

A Multivariate Study of Morphological Characters for Echinocactus horizonthalonius and E. texensis (Cactaceae) and Description of a New Subspecies, E. horizonthalonius subsp. australis


Abstract
1. Introduction
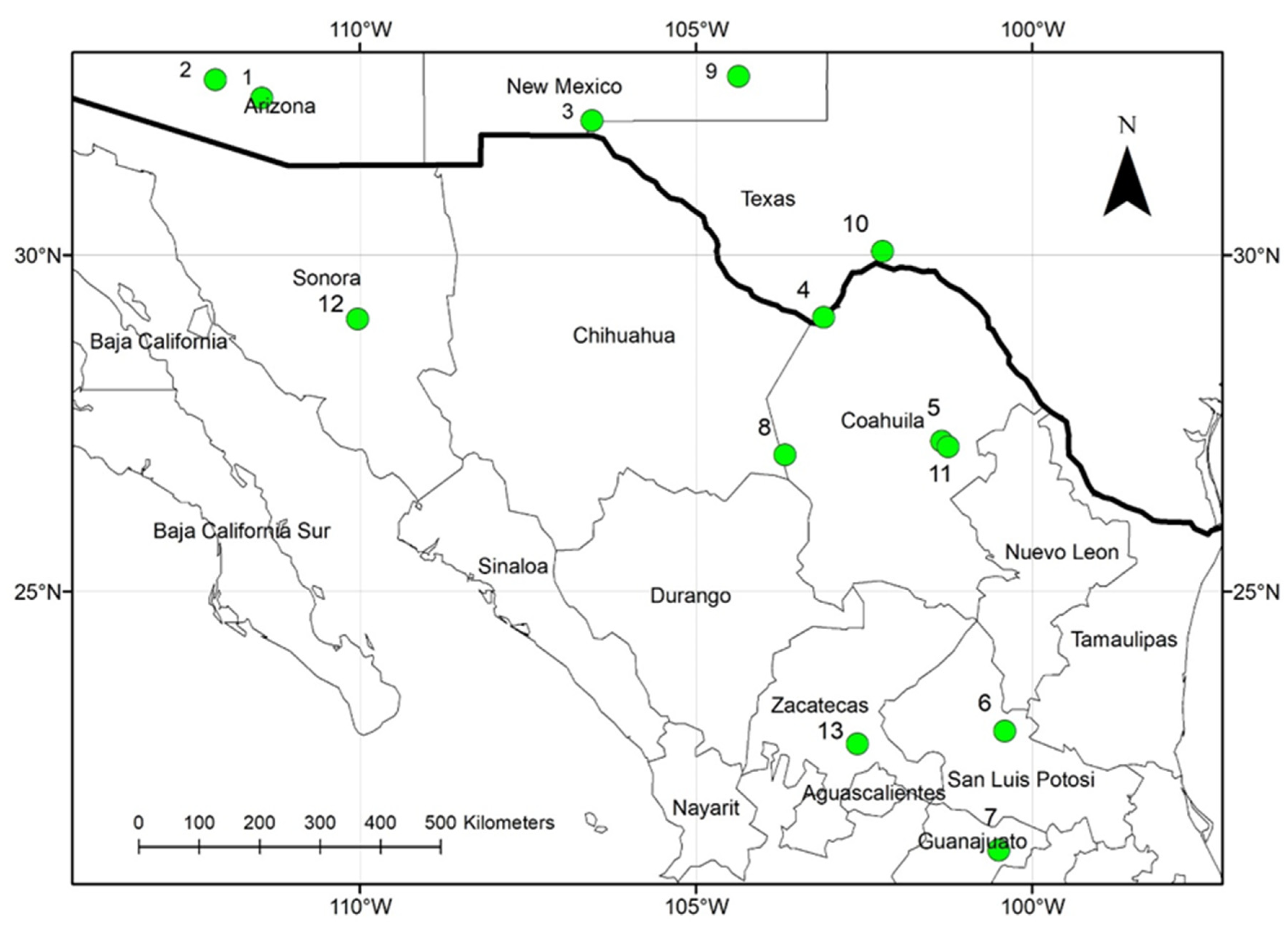
2. Methods
3. Results
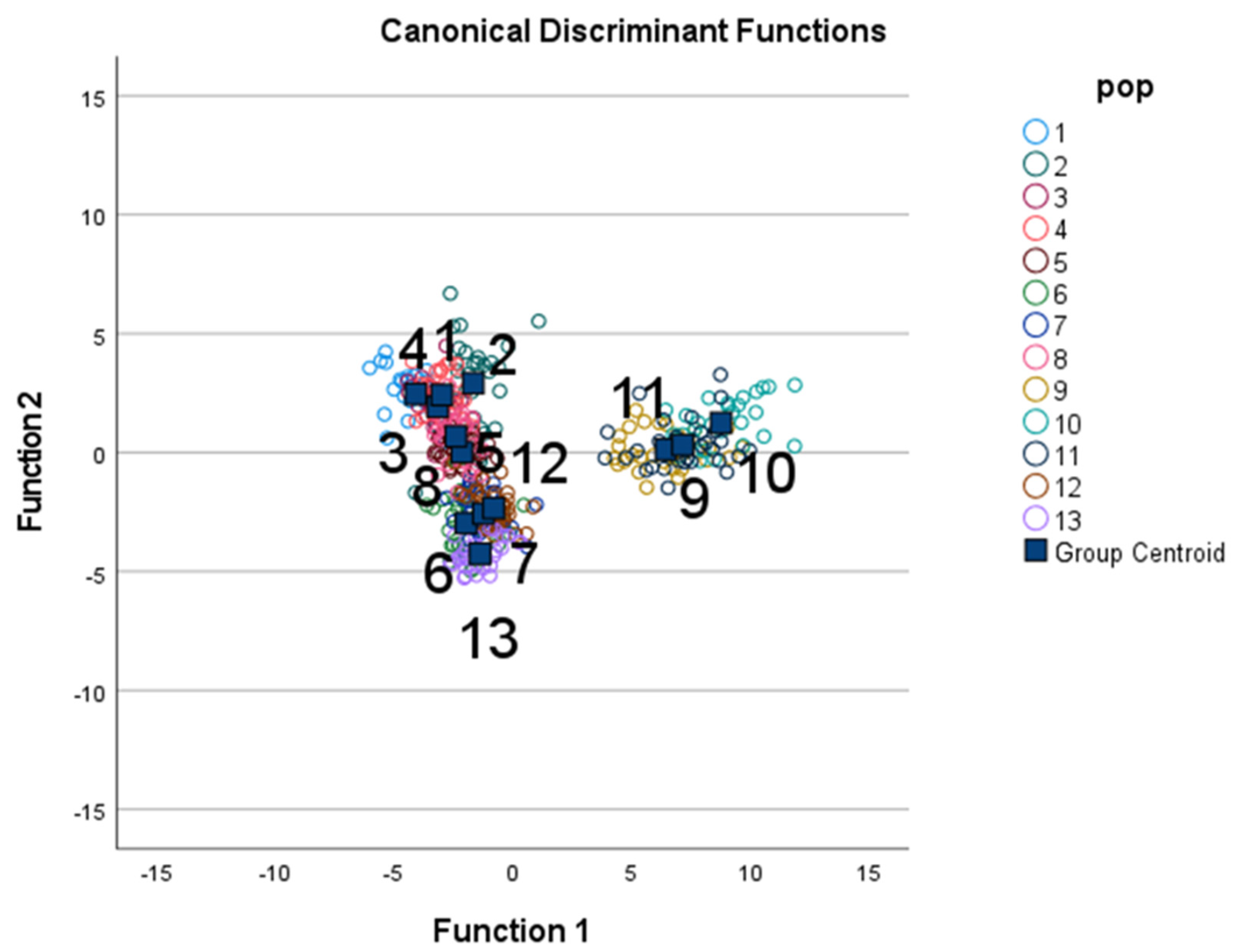
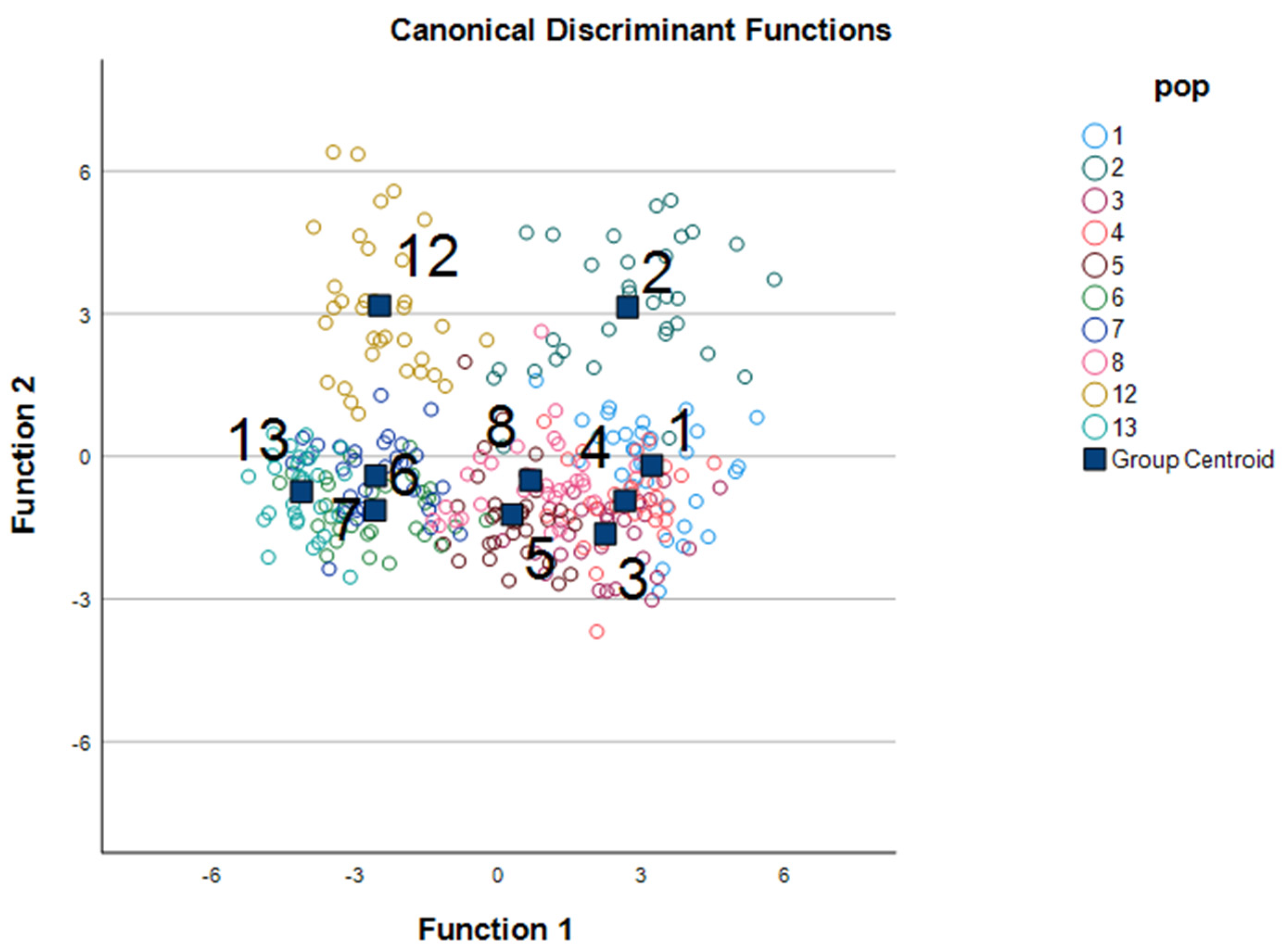
3.1. Discriminant Function Analysis
3.2. MANOVA
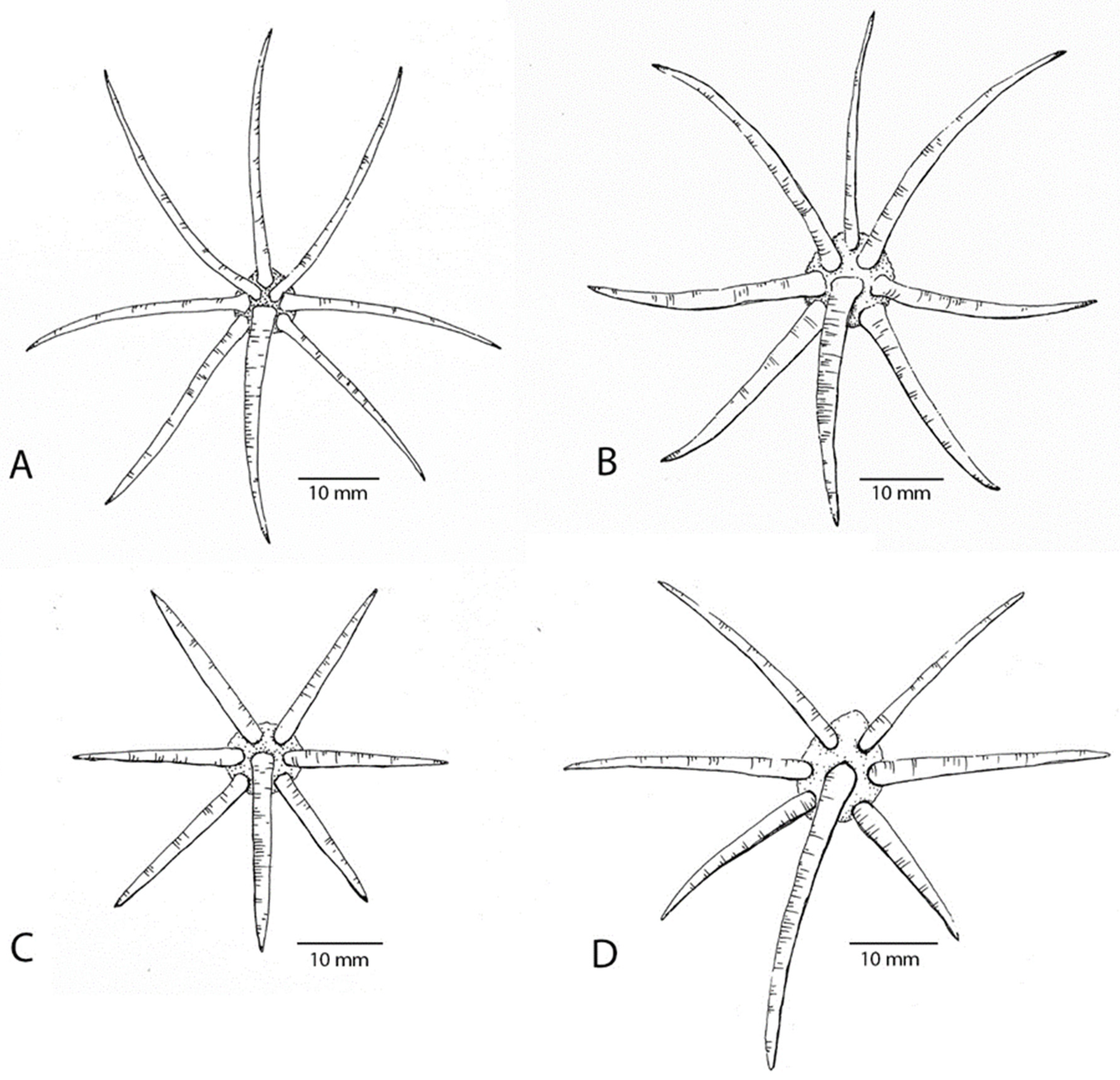
4. Discussion and Conclusions
4.1. Taxonomy
4.2. Key to the Subspecies within Echinocactus Horizonthalonius
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site No. | A priori Taxon | Locality | Habitat | Voucher |
| 1 | E. horizonthalonius var. nicholii | Arizona, Pima County, 735–780 m elevation; Waterman Mountains, ca. 50 km NW of Tucson. | Sparse scrub on limestone outcroppings and associated alluvium with Acacia constricta, Ambrosia deltoidea, Aristida purpurea, Calliandra eriophylla, Carnegiea gigantea, Cylindropuntia acanthocarpa, C. bigelovii, C. fulgida, C. leptocaulis, Dasyochloa pulchella, Echinocereus engelmannii, Encelia farinosa, Eriogonum inflatum, Ferocactus cylindraceus, Fouquieria splendens, Krameria erecta, Larrea tridentata, Cochemiea grahamii, Olneya tesota, Opuntia engelmannii, Parkinsonia microphylla, and Tiquilia canescens. | Parfitt 2788 (ASU) |
| 2 | E. horizonthalonius var. nicholii | Pinal County, Northwest end of Vekol Mountains, Tohono O’odham Nation. | Sparse scrub on lower bajada of limestone rock and gravel with Acacia biuncifera, Ambrosia deltoidea, A. dumosa, Carnegiea gigantea, Cylindropuntia acanthocarpa, C. fulgida, C. leptocaulis, Echinocereus engelmannii, Ephedra fasciculata, Eriogonum inflatum, Fouquieria splendens, Grusonia parishii, Janusia gracilis, Krameria bicolor, Larrea tridentata, Lycium berlandieri, Cochemiea grahamii, Muhlenbergia porteri, Olneya tesota, Opuntia engelmannii, Parkinsonia microphylla, and Zinnia acerosa. | Bruner 11572 (ASU) |
| 3 | E. horizonthalonius var. horizonthalonius | New Mexico, Doña Ana County, 32.006° N 106.554° W; 1255–1270 m elevation; Franklin Mountains, 2.8 km WSW of Anthony Gap, 38 km SE of Las Cruces | Larrea tridentata scrub on limestone hills with Agave lechuguilla, Aloysia wrightii, Bahia absinthifolia, Cylindropuntia leptocaulis, Echinocereus coccineus, E. dasyacanthus, Fouquieria splendens, Gutierrezia microcephala, Koeberlinia spinosa, Krameria erecta, Muhlenbergia porteri, Opuntia spinosibacca, Parthenium incanum, Thymophylla acerosa, T. pentachaeta, Tiquilia canescens, T. greggii, Yucca treculeana, and Zinnia acerosa. | Baker 16593 (ASU) |
| 4 | E. horizonthalonius var. horizonthalonius | Texas, Brewster County, 29.077° N 103.104° W; 690 m elevation; 550 m WSW of Boquillas, 500 m north of the confluence of Fresno Creek with the Rio Grande River; 1 km SE of the summit of Tally Mountain. | Limestone ridges with Acacia neovernicosa, Ariocarpus fissuratus, Bouteloua trifida, Cylindropuntia leptocaulis, Dasyochloa pulchella, Echinocactus horizonthalonius, E. dasyacanthus, Ephedra trifurca, Glandulicactus uncinatus, Guaiacum angustifolium, Leucophyllum frutescens, Opuntia spinosibacca, Porophyllum gracilis, Prosopis glandulosa, Selaginella lepidophylla, Tiquilia canescens, and Tridens mutica. | Baker 16595.2 (ASU, photos only) |
| 5 | E. horizonthalonius var. horizonthalonius | México, Coahuila, 27.231° N 101.350° W; 1135 m elevation; 3 km north of Plan de Guadalupe, 90 km NNW of Saltillo. | Larrea tridentata scrub with Agave lechuguilla, Bahia absinthifolia, Cylindropuntia imbricata, C. kleiniae, Dasyochloa pulchella, Euphorbia antisyphilitica, Fouquieria splendens, Jatropha dioica, Lippia graveolens, Lophophora williamsii, Opuntia engelmannii, O. rufida, Parkinsonia texana, Pennisetum ciliare, Thelocactus bicolor, Tiquilia canescens, and Yucca treculeana. | Baker 16612 (ASU) |
| 6 | E. horizonthalonius var. horizonthalonius | México, San Luis Potosí, 22.928° N 100.411° W; 1400 m elevation; 97 km NNE of San Luis Potosí; 1 km ENE of El Entronque, northern tip of Sierra Cuchillo de Enmedio; along Hwy 80, 5 km (by road) east of its junction with Hwy 57. | Larrea tridentata scrub with Agave scabra, Celtis pallida, Cylindropuntia kleiniae, C. leptocaulis, C. tunicata, Echinocactus platyacanthus, Echinocereus pectinatus, Jatropha dioica, Koeberlinia spinosa, Myrtillocactus geometrizans, Opuntia engelmannii, Prosopis laevigata, Thelocactus bicolor, Tiquilia canescens, and Yucca decipiens. | Baker 16119 (ASU) |
| 7 | E. horizonthalonius var. horizonthalonius | México, Guanajuato, 21.163° N 100.500° W; 2130 elevation; 15 km south of San Luis de la Paz (Mineral de la Pozos), just NE of the Ejido of Espinas Blancas. | Disclimax grassland on rocky volcanic soil with Bouteloua curtipendula, Cathestecum erectum, Coryphantha erecta, Cylindropuntia imbricata, C. tunicata, Dasyochloa pulchella, Erioneuron pilosum, Jatropha dioica, Myrtillocactus geometrizans, Opuntia engelmannii, Opuntia leucotricha, Parthenium incanum, and Yucca filifolia | R. T. Barcenas T-115 |
| 8 | E. horizonthalonius var. horizonthalonius | México, Coahuila, 27.029° N 103.682° W; 1115 m elevation; 28 km south of La Esmeralda; 5 km east of Guimbalete, 7 km SSE of Cerro el Venado Pelón. | Larrea tridentata scrub with Acacia neovernicosa, Bahia absinthifolia, Coryphantha macromeris, C. poselgeriana, Cylindropuntia leptocaulis, C. imbricata, Echinocereus stramineus, Ferocactus hamatacanthus, Euphorbia antisyphilitica, Flourensia cernua, Fouquieria splendens, Jatropha dioica, Mammillaria heyderi, Opuntia rufida, Parthenium incanum, and Viguiera stenoloba. | Baker 16608 (ASU) |
| 9 | E. texensis | New Mexico, Eddy County, 32.661° N 104.375° W; 1020 m elevation; 1 km south of Fourmile Draw, 6 km west of the Pecos River, 20 km south of the center of Artesia. | Scleropogon brevifolius grassland with Coryphantha macromeris, Croton pottsii, Flourensia cernua, Gutierrezia sarothrae, Krameria erecta, Larrea tridentata, Lesquerella fendleri, Opuntia tortispina, Pleuraphis mutica, Prosopis glandulosa, Rhus microphylla, and Yucca glauca. | Baker 16617 (ASU) |
| 10 | E. texensis | Texas, Terrell County, 30.058° N 102.235° W; 695 m elevation; 18 km ESE of Sanderson, between Hwy 90 and the railroad tracks, just west of the old Mofeta Railroad stop. | Larrea tridentata scrub with Coryphantha macromeris, Cylindropuntia leptocaulis, Echinocactus horizonthalonius, Echinocereus coccineus, Flourensia cernua, Koeberlinia spinosa, Lycium berlandieri, Opuntia engelmannii, O. mackenensii, O. strigil, Panicum hallii, Parthenium incanum, Prosopis glandulosa, Tiquilia canescens, Viguiera stenoloba, and Yucca treculeana. | Baker 16596 (ASU) |
| 11 | E. texensis | México, Coahuila, 27.153° N 101.257° W; 395 m elevation; just east of Río Monclova, 10 km SSW of Primero de Mayo, 5 km west of Cuchillo de Arco, 30 km ENE of San Bueno Ventura. | Larrea tridentata scrub with Bahia absinthifolia, Coryphantha macromeris, Cylindropuntia kleiniae, C. leptocaulis, Echinocereus enneacanthus, Flourensia cernua, Fouquieria splendens, Guaiacum angustifolium, Lycium berlandieri, Mammillaria heyderi, Muhlenbergia porteri, Opuntia engelmannii, Prosopis glandulosa, and Tiquilia canescens. | Baker 16609 (ASU) |
| 12 | E. horizonthalonius var. horizonthalonius | México, Sonora, 660 m elevation, 90 km east of Hermosillo, 11 km ENE Mazatán, 14 km ESE of the summit of Sierra Mazatán. | Low ridges of igneous and sedimentary alluvium, including a surface layer of rocks and gravel, grassland/sparse shrubland with Acacia constricta, Agave angustifolia var. angustifolia, Agave shrevei subsp. matapensis, Allionia incarnata, Aristida adscensionis, A. ternipes var. ternipes, Bouteloua barbata var. barbata, B. diversispicula, Bursera fagaroides var. elongata, Bursera laxiflora, Caesalpinia pulcherrima, Callaeum macropterum, Calliandra eriophylla, Condalia warnockii, Cottsia linearis, Dalea mollis, Ditaxis neomexicana, Evolvulus alsinoides var. angustifolia, Eysenhardtia orthocarpa var. orthocarpa, Fouquieria splendens, Guaiacum coulteri, Haematoxylum brasiletto, Hedeoma nanum, Jatropha cardiophylla, Karwinskia humboldtiana, Krameria erecta, Lysiloma watsonii, Mimosa distachya, Opuntia durangensis, Parkinsonia × sonorae, Plantago ovata, Polygala macradenia, Porophyllum gracile, and Turnera diffusa. | Baker 17574 (ASU, photos only) |
| 13 | E. horizonthalonius var. horizonthalonius | México, Zacatecas, 22.73435° N 102.60492° W (WGS84), 2385 m elevation, just south of the outskirts of the metropolis of Zacatecas, 5 km SSW of its center. | Low scrub/ grassland with Acalypha phleoides, Astragalus diphacus, Baccharis pteronioides, Bouteloua curtipendula, B. gracilis, Calylophus hartwegii, Dalea prostrata, Dasyochloa pulchella, Echeveria paniculata, Euphorbia cuphosperma, Krameria pauciflora, Lycurus phleoides, Mammillaria heyderi, Mimosa biuncifera, Oenothera kunthiana, Opuntia leucotricha, O. robustispina, Penstemon roseus, Sanvitalia procumbens, Solanum elaeagnifolium, Thelesperma megapotamicum, Viguiera dentata, Xanthisma spinulosum, and Yucca decipiens. | Photo info? Maybe reference Figure 1. |
References
- Baker, M.A.; Butterworth, C. Geographic distribution and taxonomic circumscription of populations within Coryphantha section Robustispina (Cactaceae). Am. J. Bot. 2013, 100, 984–997. [Google Scholar] [CrossRef]
- Baker, M.A.; Porter, J.M. The use of multivariate analyses of morphological characters and DNA analyses for assessing the taxonomic ranking of rare plant taxa: An example comparing populations of Echinomastus erectocentrus var. acunensis with those of its relatives E. erectocentrus var. erectocentrus and E. johnsonii. In Endangered Species: Threats, Conservation and Future Research; Quinn, M., Ed.; Nova Science Publishers, Inc., Hauppauge: New York, NY, USA, 2016; pp. 19–61. [Google Scholar]
- Majure, L.C.; Baker, M.A.; Cloud-Hughes, M.; Salywon, A.; Neubig, K.M. Phylogenomics in Cactaceae: A case study using the chollas sensu lato (Cylindropuntieae, Opuntioideae) reveals a common pattern out of the Chihuahuan and Sonoran deserts. Am. J. Bot. 2019, 106, 1327–1345. [Google Scholar] [CrossRef] [PubMed]
- Bravo-Hollis, H. Las Cactáceas de México; México UNAM: Mexico City, México, 1937. [Google Scholar]
- Powell, M.A.; Weedin, J.F. Cacti of the Trans-Pecos and Adjacent Areas; Texas Tech University Press: Lubbock, TX, USA, 2004. [Google Scholar]
- Zimmerman, A.D.; Parfitt, B.D. Cactaceae: Echinocactus. In Flora of North America Editorial Committee, Flora of North America; Missouri Botanical Garden Press: St. Louis, MO, USA, 2003; Volume 4. [Google Scholar]
- Vargas-Luna, M.D.; Hernández-Ledesma, P.; Majure, L.C.; Puente-Martínez, R.; Macías, H.M.H.; Luna, R.T.B. Splitting Echinocactus: Morphological and molecular evidence support the recognition of Homalocephala as a distinct genus in the Cactaceae. PhytoKeys 2018, 111, 31–59. [Google Scholar] [CrossRef]
- Goettsch, B.; Hilton-Taylor, C.; Cruz-Piñón, G.; Duffy, J.P.; Frances, A.; Hernández, H.M.; Inger, R.; Pollock, C.; Schipper, J.; Superina, M.; et al. High proportion of cactus species threatened with extinction. Nat. Plants 2015, 1, 15142. [Google Scholar] [CrossRef]
- Worthington, R.D. Observations on the flowering cacti from the vicinity of El Paso, Texas. Cactus Succul. J. 1986, 58, 213–217. [Google Scholar]
- Benson, L. The Cacti of Arizona, 3rd ed.; University of Arizona Press: Tucson, AZ, USA, 1969. [Google Scholar]
- Aguilar, R.P.; van Devender, T.R.; Felger, R.S. Cactáceas de Sonora, México: Su Diversidad, Uso y Conservación; Arizona-Sonora Desert Museum Press: Tucson, AZ, USA, 2000. [Google Scholar]
- Yatskievych, G.; Fischer, P.C. New plant records from the Sonoran Desert. Desert Plants 1983, 5, 180–185. [Google Scholar]
- Benson, L. The Cacti of the United States and Canada; Stanford University Press: Stanford, CA, USA, 1982. [Google Scholar]
- Anderson, E. The Cactus Family; Timber Press: Portland, OR, USA, 2001. [Google Scholar]
- U.S. Fish and Wildlife Service. Nichol Turk’s Head Cactus (Echinocactus horizonthalonius var. nicholii) Recovery Plan; U.S. Fish and Wildlife Service: Albuquerque, NM, USA, 1986. [Google Scholar]
- Butterwick, M. Nichol’s Turk’s Head Cactus Habitat Management Plan; BLM: Phoenix, AZ, USA, 1985. [Google Scholar]
- Phillips, A.M., III; Phillips, B.G.; Green, L.T., III; Mazzoni, J.; Peterson, E.M. Status Report: Echinocactus Horizonthalonius Lemaire var. nicholii L. Benson; USFWS: Albuquerque, NM, USA, 1979. [Google Scholar]
- Wallace, R.S. Molecular systematic study of the Cactaceae: Using chloroplast DNA variation to elucidate cactus phylogeny. Bradleya 1995, 13, 1–12. [Google Scholar] [CrossRef]
- IBM SPSS Statistics for Windows, Version 28.0.1.1; IBM Corp: Armonk, NY, USA, 2015.
- Baker, M.A. Morphological and cytological analyses in Cylindropuntia (Cactaceae): The taxonomic circumscription of C. echinocarpa, C. multigeniculata, and C. whipplei. J. Bot. Res. Inst. Tex. 2016, 10, 325–343. [Google Scholar]
- Baker, M.A.; Pinkava, D.J.; Cloud-Hughes, M.A. Analysis of infraspecific taxa within Cylindropuntia acanthocarpa using multivariate analysis of morphological characters. Canotia 2018, 14, 66–92. [Google Scholar]
- Stebbins, G.L. Variation and Evolution in Plants; Columbia University Press: New York, NY, USA; London, UK, 1950. [Google Scholar]
- Baker, M.A.; Cloud-Hughes, M.A.; Rebman, J.P.; Majure, L.C. Nomenclatural changes in Cylindropuntia (Cactaceae). Madroño 2019, 66, 92. [Google Scholar] [CrossRef]
- Weniger, D. Cacti of Texas and Neighboring States; University of Texas Press: Austin, TX, USA, 1984. [Google Scholar]





| Character | Explanation |
|---|---|
| Stem height from soil level | Length of stem from soil surface to apex, excluding spines |
| * Stem diameter midway | Diameter of stem midway between soil level and stem apex |
| Stem height divided by stem diameter midway | Derived character reflecting overall stem shape |
| * Stem diameter base | Diameter of stem at soil level |
| Stem diameter midway divided by stem diameter base | Derived character reflecting shape of lower portion of stem |
| Rib number | Number of ribs |
| The following characters are measured three times for each individual. Spine measurements are made on the uppermost fully mature spine clusters as determined by size and color of spines | |
| Rib height | Maximum height of rib excluding spines and pubescence |
| Rib width near stem apex | Distance from sinus to sinus along a rib at the edge of the wooly apex of the stem |
| Rib width maximum | Maximum distance from sinus to sinus along a rib, generally toward base of stem |
| Length between areoles | Maximum distance between three areoles divided by two |
| Central spine number | The number of spines with bases centrally inserted within the areole |
| Radial spine number | The number of spines with bases inserted at the edge of the areole |
| Lower central spine length | The length of the lower central spine measured along its curvature |
| Lower central spine curvature | The greatest perpendicular distance between the surface of the lower central spine (generally near the midpoint) and a line between the spine base and apex  |
| Upper central spine length | The length of the longest upper central spine |
| Upper central spine curvature | The greatest perpendicular distance between the surface of the upper central spine (generally near the midpoint) and an imaginary line between the spine base and apex |
| Radial spine length | The length of the longest radial spine |
| Radial spine curvature | The greatest perpendicular distance between the surface of the radial spine (generally near the midpoint) and an imaginary line between the spine base and apex |
| Lower central spine width | The lateral width of the lower central spine as measured near its midpoint. Generally greater than the dorso-ventral thickness |
| * Lower central spine dorso-ventral thickness | The dorso-ventral thickness of the lower central spine as measured near its midpoint |
| Lower central spine width divided by its thickness | A derived character assessing flatness of the spines |
| Upper central spine width | The lateral width of the upper central spine as measured near its midpoint |
| Radial spine lateral width | The lateral width of the radial spine as measured near its midpoint |
| Function | Eigenvalue | % of Variance | Cumulative % | Canonical Correlation |
|---|---|---|---|---|
| 1 | 12.073 | 75.3 | 75.3 | 0.961 |
| 2 | 2.571 | 16.0 | 91.3 | 0.849 |
| 3 | 1.399 | 8.7 | 100.0 | 0.764 |
| Taxon | Predicted Group Membership | Total | |||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | ||||
| Original | Count | 1 | 91 | 0 | 0 | 0 | 91 |
| 2 | 0 | 91 | 6 | 2 | 99 | ||
| 3 | 0 | 2 | 118 | 0 | 120 | ||
| 4 | 0 | 0 | 1 | 91 | 92 | ||
| Ungrouped cases | 0 | 0 | 5 | 0 | 5 | ||
| % | 1 | 100.0 | 0.0 | 0.0 | 0.0 | 100.0 | |
| 2 | 0.0 | 91.9 | 6.1 | 2.0 | 100.0 | ||
| 3 | 0.0 | 1.7 | 98.3 | 0.0 | 100.0 | ||
| 4 | 0.0 | 0.0 | 1.1 | 98.9 | 100.0 | ||
| Ungrouped cases | 0.0 | 0.0 | 100.0 | 0.0 | 100.0 | ||
| Character | E. texensis | E. horizonthalonius var. horizonthalonius | E. horizonthalonius var. nicholii | E. horizonthalonius taxon nov. |
|---|---|---|---|---|
| Stem height divided by stem diameter midway | 0.31 | 0.95 | 0.9 9 | 0.47 |
| Rib number | 17.08 * | 8.00 | 8.01 | 8.05 |
| Stem diameter midway divided by stem diameter base | 0.96 | 1.00 * | 1.04 * | 0.95 |
| Rib height | 16.80 | 16.65 | 17.57 | 12.48 * |
| Rib width near stem apex | 4.37 * | 14.9o | 15.38 | 15.55 |
| Rib width maximum | 29.63 * | 41.29 | 45.13 | 43.45 |
| Length between areoles | 64.03 * | 17.65 | 17.97 | 21.24 |
| Central spine number | 2.99 | 3.08 | 2.97 | 3.02 |
| Radial spine number | 4.22 | 5.01 | 4.80 | 4.06 * |
| Lower central spine length | 40.92 * | 29.54 | 30.53 | 22.07 * |
| Lower central spine angle | 36.60 | 30.68 | 31.09 | 13.95 * |
| Lower central spine curvature | 1.53 | 1.43 | 1.67 | 0.24 * |
| Number of lower central spine annuli | 11.92 | 12.99 | 16.69 | 15.35 |
| Upper central spine length | 29.05 | 29.77 | 35.44 * | 21.24 * |
| Upper central spine angle | 50.99 | 47.05 | 46.43 | 44.99 |
| Upper central spine curvature | 0.62 | 2.52 | 3.05 | 0.19 * |
| Radial spine length | 31.18 | 28.19 | 29.84 | 20.21 |
| Radial spine angle | 17.46 | 26.01 | 23.70 | 13.98 * |
| Radial spine curvature | 0.7440 | 1.8867 | 1.6415 | 0.5319 * |
| Lower central spine width | 277 | 168 | 238 | 163 |
| Lower central spine width divided by thickness | 1.63 | 1.55 * | 1.81 * | 1.25 |
| Upper central spine width | 164 | 112 | 156 * | 137 |
| Radial spine width | 192 | 136 | 159 | 164 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baker, M.A. A Multivariate Study of Morphological Characters for Echinocactus horizonthalonius and E. texensis (Cactaceae) and Description of a New Subspecies, E. horizonthalonius subsp. australis. Diversity 2022, 14, 1020. https://doi.org/10.3390/d14121020
Baker MA. A Multivariate Study of Morphological Characters for Echinocactus horizonthalonius and E. texensis (Cactaceae) and Description of a New Subspecies, E. horizonthalonius subsp. australis. Diversity. 2022; 14(12):1020. https://doi.org/10.3390/d14121020
Chicago/Turabian StyleBaker, Marc A. 2022. "A Multivariate Study of Morphological Characters for Echinocactus horizonthalonius and E. texensis (Cactaceae) and Description of a New Subspecies, E. horizonthalonius subsp. australis" Diversity 14, no. 12: 1020. https://doi.org/10.3390/d14121020
APA StyleBaker, M. A. (2022). A Multivariate Study of Morphological Characters for Echinocactus horizonthalonius and E. texensis (Cactaceae) and Description of a New Subspecies, E. horizonthalonius subsp. australis. Diversity, 14(12), 1020. https://doi.org/10.3390/d14121020