
Patterns of Genetic Variation in the Eisenia nordenskioldi Complex (Oligochaeta: Lumbricidae) along an Elevation Gradient in Northern China

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
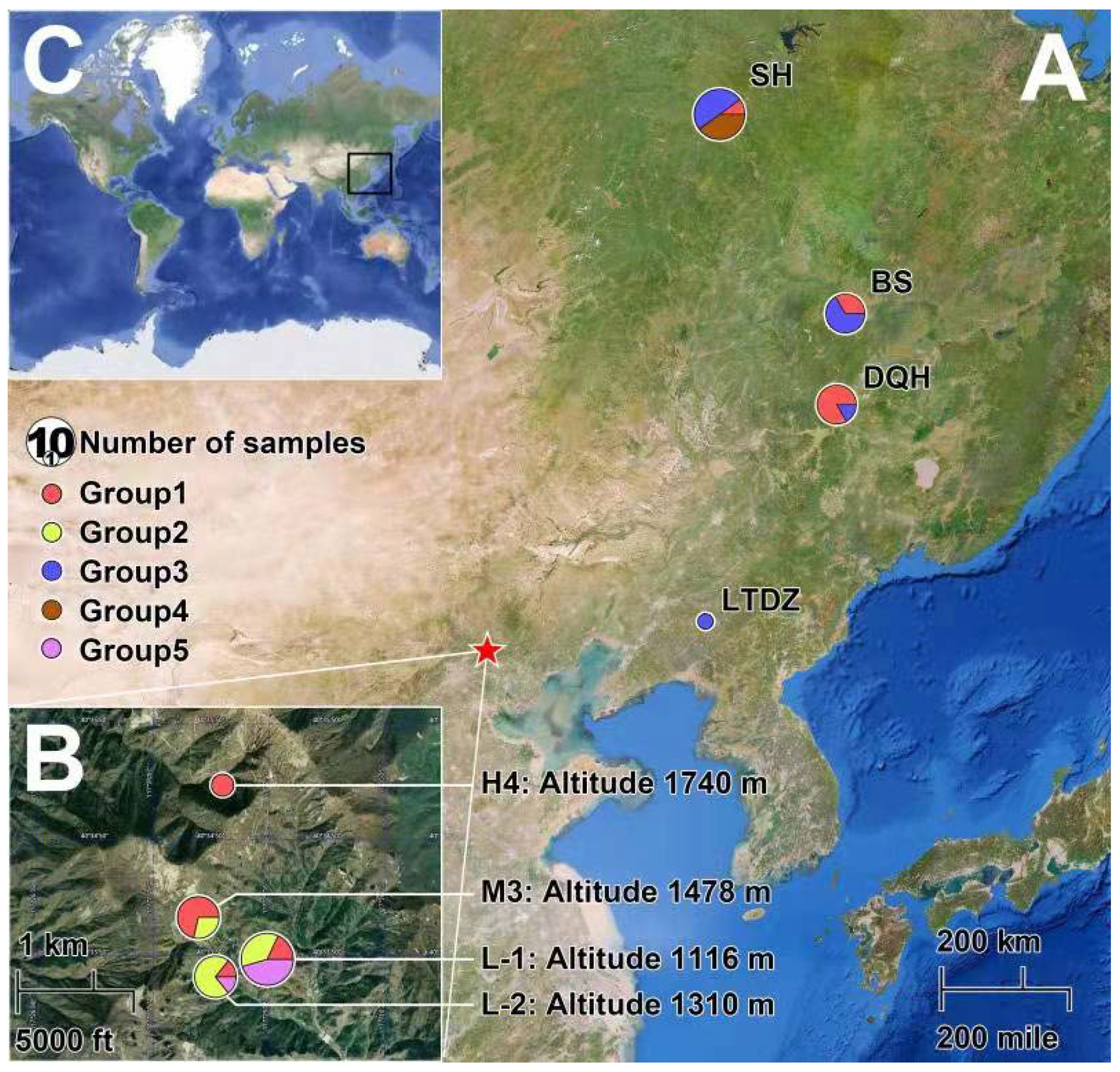
2.1. Sample Collection and DNA Extraction
2.2. Sequence Analysis
3. Results
3.1. Distribution of the E. nordenskioldi Complex along an Elevation Gradient
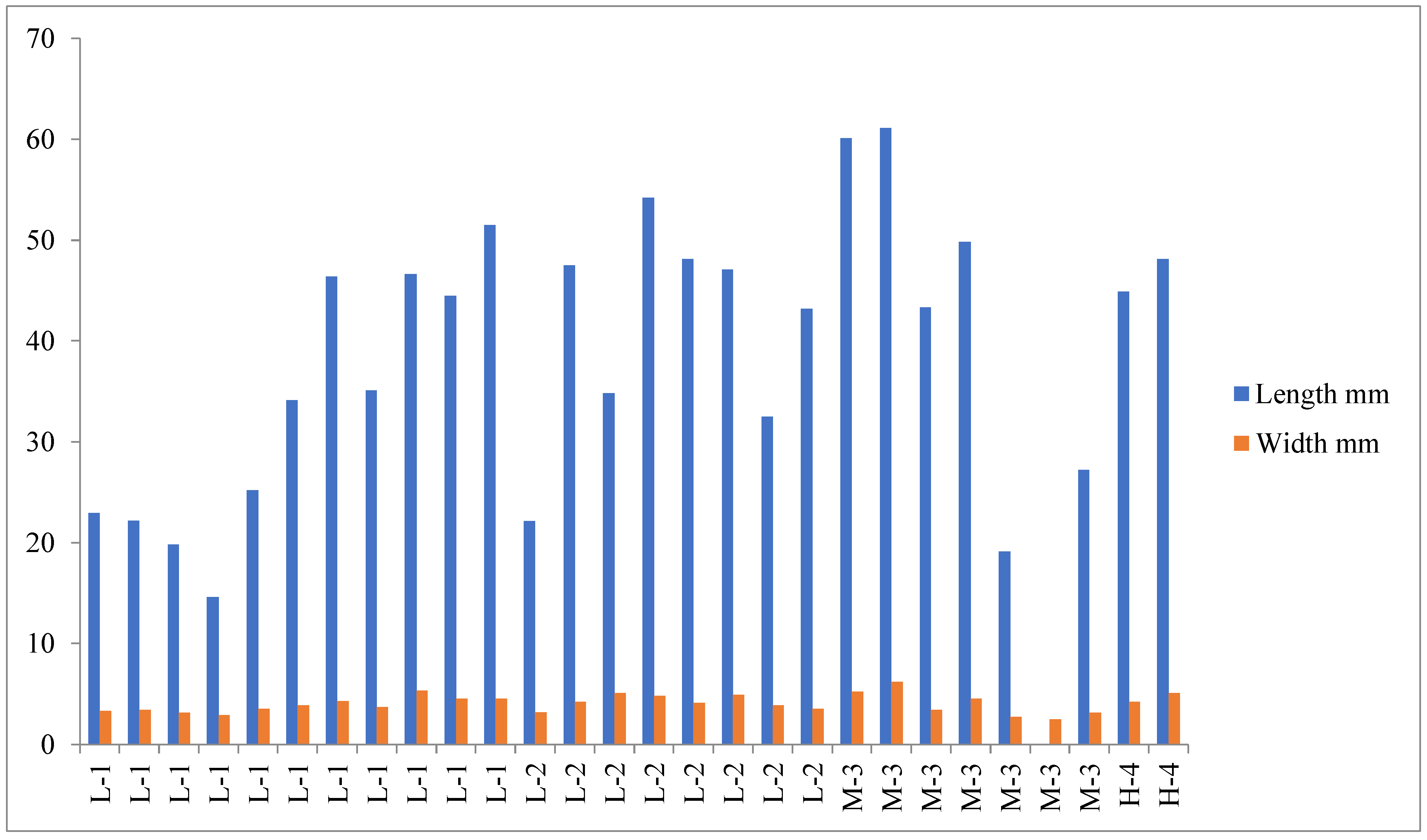
3.2. External Morphological Data of E. nordenskioldi Complex from Mt. Wuling
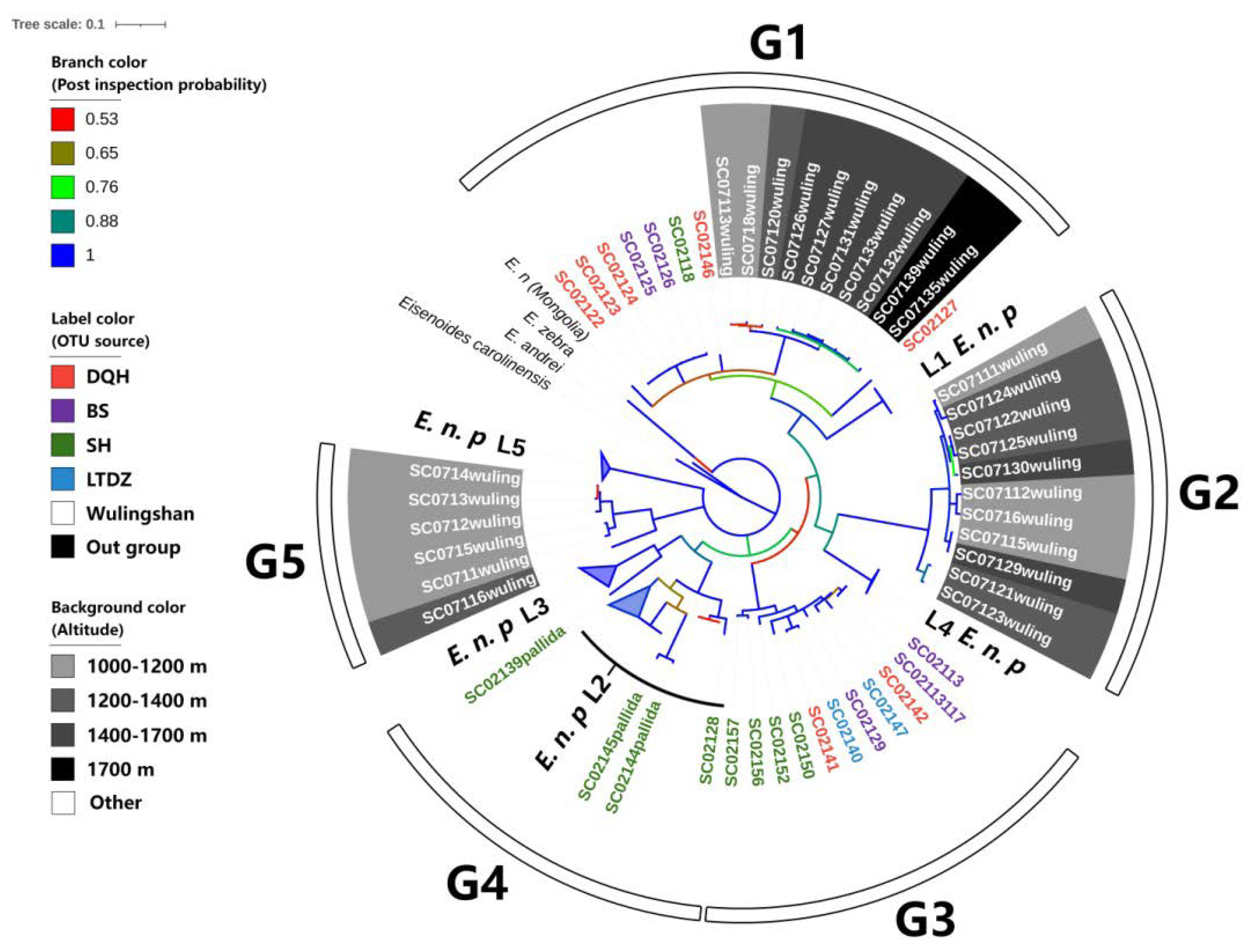
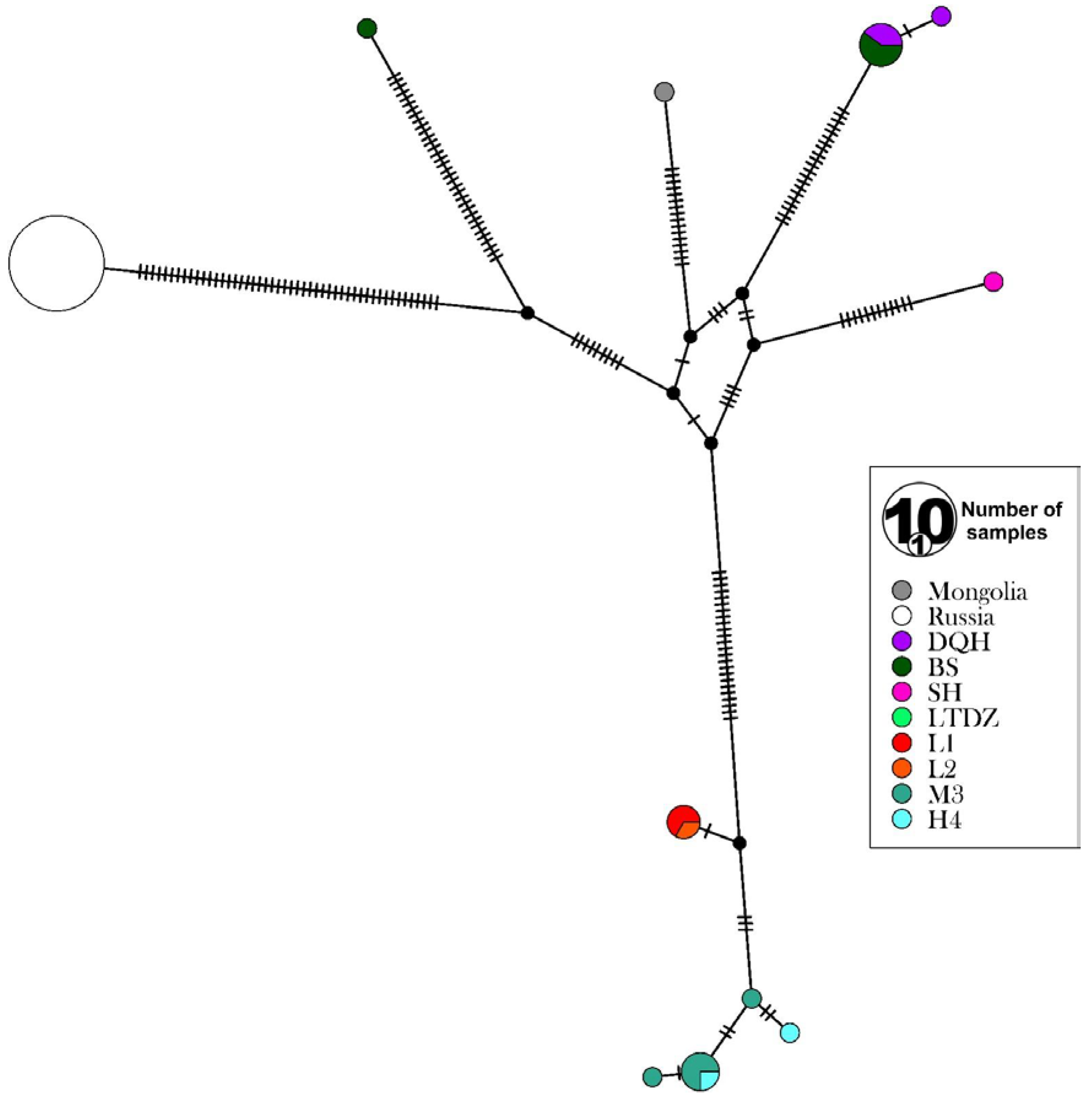
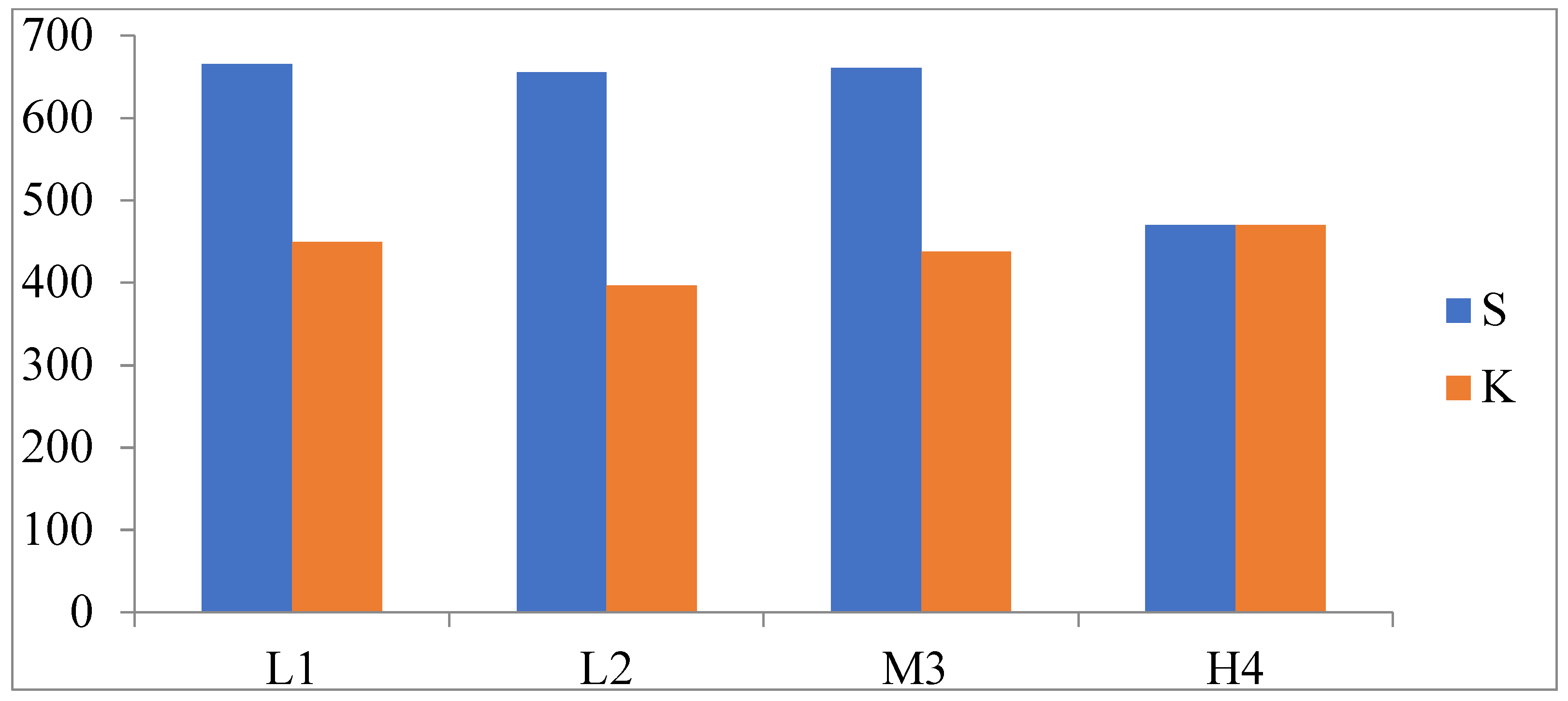
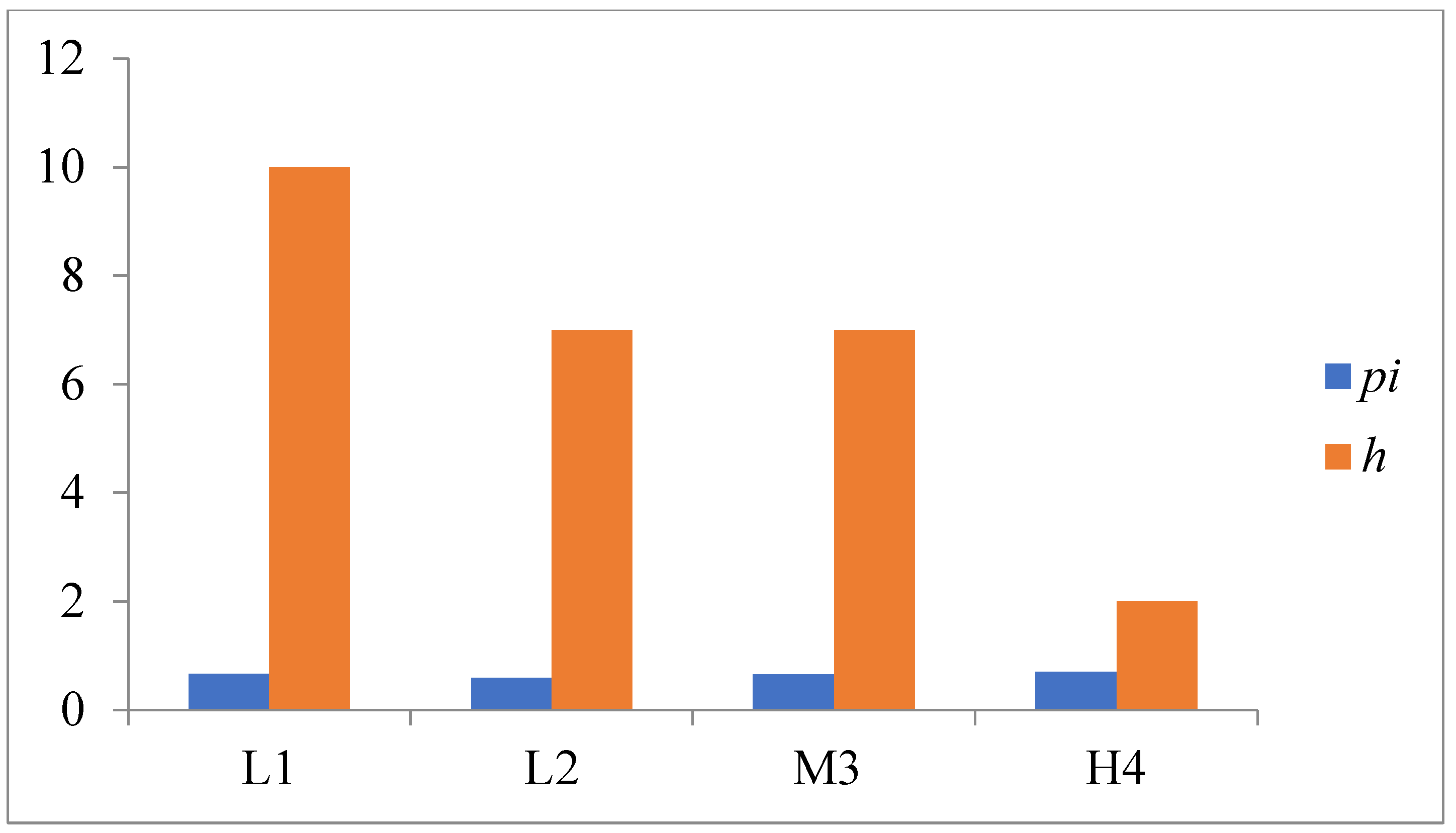
3.3. Genetic Diversity and Differentiation of the Mt. Wuling E. nordenskioldi Complex
4. Discussion
4.1. Morphological and Genetic Variations within the E. nordenskioldi Complex According to Elevation
4.2. Distribution of the E. nordenskioldi Complex from Mt. Wuling
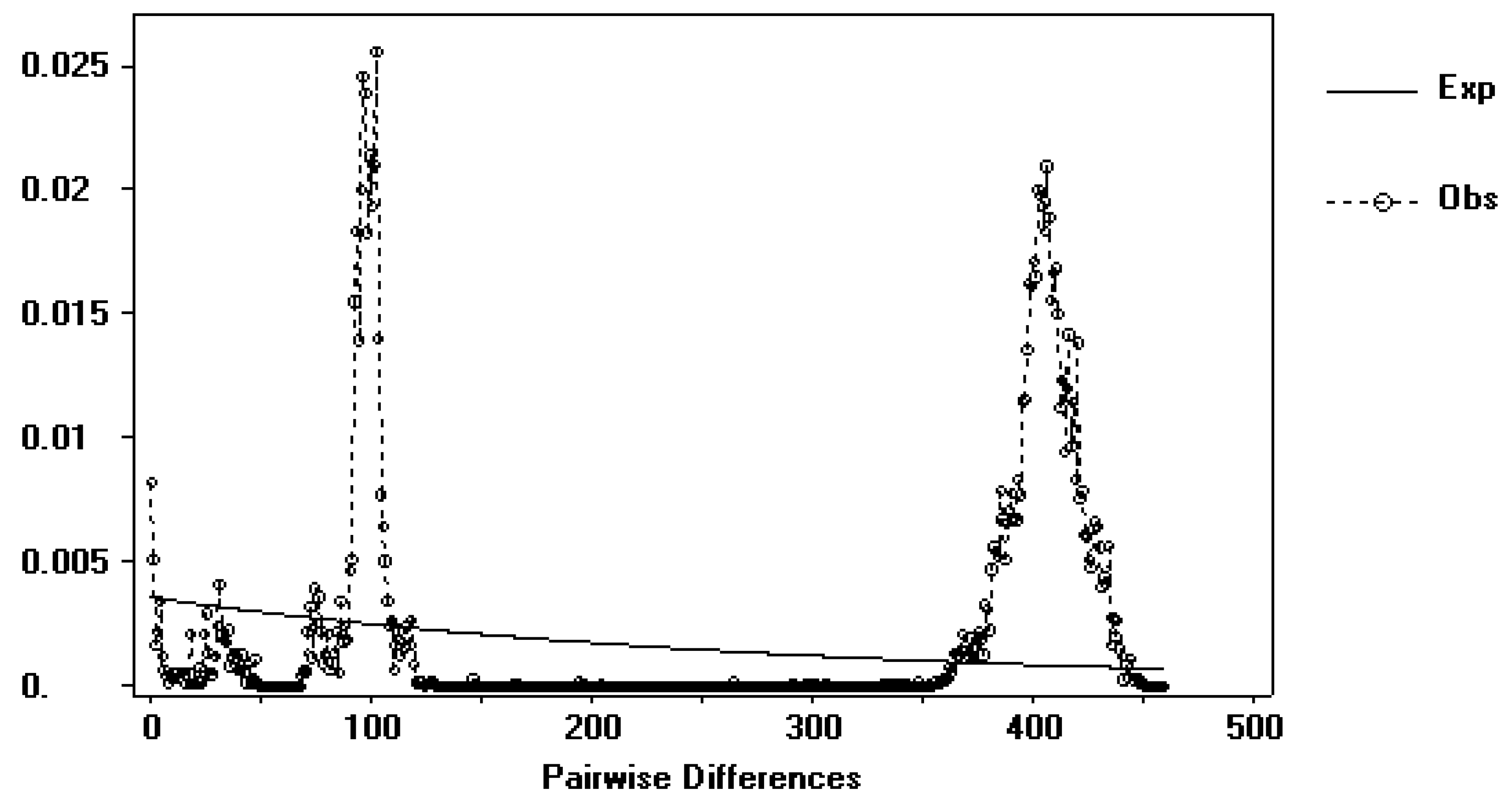
4.3. Phylogeography of the E. nordenskioldi Complex from Mt. Wuling
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Xu, S.B.; Li, Y.H.; Zhu, L.Q.; Li, L. A review on the vertical distribution patterns of soil communities in Mountains. J. Henan Univ. (Nat. Sci.) 2020, 50, 19–28. [Google Scholar]
- Xu, G.R.; Zhang, Y.X.; Zhang, S.; Ma, K. Effect of Elevation on Abundance Distribution of Different Feeding Groups in Litter-dwelling Soil Fauna. Acta Pedol. Sin. 2017, 54, 237–245. [Google Scholar]
- Novo, M.; Cunha, L.; Maceda-Veiga, A.; Talavera, J.A.; Hodson, M.E.; Spurgeon, D.; Bruford, M.W.; Morgan, A.J.; Kille, P. Multiple introductions and environmental factors affecting the establishment of invasive species on a volcanic island. Soil Biol. Biochem. 2015, 85, 89–100. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, G.; Garcia, E.; Cruz, V.; Sonia, B.; Marcela, Z.; María, M.; Rivera, M.M. Earthworms communities along an elevation gradient in Northeastern Puerto Rico. Eur. J. Soil Biol. 2007, 43, 24–32. [Google Scholar] [CrossRef]
- Jiao, L.; Wang, X.D.; Huang, L.; Fei, X. Vertical Distribution Types and Profile Characteristics of soils in Wuling Mountain. J. Cap. Norm. Univ. (Nat. Sci. Ed.) 2011, 32, 69–72. [Google Scholar]
- Vsevolodova-Perel, T.S. The Earthworms of the Fauna of Russia; Nauka: Moscow, Russia, 1997. [Google Scholar]
- Shekhovtsov, S.V.; Golovanova, E.V.; Ershov, N.I.; Poluboyarova, T.V.; Berman, D.I.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Phylogeny of the Eisenia nordenskioldi complex based on mitochondrial genomes. Eur. J. Soil Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Peltek, S.E. Phylogeography of the earthworm Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) in northeastern Eurasia. Dokl. Biol. Sci. 2015, 461, 1–4. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Bazarova, N.E.; Bulakhova, N.A.; Porco, D.; Peltek, S.E. Cryptic genetic lineages in Eisenia nordenskioldi pallida (Oligochaeta, Lumbricidae). Eur. J. Soil Biol. 2016, 75, 151–156. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Berman, D.I.; Golovanova, E.V.; Peltek, S.E. Genetic diversity of the earthworm Eisenia nordenskioldi (Lumbricidae, Annelida), Vavilovskii Zhurnal Genetiki i Selektsii, Vavilov. J. Genet. Breed. 2017, 21, 589–595. [Google Scholar] [CrossRef] [Green Version]
- Shekhovtsov, S.V.; Berman, D.I.; Bulakhova, N.A.; Vinokurov, N.N.; Peltek, S.E. Phylogeography of Eisenia nordenskioldi nordenskioldi (Lumbricidae, Oligochaeta) from the north of Asia. Polar Biol. 2018, 41, 237–247. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Golovanova, E.V.; Peltek, S.E. Cryptic diversity within the Nordenskiold’s earthworm, Eisenia nordenskioldi subsp. nordenskioldi (Lumbricidae, Annelida). Eur. J. Soil Biol. 2013, 58, 13–18. [Google Scholar] [CrossRef]
- Shekhovtsov, S.V.; Shipova, A.A.; Poluboyarova, T.V.; Geraskina, A.P.; Peltek, S.E. Species delimitation of the Eisenia nordenskioldi complex (Oligochaeta, Lumbricidae) using transcriptomic data. Front. Genet. 2020, 11, 1508. [Google Scholar] [CrossRef]
- Vsevolodova-Perel, T.S.; Leirikh, A.N. Distribution and ecology of the earthworm Eisenia nordenskioldi pallida (Oligochaeta, Lumbricidae) dominant in southern Siberia and the Russian Far East. Entomol. Rev. 2014, 93, 45–52. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Blakemore, R.J. Earthworms newly from mongolia (Oligochaeta, Lumbricidae, Eisenia). Zookeys 2013, 285, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nylander, J.A.A. MrAIC.pl. Program Distributed by the Author; Evolutionary Biology Centre, Uppsala University: Uppsala, Sweden, 2004. [Google Scholar]
- Hong, Y.; Csuzdi, C. New Data to the Earthworm Fauna of the Korean Peninsula with Redescription of Eisenia koreana (Zicsi) and Remarks on the Eisenia nordenskioldi Species Group (Oligochaeta, Lumbricidae). Zool. Stud. 2016, 55, e12. [Google Scholar] [CrossRef] [PubMed]
- Tsai, Z.F.; Shen, H.P.; Cai, S.C. Endemicity and Altitudinal Stratification in Distribution of Megascolecid Earthworms in the Centro-western Taiwan. Endem. Species Res. 2004, 6, 1–18. [Google Scholar]
- Dong, Y.; Jiang, J.B.; Yuan, Z.; Zhao, Q.; Qiu, J.P. Population Genetic Structure Reveals Two Lineages of Amynthas triastriatus (Oligochaeta:Megascolecidae) in China, with Notes on a New Subspecies of Amynthas triastriatus. Int. J. Environ. Res. Public Health 2020, 17, 1538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perel, T.S. Range and Regularities in the Distribution of Earthworms of the USSR Fauna; Nauka: Moscow, Russia, 1979. [Google Scholar]
- Elias, S.A. Alpine Insect Distribution Patterns. Encycl. World’s Biomes 2020, 1, 430–440. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Code | No. Specimens | N. Latitude | E. Longitude | Elevation m | Habitat |
|---|---|---|---|---|---|---|
| Mt. Wuling Nature Reserve in Xinglong, Hebei | L-1 | 11 | 40.5634 | 117.4873 | 1116 | Litter in mixed-forest |
| Mt. Wuling Nature Reserve in Xinglong, Hebei | L-2 | 8 | 40.5609 | 117.4777 | 1310 | Litter in mixed-forest |
| Mt. Wuling Nature Reserve in Xinglong, Hebei | M3 | 7 | 40.5694 | 117.4743 | 1478 | Litter in coniferous |
| Mt. Wuling Nature Reserve in Xinglong, Hebei | H4 | 2 | 40.5878 | 117.4795 | 1740 | Litter in coniferous |
| Danqinghe Nature Reserve in Heilongjiang | DQH | 7 | 46°37′ | 129°22′ | 482 | Litter in mixed-forest |
| Baoshan Nature Reserve in Heilongjiang | BS | 5 | 48°57′ | 128°52′ | 405 | Litter in mixed-forest |
| Shuanghe Nature Reserve in Heilongjiang | SH | 9 | 52°59′ | 125°22′ | 354 | Litter in mixed-forest |
| Laotudingzi Nature Reserve in Heilongjiang | LTDZ | 2 | 41°19′ | 124°54′ | 906 | Litter in mixed-forest |
| Russia | Mongolia | N.E China | Mt. Wuling | |
|---|---|---|---|---|
| S | 221 | 508 | 545 | 545 |
| h | 56 | 3 | 22 | 28 |
| K | 84.953 | 413.667 | 352.158 | 342.783 |
| M | 381 | 826 | 1598 | 1373 |
| Np | 221 | 577 | 545 | 545 |
| Hd | 0.979 | 1 | 0.996 | 1 |
| Pi | 0.156 | 0.759 | 0.646 | 0.629 |
| S | ps | Θ | π | D |
|---|---|---|---|---|
| 545 | 1 | 0.174417 | 0.512729 | 6.288815 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, Y.; Wu, H.; Li, C.; Aspe, N.M.; Wu, D. Patterns of Genetic Variation in the Eisenia nordenskioldi Complex (Oligochaeta: Lumbricidae) along an Elevation Gradient in Northern China. Diversity 2022, 14, 35. https://doi.org/10.3390/d14010035
Zhang Y, Zhang Y, Wu H, Li C, Aspe NM, Wu D. Patterns of Genetic Variation in the Eisenia nordenskioldi Complex (Oligochaeta: Lumbricidae) along an Elevation Gradient in Northern China. Diversity. 2022; 14(1):35. https://doi.org/10.3390/d14010035
Chicago/Turabian StyleZhang, Yufeng, Yiming Zhang, Hui Wu, Congsheng Li, Nonillon M. Aspe, and Donghui Wu. 2022. "Patterns of Genetic Variation in the Eisenia nordenskioldi Complex (Oligochaeta: Lumbricidae) along an Elevation Gradient in Northern China" Diversity 14, no. 1: 35. https://doi.org/10.3390/d14010035
APA StyleZhang, Y., Zhang, Y., Wu, H., Li, C., Aspe, N. M., & Wu, D. (2022). Patterns of Genetic Variation in the Eisenia nordenskioldi Complex (Oligochaeta: Lumbricidae) along an Elevation Gradient in Northern China. Diversity, 14(1), 35. https://doi.org/10.3390/d14010035