
Improving Taxonomic Practices and Enhancing Its Extensibility—An Example from Araneology

 ,
, 

Abstract
1. Introduction
2. Methods
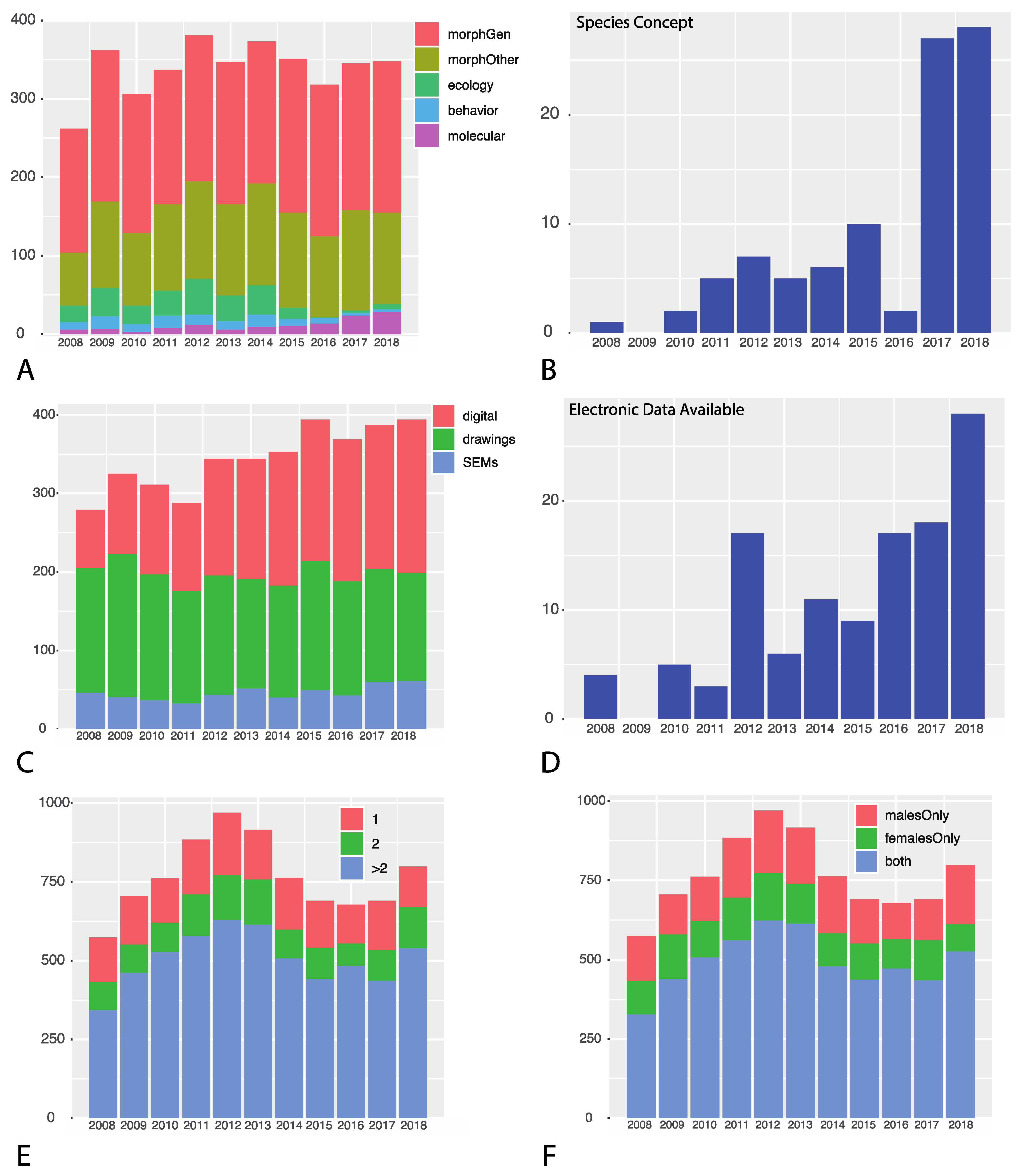
3. Results
3.1. How Integrative Is Spider Taxonomy?
3.2. Are Species Constructs Defined Conceptually?
3.3. What Data Are Being Used? What Is Available, and How Much?
4. Discussion
4.1. How Integrative Is Taxonomy?
4.2. Are Species Constructs Defined Conceptually?
4.3. Species and Data: What Is Being Used? What Is Available, and How Much?
4.4. Recommendations
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Godfray, H.C.J. Challenges for Taxonomy. Nature 2002, 417, 17–19. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, Q.D. Taxonomic Triage and the Poverty of Phylogeny. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, Q. A Taxonomic Renaissance in Three Acts. Megataxa 2020, 1, 4–8. [Google Scholar] [CrossRef][Green Version]
- Wägele, H.; Klussmann-Kolb, A.; Kuhlmann, M.; Haszprunar, G.; Lindberg, D.; Koch, A.; Wägele, J.W. The Taxonomist—An Endangered Race. A Practical Proposal for Its Survival. Front. Zool. 2011, 8, 25. [Google Scholar] [CrossRef]
- Sluys, R. The Unappreciated, Fundamentally Analytical Nature of Taxonomy and the Implications for the Inventory of Biodiversity. Biodivers. Conserv. 2013, 22, 1095–1105. [Google Scholar] [CrossRef]
- Bik, H.M. Let’s Rise up to Unite Taxonomy and Technology. PLoS Biol. 2017, 15, e2002231. [Google Scholar] [CrossRef]
- Agnarsson, I.; Kuntner, M. Taxonomy in a Changing World: Seeking Solutions for a Science in Crisis. Syst. Biol. 2007, 56, 531–539. [Google Scholar] [CrossRef]
- Bacher, S. Still Not Enough Taxonomists: Reply to Joppa et al. Trends Ecol. Evol. 2012, 27, 65–66. [Google Scholar] [CrossRef]
- Drew, L.W. Are We Losing the Science of Taxonomy? BioScience 2011, 61, 942–946. [Google Scholar] [CrossRef]
- Liu, Y.; Dietrich, C.; Braxton, S.; Wang, Y. Publishing Trends and Productivity in Insect Taxonomy from 1946 through 2012 Based on an Analysis of the Zoological Record for Four Species-Rich Families. Eur. J. Taxon. 2019, 504, 1–24. [Google Scholar] [CrossRef]
- Rodman, J.E.; Cody, J.H. The Taxonomic Impediment Overcome: NSF’s Partnerships for Enhancing Expertise in Taxonomy (PEET) as a Model. Syst. Biol. 2003, 52, 428–435. [Google Scholar] [CrossRef] [PubMed]
- Joppa, L.N.; Roberts, D.L.; Pimm, S.L. The Population Ecology and Social Behaviour of Taxonomists. Trends Ecol. Evol. 2011, 26, 551–553. [Google Scholar] [CrossRef] [PubMed]
- Bebber, D.P.; Wood, J.R.I.; Barker, C.; Scotland, R.W. Author Inflation Masks Global Capacity for Species Discovery in Flowering Plants. New Phytol. 2014, 201, 700–706. [Google Scholar] [CrossRef] [PubMed]
- Gruber, T. Academic Sell-out: How an Obsession with Metrics and Rankings Is Damaging Academia. J. Mark. High. Educ. 2014, 24, 165–177. [Google Scholar] [CrossRef]
- Bilton, D.T. What’s in a Name? What Have Taxonomy and Systematics Ever Done for Us? J. Biol. Educ. 2014, 48, 116–118. [Google Scholar] [CrossRef]
- Vink, C.; Paquin, P.; Cruickshank, R. Taxonomy and Irreproducible Biological Science. BioScience 2012, 62, 451–452. [Google Scholar] [CrossRef]
- Packer, L.; Monckton, S.K.; Onuferko, T.M.; Ferrari, R.R. Validating Taxonomic Identifications in Entomological Research. Insect Conserv. Divers. 2018, 11, 1–12. [Google Scholar] [CrossRef]
- Monckton, S.K.; Johal, S.; Packer, L. Inadequate Treatment of Taxonomic Information Prevents Replicability of Most Zoological Research. Can. J. Zool. 2020, 633–642. [Google Scholar] [CrossRef]
- Ceballos, G.; Ehrlich, P.R.; Dirzo, R. Biological Annihilation via the Ongoing Sixth Mass Extinction Signaled by Vertebrate Population Losses and Declines. Proc. Natl. Acad. Sci. USA 2017, 114, E6089–E6096. [Google Scholar] [CrossRef]
- Young, H.S.; McCauley, D.J.; Galetti, M.; Dirzo, R. Patterns, Causes, and Consequences of Anthropocene Defaunation. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 333–358. [Google Scholar] [CrossRef]
- Régnier, C.; Achaz, G.; Lambert, A.; Cowie, R.H.; Bouchet, P.; Fontaine, B. Mass Extinction in Poorly Known Taxa. Proc. Natl. Acad. Sci. USA 2015, 112, 7761–7766. [Google Scholar] [CrossRef]
- Dubois, A. Zoological Nomenclature in the Century of Extinctions: Priority vs. ‘Usage’. Org. Divers. Evol. 2010, 10, 259–274. [Google Scholar] [CrossRef]
- Bond, J. Phylogenetic Treatment and Taxonomic Revision of the Trapdoor Spider Genus Aptostichus Simon (Araneae, Mygalomorphae, Euctenizidae). ZooKeys 2012, 252, 1–209. [Google Scholar] [CrossRef]
- Troudet, J.; Vignes-Lebbe, R.; Grandcolas, P.; Legendre, F. The Increasing Disconnection of Primary Biodiversity Data from Specimens: How Does It Happen and How to Handle It? Syst. Biol. 2018, 67, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Mora, C.; Tittensor, D.P.; Adl, S.; Simpson, A.G.B.; Worm, B. How Many Species Are There on Earth and in the Ocean? PLoS Biol. 2011, 9, e1001127. [Google Scholar] [CrossRef]
- Stork, N.E. How Many Species of Insects and Other Terrestrial Arthropods Are There on Earth? Annu. Rev. Entomol. 2018, 63, 31–45. [Google Scholar] [CrossRef] [PubMed]
- World Spider Catalog. Version 20.5. Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 3 November 2021).
- Agnarsson, I.; Coddington, J.A.; Kuntner, M. Systematics: Progress in the Study of Spiders and Evolution. In Spider Research in the 21st Century: Trends and Perspectives; Siri Scientific Press: Manchester, UK, 2013; pp. 58–1111. ISBN 978-0-9574530-1-2. [Google Scholar]
- Dayrat, B. Towards Integrative Taxonomy: Integrative Taxonomy. Biol. J. Linn. Soc. 2005, 85, 407–415. [Google Scholar] [CrossRef]
- DeSalle, R.; Egan, M.G.; Siddall, M. The Unholy Trinity: Taxonomy, Species Delimitation and DNA Barcoding. Philos. Trans. R. Soc. B Biol. Sci. 2005, 360, 1905–1916. [Google Scholar] [CrossRef]
- Padial, J.M.; Miralles, A. TRehvieew Integrative Future of Taxonomy. Front. Zool. 2010, 7, 1–4. [Google Scholar] [CrossRef]
- Pomerantz, A.; Peñafiel, N.; Arteaga, A.; Bustamante, L.; Pichardo, F.; Coloma, L.A.; Barrio-Amorós, C.L.; Salazar-Valenzuela, D.; Prost, S. Real-Time DNA Barcoding in a Rainforest Using Nanopore Sequencing: Opportunities for Rapid Biodiversity Assessments and Local Capacity Building. GigaScience 2018, 7, giy033. [Google Scholar] [CrossRef]
- Mertens, J.; Van Roie, M.; Merckx, J.; Dekoninck, W. The Use of Low Cost Compact Cameras with Focus Stacking Functionality in Entomological Digitization Projects. ZooKeys 2017, 712, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Brecko, J.; Mathys, A.; Dekoninck, W.; Leponce, M.; VandenSpiegel, D.; Semal, P. Focus Stacking: Comparing Commercial Top-End Set-Ups with a Semi-Automatic Low Budget Approach. A Possible Solution for Mass Digitization of Type Specimens. ZooKeys 2014, 464, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Zamani, A.; Vahtera, V.; Sääksjärvi, I.E.; Scherz, M.D. The Omission of Critical Data in the Pursuit of ‘Revolutionary’ Methods to Accelerate the Description of Species. Syst. Entomol. 2020, 46, 1–4. [Google Scholar] [CrossRef]
- Meierotto, S.; Sharkey, M.J.; Janzen, D.H.; Hallwachs, W.; Hebert, P.D.N.; Chapman, E.G.; Smith, M.A. A Revolutionary Protocol to Describe Understudied Hyperdiverse Taxa and Overcome the Taxonomic Impediment. Dtsch. Entomol. Z. 2019, 66, 119–145. [Google Scholar] [CrossRef]
- Clerck, C. Svenska Spindlar, Uti Sina Hufvud-Slågter Indelte Samt under Några Och Sextio Särskildte Arter Beskrefne Och Med Illuminerade Figurer Uplyste; Stockholmiae: Stockholm, Sweden, 1757; pp. 1–154. [Google Scholar]
- Patzold, F.; Zilli, A.; Hundsdoerfer, A.K. Advantages of an Easy-to-Use DNA Extraction Method for Minimal-Destructive Analysis of Collection Specimens. PLoS ONE 2020, 15, e0235222. [Google Scholar] [CrossRef]
- Coates, D.J.; Byrne, M.; Moritz, C. Genetic Diversity and Conservation Units: Dealing With the Species-Population Continuum in the Age of Genomics. Front. Ecol. Evol. 2018, 6, 165. [Google Scholar] [CrossRef]
- Gómez Daglio, L.; Dawson, M.N. Integrative Taxonomy: Ghosts of Past, Present and Future. J. Mar. Biol. Assoc. UK 2019, 99, 1237–1246. [Google Scholar] [CrossRef]
- Hamilton, C.A.; Formanowicz, D.R.; Bond, J.E. Species Delimitation and Phylogeography of Aphonopelma Hentzi (Araneae, Mygalomorphae, Theraphosidae): Cryptic Diversity in North American Tarantulas. PLoS ONE 2011, 6, e26207. [Google Scholar] [CrossRef]
- Hedin, M. High-Stakes Species Delimitation in Eyeless Cave Spiders (Cicurina, Dictynidae, Araneae) from Central Texas. Mol. Ecol. 2015, 24, 346–361. [Google Scholar] [CrossRef]
- Opatova, V.; Bond, J.E.; Arnedo, M.A. Ancient Origins of the Mediterranean Trap-Door Spiders of the Family Ctenizidae (Araneae, Mygalomorphae). Mol. Phylogenet. Evol. 2013, 69, 1135–1145. [Google Scholar] [CrossRef]
- Thomas, S.M.; Hedin, M. Multigenic Phylogeographic Divergence in the Paleoendemic Southern Appalachian Opilionid Fumontana Deprehendor Shear (Opiliones, Laniatores, Triaenonychidae). Mol. Phylogenet. Evol. 2008, 46, 645–658. [Google Scholar] [CrossRef] [PubMed]
- Haszprunar, G. Species Delimitations—Not ‘Only Descriptive’. Org. Divers. Evol. 2011, 11, 249–252. [Google Scholar] [CrossRef]
- Bond, J.E.; Garrison, N.L.; Hamilton, C.A.; Godwin, R.L.; Hedin, M.; Agnarsson, I. Phylogenomics Resolves a Spider Backbone Phylogeny and Rejects a Prevailing Paradigm for Orb Web Evolution. Curr. Biol. 2014, 24, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Hormiga, G.; Giribet, G. Phylogenomic Analysis of Spiders Reveals Nonmonophyly of Orb Weavers. Curr. Biol. 2014, 24, 1772–1777. [Google Scholar] [CrossRef]
- Garrison, N.L.; Rodriguez, J.; Agnarsson, I.; Coddington, J.A.; Griswold, C.E.; Hamilton, C.A.; Hedin, M.; Kocot, K.M.; Ledford, J.M.; Bond, J.E. Spider Phylogenomics: Untangling the Spider Tree of Life. PeerJ 2016, 4, e1719. [Google Scholar] [CrossRef]
- Opatova, V.; Hamilton, C.A.; Hedin, M.; De Oca, L.M.; Král, J.; Bond, J.E. Phylogenetic Systematics and Evolution of the Spider Infraorder Mygalomorphae Using Genomic Scale Data. Syst. Biol. 2020, 69, 671–707. [Google Scholar] [CrossRef]
- Coddington, J.A.; Agnarsson, I.; Miller, J.A.; Kuntner, M.; Hormiga, G. Undersampling Bias: The Null Hypothesis for Singleton Species in Tropical Arthropod Surveys. J. Anim. Ecol. 2009, 78, 573–584. [Google Scholar] [CrossRef]
- Lim, G.S.; Balke, M.; Meier, R. Determining Species Boundaries in a World Full of Rarity: Singletons, Species Delimitation Methods. Syst. Biol. 2012, 61, 165–169. [Google Scholar] [CrossRef]
- Vogt, L.; Nickel, M.; Jenner, R.A.; Deans, A.R. The Need for Data Standards in Zoomorphology. J. Morphol. 2013, 274, 793–808. [Google Scholar] [CrossRef]
- Ramírez, M.J.; Michalik, P. The Spider Anatomy Ontology (SPD)—A Versatile Tool to Link Anatomy with Cross-Disciplinary Data. Diversity 2019, 11, 202. [Google Scholar] [CrossRef]
- Vinarski, M.V. Roots of the Taxonomic Impediment: Is the “Integrativeness” a Remedy? Integr. Zool. 2020, 15, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. Lond. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Ellis, R.; Waterton, C.; Wynne, B. Taxonomy, Biodiversity and Their Publics in Twenty-First-Century DNA Barcoding. Public Underst. Sci. 2010, 19, 497–512. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| General Descriptor | Parameters | Scoring Method |
|---|---|---|
| General information | (1) Authorship; (2) year; (3) country/region | Recorded directly |
| Species | (4) Number of new species | Exact count |
| Type of data used to delimit species | (5) Morphology—genitalic; (6) morphology—other; (7) molecular; (8) ecological; (9) behavioral | Binary scoring NO/YES (0/1) |
| Integrative | (10) Did species delimitation employ more than one data type? | Binary scoring NO/YES (0/1) |
| Species concept | (11) Was a species concept explicitly stated? (12) If species concept stated then which concept used? | (11) Binary scoring NO/YES (0/1) and (12) recorded directly |
| Illustration types used | (13) Drawings; (14) digital images; (15) scanning electron microscopy | Binary scoring NO/YES (0/1) |
| Data | (16) Are the raw data (measurements, material examined, etc.) available as a downloadable resource? | Binary scoring NO/YES (0/1) |
| Sex if specimens examined for new species description | (17) Males only; (18) females only; (19) both sexes available | Exact counts |
| Number of specimens examined for new species description | (20) 1 specimen; (21) 2 specimens; (22) 3 or more specimens | Exact counts |
| Year | # Species | # of Papers Surveyed | Genitalia | Other | Ecology | Behavior | Molecular | Integrative | Concept? |
|---|---|---|---|---|---|---|---|---|---|
| 2008 | 574 | 165 | 158 | 67 | 21 | 10 | 6 | 25 | 1 |
| 2009 | 705 | 195 | 193 | 110 | 36 | 16 | 7 | 41 | 0 |
| 2010 | 762 | 179 | 177 | 92 | 24 | 10 | 3 | 26 | 2 |
| 2011 | 884 | 177 | 171 | 110 | 32 | 16 | 8 | 41 | 5 |
| 2012 | 970 | 188 | 186 | 124 | 46 | 13 | 12 | 56 | 7 |
| 2013 | 916 | 187 | 181 | 116 | 33 | 11 | 6 | 38 | 5 |
| 2014 | 763 | 189 | 181 | 129 | 38 | 15 | 10 | 47 | 6 |
| 2015 | 691 | 205 | 196 | 121 | 14 | 9 | 11 | 27 | 10 |
| 2016 | 678 | 194 | 193 | 103 | 1 | 7 | 14 | 19 | 2 |
| 2017 | 691 | 199 | 187 | 127 | 4 | 3 | 24 | 28 | 27 |
| 2018 | 799 | 207 | 193 | 116 | 7 | 3 | 29 | 35 | 28 |
| Totals | 8433 | 2083 | 2016 | 1215 | 256 | 113 | 130 | 298 | 93 |
| Year | Drawings | Digital Images | SEM | Data Availability |
|---|---|---|---|---|
| 2008 | 159 | 74 | 46 | 4 |
| 2009 | 182 | 102 | 41 | 0 |
| 2010 | 160 | 114 | 37 | 5 |
| 2011 | 143 | 112 | 33 | 3 |
| 2012 | 152 | 148 | 44 | 17 |
| 2013 | 139 | 153 | 52 | 6 |
| 2014 | 143 | 170 | 40 | 11 |
| 2015 | 164 | 180 | 50 | 9 |
| 2016 | 145 | 181 | 43 | 17 |
| 2017 | 144 | 183 | 60 | 18 |
| 2018 | 138 | 195 | 61 | 28 |
| Totals | 1669 | 1612 | 508 | 118 |
| Year | Male Specimens Only | Female Specimens Only | Both Sexes | % Species Based on One Sex | Based on 1 Specimen | Based on 2 Specimens | Based on >2 Specimens | % Based on 2 or Fewer Specimens |
|---|---|---|---|---|---|---|---|---|
| 2008 | 139 | 107 | 328 | 42% | 140 | 90 | 344 | 40% |
| 2009 | 125 | 140 | 440 | 37% | 152 | 90 | 463 | 34% |
| 2010 | 140 | 114 | 508 | 33% | 140 | 94 | 528 | 31% |
| 2011 | 188 | 134 | 562 | 36% | 172 | 132 | 580 | 34% |
| 2012 | 197 | 149 | 624 | 35% | 197 | 142 | 631 | 35% |
| 2013 | 175 | 126 | 615 | 32% | 157 | 144 | 615 | 33% |
| 2014 | 180 | 103 | 480 | 37% | 164 | 91 | 508 | 33% |
| 2015 | 139 | 115 | 437 | 36% | 148 | 100 | 443 | 36% |
| 2016 | 113 | 93 | 472 | 30% | 123 | 71 | 484 | 29% |
| 2017 | 128 | 127 | 436 | 36% | 156 | 97 | 438 | 37% |
| 2018 | 186 | 87 | 526 | 34% | 129 | 129 | 541 | 32% |
| Totals | 1710 20% | 1295 15% | 5428 64% | 35.6% | 1678 (20%) | 1180 (14%) | 5575 66% | 34% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bond, J.E.; Godwin, R.L.; Colby, J.D.; Newton, L.G.; Zahnle, X.J.; Agnarsson, I.; Hamilton, C.A.; Kuntner, M. Improving Taxonomic Practices and Enhancing Its Extensibility—An Example from Araneology. Diversity 2022, 14, 5. https://doi.org/10.3390/d14010005
Bond JE, Godwin RL, Colby JD, Newton LG, Zahnle XJ, Agnarsson I, Hamilton CA, Kuntner M. Improving Taxonomic Practices and Enhancing Its Extensibility—An Example from Araneology. Diversity. 2022; 14(1):5. https://doi.org/10.3390/d14010005
Chicago/Turabian StyleBond, Jason E., Rebecca L. Godwin, Jordan D. Colby, Lacie G. Newton, Xavier J. Zahnle, Ingi Agnarsson, Chris A. Hamilton, and Matjaž Kuntner. 2022. "Improving Taxonomic Practices and Enhancing Its Extensibility—An Example from Araneology" Diversity 14, no. 1: 5. https://doi.org/10.3390/d14010005
APA StyleBond, J. E., Godwin, R. L., Colby, J. D., Newton, L. G., Zahnle, X. J., Agnarsson, I., Hamilton, C. A., & Kuntner, M. (2022). Improving Taxonomic Practices and Enhancing Its Extensibility—An Example from Araneology. Diversity, 14(1), 5. https://doi.org/10.3390/d14010005