
Biomass Partitioning and Morphoanatomical Traits of Six Gymnocalycium (Cactaceae) Species Occurring along a Precipitation Gradient

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
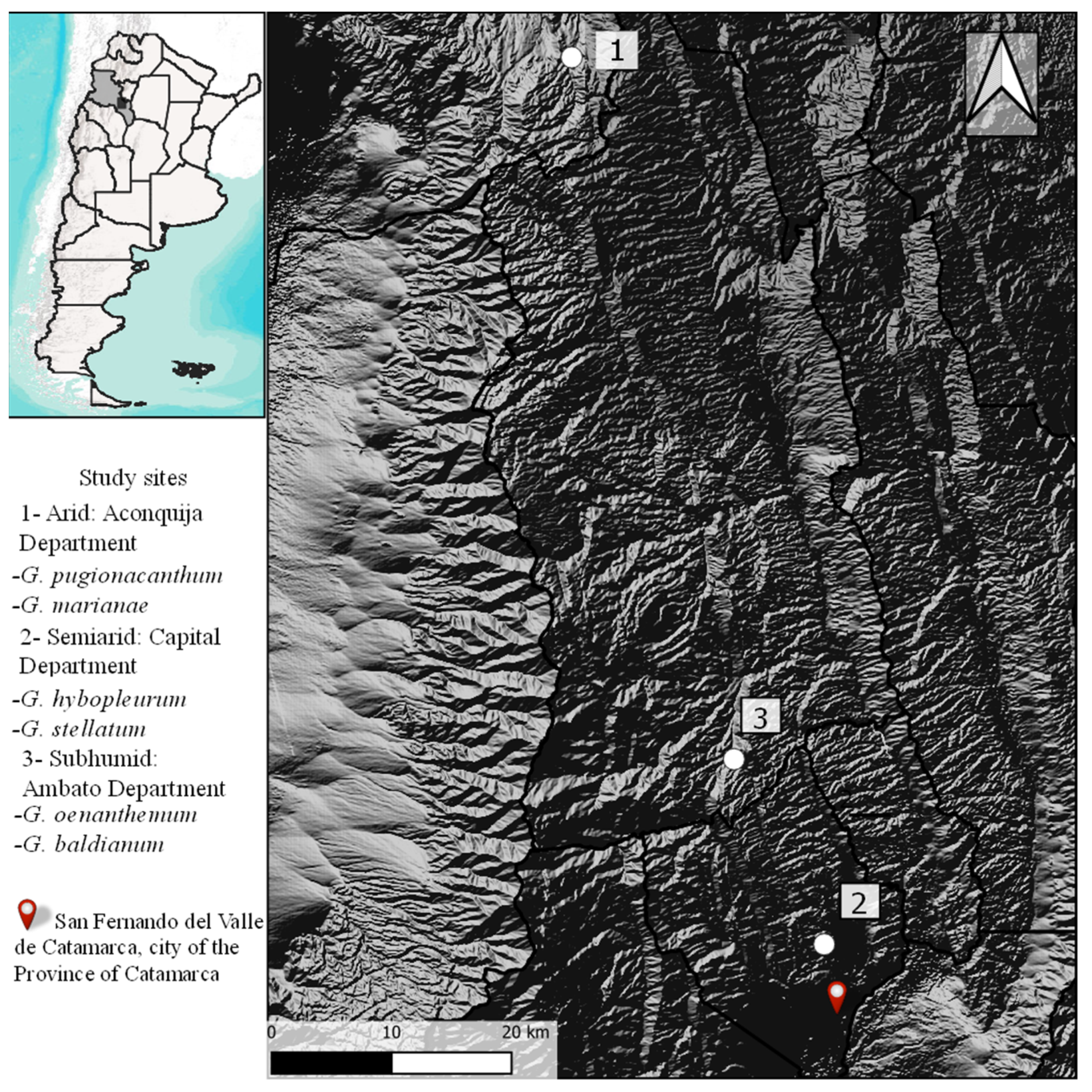
2.1. Study Species and Area
2.2. Morphological Traits
2.3. Biomass Partitioning
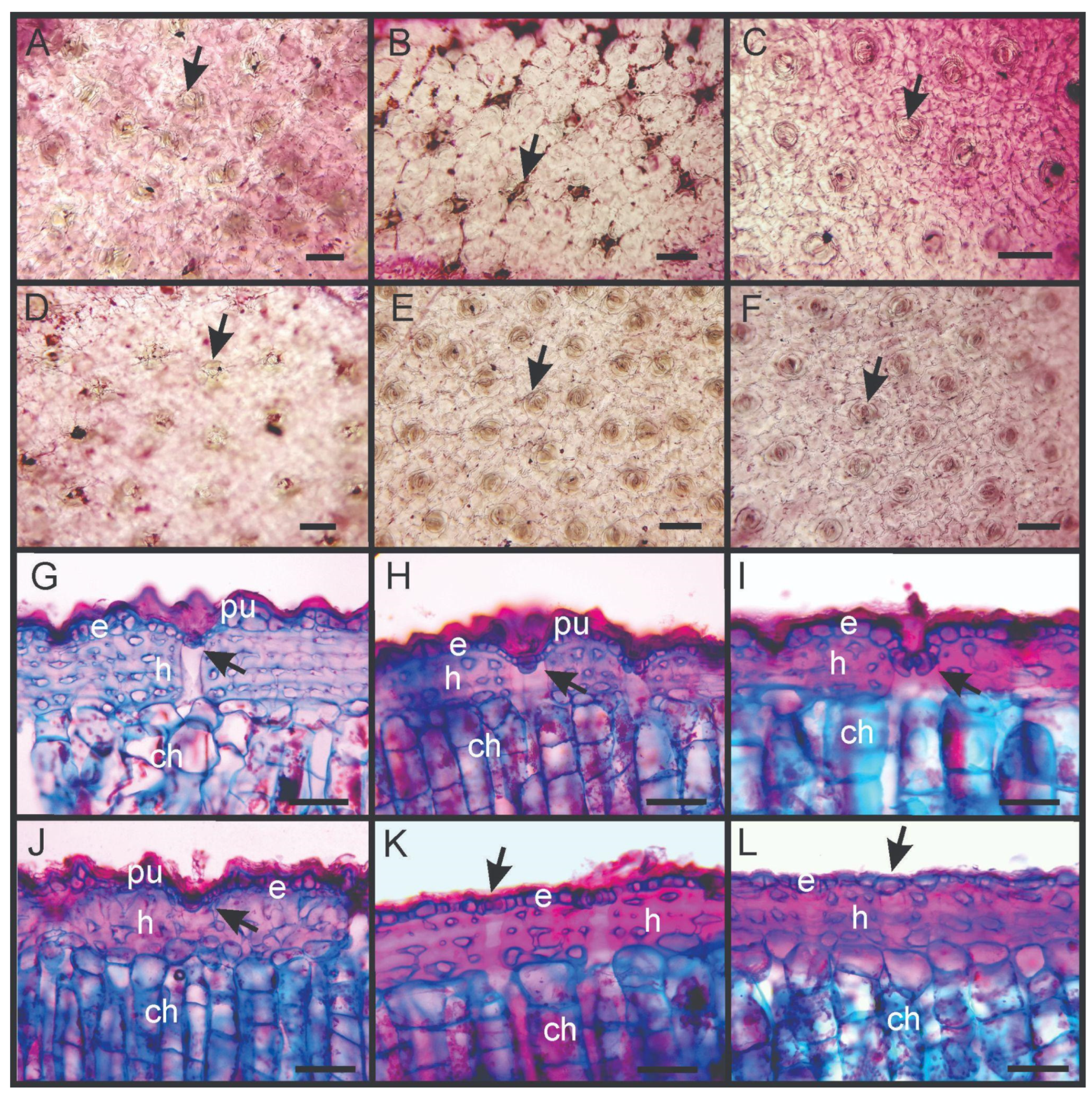
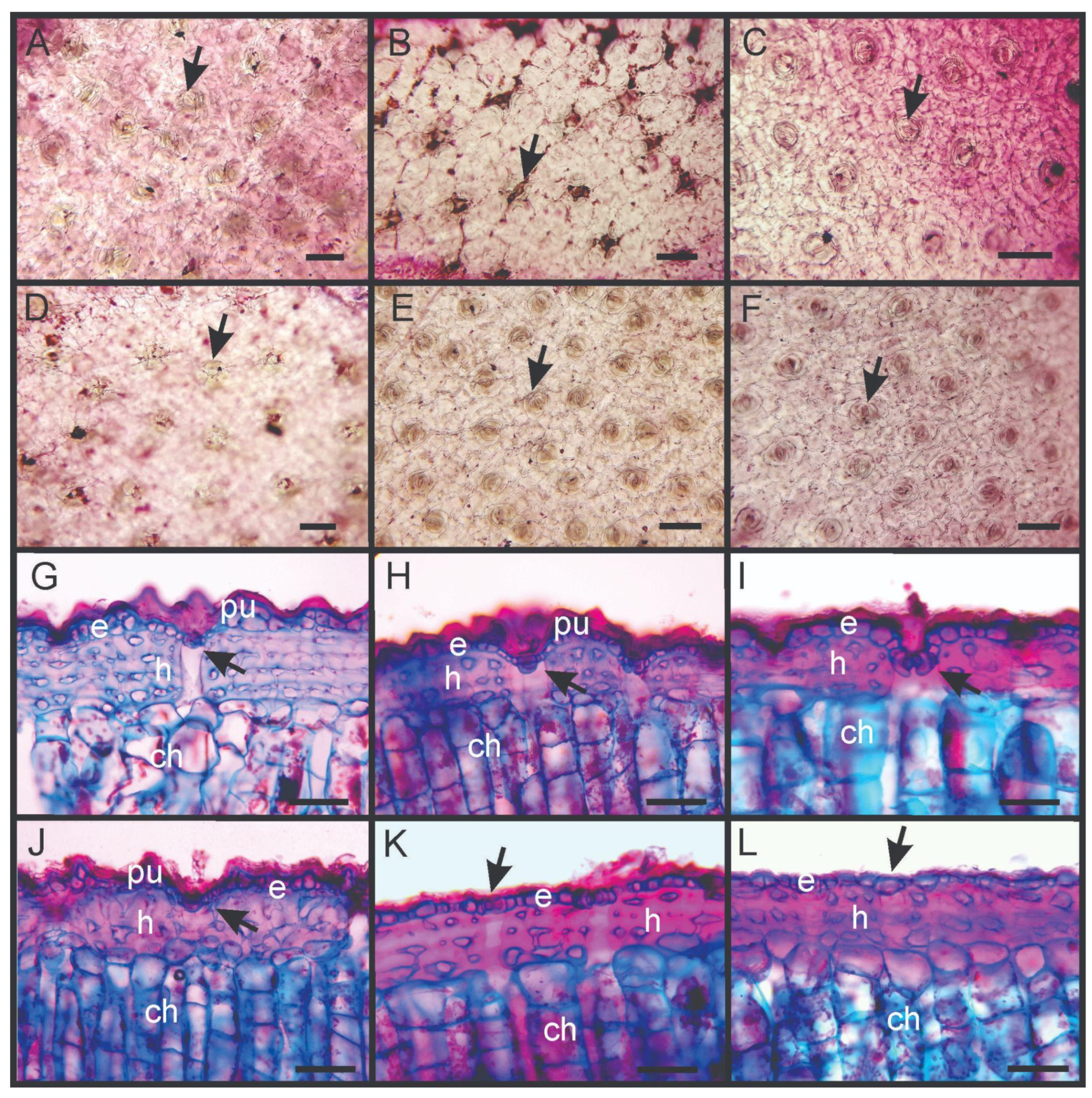
2.4. Anatomical Traits
2.5. Statistical Analysis
3. Results
3.1. Morphological Traits
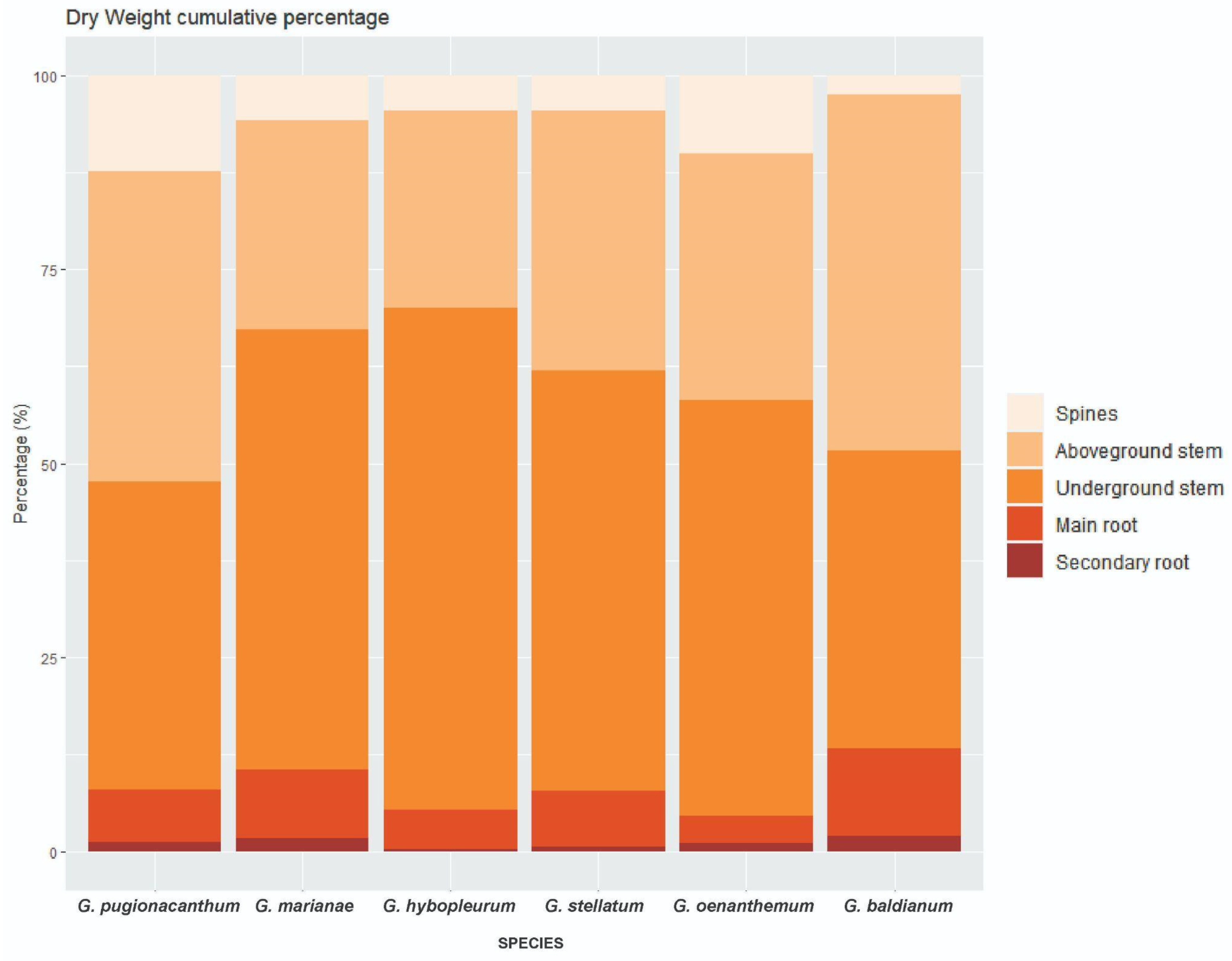
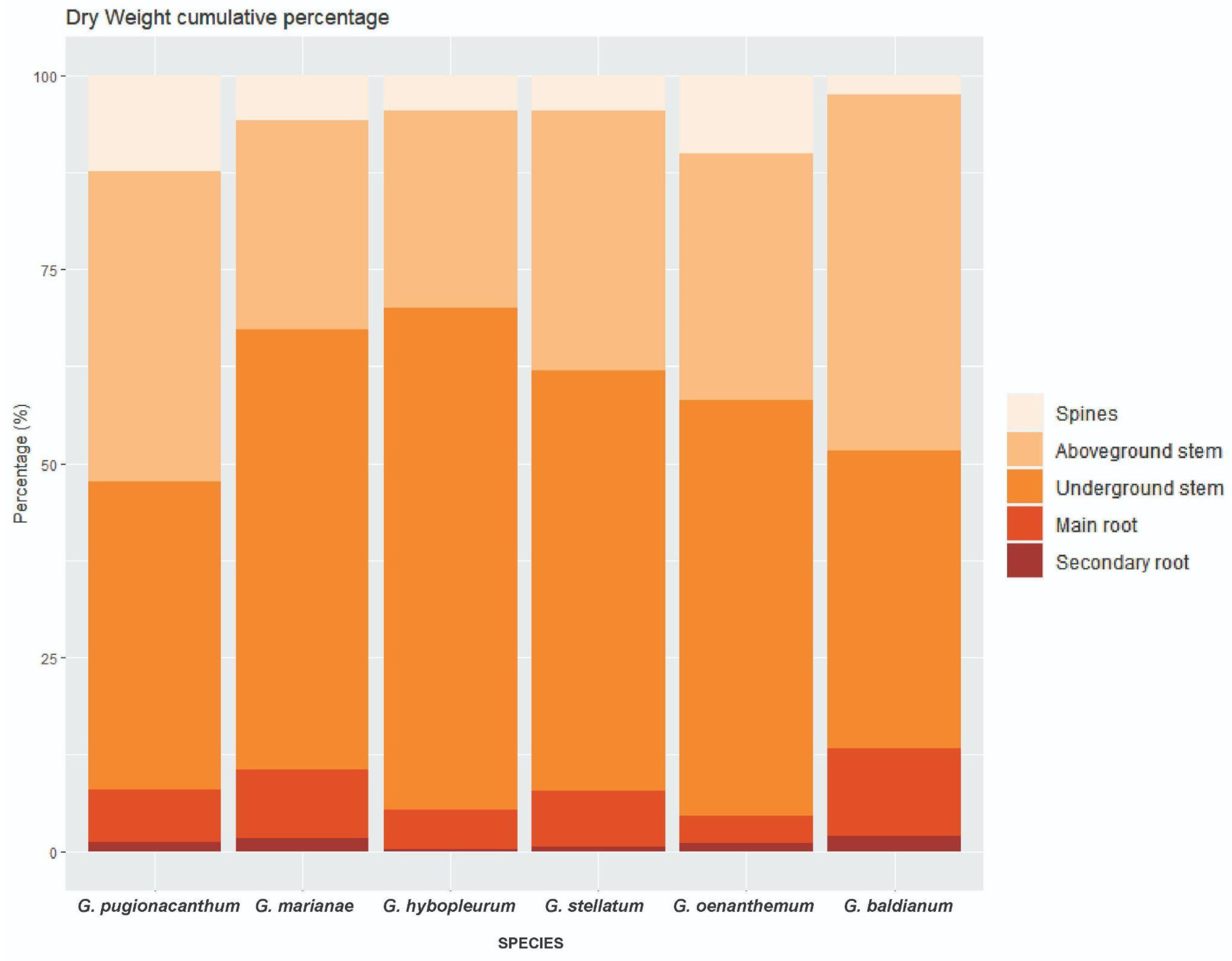
3.2. Biomass Partitioning
3.3. Anatomical Traits
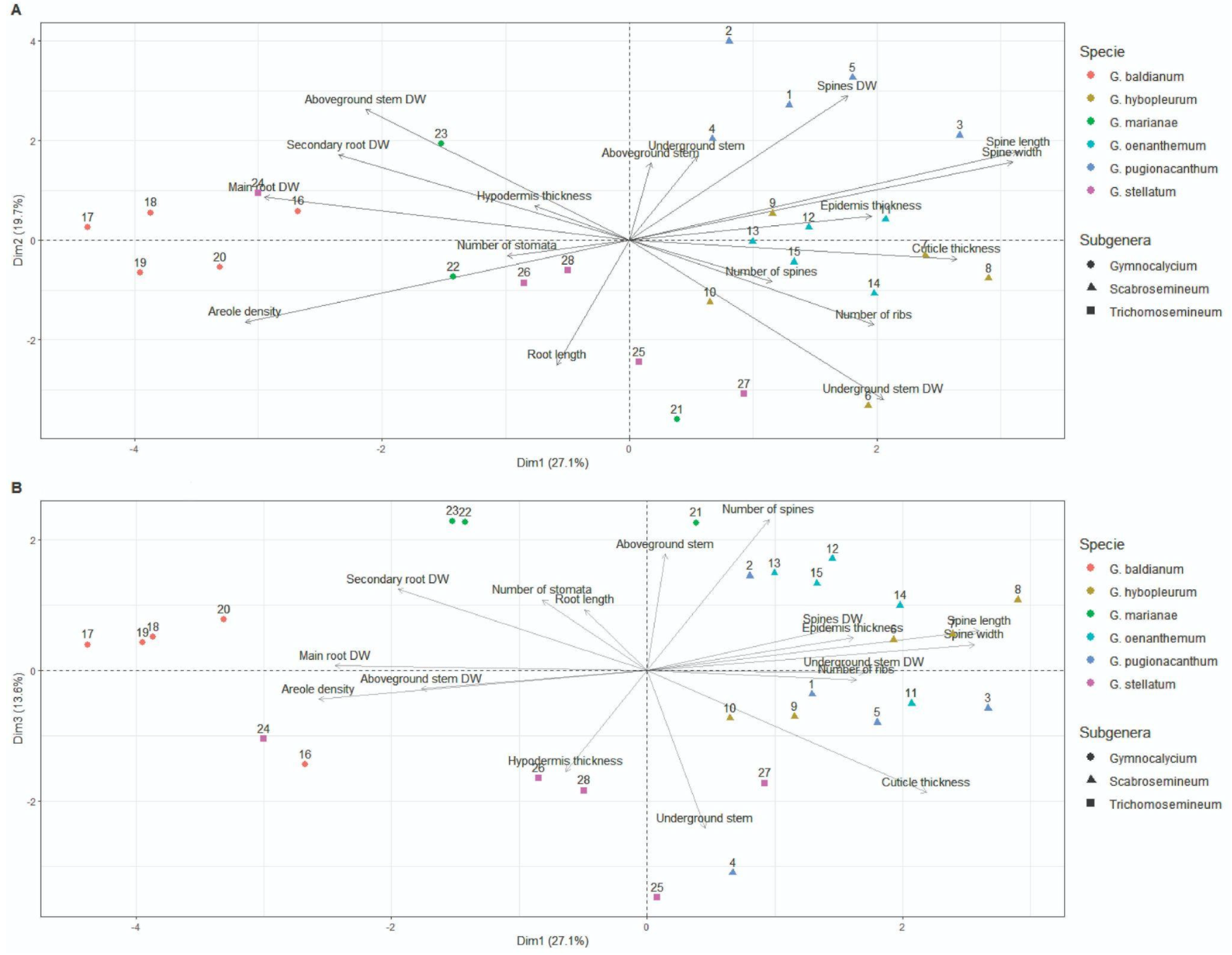
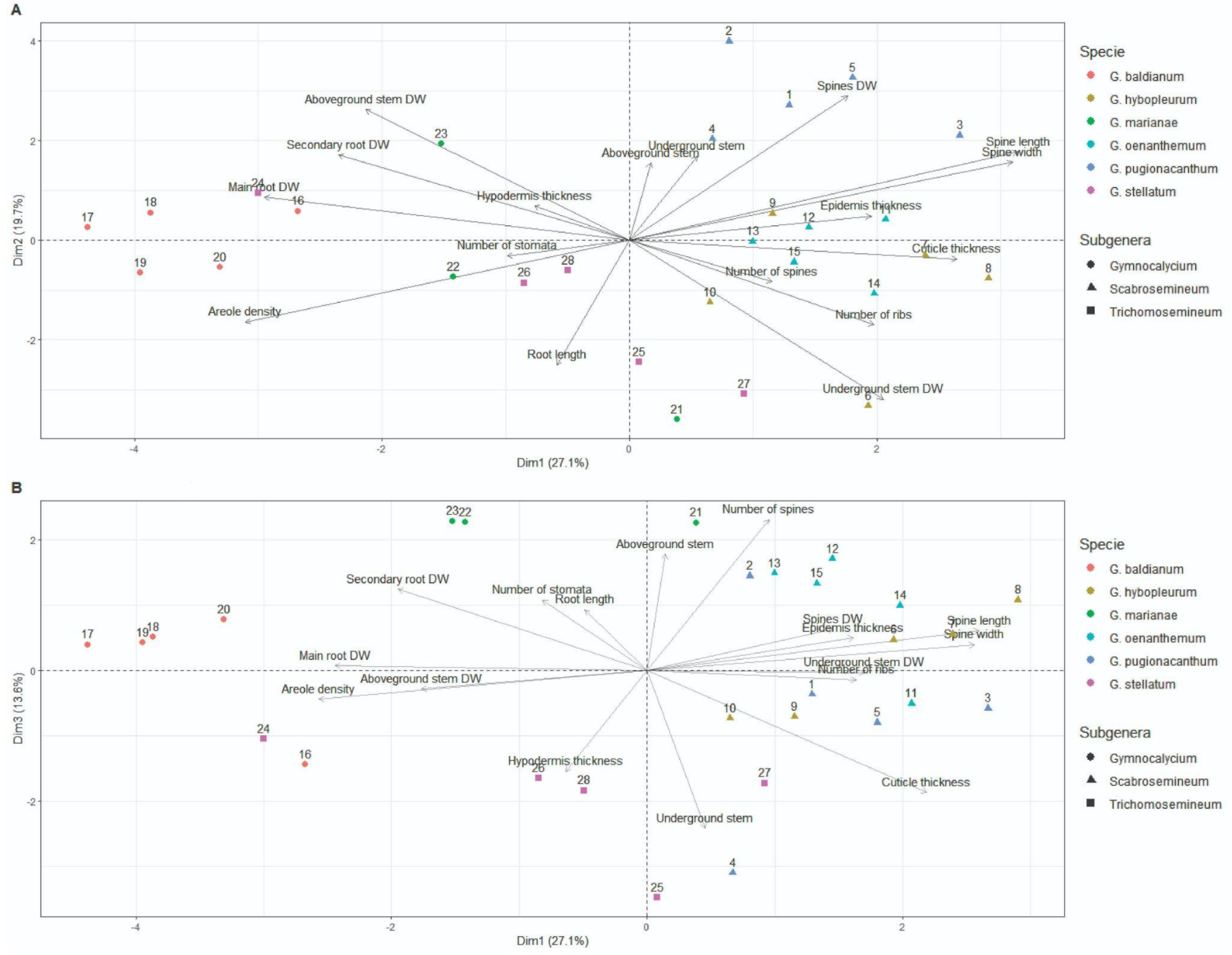
3.4. Multivariate Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Individual | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| G. pugionacanthum | 24.005 | 38.075 | 37.920 | 8 | 3 | 8 | 24.21 | 1.65 | 10.739 | 9.287 | 0.806 | 42.84 | 36.328 | 25 | 52.2 | 168.8 | 29.392 | Scabrosemineum |
| G. pugionacanthum | 38.766 | 30.053 | 31.181 | 11 | 3 | 5 | 25.17 | 1.37 | 13.997 | 5.587 | 2.837 | 47.78 | 29.8 | 14.6 | 55.6 | 122 | 24..128 | Scabrosemineum |
| G. pugionacanthum | 15.49 | 45.99 | 38.521 | 10 | 3.5 | 5 | 29.56 | 2.77 | 14.916 | 9.472 | 0.474 | 20.842 | 54.297 | 15.6 | 45 | 124.8 | 37.070 | Scabrosemineum |
| G. pugionacanthum | 19.97 | 42.222 | 37.807 | 10 | 3.5 | 5 | 25.36 | 1.17 | 10.25 | 6.003 | 1.54 | 36.969 | 45.237 | 35.4 | 19.2 | 258 | 32.244 | Scabrosemineum |
| G. pugionacanthum | 26.651 | 41.399 | 31.949 | 11 | 3 | 5 | 30.95 | 2.05 | 11.503 | 3.163 | 0.738 | 52.217 | 32.38 | 19.6 | 31.6 | 131.6 | 28.076 | Scabrosemineum |
| G. hybopleurum | 14.147 | 22.239 | 63.614 | 13 | 4.5 | 9 | 16.76 | 0.85 | 3.289 | 5.986 | 0.154 | 12.021 | 78.551 | 21.2 | 42.8 | 117.6 | 23.251 | Scabrosemineum |
| G. hybopleurum | 27.975 | 28.74 | 43.285 | 14 | 4 | 7 | 32.98 | 1..5 | 4.061 | 5.425 | 0.315 | 29.891 | 60.309 | 22 | 38.8 | 122 | 37.728 | Scabrosemineum |
| G. hybopleurum | 21.264 | 30.582 | 48.154 | 11 | 4 | 9 | 28.33 | 1.61 | 8.764 | 3.132 | 0.621 | 14.029 | 73.453 | 18.4 | 43.6 | 122.8 | 33.121 | Scabrosemineum |
| G. hybopleurum | 23.029 | 36.966 | 40.005 | 11 | 4 | 7 | 13.82 | 1.11 | 5.26 | 4.901 | 0.078 | 45.993 | 43.767 | 24.8 | 47.6 | 105.6 | 28.296 | Scabrosemineum |
| G. hybopleurum | 20.069 | 30.617 | 49.314 | 10 | 4.5 | 5 | 12.15 | 1.02 | 1.372 | 5.508 | 0.726 | 25.098 | 67.297 | 20.8 | 46.4 | 129.2 | 29.173 | Scabrosemineum |
| G. oenanthemum | 24.884 | 36.63 | 38.486 | 10 | 4 | 8 | 24.69 | 1.15 | 10.894 | 4.108 | 0.004 | 27.889 | 57.105 | 28.8 | 23.6 | 118.4 | 30.928 | Scabrosemineum |
| G. oenanthemum | 23.079 | 11.145 | 65.776 | 11 | 3 | 7 | 23.94 | 1.86 | 11.733 | 3.019 | 1.304 | 36.314 | 47.63 | 17.2 | 32.4 | 140 | 22.812 | Scabrosemineum |
| G. oenanthemum | 16.091 | 17.775 | 66.134 | 10 | 3 | 8 | 18.94 | 1.72 | 9.708 | 3.703 | 1.892 | 37.598 | 47.099 | 22 | 34 | 110.4 | 28.076 | Scabrosemineum |
| G. oenanthemum | 13.417 | 17.727 | 68.856 | 10 | 3 | 7 | 24.20 | 1.8 | 7.808 | 1.974 | 0.649 | 28.972 | 60.597 | 17.6 | 38.8 | 116.4 | 25.225 | Scabrosemineum |
| G. oenanthemum | 17.309 | 17.152 | 65.538 | 11 | 3 | 7 | 21.14 | 1.61 | 9.948 | 4.611 | 1.43 | 28.425 | 55.585 | 18 | 42.8 | 146 | 33.341 | Scabrosemineum |
| G. baldianum | 16.313 | 42.447 | 41.240 | 8 | 6 | 6 | 7.79 | 0.48 | 1.735 | 13.446 | 1.485 | 39.493 | 43.84 | 12 | 29.6 | 174 | 33.341 | Gymnocalycium |
| G. baldianum | 13.729 | 27.535 | 58.735 | 7 | 11 | 7 | 7.82 | 0.5 | 3.303 | 12.453 | 2.87 | 46.562 | 34.813 | 5.2 | 26.4 | 166.4 | 31.147 | Gymnocalycium |
| G. baldianum | 25.74 | 26.195 | 48.065 | 7 | 13 | 5 | 9.25 | 0.4 | 4.584 | 9.038 | 1.808 | 49.322 | 35.249 | 8.4 | 28.8 | 138 | 42.773 | Gymnocalycium |
| G. baldianum | 18.767 | 21.509 | 59.724 | 9 | 12 | 5 | 5.65 | 0.47 | 1.247 | 9.732 | 1.521 | 54.745 | 32.755 | 9.6 | 24 | 106.8 | 38.166 | Gymnocalycium |
| G. baldianum | 22.663 | 19.347 | 57.990 | 8 | 10 | 6 | 9.12 | 0.48 | 1.601 | 11.502 | 2.233 | 39.107 | 45.557 | 10 | 31.2 | 152.8 | 31.147 | Gymnocalycium |
| G. marianae | 32.981 | 19.137 | 47.881 | 12 | 10.5 | 11 | 11.52 | 0.66 | 1.178 | 6.68 | 0.256 | 6.011 | 85.876 | 15.2 | 34 | 115.6 | 41.018 | Gymnocalycium |
| G. marianae | 28.837 | 20.751 | 50.412 | 13 | 7.3 | 8 | 10.91 | 0.89 | 7.587 | 10.406 | 1.652 | 28.363 | 51.991 | 12 | 28 | 150.4 | 64.049 | Gymnocalycium |
| G. marianae | 37.084 | 18.772 | 44.144 | 12 | 4 | 8 | 15.14 | 0.91 | 8.675 | 9.654 | 3.051 | 46.311 | 32.309 | 10 | 30 | 164.8 | 29.831 | Gymnocalycium |
| G. stellatum | 16.587 | 35.572 | 47.841 | 10 | 8.5 | 4 | 6.07 | 0.53 | 10.346 | 17.832 | 1.015 | 40.461 | 30.346 | 10 | 23.2 | 115.2 | 23.909 | Trichomosemineum |
| G. stellatum | 10.59 | 38.435 | 50.975 | 13 | 11 | 3 | 8.51 | 0.98 | 0.491 | 4.288 | 0.439 | 27.17 | 67.612 | 30.8 | 30 | 154.8 | 27.418 | Trichomosemineum |
| G. stellatum | 17.313 | 26.863 | 55.825 | 12 | 9 | 3 | 7.77 | 0.76 | 3.883 | 5.298 | 0.567 | 42.38 | 47.871 | 17.6 | 48.8 | 176.8 | 28.076 | Trichomosemineum |
| G. stellatum | 17.146 | 26.509 | 56.344 | 15 | 7 | 5 | 9.66 | 0.74 | 1.831 | 2.553 | 0.516 | 20.253 | 74.847 | 25.6 | 29.2 | 166.4 | 24.128 | Trichomosemineum |
| G. stellatum | 16.281 | 33.393 | 50.326 | 12 | 7.5 | 4 | 6.99 | 0.8 | 5.991 | 5.584 | 0.928 | 37.407 | 50.09 | 22.4 | 36 | 155.6 | 28.515 | Trichomosemineum |
References
- Arakaki, M.; Christin, P.-A.; Nyffeler, R.; Lendel, A.; Eggli, U.; Ogburn, R.M.; Spriggs, E.; Moore, M.J.; Edwards, E.J. Contemporaneous and recent radiations of the world’s major succulent plant lineages. Proc. Natl. Acad. Sci. USA 2011, 108, 8379–8384. [Google Scholar] [CrossRef] [PubMed]
- Vendramini, F.; Díaz, S.; Gurvich, D.E.; Wilson, P.J.; Thompson, K.; Hodgson, J.G. Leaf traits as indicators of resource-use strategy in floras with succulent species. New Phytol. 2002, 154, 147–157. [Google Scholar] [CrossRef]
- Diaz, S.; Hodgson, J.G.; Thompson, K.; Cabido, M.; Cornelissen, J.H.; Jalili, A.; Montserrat-Martí, G.; Grime, J.P.; Zarrinkamar, F.; Asri, Y.; et al. The plant traits that drive ecosystems: Evidence from three continents. J. Veg. Sci. 2004, 15, 295–304. [Google Scholar] [CrossRef]
- Huber, J.; Dettman, D.L.; Williams, D.G.; Hultine, K.R. Gas exchange characteristics of giant cacti species varying in stem morphology and life history strategy. Am. J. Bot. 2018, 105, 1688–1702. [Google Scholar] [CrossRef] [PubMed]
- Anderson, E.F. The Cactus Family; Timber Press: Portland, OR, USA, 2001; pp. 15–23. [Google Scholar]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Lavorel, S.; Garnier, E. Predicting changes in community composition and ecosystem functioning from plant traits: Revisiting the Holy Grail. Funct. Ecol. 2002, 16, 545–556. [Google Scholar] [CrossRef]
- Delbón, N.; Castello, L.; Ríos Villamil, A.; Cosa, M.T.; Stiefkens, L. Surviving in semi-arid environments: Functional coordination and trade-offs in shrubs from Argentina. IAWA J. 2021, 42, 172–190. [Google Scholar] [CrossRef]
- Nunes, A.; Köbel, M.; Pinho, P.; Matos, P.; de Bello, F.; Correia, O.; Branquinho, C. Which plant traits respond to aridity? A critical step to assess functional diversity in Mediterranean drylands. Agric. For. Meteorol. 2017, 239, 176–184. [Google Scholar]
- Ogburn, M.R.; Edwards, E.J. Anatomical variation in Cactaceae and relatives: Trait lability and evolutionary innovation. Am. J. Bot. 2009, 96, 391–408. [Google Scholar] [CrossRef]
- Aliscioni, N.L.; Delbón, N.; Gurvich, D.E. Spine function in Cactaceae, a review. J. Prof. Assoc. Cactus Dev. 2021, 23, 1–11. [Google Scholar]
- Gorné, L.D.; Díaz, S. Herbivory, intraspecific trait variability and back to herbivory. Oikos 2022, 6e09054. [Google Scholar] [CrossRef]
- Drezner, T.D. Variations on saguaro cactus (Carnegiea gigantea) spines length in wet and dry portions of their range. Madroño 2017, 64, 93–98. [Google Scholar] [CrossRef]
- Gurvich, D.E.; Lorenzati, M.A.; Sosa-Pivatto, M.; Bauk, K.; Barroso, F.L. Effects of long term seed storage on germination of 13 cacti species from Central Argentina. J. Arid. Environ. 2021, 185, 104382. [Google Scholar] [CrossRef]
- Tumber-Dávila, S.; Schenk, H.; Du, E.; Jackson, R. Plant sizes and shapes above- and belowground and their interactions with climate. New Phytol. 2022, 235, 1032–1056. [Google Scholar] [CrossRef]
- Nyffeler, R.; Eggli, U. Comparative Stem Anatomy and Systematics of Eriosyce sensu lato(Cactaceae). Ann. Bot. 1997, 80, 767–786. [Google Scholar] [CrossRef]
- Loza-Cornejo, S.; Terrazas, T. Epidermal and hypodermal characteristics in North American Cactoideae (Cactaceae). J. Plant Res. 2003, 116, 27–35. [Google Scholar] [CrossRef]
- Terrazas, T.; Arias, S. Comparative Stem Anatomy in the Subfamily Cactoideae. Bot. Rev. 2003, 68, 444–473. [Google Scholar] [CrossRef]
- Soffiatti, P.; Angyalossy, V. Anatomy of Brazilian Cereeae (subfamily Cactoideae, Cactaceae): Arrojadoa Britton & Rose, Stephanocereus A. Berger and Brasilicereus Backeberg. Acta Bot. Bras. 2007, 21, 813–822. [Google Scholar]
- Arias, S.; Terrazas, T. Variación en la anatomía de la madera de Pachycereus pecten-aborigium (Cactaceae). A nales del Instituto de Biología. Universidad Nacional Autónoma de México. Ser. Botánica 2001, 72, 157–169. [Google Scholar]
- Hernandez, M.; Terrazas, T.; Alvarado, D.A.; Cavazos, L.M. Los estomas de Myrtillocactus geometrizans (Mart. ex. Pfeiff.) Console (Cactaceae): Variación en su área de distribución. Rev. Fitotec. Mex. 2007, 30, 235–240. [Google Scholar] [CrossRef]
- Aquino, D.; Moreno-Letelier, A.; González-Botello, M.A.; Arias, S. The importance of environmental conditions in maintaining lineage identy in Epithelantha (Cactacea). Ecol. Evol. 2021, 11, 4520–4531. [Google Scholar] [CrossRef]
- Darling, S. Epidermis and hypodermis of the Saguaro cactus (Cereus giganteus): Anatomy and spectral properties. Amer. J. Bot. 1989, 76, 1698–1706. [Google Scholar] [CrossRef]
- Charles, G.J. Gymnocalycium in Habitat and Culture; Graham Charles: Stamford, UK, 2008; ISBN 0956220606. [Google Scholar]
- Gurvich, D.E.; Zeballos, S.R.; Demaio, P.H. Diversity and composition of cactus species along an altitudinal gradient in the Sierras del Norte Mountains (Córdoba, Argentina). S. Afr. J. Bot. 2014, 93, 142–147. [Google Scholar] [CrossRef]
- Oyarzabal, M.; Clavijo, J.; Oakley, L.; Biganzoli, F.; Tognetti, P.; Barberis, I.; Maturo, H.M.; León, R.J.C. Vegetation units of Argentina. Ecol. Austral 2018, 028, 040–063. [Google Scholar] [CrossRef] [Green Version]
- Zarlavsky, G.E. Histología Vegetal: Técnicas Simples y Complejas, 1st ed.; Sociedad Argentina de Botánica: Buenos Aires, Argentina, 2014; pp. 1–198. [Google Scholar]
- Kraus, J.; De Sousa, H.; Rezende, M.; Castro, N.; Vecchi, C.; Luque, R. Astra Blue and Basic Fuchsin double staining of plant materials. Biotech. Histochem. 1998, 73, 235–243. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 20 June 2022).
- Bartoszek, K.; Pienaar, J.; Mostad, P.; Andersson, S.; Hansen, T.F. A phylogenetic comparative method for studying multivariate adaptation. J. Theor. Biol. 2012, 314, 204–215. [Google Scholar] [CrossRef]
- Demaio, P.; Barfuss, M.; Kiesling, R.; Till, W.; Chiapella, J. Molecular phylogeny of Gymnocalycium (Cactaceae): Assessment of alternative infrageneric systems, a new subgenus, and trends in the evolution of the genus. Am. J. Bot. 2011, 98, 1841–1854. [Google Scholar] [CrossRef]
- Ferrero, B.G. Tras una definición de las áreas protegidas. Apuntes sobre la conservación de la naturaleza en Argentina. Revista Universitaria de Geografía 2018, 27, 99–117. [Google Scholar]
- Mauseth, J.D. Water-storing and cavitation-preventing adaptations in wood of cacti. Ann. Bot. 1993, 72, 81–89. [Google Scholar] [CrossRef]
- Mauseth, J.D. Medullary bundles and the evolution of cacti. Am. J. Bot. 1993, 72, 928–932. [Google Scholar] [CrossRef]
- Sajeva, M.; Mauseth, J.D. Leaf-like structure in the photosynthetic, succulent stems of cacti. Ann. Bot. 1991, 68, 405–411. [Google Scholar] [CrossRef]
- Mauseth, J.D.; Plemons-Rodriguez, B.J. Presence of paratracheal water storage tissue does not alter vessel characters in cactus wood. Am. J. Bot. 1997, 84, 815–822. [Google Scholar] [CrossRef]
- Fahn, A.; Cutler, D.F. Xerophytes: Handbuchder Pflanzenanatomie; Gebrueder Borntraeger: Berlin/Heidelberg, Germany, 1992; Volume 13. [Google Scholar]
- Mauseth, J.D. Structure–Function Relationships in Highly Modified Shoots of Cactaceae. Ann. Bot. 2006, 98, 901–926. [Google Scholar] [CrossRef]
- Terrazas Salgado, T.; Mauseth, J.D. Shoot Anatomy and Morphology. In Cacti Biology and Uses; Nobel, P.S., Ed.; University of California Press: Oakland, CA, USA, 2002; p. 23. [Google Scholar]
- Jordan, G.J.; Dillon, R.A.; Weston, P.H. Solar radiation as a factor in the evolution of scleromorphic leaf anatomy in Proteaceae. Am. J. Bot. 2005, 92, 789–796. [Google Scholar] [CrossRef]
- Goettsch, B.; Hilton-Taylor, C.; Cruz-Piñón, G.; Duffy, J.P.; Frances, A.; Hernández, H.M.; Inger, R.; Pollock, C.; Schipper, J.; Superina, M.; et al. High proportion of cactus species threatened with extinction. Nat. Plants 2015, 1, 15142. [Google Scholar] [CrossRef]
- Gurvich, D.E. The Use of Herbicides in South American Dry Forests to Control Shrub Encroachment: A New Threat to Cactus Species? A First Assessment. Haseltonia 2019, 2019, 39–42. [Google Scholar] [CrossRef]
- Dávila, P.; Téllez-Valdés, O.; Lira, R. Impact of climate change on the distribution of populations of an endemic Mexican columnar cactus in the Tehuacán-Cuicatlán Valley, Mexico. Plant Biosyst. 2013, 147, 376–386. [Google Scholar] [CrossRef]
- Pillet, M.; Goettsch, B.; Merow, C. Elevated extinction risk of cacti under climate change. Nat. Plants 2022, 8, 366–372. [Google Scholar] [CrossRef]
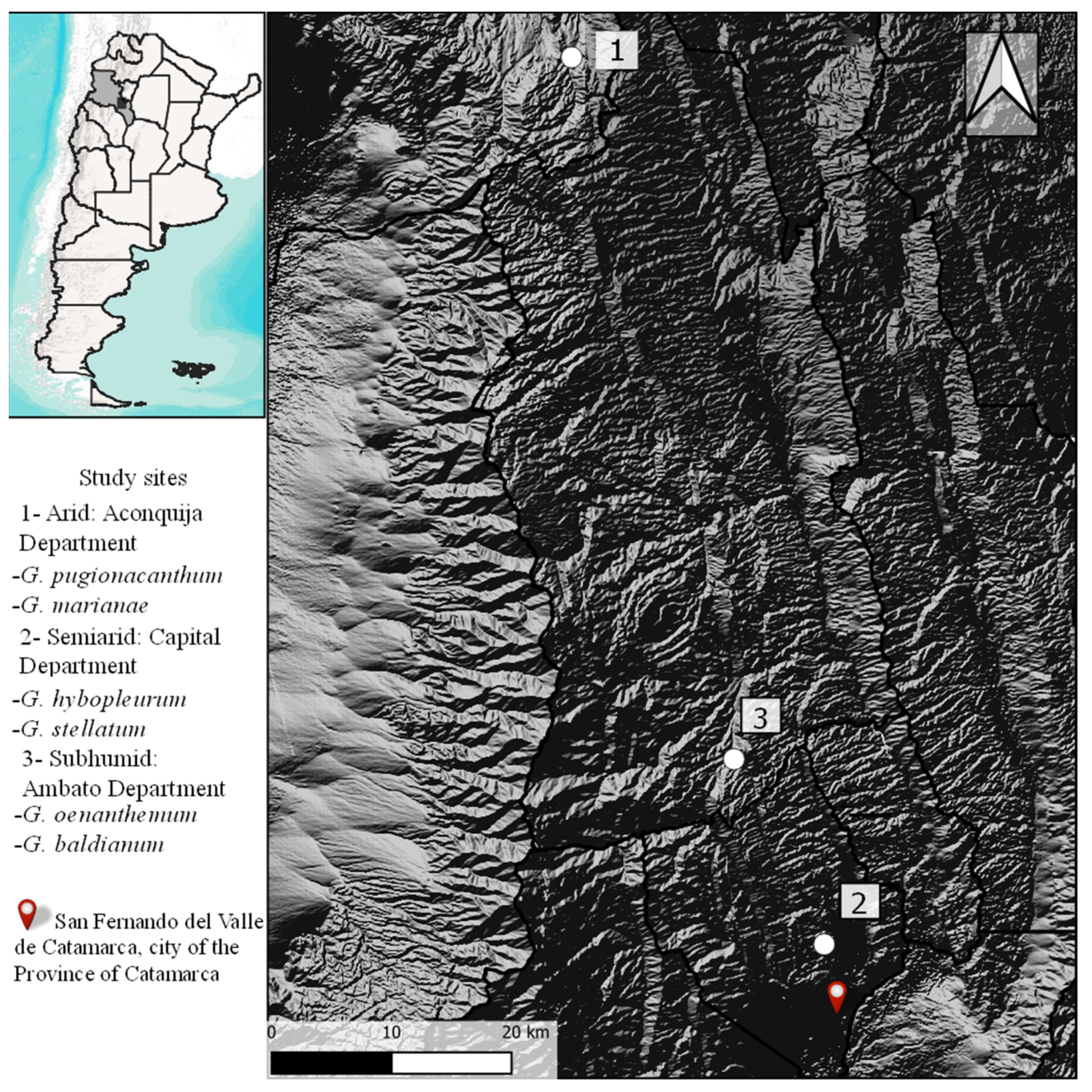

| Arid | Semiarid | Subhumid | |||||
|---|---|---|---|---|---|---|---|
| Variables | G. pugionacanthum | G. marianae | G. hybopleurum | G. stellatum | G. oenanthemum | G. baldianum | ANOVA |
| Morphological | Mean | Mean | Mean | Mean | Mean | Mean | p-Value |
| Length of aboveground stem (%) | 24.98 AB (8.79) | 32.97 B (4.12) | 21.30 A (5.01) | 15.58 A (2.82) | 18.96 A (4.84) | 19.44 A (4.82) | 0.0005 * |
| Length of underground stem (%) | 39.55 B (6.01) | 19.55 A (1.05) | 29.83 AB (5.27) | 32.15 AB (5.30) | 20.09 A (9.66) | 27.41 AB (9.05) | 0.002 * |
| Length of main root (%) | 35.49 A (3.59) | 47.48 AB (3.15) | 48.87 AB (9.05) | 52.26 B (3.68) | 60.96 B (12.63) | 53.15 B (8.15) | 0.001 * |
| Number of ribs | 10 AB (1.23) | 12.3 B (0.58) | 11.8 B (1.64) | 12.4 B (1.82) | 10.4 AB (0.55) | 7.8 A (0.84) | 0.0001 * |
| Density of areoles number/cm2 | 0.36 A (0.03) | 0.81 BC (0.36) | 0.47 AB (0.03) | 0.96 C (0.17) | 0.36 A (0.05) | 1.16 C (0.3) | <0.0001 * |
| Number of spines per areola | 5.6 AB (1.34) | 9 C (1.73) | 7.4 BC (1.67) | 3.8 A (0.84) | 7.4 BC (0.55) | 5.8 AB (0.84) | 0.0001 * |
| Spine length (mm) | 27.05 C (2.99) | 12.52 AB (2.29) | 20.81 BC (9.29) | 7.8 A (1.38) | 22.58 C (2.46) | 7.93 A (1.45) | <0.0001 * |
| Spine width (mm) | 1.8 C (0.63) | 0.82 AB (0.14) | 1.22 BC (0.32) | 0.76 AB (0.16) | 1.63 C (0.28) | 0.47 A (0.04) | <0.0001 * |
| Anatomical (µm) | |||||||
| Stomata density | 30.18 A (4.83) | 44.97 B (17.45) | 30.31 A (5.43) | 26.41 A (2.22) | 28.08 A (4.23) | 35.32 AB (5.06) | 0.017 * |
| Cuticle thickness | 22.04 B (8.52) | 12.40 AB (2.62) | 21.44 B (2.31) | 21.28 B (7.93) | 20.72 AB (4.91) | 9.04 A (2.51) | 0.005 * |
| Epidermis thickness | 40.72 (15.15) | 30.67 (3.06) | 43.84 (3.44) | 33.44 (9.71) | 34.32 (7.26) | 28.00 (2.83) | 0.073 |
| Hypodermis thickness | 161.04 (57.37) | 143.60 (25.30) | 119.44 (8.78) | 153.76 (23.36) | 126.24 (15.72) | 147.60 (26.62) | 0.274 |
| Biomass Partitioning (%) | |||||||
| Spine DW | 12.28 C (2.06) | 5.81 AB (4.05) | 4.55 AB (2.75) | 4.51 AB (3.87) | 10.02 BC (1.47) | 2.49 A (1.41) | <0.0001 * |
| Aboveground stem DW | 40.13 (12.19) | 26.9 (20.19) | 25.41 (13.72) | 33.53 (9.46) | 31.84 (4.71) | 45.85 (6.66) | 0.08 |
| Underground stem DW | 39.61 (10.08) | 56.73 (27.10) | 64.68 (13.53) | 54.15 (17.55) | 53.6 (5.98) | 38.44 (5.82) | 0.05 |
| Main root DW | 6.7 AB (2.68) | 8.91 AB (1.97) | 4.99 AB (1.11) | 7.11 AB (6.11) | 3.48 A (1.03) | 11.23 B (1.84) | 0.012 * |
| Secondary root DW | 1.28 AB (0.96) | 1.65 AB (1.40) | 0.38 A (0.28) | 0.69 AB (0.26) | 1.06 AB (0.74) | 1.98 B (0.58) | 0.027 * |
| λ (Eigenvalue) | Variance (%) | Cumulative Variance (%) | |
|---|---|---|---|
| PC1 | 4.61 | 27.11 | 27.11 |
| PC2 | 3.35 | 19.72 | 46.83 |
| PC3 | 2.31 | 13.60 | 60.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perotti, S.B.; Aliscioni, N.L.; Delbón, N.E.; Perea, M.; Hammann, A.; Gurvich, D.E. Biomass Partitioning and Morphoanatomical Traits of Six Gymnocalycium (Cactaceae) Species Occurring along a Precipitation Gradient. Diversity 2022, 14, 749. https://doi.org/10.3390/d14090749
Perotti SB, Aliscioni NL, Delbón NE, Perea M, Hammann A, Gurvich DE. Biomass Partitioning and Morphoanatomical Traits of Six Gymnocalycium (Cactaceae) Species Occurring along a Precipitation Gradient. Diversity. 2022; 14(9):749. https://doi.org/10.3390/d14090749
Chicago/Turabian StylePerotti, Solana B., Nayla L. Aliscioni, Natalia E. Delbón, Mario Perea, Ariadna Hammann, and Diego E. Gurvich. 2022. "Biomass Partitioning and Morphoanatomical Traits of Six Gymnocalycium (Cactaceae) Species Occurring along a Precipitation Gradient" Diversity 14, no. 9: 749. https://doi.org/10.3390/d14090749
APA StylePerotti, S. B., Aliscioni, N. L., Delbón, N. E., Perea, M., Hammann, A., & Gurvich, D. E. (2022). Biomass Partitioning and Morphoanatomical Traits of Six Gymnocalycium (Cactaceae) Species Occurring along a Precipitation Gradient. Diversity, 14(9), 749. https://doi.org/10.3390/d14090749