
Effects of Glyphosate Application on Physiologically Integrated Clones of the Invasive Plant Carpobrotus edulis




Abstract
:1. Introduction
2. Materials and Methods
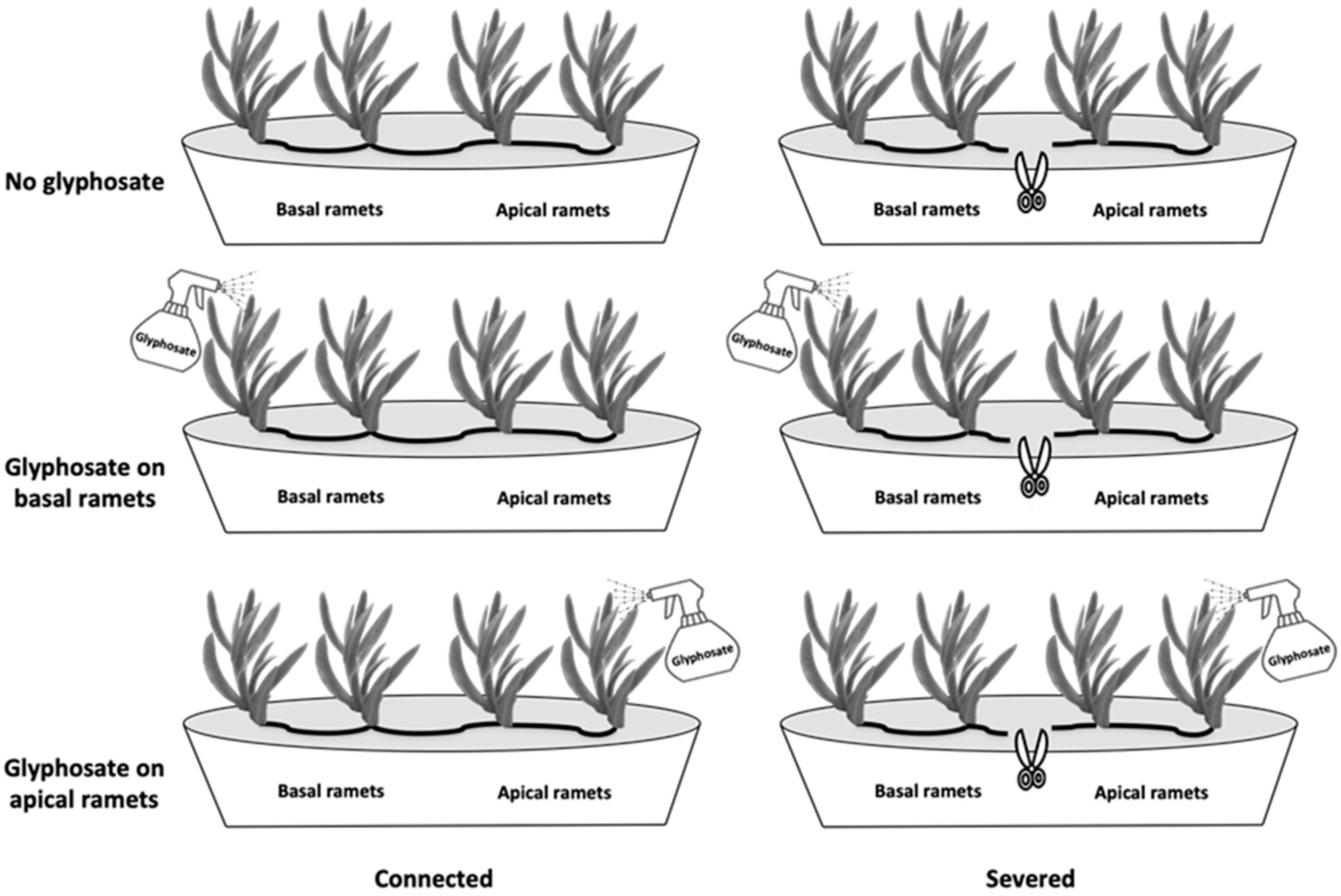
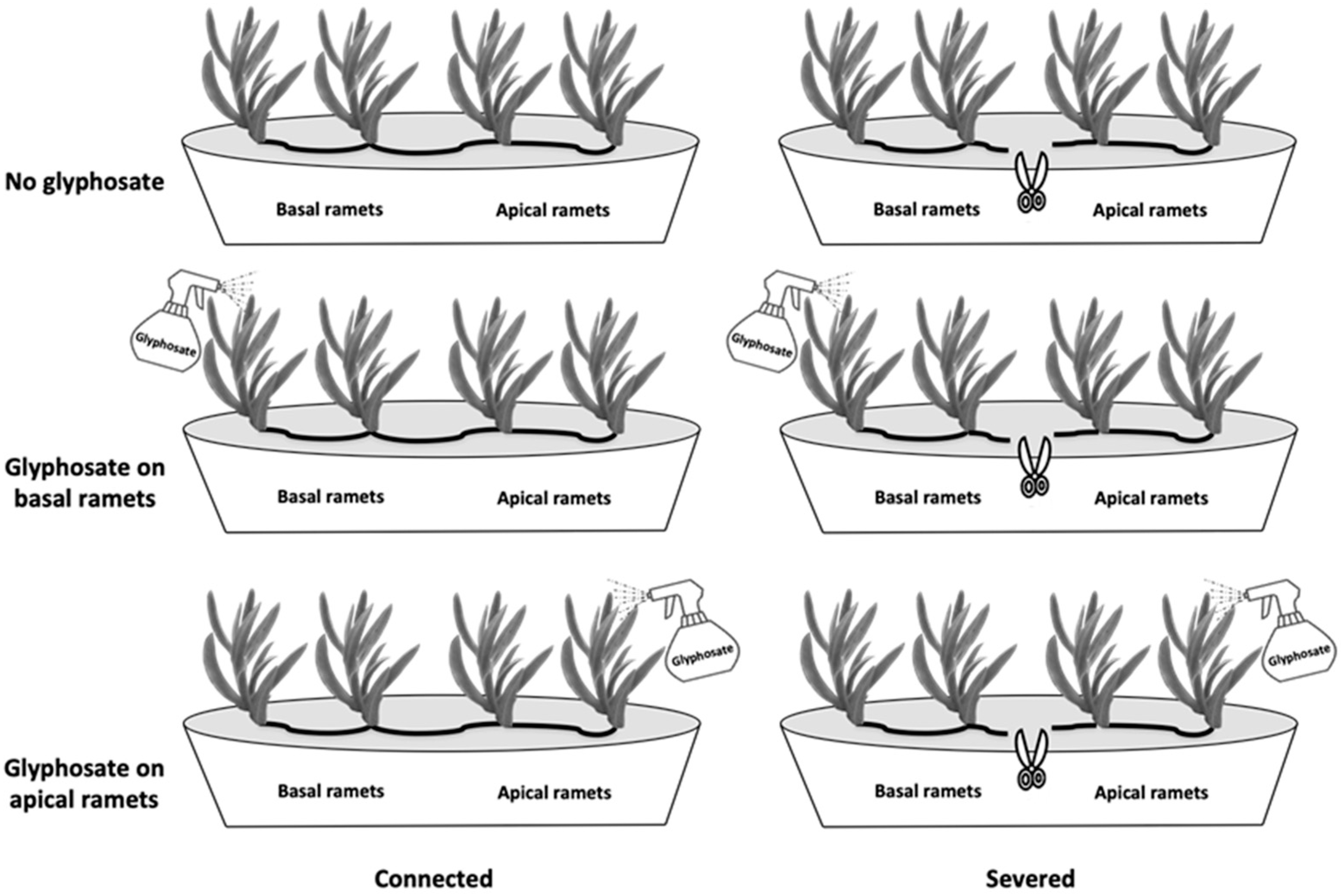
2.1. Plant Material and Experimental Design
2.2. Measurements
2.2.1. Growth and Biomass Allocation
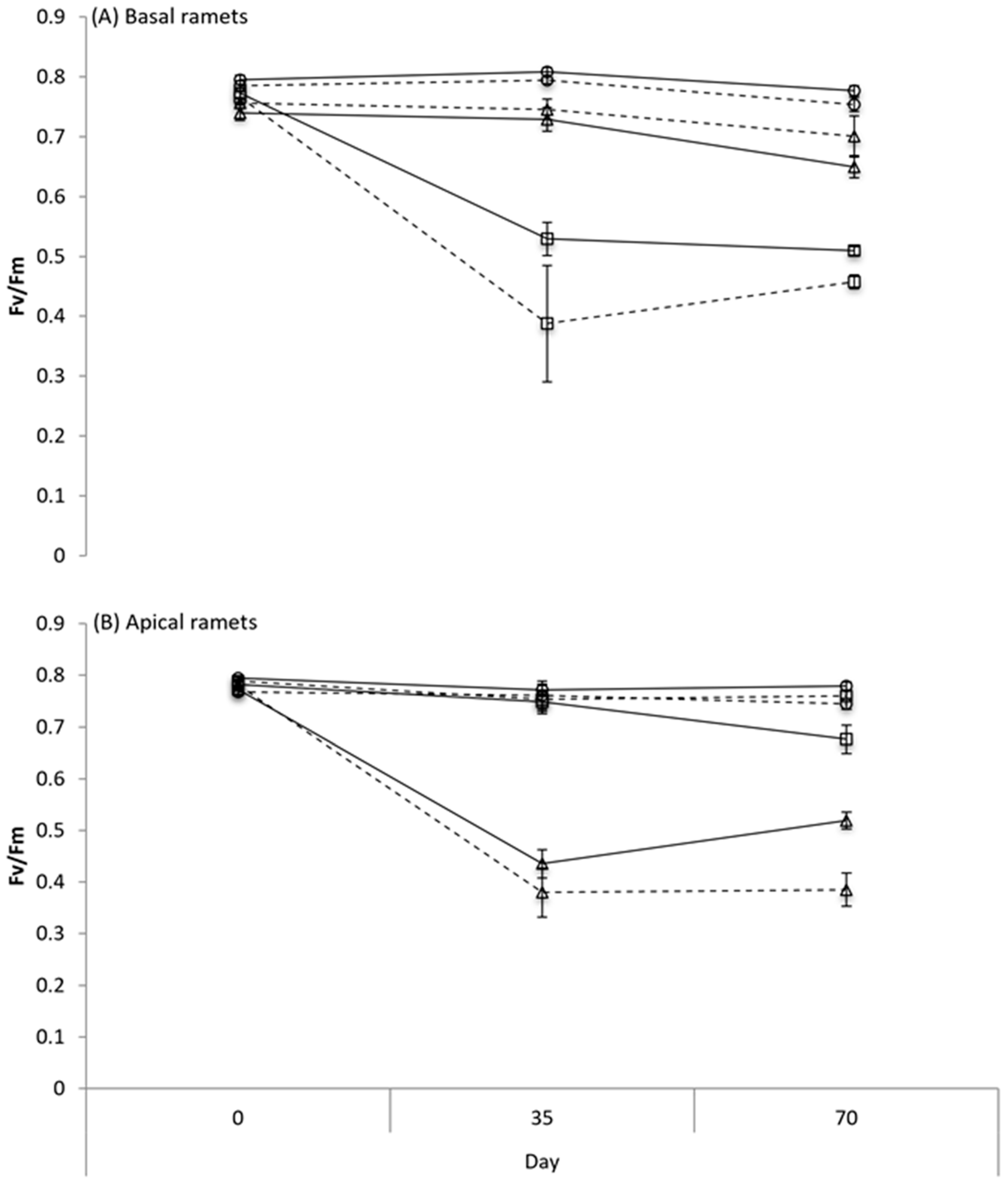
2.2.2. Chlorophyll Fluorescence
2.3. Statistical Analyses
3. Results
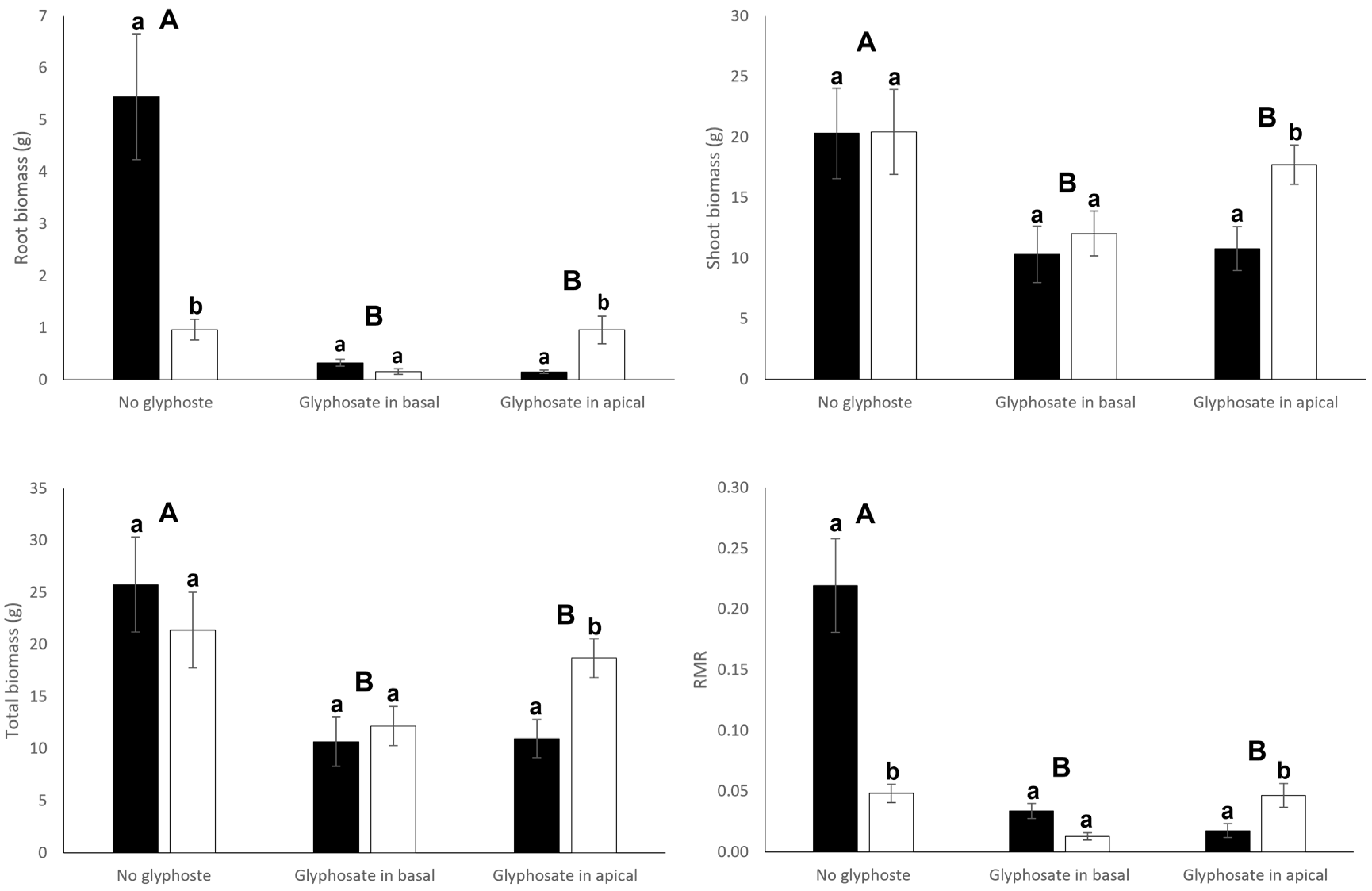
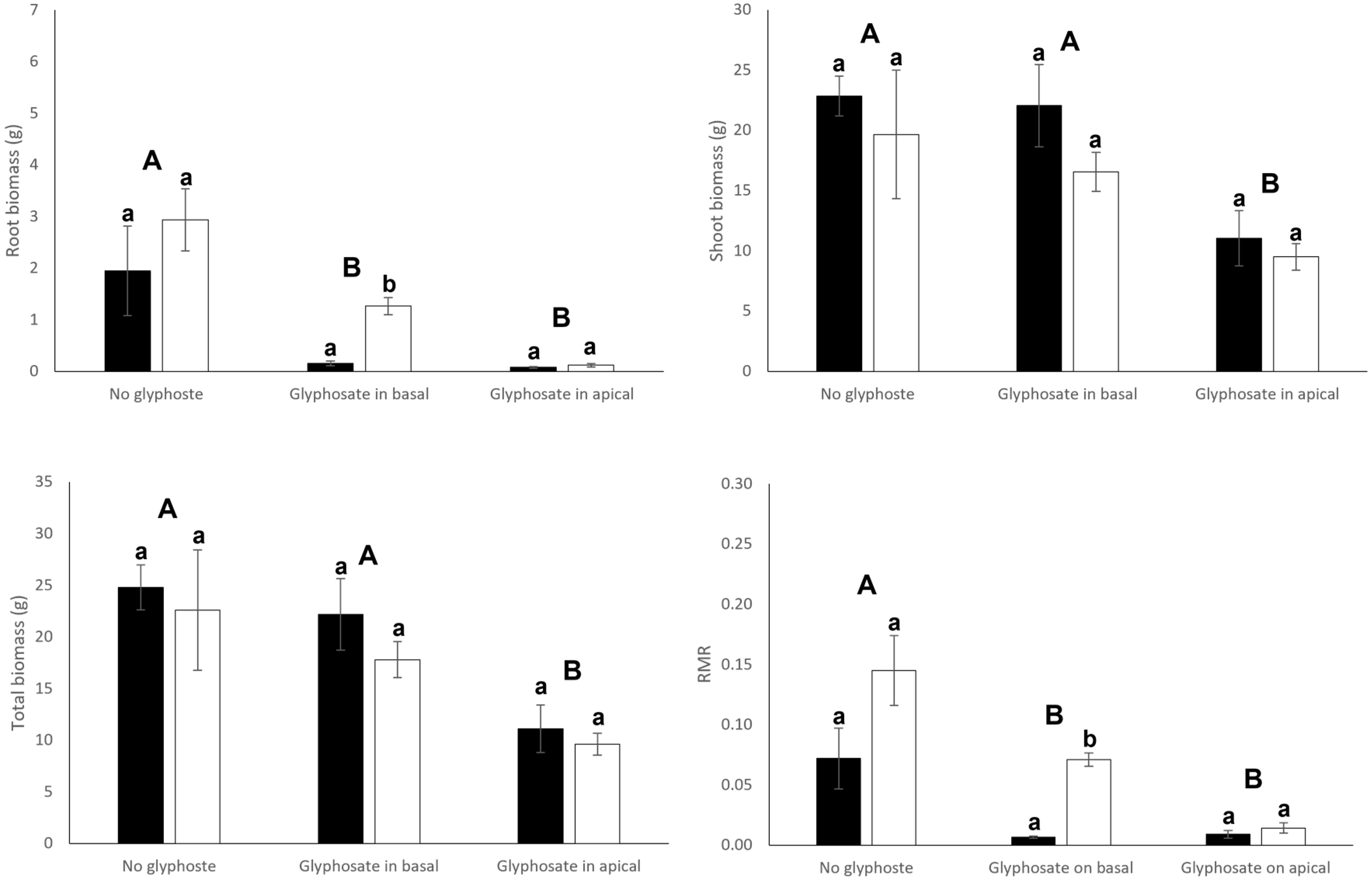
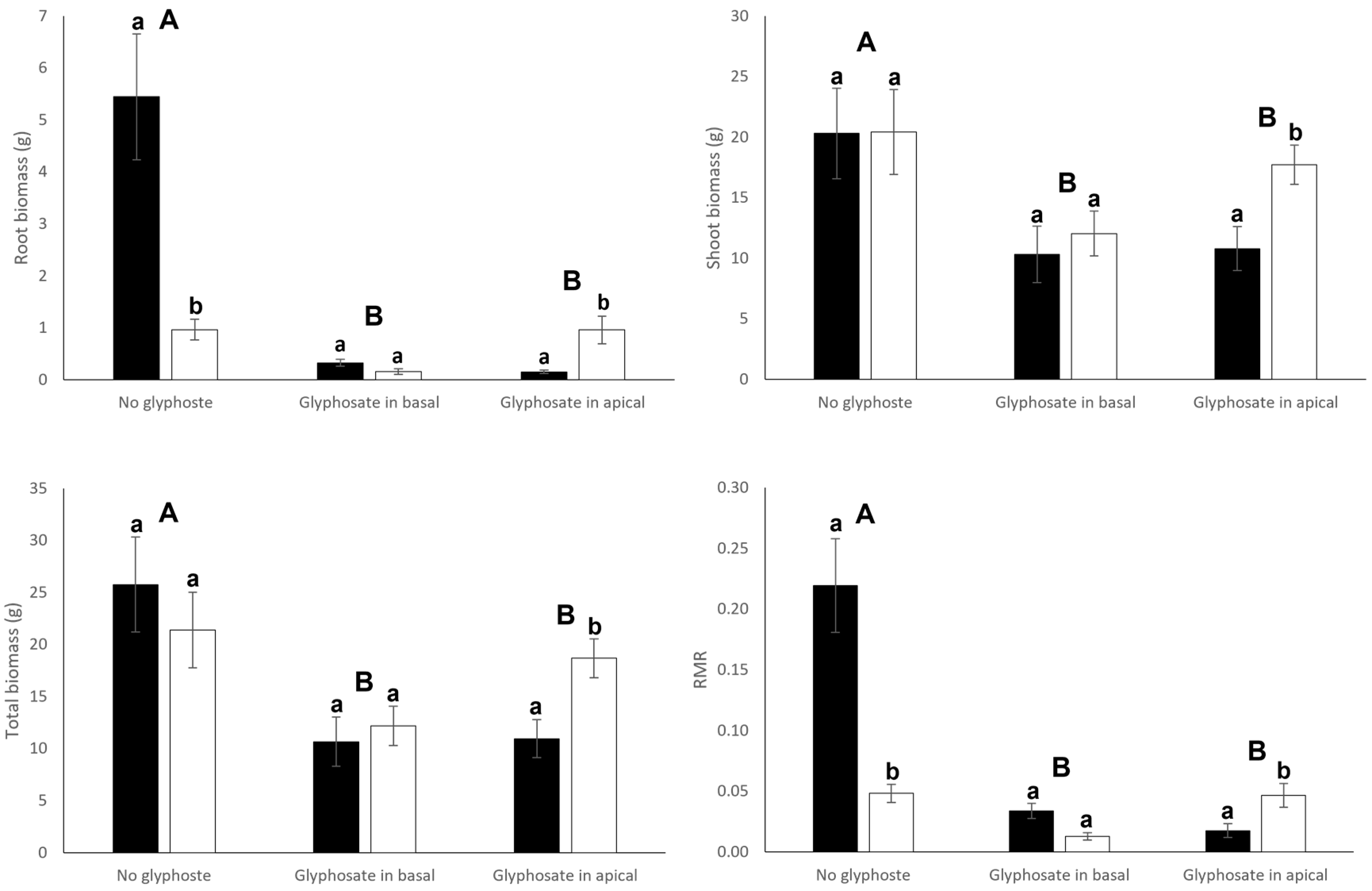
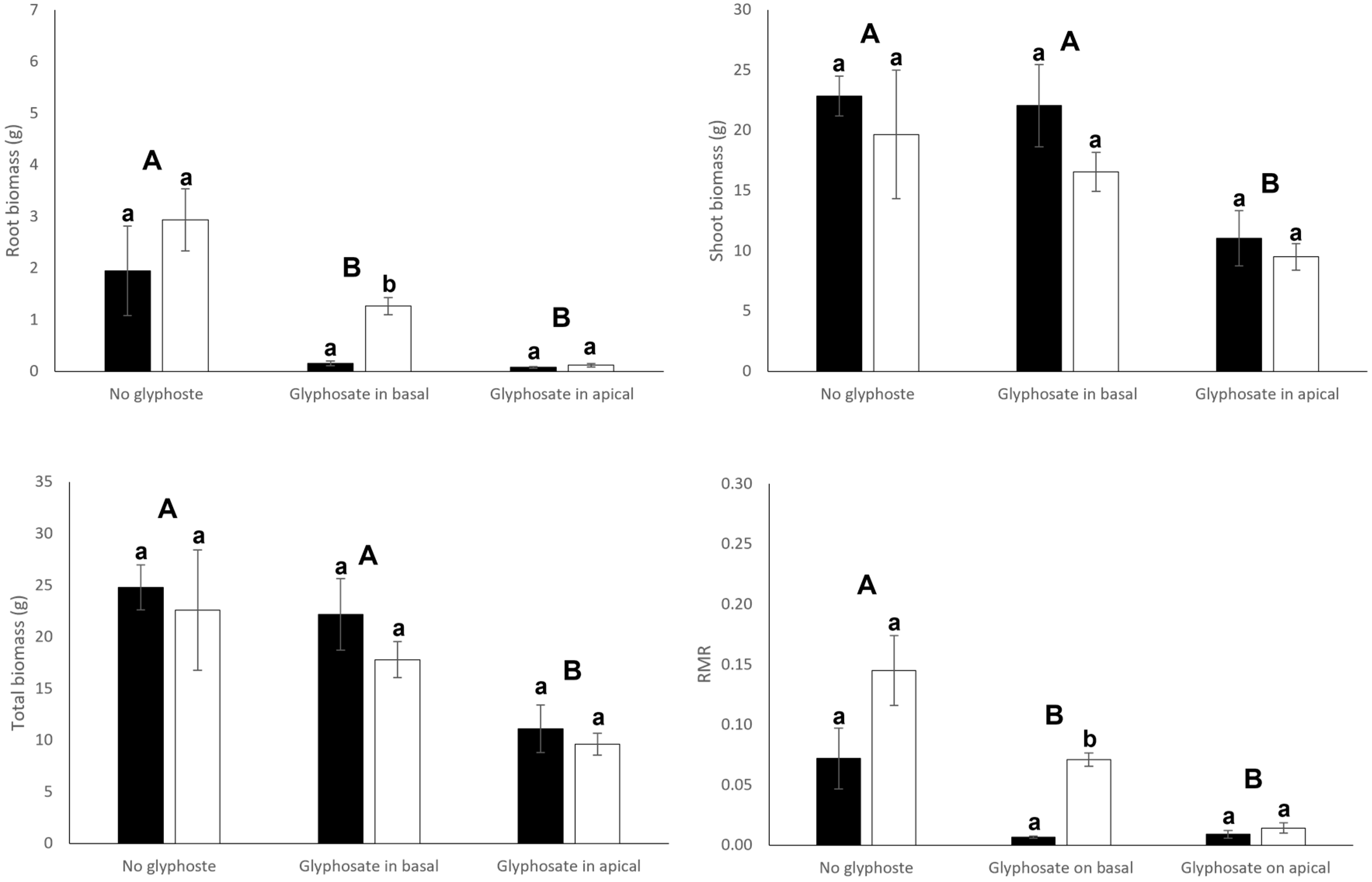
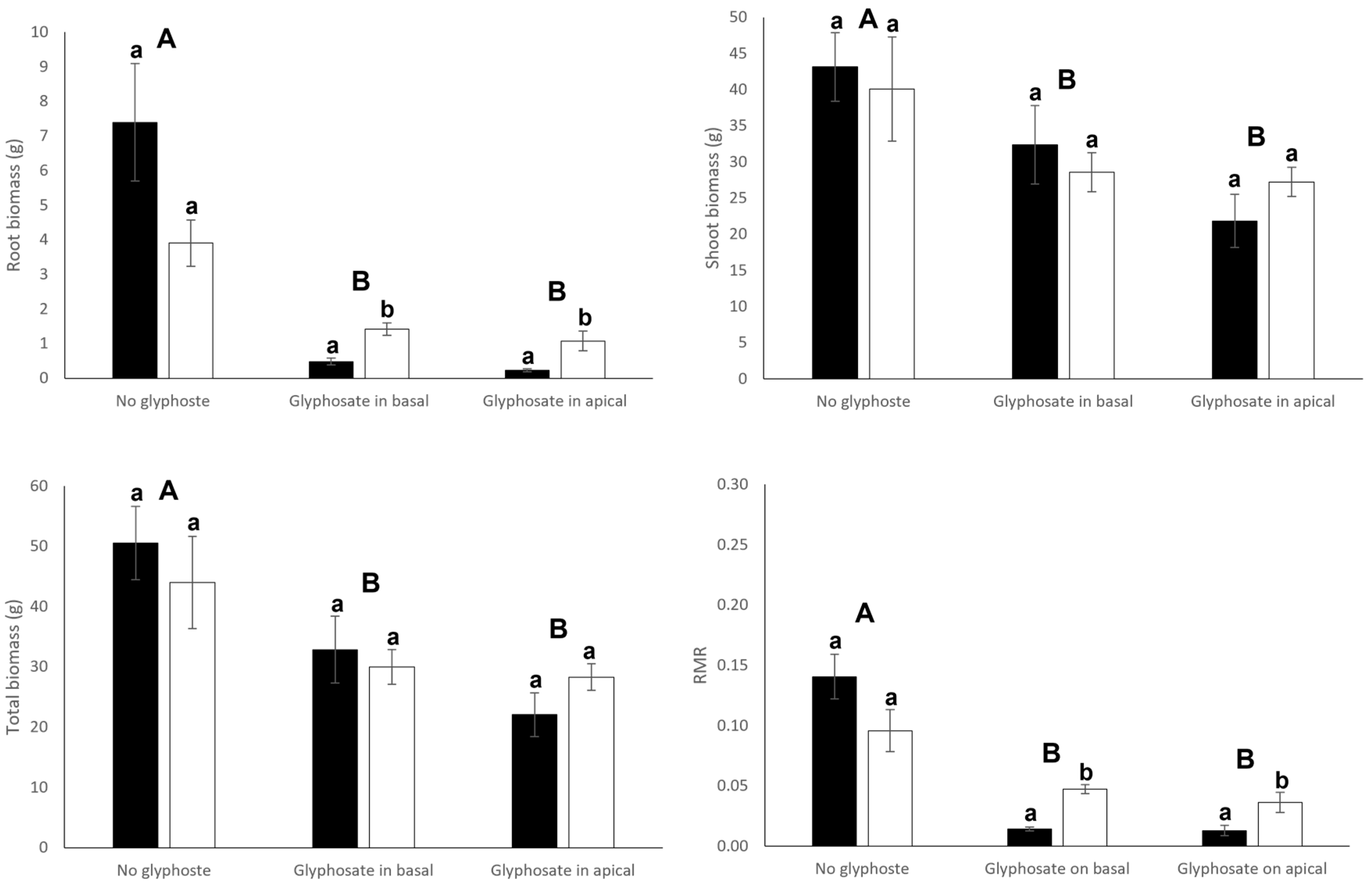
3.1. Growth and Biomass Allocation
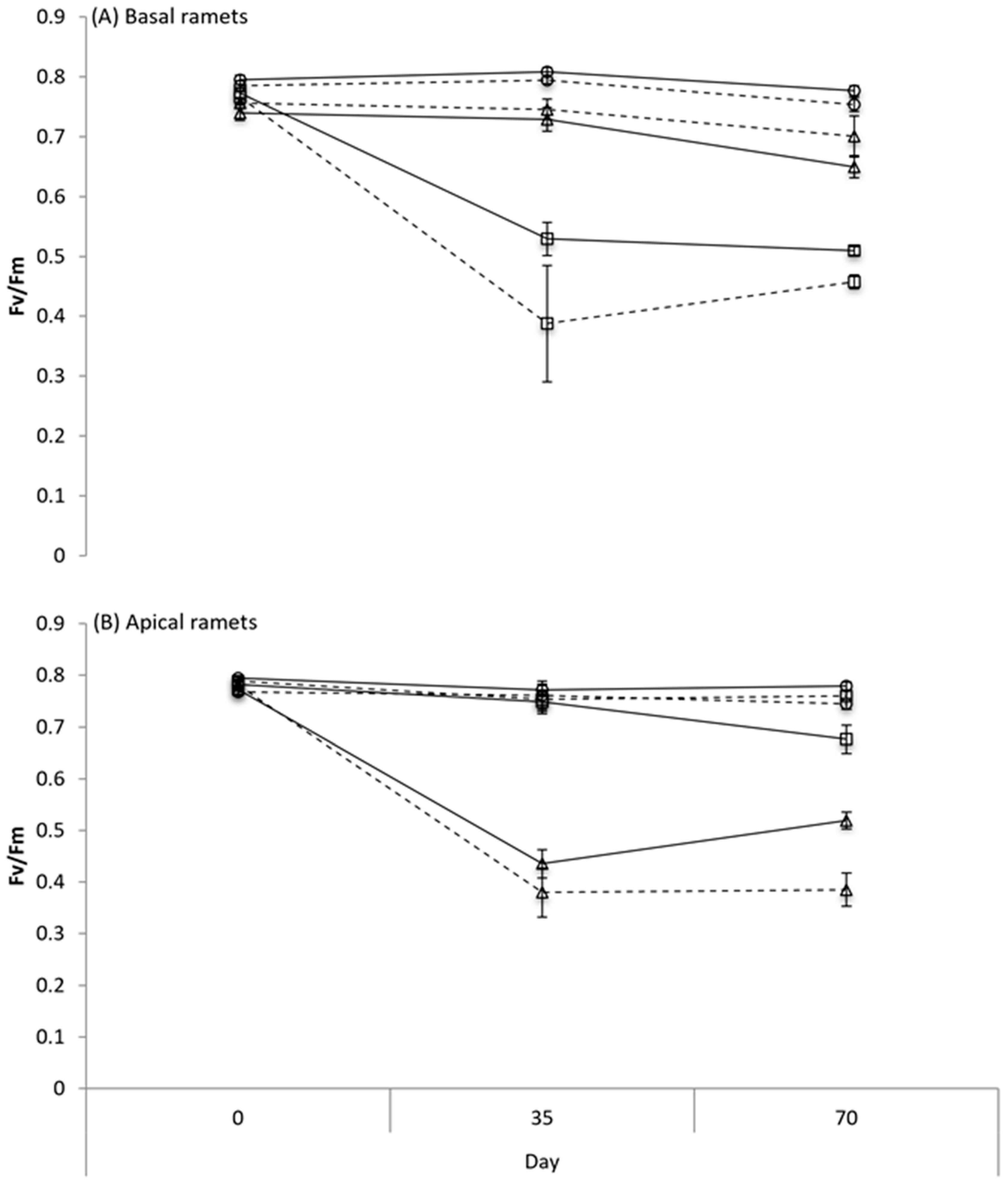
3.2. Chlorophyll Fluorescence
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martínez, M.; Psuty, N. Coastal Dunes; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Fifty years of invasion ecology—The legacy of Charles Elton. Divers. Distrib. 2008, 14, 161–168. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Strayer, D.L. Eight questions about invasions and ecosystem functioning. Ecol. Lett. 2012, 15, 1199–1210. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Pauchard, A.; Traveset, A.; Vilà, M. Linking the impacts of plant invasion on community functional structure and ecosystem properties. J. Veg. Sci. 2016, 27, 1233–1242. [Google Scholar] [CrossRef]
- Bacher, S.; Blackburn, T.M.; Essl, F.; Genovesi, P.; Heikkilä, J.; Jeschke, J.M.; Jones, G.; Keller, R.; Kenis, M.; Kueffer, C.; et al. Socio-economic impact classification of alien taxa (SEICAT). Methods Ecol. Evol. 2018, 9, 159–168. [Google Scholar] [CrossRef]
- Robertson, P.A.; Mill, A.; Novoa, A.; Jeschke, J.M.; Essl, F.; Gallardo, B.; Geist, J.; Jarić, I.; Lambin, X.; Musseau, C.; et al. A proposed unified framework to describe the management of biological invasions. Biol. Invas. 2020, 22, 2633–2645. [Google Scholar] [CrossRef]
- Diagne, C.; Leroy, B.; Vaissière, A.C.; Gozlan, R.E.; Roiz, D.; Jarić, I.; Salles, J.M.; Bradshaw, C.J.A.; Courchamp, F. High and rising economic costs of biological invasions worldwide. Nature 2021, 592, 571–576. [Google Scholar] [CrossRef]
- Campoy, J.G.; Acosta, A.T.R.; Affre, L.; Barreiro, R.; Brundu, G.; Buisson, E.; González, L.; Lema, M.; Novoa, A.; Retuerto, R.; et al. Monographs of invasive plants in Europe: Carpobrotus. Bot. Lett. 2018, 165, 440–475. [Google Scholar] [CrossRef]
- Lazzaro, L.; Tondini, E.; Lombardi, L.; Giunti, M. The eradication of Carpobrotus spp. in the sand dune ecosystem at Sterpaia (Italy, Tuscany): Indications from a successful experience. Biologia 2020, 75, 199208. [Google Scholar] [CrossRef]
- Chenot, J.; Affre, L.; Gros, R.; Dubois, L.; Malecki, S.; Passetti, A.; Aboucaya, A.; Buisson, E. Eradication of invasive Carpobrotus sp.: Effects on soil and vegetation. Restor. Ecol. 2018, 26, 106–113. [Google Scholar] [CrossRef] [Green Version]
- Fos, M.; Sanz, B.; Sanchis, E. Carpobrotus management in a Mediterranean sand dune ecosystem: Minimum effective glyphosate dose and an evaluation of tarping. J. Ecol. Eng. 2021, 22, 57–66. [Google Scholar] [CrossRef]
- Fos, M.; Sanz, B.; Sanchis, E. The use of glyphosate for Carpobrotus eradication in sand dune ecosystems: Evaluation of the potential effects on the reintroduction of native plants. Plant Biosyst. 2021. [Google Scholar] [CrossRef]
- Wisura, W.; Glen, H.F. The South African species of Carpobrotus (Mesembryanthema–Aizoaceae). Contrib. Bolus Herb. 1993, 15, 76–107. [Google Scholar]
- Roiloa, S.R.; Rodríguez-Echeverría, S.; de la Peña, E.; Freitas, H. Physiological integration increases the survival and growth of the clonal invader Carpobrotus edulis. Biol. Invas. 2010, 12, 1815–1823. [Google Scholar] [CrossRef]
- Roiloa, S.R. Clonal traits and plant invasiveness: The case of Carpobrotus N.E.Br. (Aizoaceae). Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125479. [Google Scholar] [CrossRef]
- Van Groenendael, J.M.; de Kroon, H. Clonal Growth in Plants: Regulation and Function; SPB Academic Publishing: Hague, The Netherlands, 1990. [Google Scholar]
- Si, C.; Alpert, P.; Zhang, J.F.; Lin, J.; Wang, Y.Y.; Hong, M.M.; Roiloa, S.R.; Yu, F.H. Capacity for clonal integration in introduced versus native clones of the invasive plant Hydrocotyle vulgaris. Sci. Total Environ. 2020, 745, 141056. [Google Scholar] [CrossRef]
- Wang, J.; Xu, T.; Wang, Y.; Li, G.; Abdullah, I.; Zhong, Z.; Liu, J.; Zhu, W.; Wang, L.; Wang, D.; et al. A meta-analysis of effects of physiological integration in clonal plants under homogeneous vs. heterogeneous environments. Funct. Ecol. 2021, 35, 578–589. [Google Scholar] [CrossRef]
- Lu, H.Z.; Brooker, R.; Song, L.; Liu, W.-Y.; Sack, L.; Zhang, J.-L.; Yu, F.-H. When facilitation meets clonal integration in forest canopies. New Phytol. 2020, 225, 135–142. [Google Scholar] [CrossRef]
- Gao, F.-L.; Alpert, P.; Yu, F.H. Parasitism induces negative effects of physiological integration in a clonal plant. New Phytol. 2021, 229, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Hartnett, D.C.; Bazzaz, F.A. Physiological integration among intraclonal ramets in Solidago canadensis. Ecology 1983, 64, 779–788. [Google Scholar] [CrossRef]
- Slade, A.J.; Hutchings, M.J. An analysis of the costs and benefits of physiological integration between ramets in the clonal perennial herb Glechoma hederacea. Oecologia 1987, 73, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Alpert, P. Clonal integration in Fragaria chiloensis differs between populations: Ramets from grassland are selfish. Oecologia 1999, 120, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Saitoh, T.; Seiwa, K.; Nishiwaki, A. Importance of physiological integration of dwarf bamboo to persistence in forest understorey: A field experiment. J. Ecol. 2002, 90, 78–85. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Retuerto, R. Physiological integration ameliorates effects of serpentine soils in the clonal herb Fragaria vesca. Physiol. Plantarum 2006, 128, 662–676. [Google Scholar] [CrossRef]
- Song, Y.B.; Yu, F.H.; Keser, L.H.; Dawson, W.; Fischer, M.; Dong, M.; van Kleunen, M. United we stand, divided we fall: A meta-analysis of experiments on clonal integration and its relationship to invasiveness. Oecologia 2013, 171, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Müller-Schärer, H.; van Kleunen, M.; Cai, A.M.; Zhang, P.; Yan, R.; Dong, B.C.; Yu, F.H. Invasive alien plants benefit more from clonal integration in heterogeneous environments than natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Roiloa, S.R.; Yu, F.-H.; Barreiro, R. Plant invasions: Mechanisms, impacts and management. Flora 2020, 267, 151603. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Rodríguez-Echeverría, S.; López-Otero, A.; Retuerto, R.; Freitas, H. Adaptive plasticity to heterogeneous environments increases capacity for division of labor in the clonal invader Carpobrotus edulis (Aizoaceae). Am. J. Bot. 2014, 101, 1301–1308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roiloa, S.R.; Retuerto, R.; Campoy, J.G.; Novoa, A.; Barreiro, R. Division of labour brings greater benefits to clones of Carpobrotus edulis in the non-native range: Evidence for rapid adaptive evolution. Front. Plant Sci. 2016, 7, 349. [Google Scholar] [CrossRef] [Green Version]
- Roiloa, S.R.; Alpert, P.; Barreiro, L. Differences in physiological integration between invasive and noninvasive introduced clonal species of Carpobrotus. J. Plant Ecol. 2019, 12, 972–981. [Google Scholar] [CrossRef]
- Lechuga-Lago, Y.; Sixto-Ruiz, M.; Roiloa, S.R.; González, L. Clonal integration facilitates the colonization of drought environments by plant invaders. AoB Plants 2016, 8, plw023. [Google Scholar] [CrossRef] [Green Version]
- Campoy, J.G.; Retuerto, R.; Roiloa, S.R. Resource sharing strategies in ecotypes of the invasive clonal plant Carpobrotus edulis: Specialization for abundance or scarcity of resources. J. Plant Ecol. 2017, 10, 681–691. [Google Scholar]
- Rodríguez, J.; Calbi, M.; Roiloa, S.R.; González, L. Herbivory induced non-local responses of the clonal invader Carpobrotus edulis are not mediated by clonal integration. Sci. Total Environ. 2018, 633, 1041–1050. [Google Scholar] [CrossRef]
- Stuefer, J.F.; Huber, H. The role of stolon internodes for ramet survival after clone fragmentation in Potentilla anserina. Ecol. Lett. 1999, 2, 135–139. [Google Scholar] [CrossRef]
- Latzel, V.; Klimešová, J. Fitness of resprouters versus seeders in relation to nutrient availability in two Plantago species. Acta Oecol. 2009, 35, 541–547. [Google Scholar] [CrossRef]
- Bolhàr-Nordenkampf, H.R.; Long, S.P.; Baker, N.R.; Öquist, G.; Scheiber, U.; Lechner, E.G. Chlorophyll fluorescence as a probe of the photosynthetic competence of leaves in the field: A review of current instrumentation. Funct. Ecol. 1989, 3, 497–514. [Google Scholar] [CrossRef]
- Bolhàr-Nordenkampf, H.R.; Öquist, G. Chlorophyll fluorescence—A tool in photosynthesis research. In Photosynthesis and Production in a Changing Environment; Hall, D.O., Scurlock, J.M.O., Bolhàr-Nordenkampf, H.R., Leegood, R.C., Long, S.P., Eds.; Chapman and Hall: London, UK, 1993; pp. 193–206. [Google Scholar]
- Butler, W.; Kitajima, M. Fluorescence quenching in photosystem II of chloroplasts. Biochim. Biophys. Acta 1975, 376, 116–125. [Google Scholar] [CrossRef]
- Evans, J.P. Nitrogen translocation in a clonal dune perennial Hydrocotyle bonariensis. Oecologia 1988, 77, 64–68. [Google Scholar] [CrossRef]
- Salzman, A.G.; Parker, M.A. Neighbors ameliorate local salinity stress for a rhizomatous plant in a heterogeneous environment. Oecologia 1985, 65, 273–277. [Google Scholar] [CrossRef]
- D’Hertefeldt, T.; van der Putten, W.H. Physiological integration of the clonal plant Carex arenaria and its response to soil-borne pathogens. Oikos 1998, 81, 229–237. [Google Scholar] [CrossRef]
- Portela, R.; Dong, B.C.; Yu, F.H.; Barreiro, R.; Roiloa, S.R. Effects of physiological integration on defense strategies against herbivory by the clonal plant Alternanthera philoxeroides. J. Plant Ecol. 2019, 12, 662–672. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Retuerto, R. Clonal integration in Fragaria vesca growing in metal-polluted soils: Parents face penalties for establishing their offspring in unsuitable environments. Ecol. Res. 2012, 27, 95–106. [Google Scholar] [CrossRef]
- Pitelka, L.F.; Ashmun, J.W. Population Biology and Evolution of Clonal Organisms; Jackson, J.B.C., Buss, L.W., Cook, R.E., Eds.; Yale University Press: New Haven, CT, USA, 1985; pp. 339–435. [Google Scholar]
- Jónsdóttir, I.S.; Watson, M.A. Extensive physiological integration: An adaptive trait in resource-poor environments? In The Ecology and Evolution of Clonal Plants; de Kroon, H., van Groenendael, J., Eds.; Backhuys Publishers: Leiden, The Netherlands, 1997; pp. 109–136. [Google Scholar]
- Outridge, P.M.; Hutchinson, T.C. Effects of cadmium on ramet integration and resource allocation in the clonal fern Salvinia molesta. Oecologia 1990, 84, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Ong, C.K.; Marshall, C. The growth and survival of severely-shaded tillers in Lolium perenne L. Ann. Bot. 1979, 43, 147–155. [Google Scholar] [CrossRef]
- Jónsdóttir, I.S.; Callaghan, T.V. Localized defoliation stress and the movement of 14C-photoassimilates between tillers of Carex bigelowii. Oikos 1989, 54, 211–219. [Google Scholar] [CrossRef]
- Roiloa, S.R.; Rodríguez-Echeverría, S.; Freitas, H.; Retuerto, R. Developmentally-programmed division of labour in the clonal invader Carpobrotus edulis. Biol. Invasions 2013, 15, 1895–1905. [Google Scholar] [CrossRef]
- Savini, G.; Giorgi, V.; Scarano, E.; Neri, D. Strawberry plant relationship through the stolon. Physiol. Plantarum 2008, 134, 421–429. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Connection (C) | Glyphosate (G) | C × G | ||||
|---|---|---|---|---|---|---|
| Variable | F1,30 | p | F2,30 | p | F2,30 | p |
| (A) Basal ramets | ||||||
| Root biomass | 9.33 | 0.005 | 20.09 | <0.001 | 14.99 | <0.001 |
| Shoot biomass | 1.86 | 0.182 | 6.39 | 0.005 | 0.93 | 0.407 |
| Total biomass | 0.48 | 0.492 | 9.46 | <0.001 | 2.20 | 0.128 |
| Root mass ratio | 15.40 | <0.001 | 26.24 | <0.001 | 18.89 | <0.001 |
| (B) Apical ramets | ||||||
| Root biomass | 4.00 | 0.055 | 15.45 | <0.001 | 0.90 | 0.419 |
| Shoot biomass | 2.00 | 0.167 | 7.90 | 0.002 | 0.23 | 0.796 |
| Total biomass | 1.09 | 0.306 | 9.48 | <0.001 | 0.12 | 0.891 |
| Root mass ratio | 13.30 | <0.001 | 19.58 | <0.001 | 2.68 | 0.085 |
| (C) Whole clone | ||||||
| Root biomass | 0.83 | 0.369 | 27.17 | <0.001 | 5.54 | 0.009 |
| Shoot biomass | 0.012 | 0.898 | 6.99 | 0.003 | 0.61 | 0.552 |
| Total biomass | 0.07 | 0.800 | 10.19 | <0.001 | 0.85 | 0.436 |
| Root mass ratio | 0.18 | 0.677 | 43.85 | <0.001 | 7.19 | 0.003 |
| Effect | df | F | p |
|---|---|---|---|
| (A) Basal ramets | |||
| Between-subject | |||
| Connection (C) | 1, 30 | 1.73 | 0.198 |
| Glyphosate (G) | 2, 30 | 86.10 | <0.001 |
| C x G | 2, 30 | 3.99 | 0.029 |
| Within-subject | |||
| Time (T) | 2, 60 | 39.85 | <0.001 |
| C x T | 2, 60 | 1.31 | 0.278 |
| G x T | 4, 60 | 25.73 | <0.001 |
| C x G x T | 4, 60 | 1.19 | 0.323 |
| (B) Apical ramets | |||
| Between-subject | |||
| Connection (C) | 1, 30 | 5.00 | 0.033 |
| Glyphosate (G) | 2, 30 | 335.40 | <0.001 |
| C x G | 2, 30 | 11.50 | <0.001 |
| Within-subject | |||
| Time (T) | 2, 60 | 85.66 | <0.001 |
| C x T | 2, 60 | 0.54 | 0.583 |
| G x T | 4, 60 | 50.95 | <0.001 |
| C x G x T | 4, 60 | 3.88 | 0.008 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roiloa, S.R.; Yu, F.-H.; Barreiro, R. Effects of Glyphosate Application on Physiologically Integrated Clones of the Invasive Plant Carpobrotus edulis. Diversity 2022, 14, 47. https://doi.org/10.3390/d14010047
Roiloa SR, Yu F-H, Barreiro R. Effects of Glyphosate Application on Physiologically Integrated Clones of the Invasive Plant Carpobrotus edulis. Diversity. 2022; 14(1):47. https://doi.org/10.3390/d14010047
Chicago/Turabian StyleRoiloa, Sergio R., Fei-Hai Yu, and Rodolfo Barreiro. 2022. "Effects of Glyphosate Application on Physiologically Integrated Clones of the Invasive Plant Carpobrotus edulis" Diversity 14, no. 1: 47. https://doi.org/10.3390/d14010047
APA StyleRoiloa, S. R., Yu, F.-H., & Barreiro, R. (2022). Effects of Glyphosate Application on Physiologically Integrated Clones of the Invasive Plant Carpobrotus edulis. Diversity, 14(1), 47. https://doi.org/10.3390/d14010047