

Osteichthyan Fishes from the uppermost Norian (Triassic) of the Fuchsberg near Seinstedt, Lower Saxony (Germany)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
The Fish Material
2. Material and Methods
2.1. Anatomical Terminology
2.2. Illustrations
3. Systematic Paleontology
3.1. Teleosteomorphs
3.1.1. Seinstedtia gen. nov.
3.1.2. Seinstedtia parva gen. et sp. nov.
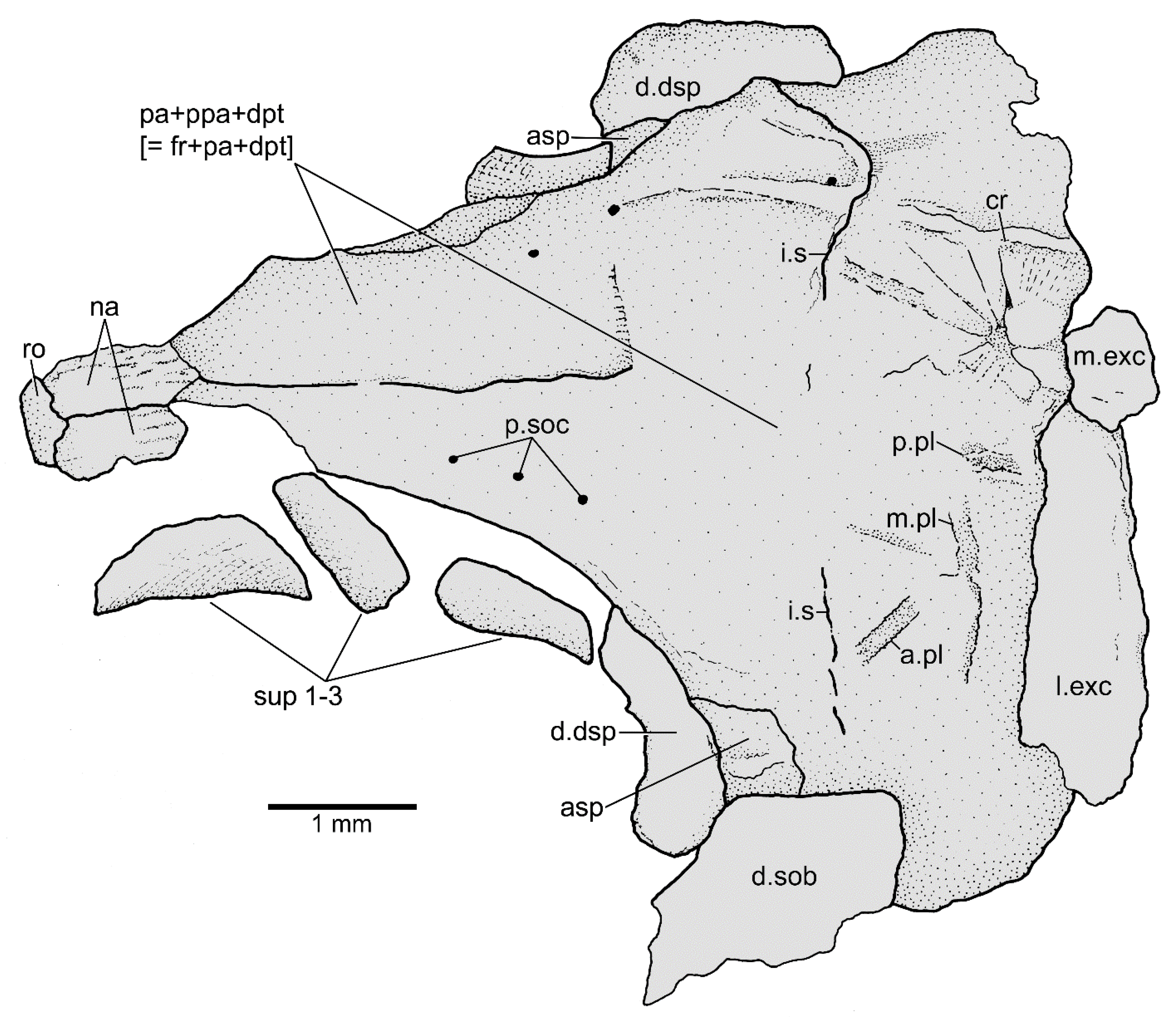
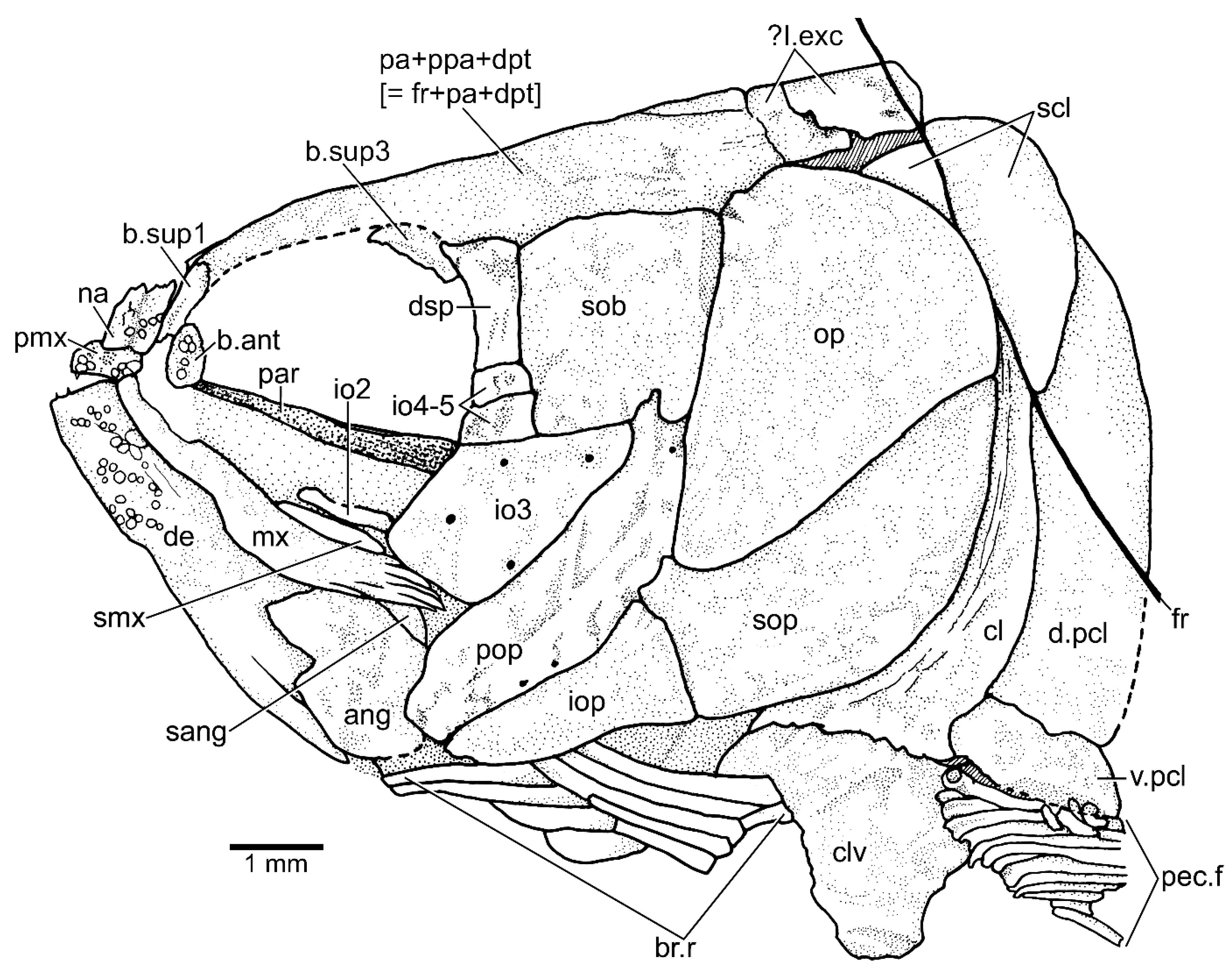
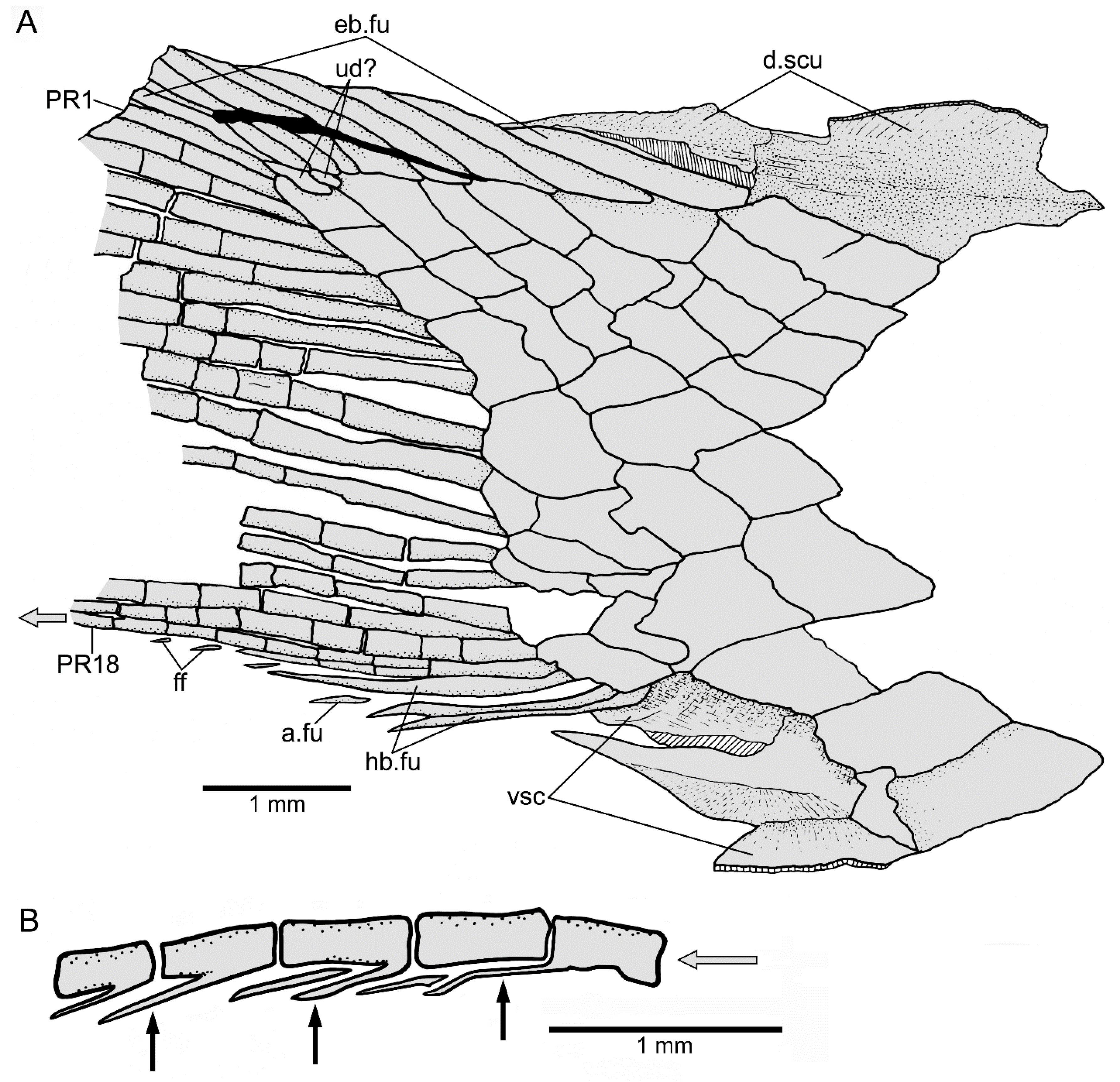
3.1.3. Description
3.1.4. Comparisons and Taxonomic Assignment
3.1.5. Miniaturization and Small Body-Sized Fishes
3.1.6. Diversity and Paleoecology
3.2. Actinistians
3.2.1. Actinistia Indet
3.2.2. Comparison and Taxonomic Remarks
3.2.3. Comments on Triassic Actinistians
3.3. Paleoecology
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- von Strombeck, A. Ueber den oberen Keuper bei Braunschweig. Z. Dtsch. Geol. Ges. 1852, 4, 54–82. [Google Scholar]
- Behrend, F. Erläuterungen zur Geologischen Karte von Preußen und Benachbarten Deutschen Ländern; Blatt Hornburg: Berlin, Germany, 1927; p. 37. [Google Scholar]
- Jüngst, H. Rät, Psilonoten-und Schlotheimienschichten im nördlichen Harzvorlande. Geol. Paläontol. Abh. 1928, 16, 1–194. [Google Scholar]
- Will, H.-J. Untersuchungen zur Stratigraphie und Genese des Oberkeupers in Nordwestdeutschland. Beih. Geol. Jb. 1969, 54, 1–240. [Google Scholar]
- Brandes, C.; Schmidt, C.; Tanner, D.C.; Winsemann, J. Paleostress pattern and salt tectonics within a developing foreland basin (north-western Subhercynian Basin, northern Germany). Int. J. Earth. Sci. 2013, 102, 2239–2254. [Google Scholar] [CrossRef]
- Benda, L. Über die Anreicherung von Uran und Thorium in Phosphoriten und Bonebeds des nördlichen Harzvorlandes. Geol. Jahrb. 1962, 80, 313–348. [Google Scholar]
- Fraas, E. Reste von Zanclodon aus dem oberen Keuper vom Langenberge bei Wolfenbüttel. Z. Dtsch. Geol. Ges. 1897, 49, 482–485. [Google Scholar]
- Hauschke, N.; Wilde, V. Paleolimulus fuchsbergensis n. sp. (Xiphosura, Merostomata) aus der oberen Trias von Nordwestdeutschland, mit einer Übersicht zur Systematik und Verbreitung rezenter Limuliden. Paläontol. Z. 1987, 61, 87–108. [Google Scholar] [CrossRef]
- Hauschke, N.; Wilde, V. Zur Verbreitung und Ökologie mesozoischer Limuliden. N. Jb. Geol. Paläont. Abh. 1991, 183, 391–411. [Google Scholar]
- Barth, G. Das Ober-Norium von Seinstedt (Niedersachsen)—Sedimentologie, Geochemie und Arthropodenfunde (Insekten, Conchostraken). Unpublished Diploma Thesis, Martin-Luther-Universität Halle-Wittenberg, Halle, Germany, 2011. [Google Scholar]
- Kumm, A. Trias und Lias. In Das Mesozoikum in Niedersachsen, 1. Abteilung, Geologie und Lagerstätten Niedersachsens; Kumm, A., Riedel, L., Schott, W., Eds.; Provinzial-Institut für Landesplanung, Landes-und Volkskunde von Niedersachsen an der Universität Göttingen: Göttingen, Germany, 1941; Volume 2, pp. 1–328. [Google Scholar]
- Appel, D. Petrographie und Genese der Sandsteine des Unter-und Mittelräts im nördlichen Harzvorland (Ostniedersachsen). Mitt. Geol. Inst. Univ. Hannover 1981, 20, 1–133. [Google Scholar]
- Hauschke, N.; Wilde, V. Fazies und Fossilführung des Fuchsberges bei Seinstedt (nördliches Harzvorland, Niedersachsen). Hall. Jb. Geowiss. B 1996, 18, 141–152. [Google Scholar]
- Barth, G.; Kozur, H.W. A latest Norian age for insect-bearing beds of the Fuchsberg und Langenberg near Seinstedt, northern foreland of the Harz Mountains (Lower Saxony, Germany). New Mex. Mus. Nat. Hist. Sci. Albuq. USA 2011, Bulletin 53, 157–165. [Google Scholar]
- Hauschke, N.; Kozur, H.W. Two new conchostracan species from the Late Triassic of the Fuchsberg, northern foreland of the Harz Mountains northeast of Seinstedt (Lower Saxony, Germany). New Mex. Mus. Nat. Hist. Sci. Albuq. USA 2011, Bulletin 53, 187–194. [Google Scholar]
- Barth, G.; Franz, M.; Heunisch, C.; Kustatscher, E.; Thies, D.; Vespermann, J.; Wolfgramm, M. Late Triassic (Norian-Rhaetian) brackish to freshwater habitats at a fluvial-dominated delta plain (Seinstedt, Lower Saxony, Germany). Palaeobio. Palaeoenv. 2014, 94, 495–528. [Google Scholar] [CrossRef]
- Barth, G.; Ansorge, J.; Brauckmann, C. First record of the genus Ipsvicia (Hemiptera: Ipsviciidae) outside Gondwana—An Australian congener from the Upper Triassic of Germany. Pol. J. Entomol. 2011, 80, 645–657. [Google Scholar] [CrossRef]
- Koch, G.; Schneider, W. Der Obere Keuper Südost-Niedersachsens in Übertage-Aufschlüssen—Fazies und Mineralbestand. Ber. Naturhist. Ges. Hannover. 1985, 128, 19–48. [Google Scholar]
- Storrs, G.W. Fossil vertebrate faunas of the British Rhaetian (latest Triassic). Zool. J. Linn. 1994, 112, 217–259. [Google Scholar] [CrossRef]
- Schultze, H.-P.; Mickle, K.E.; Poplin, C.; Hilton, E.J.; Grande, L.; Actinopterygii, I. Palaeoniscimorpha, Stem Neopterygii, Chondrostei. In Handbook of Paleoichthyology; Schultze, H.-P., Ed.; Verlag Dr. Friedrich Pfeil: München, Germany, 2021; Volume 8A, p. 299. [Google Scholar]
- Schultze, H.-P. Nomenclature and homologization of cranial bones in actinopterygians. In Mesozoic Fishes 4—Homology and Phylogeny; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2008; pp. 23–48. [Google Scholar]
- Teng, C.S.; Cavin, L.; Maxson, R.E., Jr.; Sánchez-Villagra, M.R.; Crump, J.G. Resolving homology in the face of shifting germ layer origins: Lessons from a major skull vault boundary. eLife 2019, 8, e52814. [Google Scholar] [CrossRef]
- Arratia, G. Actinopterygian postcranial skeleton with special reference to the diversity of fin ray elements, and the problem of identifying homologies. In Mesozoic Fishes 4—Homology and Phylogeny; Arratia, G., Schultze, H.-P., Wilson, M.V.H., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2008; pp. 40–101. [Google Scholar]
- Arratia, G. Identifying patterns of diversity of the actinopterygian fulcra. Acta. Zool. 2009, 90, 220–235. [Google Scholar] [CrossRef]
- Arratia, G.; Schultze, H.-P. Reevaluation of the caudal skeleton of certain actinopterygian fishes III. Salmonidae. Homologization of caudal skeletal structures. J. Morphol. 1992, 214, 187–249. [Google Scholar] [CrossRef]
- Huxley, T.H. On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proc. Zool. Soc. Lond. 1880, 1880, 649–661. [Google Scholar]
- Cope, E.D. Zittel’s Manual of Palaeontology. Am. Nat. 1887, 21, 1014–1019. [Google Scholar]
- Regan, C.T. The skeleton of Lepidosteus, with remarks on the origin and evolution of the lower Neopterygian Fishes. Proc. Zool. Soc. Lond. 1923, 93, 445–461. [Google Scholar] [CrossRef]
- Arratia, G. The sister-group of Teleostei: Consensus and disagreements. J. Vertebr. Paleontol. 2001, 21, 767–773. [Google Scholar] [CrossRef]
- Hauschke, N.; Barth, G.; Heunisch, C.; Kozur, H.W.; Wilde, V. Die Fossilvergesellschaftung in der Oberen Trias von Seinstedt (Niedersachsen). In Trias, Aufbruch in das Erdmittelalter 2; Hauschke, N., Franz, M., Bachmann, G.H., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2021; pp. 467–474. [Google Scholar]
- Jollie, M. Chordate Morphology; Reinhold Publishing: New York, NY, USA, 1962; p. 478. [Google Scholar]
- Tintori, A. Paralepidotus ornatus (AGASSIZ 1833-43): A semionotid from the Norian (Late Triassic) of Europe. In Mesozoic Fishes—Systematics and Palaeoecology; Arratia., G., Viohl, G., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 1996; pp. 167–179. [Google Scholar]
- Olsen, P.E.; McCune, A.R. Morphology of the Semionotus elegans species group from the Early Jurassic part of the Newark Supergroup of eastern North America with comments on the family Semionotidae (Neopterygii). J. Vertebr. Paleontol. 1991, 11, 269–292. [Google Scholar] [CrossRef]
- Grande, L. An empirical synthetic pattern study of gars (Lepisosteiformes) and closely related species, based mostly on skeletal anatomy: The resurrection of Holostei. Am. Soc. Ichthyol. Herpetol. 2010, 1–871. [Google Scholar]
- López-Arbarello, A. Phylogenetic Interrelationships of Ginglymodian Fishes (Actinopterygii: Neopterygii). PLoS ONE 2012, 7, e39370. [Google Scholar] [CrossRef]
- Tintori, A.; Lombardo, C. The Zorzino Limestone Actinopterygian Fauna from the Late Triassic (Norian) of the southern Alps. In The Late Triassic World; Tanner, L.H., Ed.; Topics in Geobiology; Springer: Cham, Switzerland, 2017; Volume 46, pp. 315–350. [Google Scholar] [CrossRef]
- López-Arbarello, A.; Bürgin, T.; Furrer, H.; Stockar, R. Taxonomy and phylogeny of Eosemionotus Stolley, 1920 (Neopterygii: Ginglymodi) from the Middle Triassic of Europe. Palaeontol. Electron. 2019, 22, 1–64. [Google Scholar] [CrossRef]
- Gallo, V.; Lins De Paiva, H.C.; Petra, R.; Brito, P. Lophionotus parnaibensis, sp. nov. (Semionotiformes, Semionotidae) in the Upper Jurassic of the Parnaíba Basin, northeastern Brazil. J. Vertebr. Paleontol. 2021, 41, e1994983. [Google Scholar] [CrossRef]
- Griffith, J. The Upper Triassic fishes from Polzberg bei Lunz, Austria. Zool. J. Linn. Soc. 1977, 60, 1–93. [Google Scholar] [CrossRef]
- Arratia, G.; Herzog, A. A new halecomorph fish from the Middle Triassic of Switzerland and its Systematic implications. J. Vertebr. Paleontol. 2007, 27, 838–849. [Google Scholar] [CrossRef]
- Zambelli, R. Note sui Pholidophoriformes. I. Parapholidophorus nybelini gen. n. sp. n. Rendiconti Instituto Lombardo Accademia di Science e Lettere 1975, 109, 3–49. [Google Scholar]
- Zambelli, R. Note sui Pholidophoriformes. II. Pholidoctenus serianus gen. n. sp. n. Rendiconti della Accademia Nazionale dei Lincei 1977, 5, 101–123. [Google Scholar]
- Zambelli, R. Note sui Pholidophoriformes. III contributo: Pholidophorus gervasuttii sp. n. Revista del Museo Civico di Scienze Naturali “E. Caffi” 1980, 1, 5–44. [Google Scholar]
- Zambelli, R. Note sui Pholidophoriformes. IV contributo: Pholidorhynchodon malzannia gen. n. sp. n. Revista del Museo Civico di Scienze Naturali “E. Caffi” 1980, 2, 129–168. [Google Scholar]
- Zambelli, R. Note sui Pholidophoriformes. VII contributo. Eopholidophorus forojuliensis n. g., n. sp. Gortania Atti Mus. Friul. Stor. Nat. 1990, 11, 63–76. [Google Scholar]
- Arratia, G. Morphology, taxonomy, and phylogeny of Triassic pholidophorid fishes (Actinopterygii, Teleostei). J. Vertebr. Paleontol. 2013, 33, 1–138. [Google Scholar] [CrossRef]
- Arratia, G. New Triassic teleosts (Actinopterygii, Teleosteomorpha) from northern Italy and their phylogenetic relationships among the most basal teleosts. J. Vertebr. Paleontol. 2017, 37, e1312690. [Google Scholar] [CrossRef]
- Tintori, A. The actinopterygian fish Prohalecites from the Triassic of Northern Italy. Paleontology 1990, 33, 155–174. [Google Scholar]
- Tintori, A.; Sun, Z.-Y.; Lombardo, C.; Jiang, D.-Y.; Sun, Y.-L.; Rusconi, M.; Hao, W.-C. New specialized basal neopterygians (Actinopterygii) from Triassic of the Tethys realm. Geol. Insubr. 2007, 10, 13–20. [Google Scholar]
- Arratia, G. The outstanding suction-feeder Marcopoloichthys furreri new species (Actinopterygii) from the Middle Triassic Tethys Realm of Europe and its implications for early evolution of neopterygian fishes. Mitt. Mus. Nat. Berl. Foss. Rec. 2022, 25, 231–261. [Google Scholar] [CrossRef]
- Bürgin, T. Basal ray-finned fishes (Osteichthyes: Actinopterygii) from the Middle Triassic of Monte San Giorgio (Canton Tessin, Switzerland): Systematic palaeontology with notes on functional morphology and palaeoecology. Abh. Schweiz. Paläontol. Ges. 1992, 114, 1–164. [Google Scholar]
- Sun, Z.-Y.; Tintori, A.; Lombardo, C.; Jiang, D.-Y. New miniature neopterygians from the Middle Triassic of Yunnan Province, South China. N. Jb. Geol. Paläont. Abh. 2016, 282, 135–156. [Google Scholar] [CrossRef]
- Shen, C.; Arratia, G. Re-description of the sexually dimorphic peltopleuriform fish Wusaichthys exquisitus (Middle Triassic, China): Taxonomic implications and phylogenetic relationships. J. Syst. Palaeontol. 2022, 19, 1317–1342. [Google Scholar] [CrossRef]
- Yuan, Z.; Xu, G.-H.; Dai, X.; Wang, F.; Liu, X.; Jia, E.; Miao, L.; Song, H. A new perleidid neopterygian fish from the Early Triassic (Dienerian, Induan) of South China, with a reassessment of the relationships of Perleidiformes. PeerJ 2022, 10, e13448. [Google Scholar] [CrossRef]
- Tintori, A.; Sun, Z.; Ni, P.; Lombardo, C.; Jiang, D.; Motani, R. Oldest stem Teleostei from the late Ladinian (Middle Triassic) of southern China. Riv. Ital. Paleontol. Stratigr. 2015, 121, 285–296. [Google Scholar]
- Schultze, H.-P.; Kriwet, J. Fische der Germanischen Trias. In Trias, Aufbruch in das Erdmittelalter 1; Hauschke, N., Franz, M., Bachmann, G.H., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2021; pp. 317–323. [Google Scholar]
- Romer, A.S. Herpetichthyes, Amphibioidei, Choanichthyes or Sarcopterygii? Nature 1955, 176, 126. [Google Scholar] [CrossRef]
- Cope, E.D. Contribution to the ichthyology of the Lesser Antilles. Trans. Am. Philos. Soc. 1871, 14, 445–483. [Google Scholar] [CrossRef]
- Schaeffer, B. The Triassic coelacanth fish Diplurus, with observations on the evolution of the Coelacanthini. Bull. Am. Mus. Nat. Hist. 1952, 99, 25–78. [Google Scholar]
- Mondéjar-Fernández, J.; Meunier, F.J.; Cloutier, R.; Clément, G.; Laurin, M. A microanatomical and histological study of the scales of the Devonian sarcopterygian Miguashaia bureaui and the evolution of the squamation in coelacanths. J. Anat. 2021, 239, 451–478. [Google Scholar] [CrossRef]
- Geng, B.H.; Zhu, M.; Jin, F. A revision and phylogenetic analysis of Guizhoucoelacanthus (Sarcopterygii, Actinistia) from the Triassic of China. Vert. PalAs. 2009, 47, 165–177. [Google Scholar]
- Schaeffer, B. Late Triassic fishes from the western United States. Bull. Am. Mus. Nat. Hist. 1967, 135, 285–342. [Google Scholar]
- Forey, P.L. History of the Coelacanth Fishes; Chapman & Hall: London, UK, 1998; p. 419. [Google Scholar]
- Cloutier, R.; Forey, P.L. Diversity of extinct and living actinistian fishes (Sarcopterygii). Environ. Biol. Fishes. 1991, 32, 59–74. [Google Scholar] [CrossRef]
- Schultze, H.-P. Mesozoic sarcopterygians. In Mesozoic Fishes 3—Systematics, Paleoenvironments and Biodiversity; Arratia, G., Tintori, A., Eds.; Verlag Dr. Friedrich Pfeil: München, Germany, 2004; pp. 463–492. [Google Scholar]
- Wen, W.; Zhang, Q.-Y.; Hu, S.-X.; Benton, M.J.; Zhou, C.-Y.; Tao, X.; Huang, J.-Y.; Chen, Z.-Q. Coelacanths from the Middle Triassic Luoping Biota, Yunnan, South China, with the earliest evidence of ovoviviparity. Acta Palaeontol. Pol. 2013, 58, 175–193. [Google Scholar] [CrossRef]
- Cavin, L.; Furrer, H.; Obrist, C. New coelacanth material from the Middle Triassic of eastern Switzerland, and comments on the taxic diversity of actinistans. Swiss. J. Geosci. 2013, 106, 161–177. [Google Scholar] [CrossRef]
- Ash, S.R. Geology, Paleontology, and Paleoecology of a Late Triassic Lake, Western New Mexico (Ciniza Lake Beds). B.Y.U. Geol. Stud. 1978, 25, 67–68. [Google Scholar]
- Yabumoto, Y.; Neuman, A. A coelacanth scale from the Upper Triassic Pardonet Formation, British Columbia, Canada. Paleontol. Res. 2004, 8, 337–340. [Google Scholar] [CrossRef]
- Böttcher, R. Fische des Lettenkeupers. In Der Lettenkeuper—Ein Fenster in die Zeit vor den Dinosauriern; Hagdorn, H., Schoch, R., Schweigert, G., Eds.; Staatliches Museum für Naturkunde Stuttgart und Muschelkalkmuseum Hagdorn Stadt Ingelfingen: Stuttgart, Germany, 2015; pp. 141–202. [Google Scholar]
- Schaeffer, B.; Gregory, J.T. Coelacanth fishes from the continental Triassic of the western United States. Am. Mus. Novit. 1961, 2036, 1–18. [Google Scholar]
- Hunt, A.P. A new coelacanth (Osteichthyes: Actinistia) from the continental Upper Triassic of New Mexico. New Mex. Mus. Nat. Hist. Sci. Albuq. USA 1997, Bulletin 11, 25–27. [Google Scholar]
- Martin, M. Les dipneustes et actinistians du Trias supérieur continental marocain. Stutt. Beitr. Naturkd. B. 1981, 69, 1–29. [Google Scholar]
- Dehm, R. Ein Coelacanthide aus dem Mittleren Keuper Frankens. N. Jb. Geol. Paläont. Mh. 1956, 3, 148–153. [Google Scholar]
- Dehm, R. Über einen ersten, verschollenen Coelacathiden-Fund aus dem Mittleren Keuper Frankens. N. Jb. Geol. Paläont. Mh. 1956, 11, 525–529. [Google Scholar]
- Milner, A.R.C.; Kirkland, J.I.; Birthisel, T.A. The geographic distribution and biostratigraphy of Late Triassic-Early Jurassic freshwater fish faunas of the southwestern United States. New Mex. Mus. Nat. Hist. Sci. Albuq. USA 2006, Bulletin 37, 522–529. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schultze, H.-P.; Arratia, G.; Hauschke, N.; Wilde, V. Osteichthyan Fishes from the uppermost Norian (Triassic) of the Fuchsberg near Seinstedt, Lower Saxony (Germany). Diversity 2022, 14, 901. https://doi.org/10.3390/d14110901
Schultze H-P, Arratia G, Hauschke N, Wilde V. Osteichthyan Fishes from the uppermost Norian (Triassic) of the Fuchsberg near Seinstedt, Lower Saxony (Germany). Diversity. 2022; 14(11):901. https://doi.org/10.3390/d14110901
Chicago/Turabian StyleSchultze, Hans-Peter, Gloria Arratia, Norbert Hauschke, and Volker Wilde. 2022. "Osteichthyan Fishes from the uppermost Norian (Triassic) of the Fuchsberg near Seinstedt, Lower Saxony (Germany)" Diversity 14, no. 11: 901. https://doi.org/10.3390/d14110901
APA StyleSchultze, H.-P., Arratia, G., Hauschke, N., & Wilde, V. (2022). Osteichthyan Fishes from the uppermost Norian (Triassic) of the Fuchsberg near Seinstedt, Lower Saxony (Germany). Diversity, 14(11), 901. https://doi.org/10.3390/d14110901