
Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Molecular Phylogenetics Techniques
3. Contributions of Molecular Phylogenetics to Lumbricidae Systematics
3.1. Advances
3.2. Remaining Questions
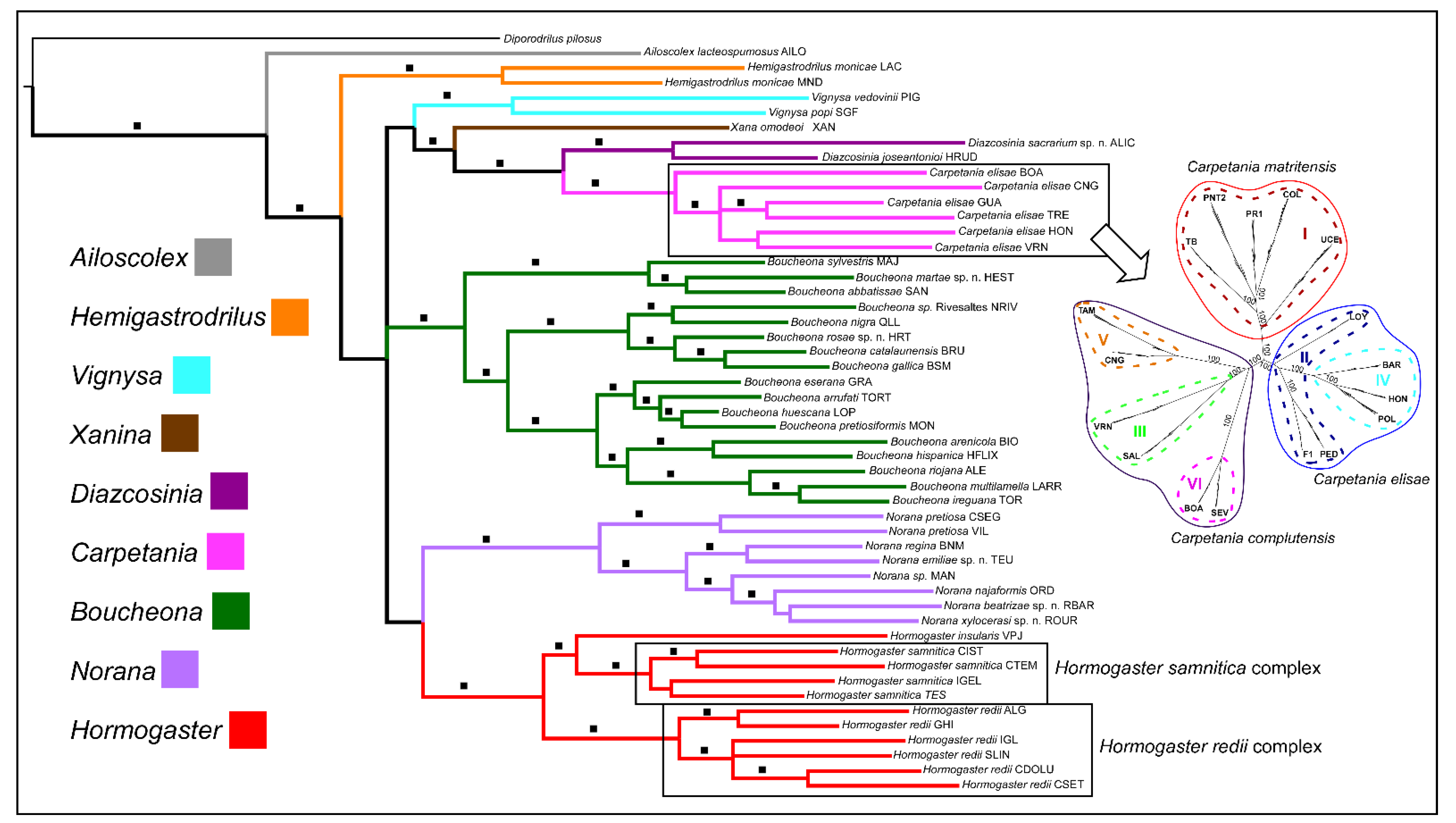
4. Contributions of Molecular Phylogenetics to the Hormogastridae Systematics
4.1. Advances

4.2. Remaining Questions
5. Remaining Questions of Lumbricoidea
6. Other Families: Remaining Questions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Pop, A.A.; Wink, M.; Pop, V.V. Use of 18S, 16S rDNA and cytochrome c oxidase sequences in earthworm taxonomy (Oligochaeta, Lumbricidae): The 7th international symposium on earthworm ecology·Cardiff·Wales·2002. Pedobiologia 2003, 47, 428–433. [Google Scholar] [CrossRef]
- Michaelsen, W. Oligochaeta. In Das Tierreich X.; Friedländer & Sohn: Berlin/Heidelberg, Germany, 1900; pp. 1–575. [Google Scholar]
- Pop, V. Zur phylogenie und Systematik der Lumbriciden. Zool. Jahrbücher Abt. Für Syst. Okol. Und Geogr. Der Tiere 1941, 74, 487–522. [Google Scholar]
- Omodeo, P. Contributo alla revisione dei Lumbricidae. Archivio Zoologico Italiano 1956, 41, 129–212. [Google Scholar]
- Gates, G.E. Contributions to a revision of the earthworm family Lumbricidae XII. Enterion mammale Savigny, 1826 and its position in the family. Megadrilogica 1975, 2, 1–5. [Google Scholar]
- Perel, T.S. A critical analysis of the system of family Lumbricidae. Zoologicheski Zhurnal 1976, 55, 823–836. (In Russian) [Google Scholar]
- Mršić, N. Monograph on Earthworms (Lumbricidae) of the Balkans I–II; Slovenska Akademija Znanosti in Umetnosti, Zazred za Naravoslovne Vede Opera. 31: Ljubljana, Slovenia, 1991; pp. 1–757. [Google Scholar]
- Qiu, J.P.; Bouché, M.B. Liste classée des taxons valides de Lombriciens (Oligochaeta: Lumbricoidea) apres l’etude des trios cinquieme d’entre-eux. Doc. Pédozoologiques Intégrologiques 1998, 4, 181–200. [Google Scholar]
- James, S.W.; Davidson, S.K. Molecular phylogeny of earthworms (Annelida: Crassiclitellata) based on 28S, 18S and 16S gene sequences. Invertebr. Syst. 2012, 26, 213–229. [Google Scholar] [CrossRef]
- Decaëns, T.; Porco, D.; Rougerie, R.; Brown, G.G.; James, S.W. Potential of DNA barcoding for earthworm research in taxonomy and ecology. Appl. Soil Ecol. 2013, 65, 35–42. [Google Scholar] [CrossRef]
- Hebert, P.D.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- King, R.A.; Tibble, A.L.; Symondson, W.O.C. Opening a can of worms: Unprecedented sympatric cryptic diversity within British lumbricid earthworms. Mol. Ecol. 2008, 17, 4684–4698. [Google Scholar] [CrossRef]
- Porco, D.; Chang, C.-H.; Dupont, L.; James, S.W.; Richard, B.; Decaëns, T. A reference library of DNA barcodes for the earthworms from Upper Normandy: Biodiversity assessment, new records, potential cases of cryptic diversity and ongoing speciation. Appl. Soil Ecol. 2018, 124, 362–371. [Google Scholar] [CrossRef]
- Novo, M.; Almodóvar, A.; Fernández, R.; Trigo, D.; Cosín, D.J.D. Cryptic speciation of hormogastrid earthworms revealed by mitochondrial and nuclear data. Mol. Phylogenet. Evol. 2010, 56, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Buckley, T.R.; James, S.; Allwood, J.; Bartlam, S.; Howitt, R.; Prada, D. Phylogenetic analysis of New Zealand earthworms (Oligochaeta: Megascolecidae) reveals ancient clades and cryptic taxonomic diversity. Mol. Phylogenetics Evol. 2011, 58, 85–96. [Google Scholar] [CrossRef]
- Marchán, D.F.; Hedde, M.; Lapied, E.; Maggia, M.E.; Novo, M.; Domínguez, J.; Decaëns, T. Contrasting phylogeographic patterns of earthworms (Crassiclitellata, Lumbricidae) on near-shore mediterranean islands. Eur. J. Soil Biol. 2020, 101, 103242. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Bloch, R.; Breinholt, J.W.; Pfenninger, M.; Domínguez, J. Taxonomic assessment of Lumbricidae (Oligochaeta) earthworm genera using DNA barcodes. Eur. J. Soil Biol. 2012, 48, 41–47. [Google Scholar] [CrossRef]
- Jamieson, B.G.; Tillier, S.; Tillier, A.; Justine, J.L.; Ling, E.; James, S.; McDonald, K.; Hugall, A.F. Phylogeny of the Megascolecidae and Crassiclitellata (Annelida, Oligochaeta): Combined versus partitioned analysis using nuclear (28S) and mitochondrial (12S, 16S) rDNA. Zoosystema 2002, 24, 707–734. [Google Scholar]
- Klarica, J.; Kloss-Brandstätter, A.; Traugott, M.; Juen, A. Comparing four mitochondrial genes in earthworms–implications for identification, phylogenetics, and discovery of cryptic species. Soil Biol. Biochem. 2012, 45, 23–30. [Google Scholar] [CrossRef]
- Fernández, R.; Almodóvar, A.; Novo, M.; Simancas, B.; Cosín, D.J.D. Adding complexity to the complex: New insights into the phylogeny, diversification and origin of parthenogenesis in the Aporrectodea caliginosa species complex (Oligochaeta, Lumbricidae). Mol. Phylogenetics Evol. 2012, 64, 368–379. [Google Scholar] [CrossRef]
- Novo, M.; Almodovar, A.; Fernandez, R.; Trigo, D.; Díaz-Cosín, D.J.; Giribet, G. Appearances can be deceptive: Different diversification patterns within a group of Mediterranean earthworms (Oligochaeta, Hormogastridae). Mol. Ecol. 2012, 21, 3776–3793. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Porto, P.G.; Cosín, D.J.D.; Pérez-Losada, M. Multigene phylogeny reveals two new isolated and relic earthworm genera (Oligochaeta: Lumbricidae). Zool. J. Linn. Soc. 2017, 182, 258–274. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, J.; Dong, Y.; Qiu, J. Complete mitochondrial genome of four pheretimoid earthworms (Clitellata: Oligochaeta) and their phylogenetic reconstruction. Gene 2015, 574, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Shekhovtsov, S.V.; Golovanova, E.V.; Ershov, N.I.; Poluboyarova, T.V.; Berman, D.I.; Bulakhova, N.A.; Szederjesi, T.; Peltek, S.E. Phylogeny of the Eisenia nordenskioldi complex based on mitochondrial genomes. Eur. J. Soil Biol. 2020, 96, 103137. [Google Scholar] [CrossRef]
- Dupont, L.; Porco, D.; Symondson, W.O.C.; Roy, V. Hybridization relics complicate barcode-based identification of species in earthworms. Mol. Ecol. Resour. 2016, 16, 883–894. [Google Scholar] [CrossRef]
- Taheri, S.; James, S.; Roy, V.; Decaëns, T.; Williams, B.W.; Anderson, F.; Rougerie, R.; Chang, C.H.; Brown, G.; Cunha, L.; et al. Complex taxonomy of the ‘brush tail’peregrine earthworm Pontoscolex corethrurus. Mol. Phylogenet. Evol. 2018, 124, 60–70. [Google Scholar] [CrossRef]
- Novo, M.; Fernández, R.; Andrade, S.C.; Marchán, D.F.; Cunha, L.; Cosín, D.J.D. Phylogenomic analyses of a Mediterranean earthworm family (Annelida: Hormogastridae). Mol. Phylogenet. Evol. 2016, 94, 473–478. [Google Scholar] [CrossRef] [PubMed]
- Anderson, F.E.; Williams, B.W.; Horn, K.M.; Erséus, C.; Halanych, K.M.; Santos, S.R.; James, S.W. Phylogenomic analyses of Crassiclitellata support major Northern and Southern Hemisphere clades and a Pangaean origin for earthworms. BMC Evol. Biol. 2017, 17, 123. [Google Scholar]
- Faircloth, B.C.; Glenn, T.C. Not All Sequence Tags Are Created Equal: Designing and Validating Sequence Identification Tags Robust to Indels. PLoS ONE 2012, 7, e42543. [Google Scholar] [CrossRef]
- Lemmon, A.R.; Emme, S.A.; Lemmon, E.M. Anchored Hybrid Enrichment for Massively High-Throughput Phylogenomics. Syst. Biol. 2012, 61, 727–744. [Google Scholar] [CrossRef] [PubMed]
- Karin, B.R.; Gamble, T.; Jackman, T.R. Optimizing Phylogenomics with Rapidly Evolving Long Exons: Comparison with Anchored Hybrid Enrichment and Ultraconserved Elements. Mol. Biol. Evol. 2020, 37, 904–922. [Google Scholar] [CrossRef]
- Crawford, N.G.; Parham, J.F.; Sellas, A.B.; Faircloth, B.C.; Glenn, T.C.; Papenfuss, T.J.; Henderson, J.B.; Hansen, M.H.; Simison, W.B. A phylogenomic analysis of turtles. Mol. Phylogenet. Evol. 2015, 83, 250–257. [Google Scholar] [CrossRef]
- Streicher, J.W.; Wiens, J.J. Phylogenomic analyses of more than 4000 nuclear loci resolve the origin of snakes among lizard families. Biol. Lett. 2017, 13, 20170393. [Google Scholar] [CrossRef]
- Phillips, A.J.; Dornburg, A.; Zapfe, K.L.; Anderson, F.E.; James, S.W.; Erséus, C.; Lemmon, E.M.; Lemmon, A.R.; Williams, B.W. Phylogenomic Analysis of a Putative Missing Link Sparks Reinterpretation of Leech Evolution. Genome Biol. Evol. 2019, 11, 3082–3093. [Google Scholar] [CrossRef]
- Emerson, K.J.; Merz, C.R.; Catchen, J.M.; Hohenlohe, P.A.; Cresko, W.A.; Bradshaw, W.E.; Holzapfel, C.M. Resolving postglacial phylogeography using high-throughput sequencing. Proc. Natl. Acad. Sci. USA 2010, 107, 16196–16200. [Google Scholar] [CrossRef] [PubMed]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.A.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef]
- Anderson, C.; Cunha, L.; Sechi, P.; Kille, P.; Spurgeon, D. Genetic variation in populations of the earthworm, Lumbricus rubellus, across contaminated mine sites. BMC Genet. 2017, 18, 97. [Google Scholar] [CrossRef]
- Marchán, D.F.; Novo, M.; Sánchez, N.; Domínguez, J.; Cosín, D.J.D.; Fernandez, R. Local adaptation fuels cryptic speciation in terrestrial annelids. Mol. Phylogenetics Evol. 2020, 146, 106767. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernández, R.; Domínguez, J.; Díaz Cosín, D.J.; Novo, M. Genome-informed integrative taxonomic description of three cryptic species in the earthworm genus Carpetania (Oligochaeta, Hormogastridae). Syst. Biodivers. 2020, 18, 203–215. [Google Scholar] [CrossRef]
- Yuan, Z.; Jiang, J.; Dong, Y.; Zhao, Q.; Sun, J.; Qiu, J. Unearthing the genetic divergence and gene flow of the earthworm Amynthas_YN2017 sp. (Oligochaeta: Megascolecidae) populations based on restriction site-associated DNA sequencing. Eur. J. Soil Biol. 2020, 99, 103210. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef]
- Marchan, D.F.; Fernández, R.; de Sosa, I.; Cosin, D.J.D.; Novo, M. Pinpointing cryptic borders: Fine-scale phylogeography and genetic landscape analysis of the Hormogaster elisae complex (Oligochaeta, Hormogastridae). Mol. Phylogenet. Evol. 2017, 112, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Novo, M.; Almodóvar, A.; Fernández, R.; Giribet, G.; Cosín, D.J.D. Understanding the biogeography of a group of earthworms in the Mediterranean basin—The phylogenetic puzzle of Hormogastridae (Clitellata: Oligochaeta). Mol. Phylogenet. Evol. 2011, 61, 125–135. [Google Scholar] [CrossRef]
- Shen, H.-P.; Chang, C.-H.; Ota, H. The biogeographical history of giant earthworms of the Metaphire formosae species group (Clitellata: Megascolecidae) in Taiwan and the Ryukyu Archipelago, with the description of a new species from Yonagunijima, Southern Ryukyus. Org. Divers. Evol. 2021. [Google Scholar] [CrossRef]
- Zhao, Q.; Cluzeau, D.; Jiang, J.; Petit, E.J.; Briard, C.; Sun, J.; Prinzing, A.; Qiu, J. Molecular Phylogeny of Pheretimoid Earthworms (Haplotaxina: Megascolecidae) Based on Mitochondrial DNA in Hainan Island, China. Mol. Biol. 2015, 4, 4. [Google Scholar] [CrossRef]
- De Sosa, I.; Cosín, D.J.D.; Csuzdi, C.; Paoletti, M.G.; Marchán, D.F. Placing Eophila tellinii (Oligochaeta, Lumbricidae) in a molecular phylogenetic context advances the century-old controversy around the problematic genus. Eur. J. Soil Biol. 2019, 94, 103114. [Google Scholar] [CrossRef]
- Marchán, D.F.; Csuzdi, C.; Decaëns, T.; Szederjesi, T.; Pizl, V.; Domínguez, J. The disjunct distribution of relict earthworm genera clarifies the early historical biogeography of the Lumbricidae (Crassiclitellata, Annelida). J. Zoöl. Syst. Evol. Res. 2021, 59, 1703–1717. [Google Scholar] [CrossRef]
- Chen, D.; Hosner, P.A.; Dittmann, D.L.; O’Neill, J.P.; Birks, S.M.; Braun, E.L.; Kimball, R.T. Divergence time estimation of Galliformes based on the best gene shopping scheme of ultraconserved elements. BMC Ecol. Evol. 2021, 21, 209. [Google Scholar] [CrossRef]
- Domínguez, J.; Aira, M.; Breinholt, J.W.; Stojanovic, M.; James, S.W.; Pérez-Losada, M. Underground evolution: New roots for the old tree of lumbricid earthworms. Mol Phylogenet Evol. 2015, 83, 7–19. [Google Scholar] [CrossRef]
- Jiménez Pinadero, S.; Marchán, D.F.; Novo, M.; Trigo, D.; Domínguez, J.; Cosín, D.J.D. Sorry atlanticus, you are not my type: Molecular assessment splits Zophoscolex (Lumbricidae: Crassiclitellata) into French and Iberian genera. Zool. J. Linn. Soc. 2021. [Google Scholar] [CrossRef]
- Marchán, D.F.; Jiménez, S.; Decaëns, T.; Domínguez, J. Systematic revision of Gatesona (Crassiclitellata, Lumbricidae), an endemic earthworm genus from the Massif Central (France). PLoS ONE 2021, 16, e0255978. [Google Scholar] [CrossRef]
- Csuzdi, C.; Chang, C.-H.; Pavlícek, T.; Szederjesi, T.; Esopi, D.; Szlavecz, K. Molecular phylogeny and systematics of native North American lumbricid earthworms (Clitellata: Megadrili). PLoS ONE 2017, 12, e0181504. [Google Scholar] [CrossRef]
- Pérez-Losada, M.; Ricoy, M.; Marshall, J.C.; Domínguez, J. Phylogenetic assessment of the earthworm Aporrectodea caliginosa species complex (Oligochaeta: Lumbricidae) based on mitochondrial and nuclear DNA sequences. Mol. Phylogenet. Evol. 2009, 52, 293–302. [Google Scholar] [CrossRef]
- Fernández, R.; Novo, M.; Marchán, D.F.; Cosín, D.J.D. Diversification patterns in cosmopolitan earthworms: Similar mode but different tempo. Mol. Phylogenet. Evol. 2016, 94, 701–708. [Google Scholar] [CrossRef]
- Dupont, L.; Lazrek, F.; Porco, D.; King, R.; Rougerie, R.; Symondson, W.; Livet, A.; Richard, B.; Decaëns, T.; Butt, K.; et al. New insight into the genetic structure of the Allolobophora chlorotica aggregate in Europe using microsatellite and mitochondrial data. Pedobiologia 2011, 54, 217–224. [Google Scholar] [CrossRef]
- Marchán, D.F.; Cosín, D.J.D.; Novo, M. Why are we blind to cryptic species? Lessons from the eyeless. Eur. J. Soil Biol. 2018, 86, 49–51. [Google Scholar] [CrossRef]
- Szederjesi, T.; Pop, V.V.; Pavlíček, T.; MÁrton, O.; KrÍzsik, V.; Csuzdi, C. Integrated taxonomy reveals multiple species in the Dendrobaena byblica (Rosa, 1893) complex (Oligochaeta: Lumbricidae). Zool. J. Linn. Soc. 2018, 182, 500–516. [Google Scholar] [CrossRef]
- Martinsson, S.; Klinth, M.; Erséus, C. Distribution and genetic diversity of two earthworms, Helodrilus oculatus and Satchellius mammalis (Clitellata: Lumbricidae) in Scandinavia. Fauna Nor. 2021, 41, 1–14. [Google Scholar] [CrossRef]
- Marchán, D.F.; Decaëns, T.; Cosín, D.J.D.; Hedde, M.; Lapied, E.; Domínguez, J. French Mediterranean islands as a refuge of relic earthworm species: Cataladrilus porquerollensis sp. nov. and Scherotheca portcrosana sp. nov. (Crassiclitellata, Lumbricidae). Eur. J. Taxon. 2020, 701, 1–22. [Google Scholar] [CrossRef]
- Bache, F.; Olivet, J.L.; Gorini, C.; Aslanian, D.; Labails, C.; Rabineau, M. Evolution of rifted continental margins: The case of the Gulf of Lions (Western Mediterranean Basin). Earth Planet. Sci. Lett. 2010, 292, 345–356. [Google Scholar] [CrossRef]
- Cosin, D.D.; Novo, M.; Fernández, R.; Marchán, D.F.; Gutiérrez, M. A new earthworm species within a controversial genus: Eiseniona gerardoi sp. n. (Annelida, Lumbricidae)—Description based on morphological and molecular data. ZooKeys 2014, 399, 71–87. [Google Scholar] [CrossRef]
- Novo, M.; Fernández, R.; Marchán, D.F.; Gutiérrez, M.; Cosín, D.J.D. Compilation of morphological and molecular data, a necessity for taxonomy: The case of Hormogaster abbatissae sp. n.(Annelida, Clitellata, Hormogastridae). ZooKeys 2012, 242, 1. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernández, R.; Novo, M.; Cosin, D.D. New light into the hormogastrid riddle: Morphological and molecular description of Hormogaster joseantonioi sp. n. (Annelida, Clitellata, Hormogastridae). ZooKeys 2014, 414, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Rota, E.; Marchan, D.F.; Omodeo, P. Hormogaster regina sp. n. (Annelida: Clitellata: Hormogastridae): A giant earthworm from Spanish Catalonia, described from morphological and molecular evidence. Zool. Anz. A J. Comp. Zool. 2016, 261, 56–65. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernandez, R.; Sánchez, N.; De Sosa, I.; Cosín, D.J.D.; Novo, M. Insights into the diversity of Hormogastridae (Annelida, Oligochaeta) with descriptions of six new species. Zootaxa 2018, 4496, 65–95. [Google Scholar] [CrossRef]
- Marchan, D.F.; Novo, M.; Fernández, R.; De Sosa, I.; Trigo, D.; Cosín, D.J.D. Evaluating evolutionary pressures and phylogenetic signal in earthworms: A case study—The number of typhlosole lamellae in Hormogastridae (Annelida, Oligochaeta). Zool. J. Linn. Soc. 2016, 178, 4–14. [Google Scholar] [CrossRef]
- Marchán, D.F.; Sánchez, N.; Novo, M.; Fernández, R.; Pardos, F.; Cosín, D.J.D. Cryptic characters for cryptic taxa: On the taxonomic utility of the genital chaetae in earthworms (Oligochaeta, Hormogastridae). Zool. Anz. A J. Comp. Zool. 2016, 264, 17–28. [Google Scholar] [CrossRef]
- Marchán, D.F.; Fernández, R.; de Sosa, I.; Sánchez, N.; Cosín, D.J.D.; Novo, M. Integrative systematic revision of a Mediterranean earthworm family: Hormogastridae (Annelida, Oligochaeta). Invertebr. Syst. 2018, 32, 652–671. [Google Scholar] [CrossRef]
- Novo, M.; Fernández, R.; Fernandez Marchan, D.; Trigo, D.; Diaz Cosin, D.J.; Giribet, G. Unearthing the historical biogeography of Mediterranean earthworms (Annelida: Hormogastridae). J. Biogeogr. 2015, 42, 751–762. [Google Scholar] [CrossRef]
- Tilikj, N.; Novo, M. How to resist soil desiccation: Transcriptional changes in a Mediterranean earthworm during aestivation. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2021, 264, 111112. [Google Scholar] [CrossRef]
- Fernández, R.; Almodóvar, A.; Novo, M.; Gutiérrez, M.; Cosín, D.J.D. Earthworms, good indicators for palaeogeographical studies? Testing the genetic structure and demographic history in the peregrine earthworm Aporrectodea trapezoides (Dugès, 1828) in southern Europe. Soil Biol. Biochem. 2013, 58, 127–135. [Google Scholar] [CrossRef]
- Taheri, S.; Decaëns, T.; Cunha, L.; Brown, G.G.; Da Silva, E.; Bartz, M.; Baretta, D.; Dupont, L. Genetic evidence of multiple introductions and mixed reproductive strategy in the peregrine earthworm Pontoscolex corethrurus. Biol. Invasions 2020, 22, 2545–2557. [Google Scholar] [CrossRef]
- Aspe, N.M.; James, S.W. Molecular phylogeny and biogeographic distribution of pheretimoid earthworms (clitellata: Megascolecidae) of the Philippine archipelago. Eur. J. Soil Biol. 2018, 85, 89–97. [Google Scholar] [CrossRef]
- Jiang, J.; Qiu, J. Origin and evolution of earthworms belonging to the family Megascolecidae in China. Biodivers. Sci. 2018, 26, 1074–1082. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marchán, D.F.; Decaëns, T.; Domínguez, J.; Novo, M. Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions. Diversity 2022, 14, 30. https://doi.org/10.3390/d14010030
Marchán DF, Decaëns T, Domínguez J, Novo M. Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions. Diversity. 2022; 14(1):30. https://doi.org/10.3390/d14010030
Chicago/Turabian StyleMarchán, Daniel Fernández, Thibaud Decaëns, Jorge Domínguez, and Marta Novo. 2022. "Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions" Diversity 14, no. 1: 30. https://doi.org/10.3390/d14010030
APA StyleMarchán, D. F., Decaëns, T., Domínguez, J., & Novo, M. (2022). Perspectives in Earthworm Molecular Phylogeny: Recent Advances in Lumbricoidea and Standing Questions. Diversity, 14(1), 30. https://doi.org/10.3390/d14010030