in front view; without germ pore, hyaline, slightly thick-walled (about 0.5 μm); dextrinoid, metachromatic in Cresyl Blue. Basidia, 16.0–20.5 × 7.0–9.5 μm, clavate, hyaline, four-spored. Cheilocystidia, 29.0–42.0 × 8.0–12.5 μm, mostly narrowly clavate, occasionally with a constricted apex. Pleurocystidia, absent. Lamellar trama, trabecular. Pileus surface composed of 5.0–13.0 μm wide repent-hyphae, subcylindrical, thin-walled, hyaline. Stipitipellis composed of subcylindric hyphae, 4.0–8.5 μm in width with yellowish intracellular pigment. Clamp connections absent in all parts of the basidiomata.
Habitat and Habit: solitary to scattered on soil in summer.
Other specimens examined: CHINA. Inner Mongolia Autonomous Region: Daheishan National Nature Reserve, Aohan Banner, Chifeng City, 22 August 2016, T.Z. Liu, S.L. Zhang, Y.X. Yang (CFSZ 11287). Inner Mongolia Autonomous Region: Sidaogou, Heilihe National Nature Reserve, Ningcheng County, Chifeng City, 22 August 2019, T.Z. Liu, T. Li (CFSZ 20991).
Comments:
Leucoagaricus candidus is morphologically similar to
Leucoagaricus nivalis,
Leucoagaricus serenus (Fr.) Bon & Boiffard and
Leucoagaricus sericifer on the overall whitish to white basidiomata. However,
La. candidus differs from them by having ellipsoid to broadly ellipsoid basidiospores and a pileus surface composed of wider hyphae (5.0–13.0 μm). In addition, both
La. serenus and
La. sericifer were originally described in Europe and are widespread in Europe [
6,
31,
35].
Two white European species,
La. subvolvatus (Malencon & Bertault) Bon and
La. menieri (Sacc.) Singer, are also similar to
La. candidus in the overall white basidiomata and similar basidiospores. However,
La. subvolvatus has white to cream pileus and cheilocystidia with crystals at the apex [
32];
La. menieri has fragile and sericeous pileus and cheilocystidia with crystals at the apex [
32].
Figure 10.
Basidiomata of Leucoagaricus centricastaneus. (A) SYAU FUNGI 072; (B,C) SYAU FUNGI 075; (D) SYAU FUNGI 076). Scale bars: 1 cm.
Figure 10.
Basidiomata of Leucoagaricus centricastaneus. (A) SYAU FUNGI 072; (B,C) SYAU FUNGI 075; (D) SYAU FUNGI 076). Scale bars: 1 cm.
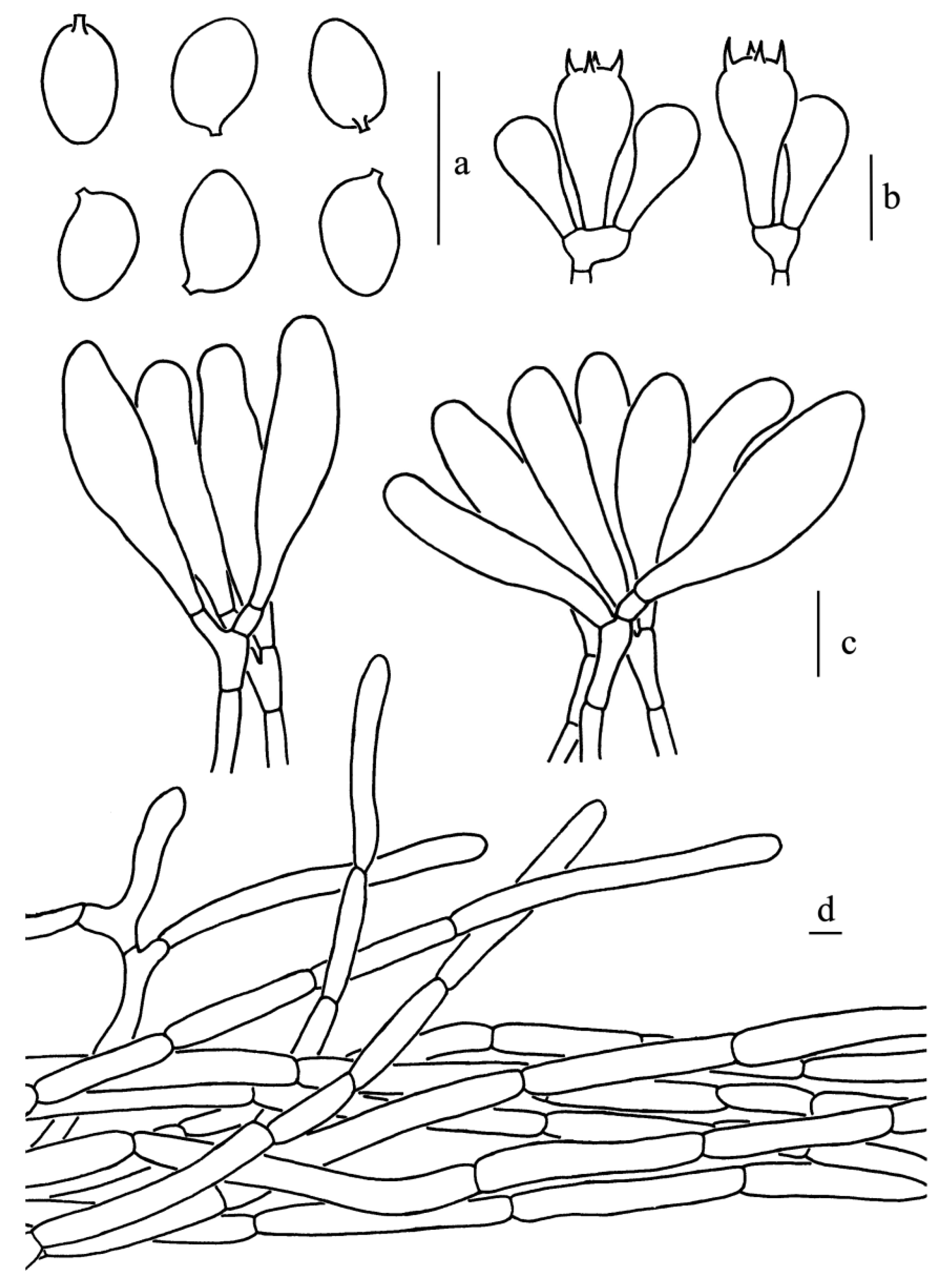
Figure 11.
Leucoagaricus centricastaneus (SYAU FUNGI 076). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Figure 11.
Leucoagaricus centricastaneus (SYAU FUNGI 076). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Diagnosis: Leucoagaricus centricastaneus is characterized by its pileus with orange–brown, yellow–brown or red–brown floccose squamules, ovoid to ellipsoid basidiospores and clavate to fusiform cheilocystidia with refractive contents on the apex.
Type: CHINA. Liaoning Province: Shenyang, Shenyang Agricultural University, August 2020, X.D. Yu (SYAU FUNGI 076, holotype).
GenBank Acc. No.: ITS = OM976855, LSU = OM976871
Description: Pileus, 43–62 mm in diameter; slightly umbonate, covered with orange-brown (5B6), yellow-brown (3B6) or red-brown (9D7–9D8) floccose squamules. The color of the squamules washes out towards the margin. Context, white (1A1); fleshy, no change in color when bruised. Lamellae-free, white (1A1), crowded, up to 3 mm, edge finely scalloped. Stipe, 47–73 × 3–6 mm; white (1A1), nearly smooth or slightly fibrillose, enlarged to subclavate toward base. Annulus, simple; white (1A1), membranous, located in the middle to upper part of the stipe. Flavor and odor not recorded. Spore print not recorded.
Basidiospores, [60/3/2], (5.0)5.5–7.0(7.5) × (4.0)4.5–5.0 μm; Q = (1.20)1.22–1.44(1.50); Qm = 1.32 ± 0.09; ovoid to ellipsoid in side view; ellipsoid in front view; dextrinoid, metachromatic in Cresyl Blue, without germ pore, hyaline, slightly thick-walled (about 0.5 μm). Basidia, 17.5–24.0 × 8.5–10.5 μm; clavate, hyaline, four-spored; sterigmata, 2.5 μm long. Cheilocystidia, 28.5–43.5 × 9.0–15.0 μm; clavate to broadly clavate, at times fusiform, hyaline, forming clusters. Some cheilocystidia have obvious refractive contents at the apex. Pleurocystidia, absent. Lamellar trama, trabecular. Pileus squamules made up of subcylindrical septate hyphae 6.5–10.5 μm in diameter, with terminal elements barely differentiated, narrowly clavate to slightly fusiform (42.0–72.5 × 12.5–24.0 μm). Stipitipellis composed of subcylindrical to long ellipsoid hyphae, 5.5–13.0 μm in diameter, hyaline. Clamp connections absent in all parts of the basidiomata.
Habitat and Habit: solitary in grass in summer. So far, known only in Shenyang, Liaoning Province in northern China.
Other specimens examined: CHINA. Liaoning Province: Shenyang, Shenyang Agricultural University, August 2020, X.D. Yu (SYAU FUNGI 075). Liaoning Province: Shenyang, Shenyang Agricultural University, August 2020, X.D. Yu (SYAU FUNGI 072).
Comments:
Leucoagaricus centricastaneus is very close to
Leucoagaricus lahorensis Qasim, T. Amir & Nawaz and
Leucoagaricus lahorensiformis S. Hussain, H. Ahmad, Afshan & Khalid in the phylogenetic analysis (
Figure 1). All three species have subclavate cheilocystidia. However,
La. lahorensis and
La. lahorensiformis, originally described in Pakistan, have a differently colored pileus (orange, orange–reddish or reddish–brown) and amygdaliform to ellipsoid basidiospores, and the cheilocystidia of
La. lahorensis has no refractive contents on the apex [
19,
36].
Leucoagaricus japonicus,
Leucoagaricus rubrotinctus (Peck) Singer and
Leucoagaricus vassiljevae E.F. Malysheva, Svetash. & Bulakh are similar to
La. centricastaneus in having similarly shaped cheilocystidia. However, the cheilocystidia of these species have no refractive contents on the apex [
32,
33,
34]. In addition,
La. japonicus, originally described in Japan, has a testaceous, scaly pileus; longer cheilocystidia; and amygdaliform basidiospores [
34].
La. rubrotinctus, originally described in the United States, has an orange–reddish pileus and larger amygdaliform basidiospores [
32].
La. vassiljevae, described in the Russian far east, has appressed red–brown or crimson–brown fibrils and broadly clavate basidia [
33].
Figure 12.
Basidiomata of Leucoagaricus virens. (A–C) CFSZ 19869. Scale bars: 1 cm.
Figure 12.
Basidiomata of Leucoagaricus virens. (A–C) CFSZ 19869. Scale bars: 1 cm.
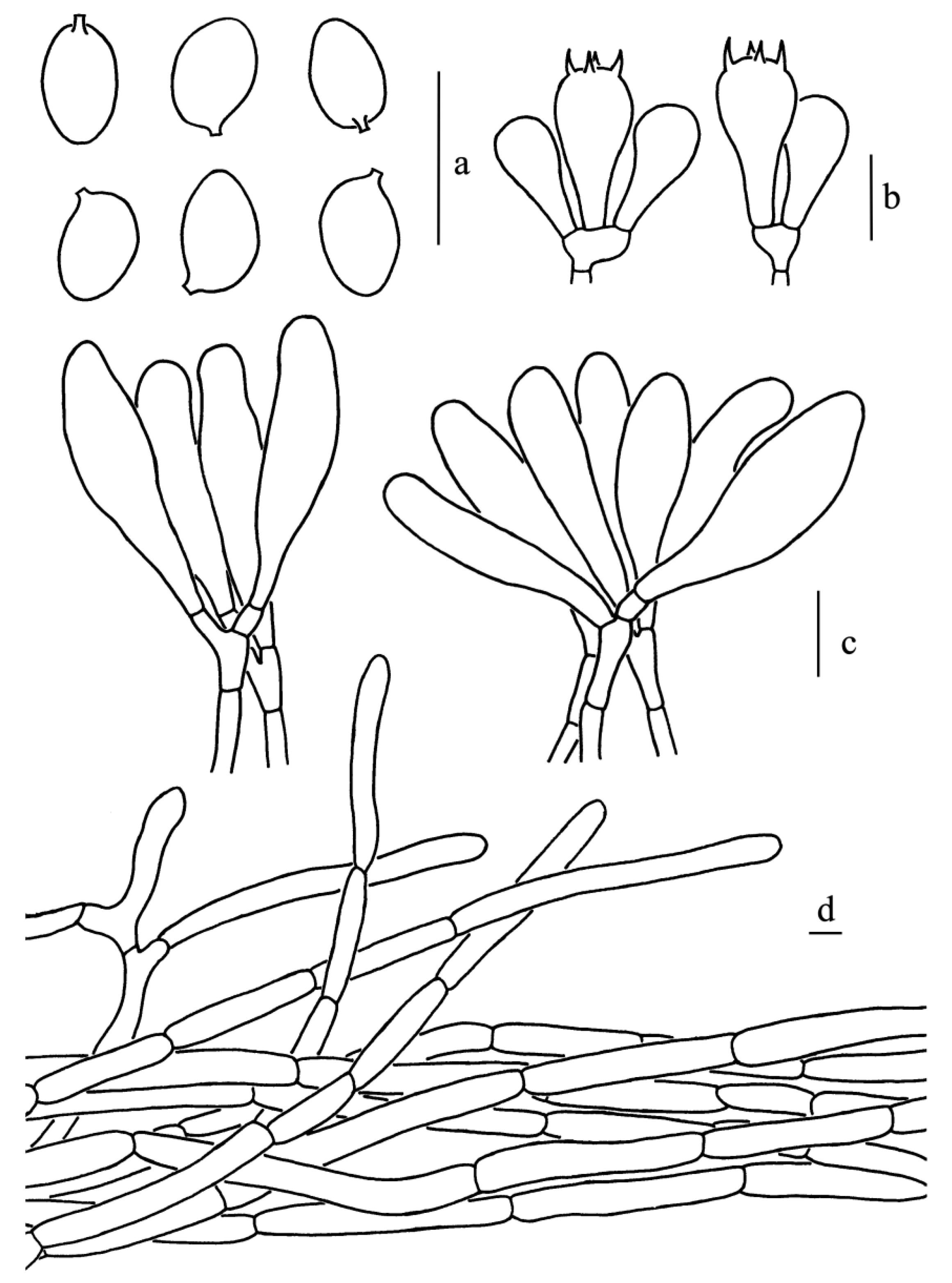
Figure 13.
Leucoagaricus virens (CFSZ19869). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Figure 13.
Leucoagaricus virens (CFSZ19869). (a) Basidiospores. (b) Basidia. (c) Cheilocystidia. (d) Elements of pileus squamules. Scale bars: 10 μm.
Diagnosis: Leucoagaricus virens is characterized by dark green (27E6) to light green (27B3) changes of pileus when bruised or dried; amygdaliform to ellipsoid basidiospores; and narrowly clavate, clavate to fusiform cheilocystidia.
Type: CHINA. Inner Mongolia Autonomous Region: Sandaohe, Heilihe National Nature Reserve, Ningcheng County, Chifeng City, 21 August 2018, T.Z. Liu and Y.X. Chen (CFSZ 19869, holotype).
GenBank Acc. No.: ITS = OM976881, LSU = OM976867 (CFSZ19794-2)
Description: Pileus, 7–13 mm in diameter; at first, nearly campanulate, becoming convex with age; center slightly umbonate, sometimes margin with inconspicuous striations; surface, whitish (1A2) to yellowish (2A3), covered with brown (6C3) to grey–brown (6E3), fibrillose, more or less radially arranged squamules; the central umbo is uniformly colored. The surface of the pileus discolors dark green (27E6) to light green (27B3) when bruised or dried. Context, thin; 1–2 mm, white (1A1), becoming dark green to light green when bruised or dried. Lamellae-free, crowded, white (1A1), slightly ventricose, with entire edges; discolors dark green to light green where bruised or dried. Stipe, 37–50 × 1–3 mm; subcylindrical, hollow, light yellowish brown (4B3), nearly smooth. Annulus, white (1A1), membranous, located at the upper part of the stipe. Flavor and odor not recorded. Spore print not observed.
Basidiospores [60/3/2], (7.5)8.0–10.0(10.5) × (4.0)4.5–5.0(5.5) μm; Q = (1.50)1.60–2.10(2.13); Qm = 1.85 ± 0.14; amygdaliform to elongate in side view; ellipsoid to elongate in front view; without germ pore, hyaline, slightly thick-walled (about 0.5–1.0 μm), dextrinoid, metachromatic in Cresyl Blue. Basidia, 16.5–27.5 × 8.5–10.0 μm; clavate, hyaline, four-spored; sterigmata, 4.5 μm long. Cheilocystidia, 27.5–52.0 × 9.5–17.0 μm; narrowly clavate to clavate, sometimes fusiform, hyaline, thin-walled. Pleurocystidia, absent. Lamellar trama, trabecular. Pileus squamules composed of loosely arranged subcylindrical hyphae with light brown (3B4) intracellular and extracellular pigments; terminal elements barely differentiated, subcylindrical to narrowly clavate (31.5–53.5 × 8.0–13.0 μm). Stipitipellis composed of hyaline, subcylindrical hyphae, 4.5–11.5 μm in diameter. Clamp connections absent in all parts of the basidiomata.
Habitat and Habit: solitary to scattered on soil in summer. So far, known only in Chifeng, Inner Mongolia Autonomous Region, China.
Other specimens examined: CHINA. Inner Mongolia Autonomous Region: Sandaohe, Heilihe National Nature Reserve, Ningcheng County, Chifeng City, 20 August 2018, T.Z. Liu, L. Zhang and Y. Bai (CFSZ 19794-2).
Comments: Basidiomata of
Leucoagaricus virens turn dark green to light green when bruised or dried. This feature is similar to those of
Leucoagaricus viriditinctus (Berk. & Broome) J.F. Liang, Zhu L. Yang & J. Xu, and
Leucoagaricus flavovirens J.F. Liang, Zhu L. Yang & J. Xu.
La. viriditinctus is also similar to
La. virens in the overall appearance of basidiomata. However,
La. viriditinctus, a species originally described in Sri Lanka, has a light blue to dark blue context when bruised or dried, and its cheilocystidia are broadly clavate to pyriform [
37].
Leucoagaricus virens is also similar to
La. flavovirens, a species originally described in China. Both species have amygdaliform to ellipsoid basidiospores and squamules composed of subcylindrical hyphae. However,
La. flavovirens has bluish green (27A7–8) changes of basidiomata when bruised [
37].
Leucocoprinus viridiflavus (Petch) E. Ludw. also has this feature.
Lc. viridiflavus is similar to
La. virens in having olive (1E4–1E5) to dark olive (2F8) changes of basidiomata on exposure of context. However,
Lc. viridiflavus have germ pore and utriform, ventricose or broadly clavate cheilocystidia [
38].
Leucoagarius atroazureus J.F. Liang, Zhu L. Yang & J. Xu and
La. caerulescens (Peck) J.F. Liang, Zhu L. Yang & J. Xu, both described in southern China, are also similar to
La. virens in having white context, brownish squamules on the pileus and similarly shaped basidiospores. However,
La. atroazureus has smaller basidiospores (5.5–8.0 × 3.5–5.5 μm) and dark blue changes of basidiomata when bruised or dried;
La. caerulescens has clavate to broadly clavate cheilocystidia and bluish green changes of basidiomata when dried [
37].





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}