
Short-Lived Aggregations of Filograna/Salmacina Tube Worms in the Gulf of Oman

 , , , and
, , , and 
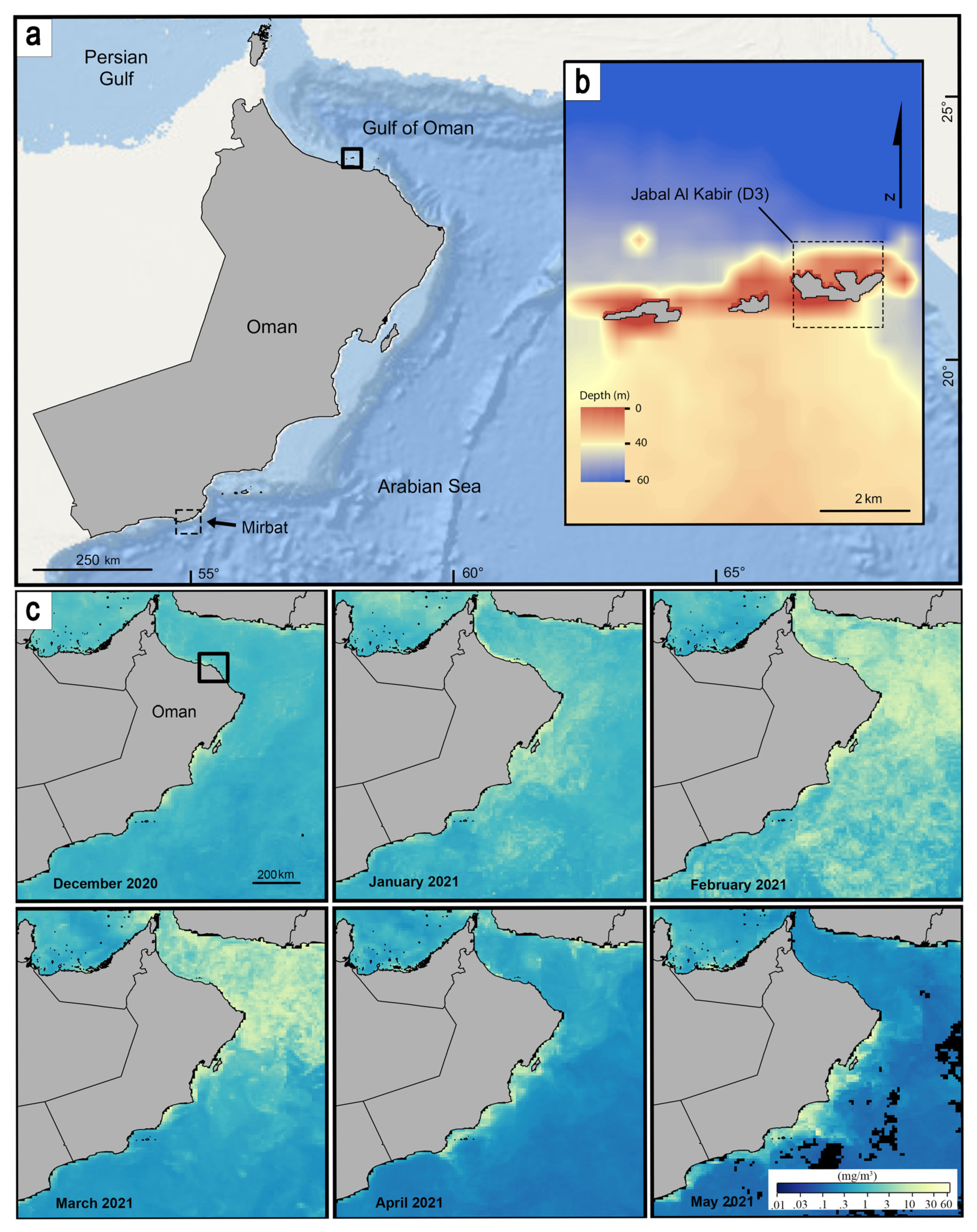{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoana, D.-P.-R.; Jose Antonio, D.-O.-C.; Angel, L.-F.; Luis Miguel, F.-V.; Francisca, G.-C.; Jose Luis, S.-L. Assessment of soft-bottom Polychaeta assemblage affected by a spatial confluence of impacts: Sewage and brine discharges. Mar. Pollut. Bull. 2009, 58, 776–782. [Google Scholar] [CrossRef]
- Elías, R.; Méndez, N.; Muniz, P.; Cabanillas, R.; Gutiérrez-Rojas, C.; Rozbaczylo, N.; Londoño-Mesa, M.H.; Gárate Contreras, P.J.; Cárdenas-Calle, M.; Villamar, F.; et al. Los poliquetos como indicadores biológicos en Latinoamérica y el Caribe. Mar. Fish. Sci. 2020, 34, 37–107. [Google Scholar] [CrossRef]
- Van der Schoot, R.J.; Hoeksema, B.W. Abundance of coral-associated fauna in relation to depth and eutrophication along the leeward side of Curaçao, southern Caribbean. Mar. Environ. Res. 2022, 181, 105738. [Google Scholar] [CrossRef]
- Tovar-Hernández, M.A.; Méndez, N.; Villalobos-Guerrero, T.F. Fouling polychaete worms from the Southern Gulf of California: Sabellidae and Serpulidae. Syst. Biodivers. 2009, 7, 319–336. [Google Scholar] [CrossRef]
- Musco, L.; Licciano, M.; Giangrande, A. Diversity and distribution of Sabellida (Annelida) under protection regimes. Water 2021, 13, 1491. [Google Scholar] [CrossRef]
- Ben-Eliahu, M.N.; ten Hove, H.A. Serpulidae (Annelida: Polychaeta) from the Suez Canal—From a Lessepsian migration perspective (a Monograph). Zootaxa 2011, 2848, 1. [Google Scholar] [CrossRef]
- Grosse, M.; Pérez, R.; Juan-Amengual, M.; Pons, J.; Capa, M. The elephant in the room: First record of invasive gregarious species of serpulids (calcareous tube annelids) in Majorca (western Mediterranean). Sci. Mar. 2021, 85, 15–28. [Google Scholar] [CrossRef]
- Çinar, M.E. Alien polychaete species worldwide: Current status and their impacts. J. Mar. Biol. Assoc. UK 2013, 93, 1257–1278. [Google Scholar] [CrossRef]
- Ten Hove, H.A.; van den Hurk, P. A review of Recent and fossil serpulid “reefs”; actuopalaeontology and “upper Malm” serpulid limestones in NW Germany. Geol. Mijnb. 1993, 72, 23–67. [Google Scholar]
- Ben-Eliahu, M.N.; Dafni, J. A new reef-building serpulid genus and species from the Gulf of Eilat and the Red Sea, with notes on other gregarious tubeworms from Israeli waters. Isr. J. Ecol. Evol. 1979, 28, 199–208. [Google Scholar]
- Ten Hove, H.A. Different causes of mass occurrence in serpulids. In Biology and Systematics of Colonial Organisms; Larwood, G.P., Rosen, B.R., Eds.; Academic Press: London, UK; New York, NY, USA, 1979; pp. 281–298. [Google Scholar]
- Ferrero, L.; Obenat, S.; Zárate, M.A. Mid-Holocene serpulid build-ups in an estuarine environment (Buenos Aires Province, Argentina). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 222, 259–271. [Google Scholar] [CrossRef]
- Glumac, B.; Berrios, L.; Greer, L.; Curran, H.A. Holocene tufa-coated serpulid mounds from the Dominican Republic: Depositional and diagenetic history, with comparison to modern serpulid aggregates from Baffin Bay. In Proceedings of the 11th Symposium on the Geology of the Bahamas and Other Carbonate Regions, San Salvadore, Bahamas; Gerace Research Center: San Salvadore, Bahamas, 2004; pp. 49–65. [Google Scholar]
- Moore, C.G.; Saunders, G.R.; Harries, D.B. The status and ecology of reefs of Serpula vermicularis L. (Polychaeta: Serpulidae) in Scotland. Aquat. Conserv. 1998, 8, 645–656. [Google Scholar] [CrossRef]
- Searle, M. Geology of the Oman Mountains, Eastern Arabia; Springer International Publishing: Cham, Switzerland, 2019. [Google Scholar]
- Wehe, T.; Fiege, D. Annotated checklist of the polychaete species of the seas surrounding the Arabian Peninsula: Red Sea, Gulf of Aden, Arabian Sea, Gulf of Oman, Arabian Gulf. Fauna Arabia 2002, 19, 7–238. [Google Scholar]
- Read, G.; Fauchald, K. World Polychaeta Database. Available online: https://www.marinespecies.org/polychaeta (accessed on 28 August 2022).
- Simon, C.A.; Helene van Niekerk, H.; Burghardt, I.; ten Hove, H.A.; Kupriyanova, E.K. Not out of Africa: Spirobranchus kraussii (Baird, 1865) is not a global fouling and invasive serpulid of Indo-Pacific origin. Aquat. Invasions 2019, 14, 221–249. [Google Scholar] [CrossRef]
- Pazoki, S.; Rahimian, H.; Struck, T.H.; Katouzian, A.R.; Kupriyanova, E.K. A New Species of the Spirobranchus kraussii-Complex (Annelida, Serpulidae) from the Persian Gulf and Gulf of Oman. Zootaxa 2020, 4748, 401–430. [Google Scholar] [CrossRef]
- Kupriyanova, E.K.; Flaxman, B.; Burghardt, I. A puzzle no more: The identity of Spirobranchus tetraceros (Schmarda, 1861), Annelida, Serpulidae, is resolved. Rec. Aust. Mus. 2022; in press. [Google Scholar]
- Ten Hove, H.A.; Kupriyanova, E.K. Taxonomy of Serpulidae (Annelida, Polychaeta): The state of affairs. Zootaxa 2009, 2036, 1–126. [Google Scholar] [CrossRef]
- Miguel De Matos Nogueira, J.; ten Hove, H.A. On a new species of Salmacina Claparède, 1870 (Polychaeta: Serpulidae) from São Paulo State, Brazil. Beaufortia 2000, 50, 151–161. [Google Scholar]
- Nishi, E.; Nishihira, M. Multi-clonal pseudo-colony formation in the calcareous tube worm Salmacina dysteri (Huxley) (Serpulidae, Polychaeta). Nat. Hist. Res. 1997, 4, 93–100. [Google Scholar]
- Pernet, B. Escape hatches for the clonal offspring of serpulid polychaetes. Biol. Bull. 2016, 200, 107–117. [Google Scholar] [CrossRef]
- Hanson, J. Formation and breakdown of serpulid tubes. Nature 1948, 161, 610–611. [Google Scholar] [CrossRef]
- Ingrosso, G.; Abbiati, M.; Badalamenti, F.; Bavestrello, G.; Belmonte, G.; Cannas, R.; Benedetti-Cecchi, L.; Bertolino, M.; Bevilacqua, S.; Bianchi, C.N.; et al. Mediterranean bioconstructions along the Italian coast. Adv. Mar. Biol. 2018, 79, 61–136. [Google Scholar] [PubMed]
- Dalyell, J.G. The Powers of the Creator Displayed in the Creation; or, Observations on Life Amidst the Various Forms of the Humbler Tribes of Animated Nature; Volume I-III, J. van Voorst: London, UK, 1853. [Google Scholar]
- Bianchi, C.N. Policheti Serpuloidei. Guide per il Riconoscimento delle Specie Animali delle Acque Lagunari e Costiere Italiane. 5. Collana del Progetto Finalizzato “Promozione della Qualità Dell’ambiente”, Serie AQ/1/96; CNR: Roma, Italy, 1981; pp. 1–187. [Google Scholar]
- Enrichetti, F.; Dominguez-Carrio’, C.; Toma, M.; Bavestrello, G.; Betti, F.; Canese, S.; Bo, M. Megabenthic communities of the Ligurian deep continental shelf and shelf break (NW Mediterranean Sea). PLoS ONE 2019, 14, e0223949. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C. Serpuloidea (Annelida: Polychaeta) from Milos, an island in the Aegean Sea with submarine hydrothermalism. J. Mar. Biol. Assoc. UK 2000, 80, 259–269. [Google Scholar] [CrossRef]
- Chan, B. Climate Change Impacts on the Serpulid Tubeworm Hydroides elegans: A Biomineralization Perspective. Ph.D. Thesis, University of Hong Kong, Pokfulam, Hong Kong, China, 2013. [Google Scholar] [CrossRef]
- Aliani, S.; Bianchi, C.N.; de Asmundis, C.; Meloni, R. Scanning electron microscope observations on the tube of the reef-forming serpulid Ficopomatus enigmaticus (Fauvel) (Annelida, Polychaeta). Boll. Zool. 1995, 62, 363–367. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Aliani, S.; Morri, C. Present-day serpulid reefs, with reference to an on-going research project on Ficopomatus enigmaticus. Publ. Serv. Géol. Luxemb. 1995, 29, 61–65. [Google Scholar]
- Hoeksema, B.W.; ten Hove, H.A. Aggregation of the reef-building tube worm Filogranella elatensis at Semporna, eastern Sabah, Malaysia. Coral Reefs 2011, 30, 839. [Google Scholar] [CrossRef]
- Holt, T.J.; Rees, E.I.; Hawkins, S.J.; Seed, R. Biogenic Reefs, An Overview of Dynamic and Sensitivity Characteristics for Conservation Management of Marine SACs; Port Erin Marine Laboratory, University of Liverpool: Liverpool, UK, 1998. [Google Scholar]
- Tudhope, A.W.; Lea, D.W.; Shimmield, G.B.; Chilcott, C.P.; Head, S. Monsoon climate and Arabian Sea coastal upwelling recorded in massive corals from southern Oman. Palaios 1996, 11, 347–361. [Google Scholar] [CrossRef]
- Claereboudt, M.R. Oman. In World Seas: An Environmental Evaluation Volume II: The Indian Ocean to the Pacific; Sheppard, C.R.C., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 25–47. [Google Scholar] [CrossRef]
- Al-Azri, A.; Piontkovski, S.; Al-Hashmi, K.; Al-Gheilani, H.; Al-Habsi, H.; Al-Khusaibi, S.; Al-Azri, N. The occurrence of algal blooms in Omani coastal waters. Aquat. Ecosyst. Health Manag. 2012, 15, 56–63. [Google Scholar] [CrossRef]
- Wei, Q.; Wang, B.; Yao, Q.; Xue, L.; Sun, J.; Xin, M.; Yu, Z. Spatiotemporal variations in the summer hypoxia in the Bohai Sea (China) and controlling mechanisms. Mar. Pollut. Bull. 2019, 138, 125–134. [Google Scholar] [CrossRef]
- Xin, M.; Wang, B.; Xie, L.; Sun, X.; Wei, Q.; Liang, S.; Chen, K. Long-term changes in nutrient regimes and their ecological effects in the Bohai Sea, China. Mar. Pollut. Bull. 2019, 146, 562–573. [Google Scholar] [CrossRef] [PubMed]
- Stachowitsch, M.; Riedel, B.; Zuschin, M.; Machan, R. Oxygen depletion and benthic mortalities: The first in situ experimental approach to documenting an elusive phenomenon. Limnol. Oceanogr. Methods 2007, 5, 344–352. [Google Scholar] [CrossRef]
- Richlen, M.L.; Morton, S.L.; Jamali, E.A.; Rajan, A.; Anderson, D.M. The catastrophic 2008–2009 red tide in the Arabian Gulf region, with observations on the identification and phylogeny of the fish-killing dinoflagellate Cochlodinium polykrikoides. Harmful Algae 2010, 9, 163–172. [Google Scholar] [CrossRef]
- Emery, N.C.; Ewanchuk, P.J.; Bertness, M.D. Competition and salt-marsh plant zonation: Stress tolerators may be dominant competitors. Ecology 2001, 82, 2471–2485. [Google Scholar] [CrossRef]
- Claereboudt, M.; Hermosa, G.; Mclean, E. Plausible cause of massive fish kills in the Gulf of Oman. In Proceedings of the First International Conference on Fisheries, Aquaculture and Environment in the Northern Indian Ocean, Muscat, Oman; 2001; pp. 123–132. [Google Scholar]
- Doney, S.C. The growing human footprint on coastal and open-ocean biogeochemistry. Science 2010, 328, 1512–1516. [Google Scholar] [CrossRef]
- Chlorophyll Concentration (1 Month–Aqua/MODIS) NASA. Available online: https://neo.gsfc.nasa.gov/view.php?datasetId=MY1DMM_CHLORA (accessed on 29 August 2022).
- Leung, J.Y.S.; Cheung, N.K.M. Feeding behaviour of a serpulid polychaete: Turning a nuisance species into a natural resource to counter algal blooms? Mar. Pollut. Bull. 2017, 115, 376–382. [Google Scholar] [CrossRef]
- Palmer, T.A.; Breaux, N.; Lebreton, B.; Guillou, G.; Beseres Pollack, J. Importance of serpulid reef to the functioning of a hypersaline estuary. Estuaries Coasts 2022, 45, 603–618. [Google Scholar] [CrossRef]
- Perry, O.; Sapir, Y.; Perry, G.; ten Hove, H.; Fine, M. Substrate selection of Christmas tree worms (Spirobranchus spp.) in the Gulf of Eilat, Red Sea. J. Mar. Biol. Assoc. UK 2018, 98, 791–799. [Google Scholar] [CrossRef]
- Hoeksema, B.W.; Wels, D.; van der Schoot, R.J.; ten Hove, H.A. Coral injuries caused by Spirobranchus opercula with and without epibiotic turf algae at Curaçao. Mar. Biol. 2019, 166, 60. [Google Scholar] [CrossRef]
- Samimi-Namin, K.; Risk, M.J.; Hoeksema, B.W.; Zohari, Z.; Rezai, H. Coral mortality and serpulid infestations associated with red tide, in the Persian Gulf. Coral Reefs 2010, 29, 509. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Samimi-Namin, K.; ten Hove, H.A.; Claereboudt, M.R.; Paulay, G.; Hoeksema, B.W. Short-Lived Aggregations of Filograna/Salmacina Tube Worms in the Gulf of Oman. Diversity 2022, 14, 902. https://doi.org/10.3390/d14110902
Samimi-Namin K, ten Hove HA, Claereboudt MR, Paulay G, Hoeksema BW. Short-Lived Aggregations of Filograna/Salmacina Tube Worms in the Gulf of Oman. Diversity. 2022; 14(11):902. https://doi.org/10.3390/d14110902
Chicago/Turabian StyleSamimi-Namin, Kaveh, Harry A. ten Hove, Michel R. Claereboudt, Gustav Paulay, and Bert W. Hoeksema. 2022. "Short-Lived Aggregations of Filograna/Salmacina Tube Worms in the Gulf of Oman" Diversity 14, no. 11: 902. https://doi.org/10.3390/d14110902
APA StyleSamimi-Namin, K., ten Hove, H. A., Claereboudt, M. R., Paulay, G., & Hoeksema, B. W. (2022). Short-Lived Aggregations of Filograna/Salmacina Tube Worms in the Gulf of Oman. Diversity, 14(11), 902. https://doi.org/10.3390/d14110902