Abstract
Coastal dunes of the southern Buenos Aires province, in Argentina, are one of the last remnants of biodiversity in the southern Pampa unit, within the Rio de la Plata Grasslands. While the direct loss of grasslands due to the advance of cities and afforestation is conspicuous, the negative effects of the subdivision of the remaining environments on biodiversity and ecosystem dynamics are less considered. Our work aimed to analyze the combined effect of fires and subdivision of grasslands by invasive alien trees. Our results suggest that fragmentation affects the resilience of coastal grasslands after fire and affects the course of succession, promoting the establishment of invasive alien plants. We also suggest that fires favor the advance of pines and acacias towards the interior of grassland remnants, further reducing their area. The effective conservation of the components of Pampas biodiversity that still persist in these coastal ecosystems will depend on preventing, mitigating and compensating the insularization effects associated with forest plantations and the expansion of invasive trees, by means of adequate territorial planning that allows remnants to be maintained in a good state of conservation.
1. Introduction
Natural grasslands of temperate and subtropical regions of the world have been almost completely transformed into agriculture, afforestation and pastures, due to the suitability of their soils and climate [1,2,3,4]. This has caused them to be recognized as a terrestrial biome, where biodiversity and ecosystem services are most at risk on a global scale [5,6]. This is the case for the “Rio de la Plata Grasslands”, originally comprising natural prairies of central-eastern Argentina, Uruguay and southern Brazil [7], that have been almost completely replaced by crops, implanted pastures and afforestation during the last two centuries. As a consequence, Argentinian Pampas are currently represented by scarce and small remnants of natural and semi-natural grasslands, immersed in a matrix of different land uses [8,9]. The few grassland relicts in a good state of conservation are restricted to habitats that are not suitable for agriculture, such as rocky outcrops, flooding areas and coastal sand dunes [10]. Coastal dunes of the southern Pampas treasure a high biological and environmental richness and provide key ecological system services, including freshwater storage and coastal protection [11,12]. Unfortunately, these areas are losing the battle against the establishment and expansion of urbanization, afforestation and the spontaneous spread of invasive alien trees [13,14]. The introduction of forest plantations in the coastal region of the Pampas began in the early nineteenth century to increase its touristic appeal [15], representing the introduction of a completely new life form, in environments where native trees are rare or absent. Some of the most commonly introduced trees include Acacia and Pinus species, which have expressed aggressive, invasive behavior in coastal dunes [10]. Trees stand out among invasive species for the extent and intensity of the changes they produce on natural grasslands, replacing native biota and promoting monospecific communities [16,17]. In this context, their dispersion represents one of the main threats to coastal dune conservation [18,19]. Both dune afforestation and further spontaneous expansion of the trees have direct negative impacts, including alteration of sand dynamics; increases in the frequency and intensity of fires, in most cases of anthropic origin; and replacement and fragmentation of native plant communities [19,20].
The fragmentation of natural habitats is one of the main processes responsible for the loss of global biodiversity [21,22]. Different authors define fragmentation as the landscape dynamics by which habitat loss results in the division of a continuous natural environment into two or more smaller fragments, isolated from each other by a matrix of dissimilar habitats. This highlights the importance of the size and spatial arrangement of the fragments as determinants of the type and intensity of the changes they undergo [23,24]. This process also implies the generation of new edges through which the matrix has a series of effects, generally negative, on the remnants of original vegetation [25]. While the direct loss of grasslands due to the advance of afforestation is conspicuous and evident, the effects of the subdivision of the remaining environments are less considered but could surpass the former in terms of their ability to affect the composition and dynamics of natural communities [19,21,26]. Fragmentation commonly results in sensible reductions in species richness [22,27]. Nevertheless, changes in species composition is a more precise indicator of the alterations associated with the fragmentation of grasslands, and it allows the proposal of possible mechanisms underlying the observed changes in plant communities [28]. In fragmented environments, the proximate causes of changes in the composition of plant communities lie in the differential responses of species to the changes in biotic and abiotic processes that result from the transition between the original landscape and the matrix [29].
On the other hand, fragmentation increases the vulnerability of fragmented vegetation to extrinsic disturbances, such as fire [30]. In this context, habitat fragmentation can alter successional trajectories, and the intensity of these changes will depend on the intensity of the disturbance and the size of the area that is being disturbed [22]. However, the interaction between fragmentation and an extrinsic disturbance, such as fire, have not been analyzed so far [21]. Fire is an important component of the ecological succession of plant communities in grassland ecosystems. Native species have evolved in the presence of this natural disturbance and therefore acquired adaptations that allow them to survive [31,32]. However, its effects on the species composition of vegetation and ecosystem functions vary with its intensity, frequency, extension and duration [33,34]. Afforestation with woody species in grassland environments and their eventual spontaneous spread generates particularly significant changes in fire dynamics [31,32,35,36]. This is because the temperature and the severity of the fire increases with the amount of easily combusted material accumulated, whereas grasslands usually reach lower burn temperatures that do not completely eliminate the vegetation cover and do not affect the organic carbon reservoirs in the soil. However, afforestation not only offers a quantity of fuel that allows it to reach higher temperature levels, it is also accompanied by an absence or a significant reduction in an understory capable of resisting or rapidly recovering after fire, so the possibilities of soil erosion and damage to the ecosystem increase [2]. The coexistence of fire and afforestation implies a risk because fire may facilitate the spontaneous spread of the trees outside the planted areas, which is particularly evident in the case of pines [2,37,38,39]. A strong correlation between the loss and fragmentation of native habitats and the increase in the abundance of invasive species has been reported for altered environments [24,40]. The transformation of the landscape by human activities favors the spread of invasive species through changes in the conditions associated with borders, increasing the arrival of propagules and opening invasion windows [41,42].
The coastal dunes of the province of Buenos Aires are well preserved in comparison to the rest of the grasslands of the southern Pampas and are dominated by native species [10,12]. Nevertheless, they are threatened by urban development, off-road vehicles, afforestation and the spontaneous expansion of exotic trees [10]. During the last ten years, we have studied the effects of the subdivision of coastal grasslands due to afforestation with exotic pines, with the objective of evaluating the effects on different biological levels and their interaction with fire [28,43,44,45]. In this communication, we integrate the analysis of the effects of fire and fragmentation by invasive alien trees, evaluating the post-fire recovery processes in terms of plant composition and the ability of exotic woody plants to advance on remnants of grasslands after the fire. We expect that: 1. fragmentation alters the composition of the plant communities, favoring exotic species, 2. there is a minimum fragment size, below which the effects of fragmentation become particularly intense, 3. fragmented grasslands are less resilient to fire than equivalent areas of continuous grassland and 4. fire in fragments further promotes the advance of alien plants.
2. Materials and Methods
2.1. Study Area
The coastal dunes of the southern Pampas [46] occupy the southwest of the Buenos Aires province, Argentina. This coastal strip ranges between two and seven kilometers in width, including beaches; bare dunes; dunes covered with vegetation, mostly psammophilous grasslands; and depressions between the dunes with ponds associated with humid grasslands [12]. The climate is temperate, influenced by the proximity to the sea [47]. Plant communities are dominated by Hyalis argentea and Panicum urvilleanum and include local endemics such as Neosparton darwinii and Senecio bergii. Interdunal depressions are covered by Cortaderia selloana, Juncus acutus, Schoenoplectus americanus and other hygrophilous herbs [11,48].
This study was carried out in a set of remnants of coastal grasslands of different areas, surrounded by a plantation of Pinus pinaster (38°54′60″ S; 60°32′60″ W) covering 30 ha, and in adjacent continuous grassland. The area also included scattered plantations of Acacia longifolia; both are species with antecedents of invasions in these ecosystems [10,20] and are also associated with changes in the dynamics of fire [38,49]. Our studies started in 2011 and extended for seven years. On January 6th, 2014, the area was completely burnt by a wildfire that also affected neighboring continuous grasslands.
2.2. Study Design and Field Sampling
Thirty-nine remnants of natural vegetation, surrounded by the pine plantation, were identified from a satellite image of the site, taken from Google Earth and dated December 14th, 2006. The area of each fragment was measured using Google Earth Pro, and fifteen sample units were selected: six small size fragments (<0.1 ha), three medium size (0.1–0.5 ha), and six large size (0.5–2.5 ha). Fifteen controls of equivalent areas were randomly selected in a neighboring continuous grassland, free of trees [50] (Figure 1). In 2011, the fragments were identified in the field, confirming that there had been no changes in their area and shape since the capture of the satellite images. Plant composition at the remnants and controls was assessed in April 2011, before the fire (t0), and three (t1) and fifteen (t2) months after it (April 2014 and April 2015, respectively). In each case, we estimated the percentage cover of all plant species present, following the method of Braun Blanquet [51] in each sample unit (each whole fragment and each whole control in the continuous grassland). The composition of plant species was assessed in the whole fragments, and we assumed that the size ranges used to group the fragments resulted in homogeneous effects of the area within each category. Plants were identified to the species and/or genus level [52] and were classified as native or exotic. The Catalog of Vascular Plants of the Southern Cone [53] was used for nomenclature.
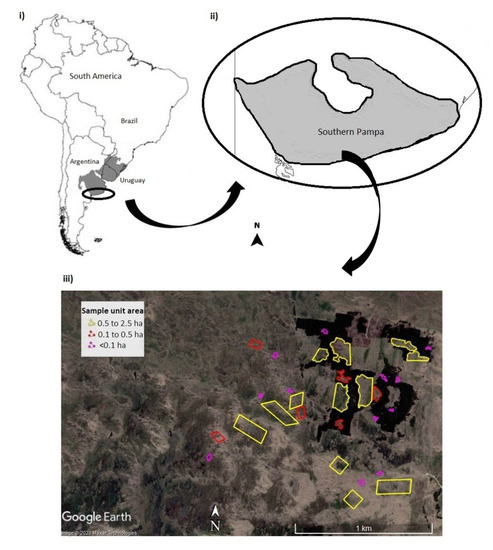
Figure 1.
Location of the study area in the coastal grasslands of the southern Pampa, Buenos Aires, Argentina (i,ii). The arrangement of the samples of different sizes (less than 0.1 ha, 0.1 to 0.5 ha and 0.5 to 2.5 ha) is indicated, including fragments surrounded by afforestation and controls in the continuous grassland (iii). In gray, delimitation of the Grasslands of the Río de la Plata (Adapted from Soriano et al. (1992)). Image from Google Earth (2021).
In 2017, three and a half years after the fire (t3), we measured the area of the fifteen fragments in the field using Google Earth Pro and estimated the cover of pines and acacias in each of them to evaluate the ability of exotic woody plants to advance over the remnants after the fire [45].
2.3. Data Analysis
Species composition of the sampling sites was compared by means of a Principal Component Analysis (PCA), based on the covariance matrix of the species cover data, with the objective of identifying sets of species associated with the size and context of the samples and with the different times with respect to fire. Data were previously transformed to the arc sine of the square root to correct the distortion of the Euclidean distance in its spatial representation [54]. To maintain an appropriate balance between the number of samples and variables (coverage by species), the plants present in less than 33% of the samples (75 species) were excluded from the analysis. Multivariate analysis of variance (MANOVA) was conducted on the transformed data to compare species composition between the different treatments (context, size of the areas and time with respect to fire). Relevant comparisons were performed through the Wilks’ Lambda test.
We compared changes in landscape configuration in the fifteen grassland fragments before (t0) and after the fire (t3) using Google Earth Pro tools and trend analysis of three landscape variables: total area of grasslands, average area of grassland fragments and average distance between them [55]. The Wilcoxon test was used to assess whether the difference between these landscape variables, before and after the fire (t0 and t3, respectively), were significant. In addition, we compared the relative frequency of fragments of different sizes. A two-way factorial ANOVA and Bonferroni method [56] were applied to analyze the effects of fire (before and after) and the size of the fragments; and their interactions on the percentage cover of P. pinaster and A. longifolia. The data were transformed to the arcsine of the square root to meet the assumptions of normality and homoscedasticity. The normality of the data was evaluated by the study of residuals and by normal probability plots. The Levene test was used to verify homogeneity of variances. All analyses and graphs were performed using the statistical packages Infostat [57] and XLSTAT (versions 5.7.2 and 2018.2.50198).
3. Results
The first component of the PCA (20.04% of the total variance) grouped the samples according to successional stages before the fire (t0) and separated the smaller fragments surrounded by the forest plantation (grouped in the third quadrant) with respect to the smaller continuous grassland controls (distributed in the first and fourth quadrants, Figure 2a). While the fragments were characterized by a greater abundance of the native species Cyperus reflexus and the exotic Senecio madagascariensis and Hypochaeris radicata, the controls were associated with high abundances of grasses and other native species typical of the Pampean grassland, such as Aristida spegazzinii, Panicum urvilleanum, Schizachyrium plumigerum, Margyricarpus pinnatus and Discaria americana (Figure 2a, Table S1). The large pre-fire samples (t0), both fragments and plots in continuous grassland, formed a group in the second quadrant, characterized by greater abundances of three perennial grasses: Cortaderia selloana, Eragrostis airoides and Imperata brasiliensis, plus Baccharis glutinosa, a perennial herb (Figure 2a, Table S1). The second component (16.56% of the total variance) separated smaller and medium fragments from large fragments and controls, three months after the fire (t1), grouping the corresponding samples in the second and in the first quadrants, respectively (Figure 2b). Samples in the second quadrant were associated with high coverage values of the exotic species Eragrostis curvula and the native Hydrocotyle bonariensis, while the first quadrant was characterized by greater abundances of the native species Eragrostis airoides, Juncus acutus, Ambrosia tenuifolia and Tessaria absinthioides; these last two species are typically associated with disturbances (Figure 2b, Table S1). The second component also separated fragments fifteen months after the fire (t2; on the third quadrant) from controls at the same sampling time (on the fourth quadrant), throughout the range of sizes analyzed (Figure 2b). Fragments were associated with higher cover of the exotic species Acacia longifolia, Pinus pinaster and Hypochaeris radicata, and the native Schoenoplectus americanus and Conyza bonariensis, while the controls were characterized by greater abundance of the natives Imperata brasiliensis, Cortaderia selloana, Panicum urvilleanum, Achyrocline satureioides, Solidago chilensis, Baccharis glutinosa and Schizachyrium plumigerum (Figure 2b, Table S1).
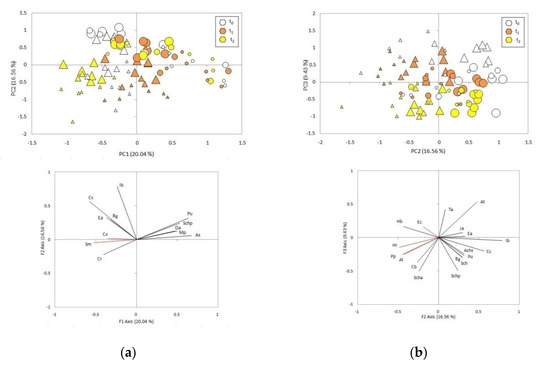
Figure 2.
Principal Component Analysis (PCA) of sampling units of different sizes and at different times in relation to fire, located in continuous grassland (circles) and surrounded by a pine plantation (triangles), and vectors of plant species with the abbreviations of the names (see references in Table S1; exotic species in red). (a) PC1 and PC2, (b) PC2 and PC3. Symbols of different sizes represent sample units of different areas (small = up to 0.1 ha; medium = 0.1 to 0.5 ha; large = 0.5 to 2.5 ha) and colors represent different times: before the fire (t0, white), and three (t1, orange) and fifteen months (t2, yellow) after the fire.
The Principal Components Analysis resulted in a grouping of the samples according to the context (fragmented grassland and continuous grassland) and the size of the areas, in the different times with respect to the fire. This was further confirmed by the multivariate analysis of variance (MANOVA) test. The results of the MANOVA indicate that there are significant differences in the mean percentage cover of the species according to the context, the size of the areas and the time with respect to the fire (Table 1). Table 1 indicates that there is significant interaction effect of context (fragmented grassland and continuous grassland) with the size of the areas and with the time with respect to the fire (p = 0.0006 and p< 0.0001, respectively).

Table 1.
MANOVA tests.
Landscape variables remained stable during the years before the fire (t0; Figure 3). Three and a half years after (t3), the total area of grassland immersed in the afforestation decreased from 11 to 8 ha. The average size of grassland fragments decreased (t0: 0.70 ± 0.82 ha; t3: 0.54 ± 0.71 ha) and the average distance between them increased (t0: 346 ± 210.55 m; t3: 369 ± 214.2 m). Both changes were statistically significant (T = 120, p = 0.001 for size; T = 253, p< 0.0001 for distance). The total number of fragments decreased after the fire (from 15 to 13). A decrease was also found in the number of the larger and smaller fragments and an increase in medium size fragments (Table 2). A significant interaction between the area and the time in relation to the fire was observed on the cover of P. pinaster and A. longifolia (F = 4.81, p = 0.007). The percentage cover of Pinus and Acacia was greater after than before the fire at the small fragments (up to 0.1 ha) but showed no differences at the medium and larger sizes (Figure 4).
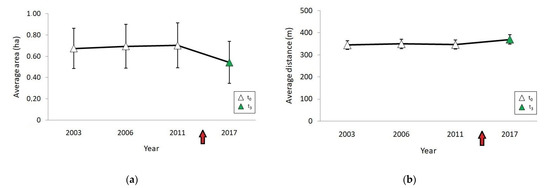
Figure 3.
Changes in (a) average area of grassland fragments, and (b) average distance between grassland fragments before (t0, white) and after (t3, green) the fire. Red arrow on the x-axis indicates the occurrence of the fire.
Table 2.
Comparison of the relative frequency of grassland fragments of different area at different times in relation to fire (t0 and t3; before and after, respectively). Adapted from Yezzi, Nebbia and Zalba (2020).
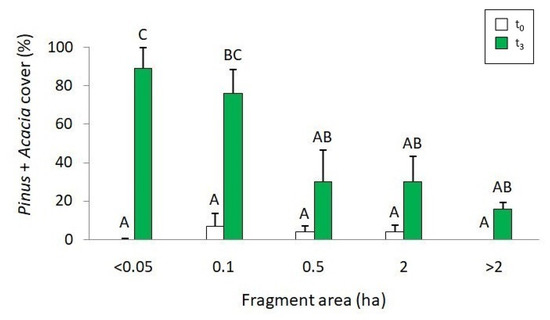
Figure 4.
Comparison between the percentage cover of P. pinaster and A. longifolia in grassland fragments of different size at different times in relation to fire (t0 = white and t3 = green, before and after fire, respectively). Different letters indicate significant differences (p < 0.05); bars represent the standard error.
4. Discussion
Fragmentation currently operates simultaneously with other disturbance factors, many of which are new to the ecosystem or have been altered by human activity in their frequency, intensity or extent [58,59]. The synergy between these factors may be more serious than their separate effects [23,60]. The invasion by exotic plants has a similar relationship with the alteration of the disturbance regime and with the fragmentation of habitats [61,62]. In particular, changes in the nature, intensity and frequency of disturbances commonly enhances ecosystem vulnerability to invasion [63,64]. Our work contributes to the understanding of the joint effects of fire and fragmentation on coastal grasslands surrounded by forest plantations and on the influence of fire on the expansion of invasive pines and acacias. Evidence is provided about how the recovery of coastal grasslands after a fire varies depending on whether they are part of a continuous area or are immersed in a forest matrix, about the reduction in the resilience of the fragmented grassland against fires and on the increase in its susceptibility to invasion by exotic species. In addition, circumstantial evidence is provided about the intensification of grassland fragmentation as a consequence of the acceleration of the advance of invasive woody species after a fire.
Before the occurrence of the fire, we found differences in the composition between smaller fragments surrounded by the forest plantation and smaller continuous grassland controls, a difference that was not evident between larger fragments and controls. Large fragments (0.5 to 2.5 ha) seemed to be more resistant to fragmentation, maintaining a composition of species similar to that of the continuous grassland, which can be considered as an initial indication of a minimum hypothetical size for the conservation of coastal grassland fragments. However, fragments smaller than 0.5 ha must not be discarded and should also be considered for conservation, not only because of their intrinsic value for certain species, but also for their possible role as stepping stones that allow the protection of species of pollinators, seed dispersers, and herbivores. In particular, we observed that the exotic species Senecio madagascariensis and Cirsium vulgare were dominant in small fragments but were practically absent or less represented in the controls of continuous grassland of equivalent areas. The tree matrix surrounding grassland fragments could modify environmental parameters such as temperature and humidity, as well as influence the dispersal of propagules [65,66], favoring the establishment of exotic species, which typically behave opportunistically [67,68,69]. However, these differences became widespread throughout the whole size range (small, medium and large areas) one year after the fire. In this way, the effects of fragmentation can go unnoticed at certain successional stages and become evident in the successional stages that follow a disturbance. This finding supports the decades-old suggestion that habitat fragmentation may alter the capacity of biodiversity to recover from additional disturbances [70] and also supports more recent findings showing how ecosystem fragmentation can modify successional trajectories that occur after a disturbance [22,71].
We found changes in species composition along the successional trajectory of grasslands affected by fire that provide evidence on the effects of fragmentation on the resilience of plant communities. Continuous grassland showed a marked constancy in the identity of dominant species, such as Cortaderia selloana and Imperata brasiliensis, before the fire and fifteen months later, evidencing a capacity of recovery that was much less obvious in the fragments surrounded by the forest plantation. The increase in the percentage cover of exotic plants, such as Hypochaeris radicata, in fragmented habitats fifteen months after the fire contrasts with its constancy throughout the succession for the plots surrounded by continuous grassland. This result indicates that fire could increase the vulnerability of fragmented coastal grasslands to invasions, while intact grasslands appear to be more resistant, as has been reported for similar fire-affected habitats in South Africa [36]. At this point, it seems important to note that the forest matrix could have affected the recovery capacity of grassland fragments not only by the insularization effects themselves but also due to the increase in temperature that a forest fire can reach compared to typical grassland fire conditions [72].
The effects of fire as a promoter of the expansion of invasive species were especially evident in the case of Pinus pinaster and Acacia longifolia, that expanded their distribution results from the forest matrix into the remnant grassland fragments after the disturbance. Pines and acacias have evolved in fire-adapted ecosystems and different authors have described the existence of positive feedback between fire and the invasion of these species [31,38,49]. Pines typically release large quantities of seeds after a fire, and the post-fire environmental conditions also favor their germination and establishment [73,74], while Acacia longifolia also benefits from fire events in terms of a massive germination of seeds from the seed bank in the soil [49]. This resulted, in our case, in the intensification of the fragmentation through the complete disappearance of some of the smallest remnants and the decrease in the area of the largest ones. The expansion of exotic woody species over areas originally covered by grassland limits native vegetation to increasingly smaller and more isolated areas, with the consequent increase in the edge effect, the loss of individuals and the limitation of biological interactions [60]. Importantly, these synergies could be magnified over time [21], since invasions and fires generate positive feedback that accelerates the process of homogenization of biodiversity in degraded grasslands [75]. One of the current challenges for biodiversity conservation is how to deal with the synergy of factors that promote ecosystem homogenization, that becomes even more important in fragmented landscapes [24,76]. In this context, it is essential to understand the interactions between invasive species and invaded ecosystems in order to detect and reduce their impacts on biodiversity [19,77,78].
The results of this work allow the identification of key information gaps for the effective conservation of coastal grassland relicts that indicate future research directions. Among them is the need to define minimum area thresholds so that the grassland fragments not only preserve their ecological integrity but also allow the recovery of the composition of their vegetation and the persistence of native species in the face of recurrent disturbances such as fires. Another priority will be to identify forest species with less spontaneous dispersal capacity over the dune ecosystem to promote the replacement of pines and acacias in the design of the coastal landscape.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/d13120637/s1, Table S1: correlation coefficients between the abundance of plant species and the first three components of the PCA (PC1, PC2 and P3).
Author Contributions
All authors contributed to the study conception and design. Data collection and experiments were performed by A.Y. All authors analyzed the results and contributed to the writing of the paper. All authors have read and agreed to the published version of the manuscript.
Funding
The work included in this paper received financial support from Universidad Nacional del Sur and CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas), Argentina.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data can be obtained from the corresponding author upon request. Data is not publicly available due to its usage in the ongoing study.
Acknowledgments
We thank to CONICET (Consejo Nacional de Investigaciones Científicas y Técnicas) and the Department of Biology, Biochemistry and Pharmacy of the Universidad Nacional del Sur.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Baldi, G.; Guerschman, J.P.; Paruelo, J.M. Characterizing fragmentation in temperate South America grasslands. Agric. Ecosyst. Environ. 2006, 116, 197–208. [Google Scholar] [CrossRef]
- Jobbágy, E.G.; Vasallo, M.; Farley, K.A.; Piñeiro, G.; Garbulsky, M.F.; Nosetto, M.D.; Jackson, R.B.; Paruelo, J.M. Forestación en pastizales: Hacia una visión integral de sus oportunidades y costos ecológicos. Agrociencias 2006, 10, 109–124. [Google Scholar]
- Modernel, P.; Rossing, W.H.A.; Dogliotti, S.; Picasso, V.; Tittonell, P. Land use change and ecosystem service provision in Pampas and Campos grasslands of southern South America. Environ. Res. Lett. 2016, 11, 113002. [Google Scholar] [CrossRef]
- Baeza, S.; Paruelo, J.M. Land use/land cover change (2000–2014) in the Rio de la Plata grasslands: An analysis based on MODIS NDVI time series. Remote Sens. 2020, 12, 381. [Google Scholar] [CrossRef] [Green Version]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- Suttie, J.M.; Reynolds, S.G.; Batello, C. Grasslands of the World; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2005. [Google Scholar]
- Soriano, A. Rio de la Plata Grasslands. In Natural Grasslands; Coupland, R.T., Ed.; Elsevier: Amsterdam, The Netherlands, 1991; pp. 367–407. [Google Scholar]
- Burkart, R. Las Áreas Protegidas de la Argentina. In La Situación Ambiental Argentina 2005; Brown, A., Martínez Ortíz, U., Acerbiand, M., Corcuera, J., Eds.; Fundación Vida Silvestre: Buenos Aires, Argentina, 2006; pp. 399–403. [Google Scholar]
- Pretelli, M.G.; Isacch, J.P.; Cardoni, D.A. Species-area relationships of specialist versus opportunistic pampas grassland birds depend on the surrounding landscape matrix. Ardeola 2018, 65, 3–23. [Google Scholar] [CrossRef]
- Zalba, S.M.; Villamil, C.B. Woody plant invasion in relictual grasslands. Biol. Invasions 2002, 4, 55–72. [Google Scholar] [CrossRef]
- Kittlein, M.J.; Mora, M.S.; Vassallo, A.I.; de Durana, F.; Ricciardulli, M.G.; Tizón, F.R. Dunas del Sureste Bonaerense. In Identificación de Áreas Valiosas de Pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil; Bilenca, D., Miñarro, F., Eds.; Fundación Vida Silvestre: Buenos Aires, Argentina, 2004; pp. 76–77. [Google Scholar]
- Celsi, C.E.; Mac-Lean, D.; Yezzi, A.; Triches, M. Dunas Costeras de la Pampa Austral. Biodiversidad, Ecología y Conservación Entre el río Quequén Salado y el Balneario Pehuen-Có; Fundación de Historia Natural Félix de Azara: Buenos Aires, Argentina, 2010. [Google Scholar]
- Bilenca, D.; Miñarro, F. Identificación de Áreas Valiosas de Pastizal (AVPs) en las Pampas y Campos de Argentina, Uruguay y sur de Brasil; Fundación Vida Silvestre: Buenos Aires, Argentina, 2004. [Google Scholar]
- Guadagnin, D.L.; Zalba, S.M.; Costa Gorriz, B.; Fonseca, C.R.; Nebbia, A.J.; Cuevas, Y.A.; Emer, C.; Germain, P.; Wendland, E.; Perello, L.F.; et al. Árvores e Arbustos Exóticos Invasores no Bioma Pampa—Questões Ecológicas, Culturais e Socioeconômicas de Umdesafiocrescente. In Campos Sulinos: Conservação e uso Sustentable da Biodiversidade; de Patta Pillar, V., Müller, S.C., de Souza, Z.M., Ávila Jacques, A.V., Eds.; Ministerio do Meio Ambiente: Brasilia, Brasil, 2009; pp. 300–316. [Google Scholar]
- Dadon, J.R. El Impacto del Turismo Sobre los Recursos Naturales Costeros en la Costa Pampeana. In Zona Costera de la Pampa Argentina; Dadon, J.R., Matteucci, S.D., Eds.; Lugar Editorial: Buenos Aires, Argentina, 2002; pp. 101–121. [Google Scholar]
- Cuevas, Y.A.; Zalba, S.M. Control de pinos invasores en el parque provincial Ernesto Tornquist (Buenos Aires): Áreas prioritarias y análisis de costos. BioScriba 2009, 2, 76–89. [Google Scholar]
- Fonseca, C.; Guadagnin, D.L.; Emer, C.; Masciadri, S.; Germain, P.; Zalba, S.M. Invasive alien plants in the Pampas grasslands: A tri-national cooperation challenge. Biol. Invasions 2013, 15, 1751–1763. [Google Scholar] [CrossRef]
- Marchante, H.; Marchante, E.; Freitas, F. Invasion of the Portuguese Dune Ecosystem by the Exotic Species Acacia longifolia (Andrews) Wild: Effects at Community Level. In Plant Invasions: Ecological Threats and Management Solutions; Child, L.E., Brock, J.H., Brundu, G., Prach, K., Pysek, P., Wade, P.M., Williamson, M., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2003; pp. 75–85. [Google Scholar]
- Pauchard, A.; García, A.; Zalba, S.; Sarasola, M.; Zenni, R.; Ziller, S.; Nuñez, M.A. Pine Invasions in South America: Reducing Their Ecological Impacts Through Active Management. In Biological Invasions in Changing Ecosystems: Vectors, Ecological Impacts, Management and Predictions; Canning Clode, J., Ed.; De Gruyter Open Ltd.: Warsaw, Poland, 2016; pp. 318–342. [Google Scholar]
- Alberio, C.; Comparatore, V. Patterns of woody plant invasion in an Argentinean coastal grassland. Acta Oecologica 2014, 54, 65–71. [Google Scholar] [CrossRef]
- Haddad, N.M.; Brudvig, L.A.; Clobert, J.; Davies, K.F.; Gonzalez, A.; Holt, R.D.; Lovejoy, T.E.; Sexton, J.O.; Austin, M.P.; Collins, C.D.; et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci. Adv. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilson, M.C.; Chen, X.; Corlett, R.T.; Didham, R.K.; Ding, P.; Holt, R.D.; Holyoak, M.; Hu, G.; Hughes, A.C.; Jiang, L.; et al. Habitat fragmentation and biodiversity conservation: Key findings and future challenges. Landsc. Ecol. 2016, 31, 219–227. [Google Scholar] [CrossRef] [Green Version]
- Ewers, R.M.; Didham, R.K. Confounding factors in the detection of species responses to habitat fragmentation. Biol. Rev. 2006, 81, 117–142. [Google Scholar] [CrossRef] [PubMed]
- Didham, R.K. Ecological Consequences of Habitat Fragmentation. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons: Hoboken, NJ, USA, 2010. [Google Scholar] [CrossRef]
- Saunders, D.A.; Hobbs, R.J.; Margules, C.R. Biological consequences of ecosystem fragmentation: A review. Conserv. Biol. 1991, 5, 18–32. [Google Scholar] [CrossRef]
- Archer, S.; Boutton, T.W.; Hibbard, K.A. Trees in Grasslands: Biogeochemical Consequences of Woody Plant Expansion. In Global Biogeochemical Cycles and Their Interrelationship with Climate; Schulze, E.A., Harrison, S.P., Heimann, M., Holland, E.A., Lloyd, J., Prentice, I.C., Schimel, D.S., Eds.; Academic Press: London, UK, 2001; pp. 115–130. [Google Scholar]
- Primack, R.; Rozzi, R.; Feinsinger, P.; Dirzo, R.; Massardo, F. Fundamentos de Conservación Biológica: Perspectivas Latinoamericanas; Fondo de Cultura Económico: Mexico City, Mexico, 2001. [Google Scholar]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Fragmentación de pastizales psamófilos por plantaciones de pinos: Efectos sobre la riqueza y la composición vegetal. Ecol. Austral. 2018, 28, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, P.R.; Rodríguez, M.E. Determinantes de la composición florística y efecto de borde en un fragmento de bosque en el Guaviare, Amazonia colombiana. Colomb. For. 2008, 11, 5–18. [Google Scholar] [CrossRef]
- Lord, J.M.; Norton, D.A. Scale and the spatial concept of fragmentation. Conserv. Biol. 1990, 4, 197–202. [Google Scholar] [CrossRef]
- Grace, J.B.; Smith, M.D.; Grace, S.L.; Stohlgren, T.J. Interactions between Fire and Invasive Plants in Temperate Grasslands of North America. In Proceedings of the Invasive Species Workshop: The Role of Fire in the Control and Spread of Invasive Species; Galley, K.E.M., Wilson, T., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2001; pp. 40–65. [Google Scholar]
- Vega, E.; Peters, E. Conceptos Generales Sobre el Disturbio y sus Efectos en los Ecosistemas. In Conservación de Ecosistemas Templados de Montaña en México; Sánchez, O., Vega, E., Peters, E., Monroy-Vilchis, O., Eds.; Secretaría de Medio Ambiente y Recursos Naturales; Instituto Nacional de Ecología; Secretaría de Medio Ambiente; Recursos Naturales y Pesca; Comisión Nacional para el conocimiento y Uso de la Biodiversidad: Mexico City, Mexico, 2003; pp. 137–150. [Google Scholar]
- Wu, G.L.; Zhao, L.P.; Wang, D.; Shi, Z.H. Effects of time-since-fire on vegetation composition and structures in semi-arid perennial grassland on the Loess Plateau, China. Clean Soil Air Water 2014, 42, 98–103. [Google Scholar] [CrossRef]
- Van Langevelde, F.; de Groot, C.; Groen, T.A.; Heitkonig, I.M.A.; Gaigher, I. Effect of patches of woody vegetation on the role of fire in tropical grasslands and savannas. Int. J. Wildland Fire 2014, 23, 410–416. [Google Scholar] [CrossRef]
- Deák, B.; Valkó, O.; Török, P.; Végvári, Z.; Hartel, T. Grassland fires in Hungary Experiences of nature conservationists on the effects of fire on biodiversity. Appl. Ecol. Environ. Res. 2014, 12, 267–283. [Google Scholar] [CrossRef]
- Bachinger, L.M.; Brown, L.R.; van Rooyen, M.W. The effects of fire-breaks on plant diversity and species composition in the grasslands of the Loskop Dam Nature Reserve, South Africa. Afr. J. Range Forage Sci. 2016, 33, 21–32. [Google Scholar] [CrossRef]
- Richardson, D.M. Forestry trees as invasive aliens. Conserv. Biol. 1998, 12, 18–26. [Google Scholar] [CrossRef]
- Zalba, S.M.; Cuevas, Y.A.; Boó, R. Aleppo pine invasion after a wildfire in an Argentinean grassland nature reserve. J. Environ. Manag. 2008, 88, 539–546. [Google Scholar] [CrossRef]
- Cuevas, Y.A.; Zalba, S.M. Recovery of native grasslands after removing invasive pines. Restor. Ecol. 2010, 18, 711–719. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Gemmell, N.J.; Rand, T.A.; Ewers, R.M. Interactive effects of habitat modification and species invasion on native species decline. Trends Ecol. Evol. 2007, 22, 489–496. [Google Scholar] [CrossRef] [PubMed]
- With, K.A. Assessing the risk of invasive spread in fragmented landscapes. Risk Anal. 2004, 24, 803–815. [Google Scholar] [CrossRef] [PubMed]
- Yates, E.D.; Levia, D.F.; Williams, C.L. Recruitment of three non-native invasive plants into a fragmented forest in southern Illinois. For. Ecol. Manag. 2004, 190, 119–130. [Google Scholar] [CrossRef]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Fragmentation and grassland plants: Individual and transgenerational effects. Plant Ecol. 2020, 12, 1275–1291. [Google Scholar] [CrossRef]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Interaction between fire and fragmentation in the successional stages of coastal dune grasslands of the southern Pampas, Argentina. Sci. Rep. 2019, 9, 15109. [Google Scholar] [CrossRef] [Green Version]
- Yezzi, A.; Nebbia, A.J.; Zalba, S.M. Synergistic Interactions of Grassland Fragmentation, Fire and Invasive Woody Plants in the Argentine Pampas. In Advances in Environmental Research Volume 77; Daniels, J.A., Ed.; Nova Science Publishers Inc.: New York, NY, USA, 2020; pp. 43–70. [Google Scholar]
- Cabrera, A.L. Fitogeografía de la República Argentina. Boletín De La Soc. Argent. De Botánica 1971, 14, 1–42. [Google Scholar]
- Campo de Ferreras, A.; Capelli de Steffens, A.; Diez, P. El Clima del Suroeste Bonaerense; Departamento de Geografía y Turismo, Universidad Nacional del Sur, Bahía Blanca: Buenos Aires, Argentina, 2004. [Google Scholar]
- Zalba, S.M.; Nebbia, A.J. Neosparton darwinii (Verbenaceae) a restricted endemic species. Is it also endangered? Biodivers. Conserv. 1999, 8, 1585–1593. [Google Scholar] [CrossRef]
- Pieterse, P.J.; Cairns, A.L. The population dynamics of the weed Acacia longifolia (Fabaceae) in the absence and presence of fire. Siud Afrik. Basboutydskrif 1988, 145, 25–27. [Google Scholar]
- Bowers, M.A.; Dooley, J.L. A controlled, hierarchical study of habitat fragmentation: Responses at the individual, patch, and landscape scale. Landsc. Ecol. 1999, 14, 381–389. [Google Scholar] [CrossRef]
- Mueller-Dombois, D.; Ellenberg, E. Aims and Methods of Vegetation Ecology; John Wiley & Sons: Hoboken, NJ, USA, 1974. [Google Scholar]
- Cabrera, A.L. Flora de la Provincia de Buenos Aires; Colección Científica del INTA: Buenos Aires, Argentina, 1965. [Google Scholar]
- Zuloaga, F.O.; Morrone, O.; Belgrano, M. Catálogo de las Plantas Vasculares del Cono Sur; Monographs in Systematic Botany 107; MO Botanical Garden Press: St. Louis, MI, USA, 2008. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Santos, T.; Tellería, J.L. Pérdida y fragmentación del hábitat: Efecto sobre la conservación de las especies. Ecosistemas 2006, 15, 3–12. [Google Scholar]
- Zar, J.H. Biostatistical Analysis; Prentice Hall: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat; Universidad Nacional de Córdoba: Córdoba, Argentina, 2008. [Google Scholar]
- Regan, H.M.; Crookston, J.B.; Swab, R.; Franklin, J.; Lawson, W.M. Habitat fragmentation and altered fire regime create trade-offs for an obligate seeding shrub. Ecology 2010, 91, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Driscoll, D.A.; Armenteres, D.; Bennett, A.F.; Brotons, L.; Clarke, M.F.; Doherty, T.S.; Haslem, A.; Kelly, L.K.; Sato, C.F.; Sitters, H.; et al. How fire interacts with habitat loss and fragmentation. Biol. Rev. 2021, 96, 976–998. [Google Scholar] [CrossRef]
- García, D. Efectos biológicos de la fragmentación de hábitats: Nuevas aproximaciones para resolver un viejo problema. Ecosistemas 2011, 20, 1–10. [Google Scholar]
- Fox, B.J.; Fox, M.D. The Susceptibility of Natural Communities to Invasion. In Ecology of Biological Invasions: An Australian Perspective; Groves, R.H., Burdon, J.J., Eds.; Australian Academy of Science: Canberra, Australia, 1986; pp. 57–66. [Google Scholar]
- Lozon, J.D.; MacIsaac, H.J. Biological invasions: Are they dependent on disturbance? Environ. Rev. 1997, 5, 131–144. [Google Scholar] [CrossRef]
- Didham, R.K.; Tylianakis, J.M.; Hutchison, M.A.; Ewers, R.M.; Gemmell, N.J. Are invasive species the drivers of ecological change? Trends Ecol. Evol. 2005, 20, 471–474. [Google Scholar] [CrossRef]
- Keeley, J.E. Fire and Invasive Species in Mediterranean-Climate Ecosystems of California. In Proceedings of the Invasive Species Workshop: The Role of Fire in the Control and Spread of Invasive Species; Galley, K.E.M., Wilson, T.P., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2001; pp. 81–94. [Google Scholar]
- Amodeo, M.R.; Vázquez, B.; Zalba, S.M. Generalist dispersers promote germination of an alien fleshy-fruited tree invading natural grasslands. PLoS ONE 2017, 12, e0172423. [Google Scholar] [CrossRef]
- Marozas, V. Effect of the coniferous forest–grassland edge on ground vegetation in the mixed European forest zone, Lithuania. Dendrobiology 2014, 71, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Gross, K.L.; Mittelbach, G.G.; Reynolds, H.L. Grassland invasibility and diversity: Responses to nutrients, seed input, and disturbance. Ecology 2005, 86, 476–486. [Google Scholar] [CrossRef]
- MacDougall, A.S.; Turkington, R. Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 2005, 86, 42–55. [Google Scholar] [CrossRef] [Green Version]
- Dechoum, M.; Rejmánek, M.; Castellani, T.T.; Zalba, S.M. Limited seed dispersal may explain differences in forest colonization by the Japanese raisin tree (Hovenia dulcis Thunb.), an invasive alien tree in Southern Brazil. Trop. Conserv. Sci. 2015, 8, 610–622. [Google Scholar] [CrossRef] [Green Version]
- Clements, F.E. Nature and structure of the climax. J. Ecol. 1936, 24, 252–284. [Google Scholar] [CrossRef]
- Cook, W.M.; Yao, J.; Foster, B.L.; Holt, R.D.; Patrick, L.B. Secondary succession in an experimentally fragmented landscape: Community patterns across space and time. Ecology 2005, 86, 1267–1279. [Google Scholar] [CrossRef] [Green Version]
- De las Heras, J.; Martínez, J.J.; Herranz, J.M. Impacto ecológico de los incendios forestales. Rev. De Estud. 1991, 29, 105–117. [Google Scholar]
- Rejmánek, M.; Richardson, D.M. What attributes make some plant species more invasive? Ecology 1996, 77, 1655–1661. [Google Scholar] [CrossRef]
- Tapias, R.; Gil, L.; Fuentes-Utrilla, P.; Pardos, J.A. Canopy seed banks in Mediterranean pines of south-eastern Spain: A comparison between Pinus halepensis Mill., P. pinaster Ait., P. nigra Arn. and P. pinea L. J. Ecol. 2001, 89, 629–638. [Google Scholar] [CrossRef]
- Mandle, L.; Bufford, J.; Schmidt, I.; Daehler, C. Woody exotic plant invasions and fire: Reciprocal impacts and consequences for native ecosystems. Biol. Invasions 2011, 13, 1815–1827. [Google Scholar] [CrossRef]
- García, R.A.; Franzese, J.; Policelli, N.; Sasal, Y.; Zenni, R.D.; Nuñez, M.A.; Taylor, K.; Pauchard, A. Non-Native Pines are Homogenizing the Ecosystems of South America. In From Biocultural Homogenization to Biocultural Conservation. Ecology and Ethics; Rozzi, R., May, R.H., Chapin, F.S., Massardo, F., Gavin, M.C., Klaver, I.J., Pauchard, A., Nuñez, M.A., Simberloff, D., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Elton, C.S. The Ecology of Invasions by Animals and Plants; Chapman and Hall: London, UK, 1958. [Google Scholar]
- Tilman, D. Community invasibility, recruitment limitation and grassland biodiversity. Ecology 1997, 78, 81–92. [Google Scholar] [CrossRef]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).