
The Joint Contributions of Environmental Filtering and Spatial Processes to Macroinvertebrate Metacommunity Dynamics in the Alpine Stream Environment of Baima Snow Mountain, Southwest China

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
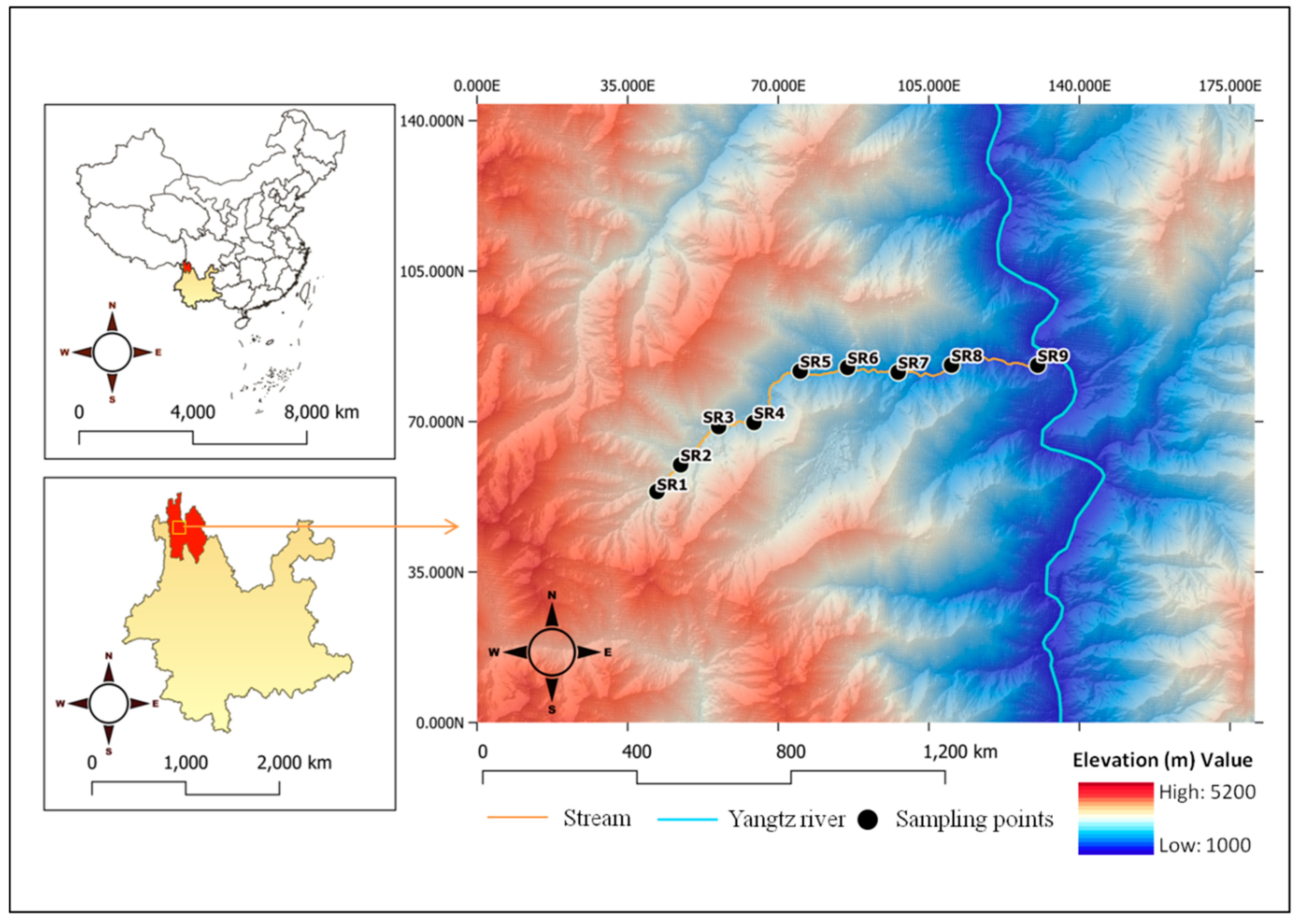
2.1. Study Area
2.2. Environmental Factors and Benthic Macroinvertebrates
2.3. Spatial Factors
2.4. Data Analysis
3. Results
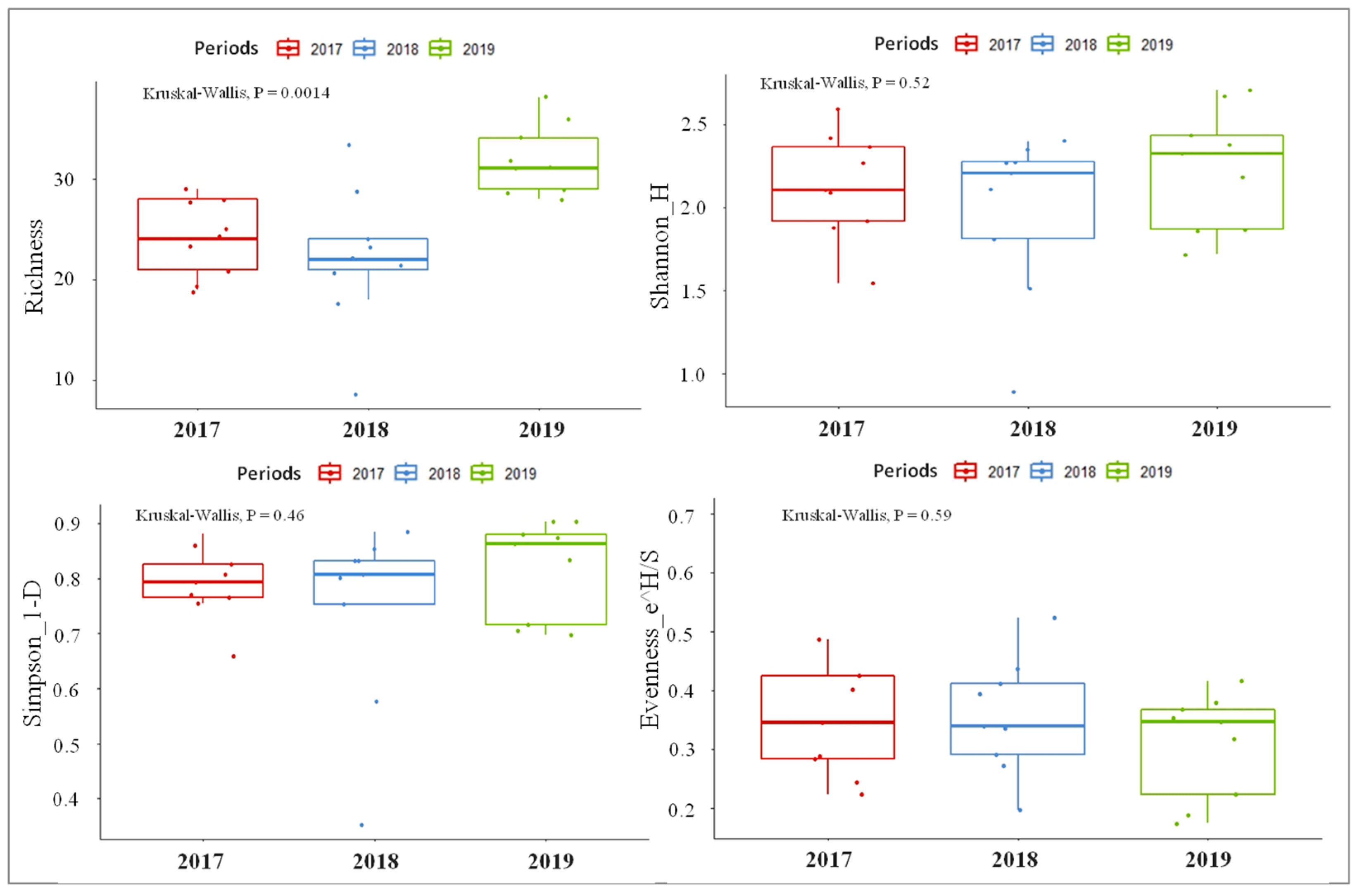
3.1. Environmental Conditions
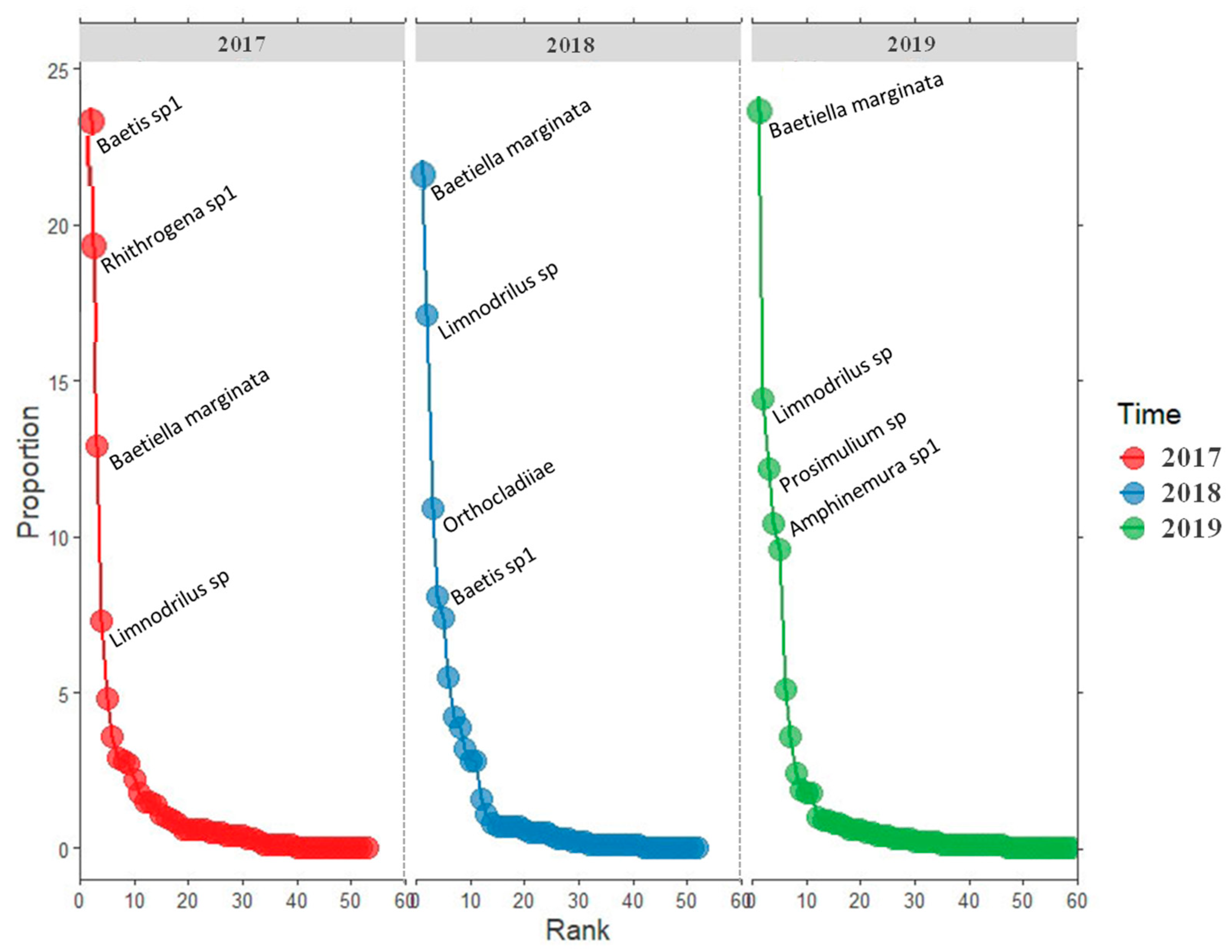
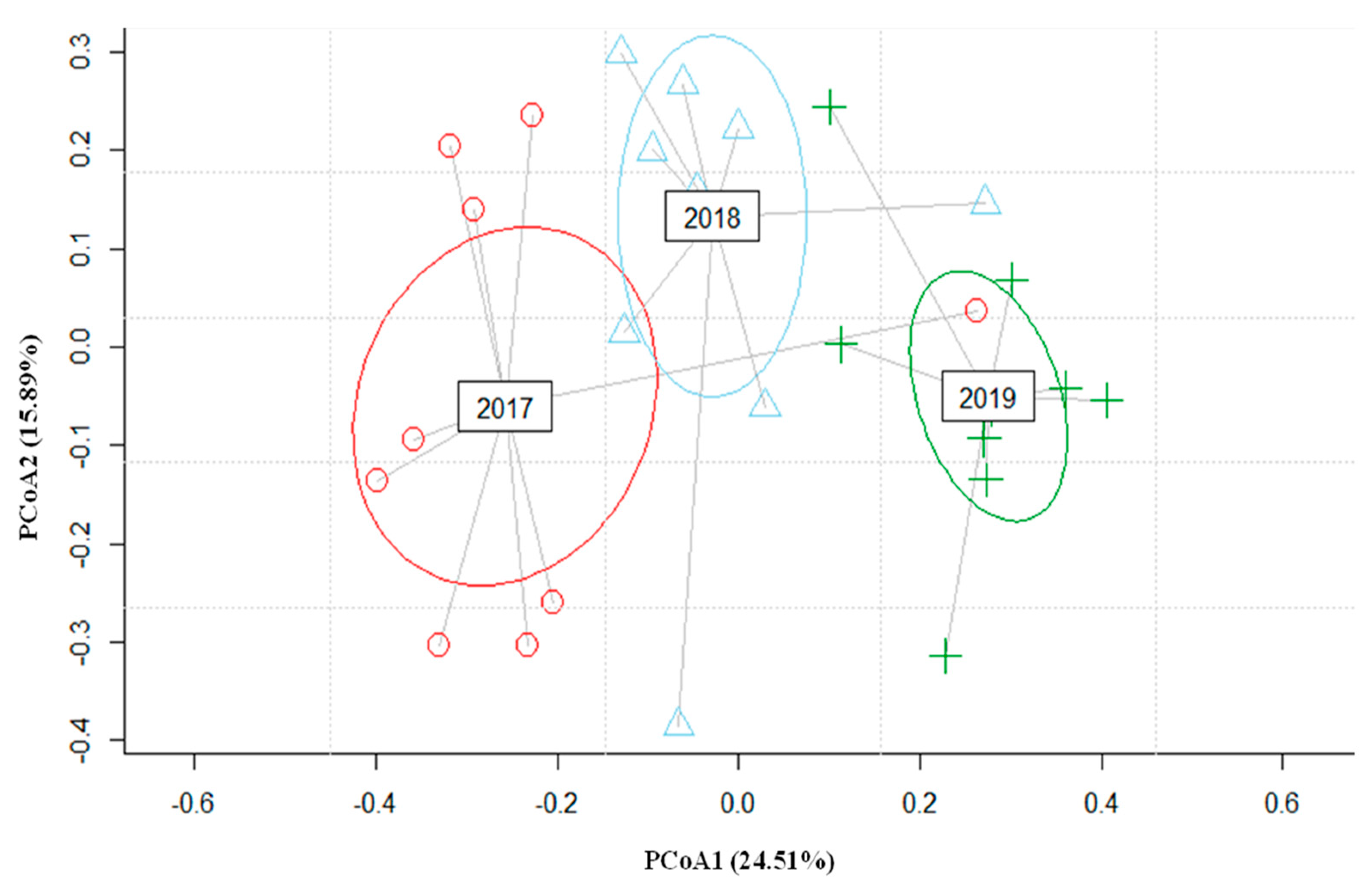
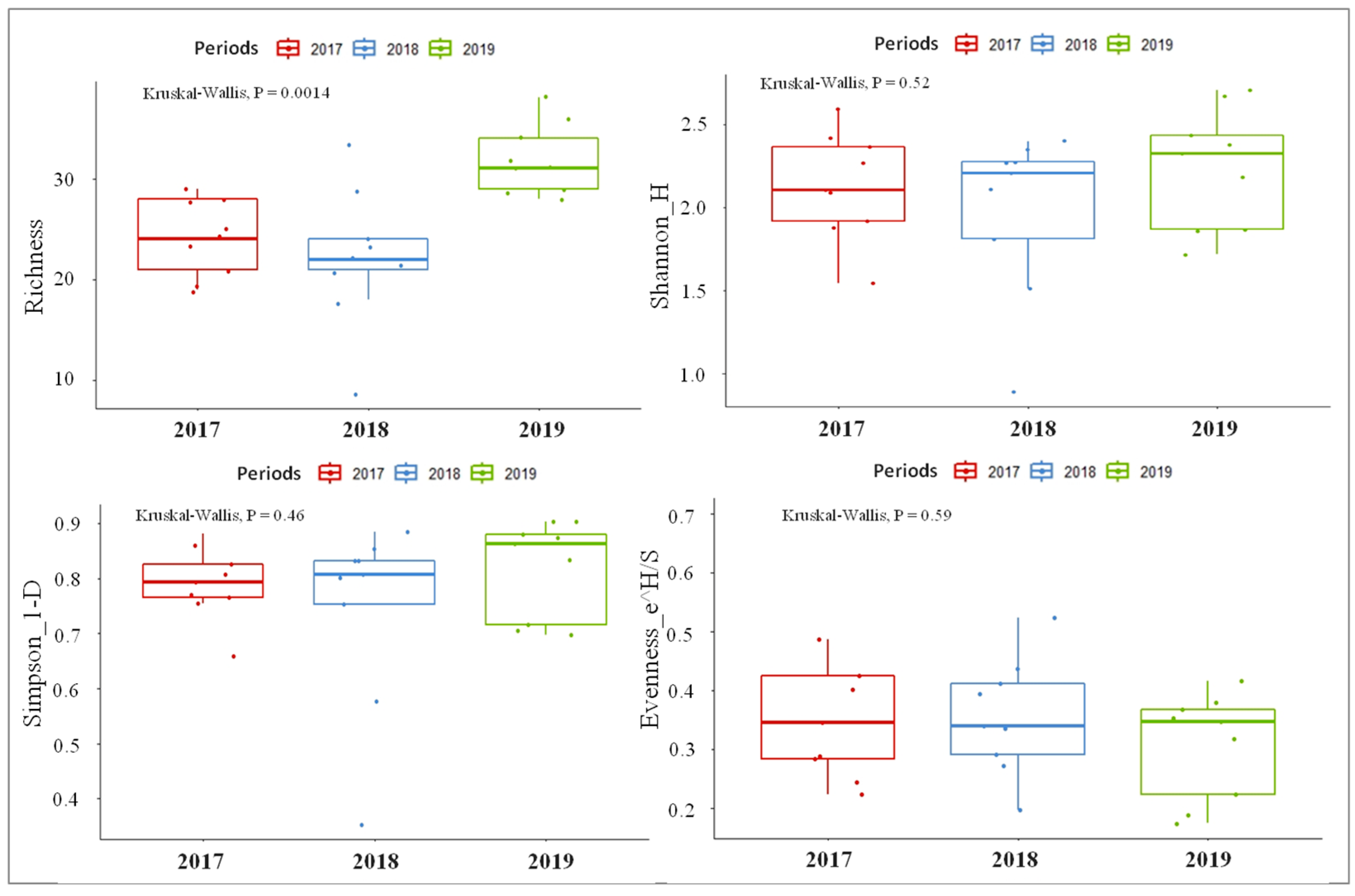
3.2. Macroinvertebrates Composition
3.3. Important Local Environmental Variables and Spatial Components
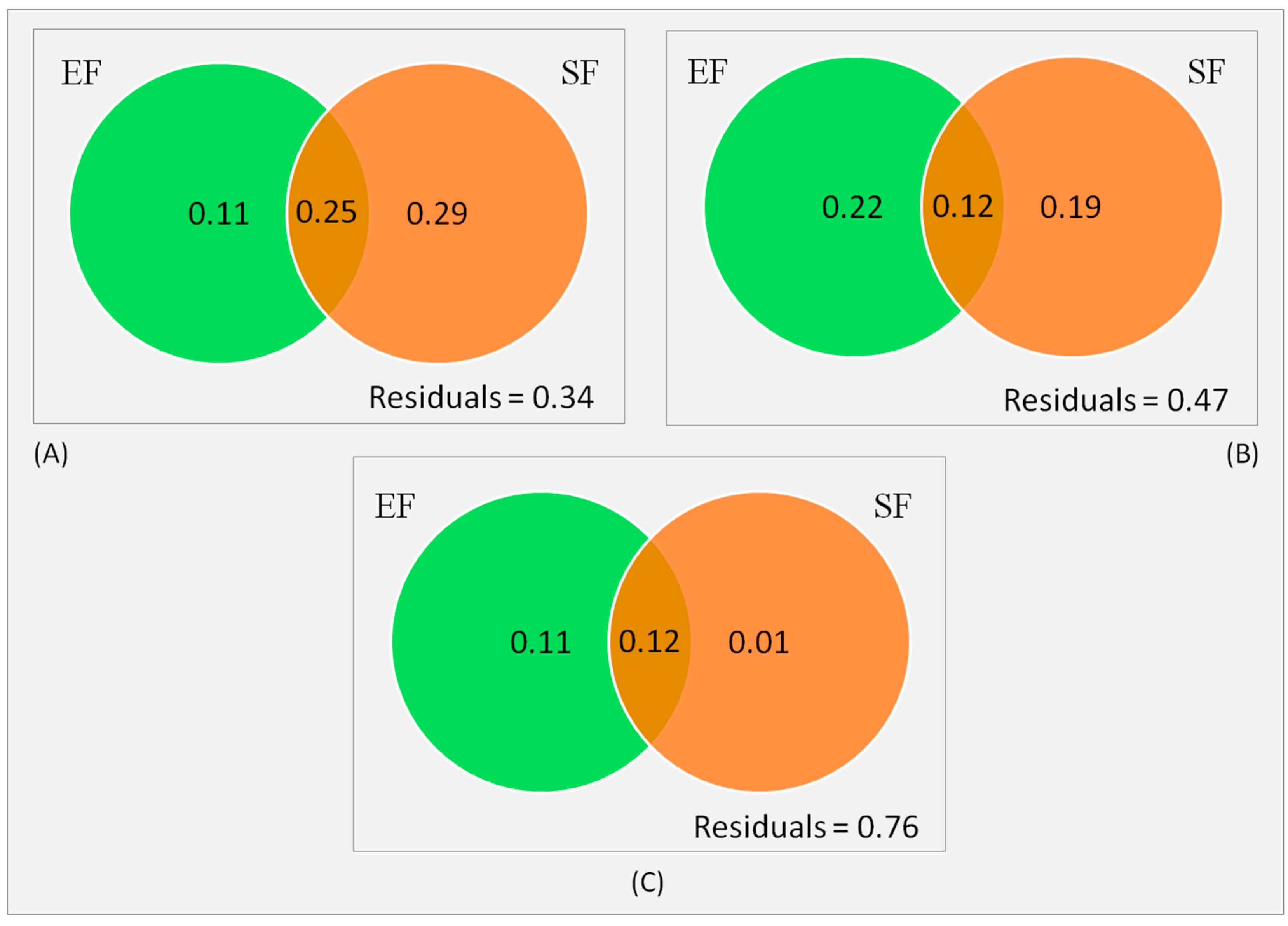
3.4. Relative Roles of Environmental and Spatial Components
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Tonkin, J.D.; Stoll, S.; Jähnig, S.C.; Haase, P. Elements of metacommunity structure of river and riparian assemblages: Communities, taxonomic groups and deconstructed trait groups. Ecol. Complex. 2016, 25, 35–43. [Google Scholar] [CrossRef]
- Keddy, P.A. Assembly and response rules: Two goals for predictive community ecology. J. Veg. Sci. 1992, 3, 157–164. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, J.C.; Malumbres-Olarte, J.; Arnedo, M.A.; Crespo, L.C.; Domenech, M.; Cardoso, P. Taxonomic divergence and functional convergence in Iberian spider forest communities: Insights from beta diversity partitioning. J. Biogeogr. 2020, 47, 288–300. [Google Scholar] [CrossRef]
- Hubbell, S.P. The unified neutral theory of biodiversity and biogeography: Reply. Ecology 2004, 85, 3175–3178. [Google Scholar] [CrossRef]
- Chase, J.M.; Jeliazkov, A.; Ladouceur, E.; Viana, D.S. Biodiversity conservation through the lens of metacommunity ecology. Ann. N. Y. Acad. Sci. 2020, 1469, 86–104. [Google Scholar] [CrossRef]
- Leibold, M.A.; Chase, J.M. Metacommunity Ecology; Princeton University Press: Priceton, NJ, USA, 2017. [Google Scholar]
- Logue, J.B.; Mouquet, N.; Peter, H.; Hillebrand, H.; The Metacommunity Working Group. Empirical approaches to metacommunities: A review and comparison with theory. Trends Ecol. Evol. 2011, 26, 482–491. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Ren, K.; Isabwe, A.; Chen, H.; Liu, M.; Yang, J. Stochastic processes shape microeukaryotic community assembly in a subtropical river across wet and dry seasons. Microbiome 2019, 7, 1–16. [Google Scholar]
- Gray, D.K.; Arnott, S.E. Does dispersal limitation impact the recovery of zooplankton communities damaged by a regional stressor? Ecol. Appl. 2011, 21, 1241–1256. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Legendre, P.; Soininen, J.; Yeh, C.F.; Graham, E.; Stegen, J.; Casamayor, E.O.; Zhou, J.; Shen, J.; Pan, F. Temperature drives local contributions to beta diversity in mountain streams: Stochastic and deterministic processes. Glob. Ecol. Biogeogr. 2020, 29, 420–432. [Google Scholar] [CrossRef]
- Jamoneau, A.; Passy, S.I.; Soininen, J.; Leboucher, T.; Tison-Rosebery, J. Beta diversity of diatom species and ecological guilds: Response to environmental and spatial mechanisms along the stream watercourse. Freshw. Biol. 2018, 63, 62–73. [Google Scholar] [CrossRef] [Green Version]
- Westveer, J.J.; van der Geest, H.G.; van Loon, E.E.; Verdonschot, P.F. Verdonschot, Connectivity and seasonality cause rapid taxonomic and functional trait succession within an invertebrate community after stream restoration. PLoS ONE 2018, 13, e0197182. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.M.; Foster, B.L.; Ballantyne IV, F.; Collins, C.D.; Antonovics, J.; Holt, R.D. Metapopulations and metacommunities: Combining spatial and temporal perspectives in plant ecology. J. Ecol. 2012, 100, 88–103. [Google Scholar] [CrossRef]
- Helsen, K.; Hermy, M.; Honnay, O. Spatial isolation slows down directional plant functional group assembly in restored semi-natural grasslands. J. Appl. Ecol. 2013, 50, 404–413. [Google Scholar] [CrossRef]
- Chang, C.; HilleRisLambers, J. Integrating succession and community assembly perspectives. F1000Research 2016, 5, 2294. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Xing, Y.; Liu, Z.; Chen, X.; Jiang, X.; Xie, Z.; Heino, J. Seasonal changes in metacommunity assembly mechanisms of benthic macroinvertebrates in a subtropical river basin. Sci. Total Environ. 2020, 729, 139046. [Google Scholar] [CrossRef] [PubMed]
- Beche, L.A.; Mcelravy, E.P.; Resh, V.H. Long-term seasonal variation in the biological traits of benthic-macroinvertebrates in two Mediterranean-climate streams in California, USA. Freshw. Biol. 2006, 51, 56–75. [Google Scholar] [CrossRef]
- Jiang, X.; Xiong, J.; Xie, Z. Longitudinal and seasonal patterns of macroinvertebrate communities in a large undammed river system in Southwest China. Quat. Int. 2017, 440, 1–12. [Google Scholar] [CrossRef]
- Bini, L.M.; Landeiro, V.L.; Padial, A.A.; Siqueira, T.; Heino, J. Nutrient enrichment is related to two facets of beta diversity for stream invertebrates across the United States. Ecology 2014, 95, 1569–1578. [Google Scholar] [CrossRef] [Green Version]
- Heino, J.; Melo, A.S.; Bini, L.M.; Altermatt, F.; Al-Shami, S.A.; Angeler, D.G.; Bonada, N.; Brand, C.; Callisto, M.; Cottenie, K.; et al. A comparative analysis reveals weak relationships between ecological factors and beta diversity of stream insect metacommunities at two spatial levels. Ecol. Evol. 2015, 5, 1235–1248. [Google Scholar] [CrossRef] [Green Version]
- Cilleros, K.; Allard, L.; Vigouroux, R.; Brosse, S. Disentangling spatial and environmental determinants of fish species richness and assemblage structure in Neotropical rainforest streams. Freshw. Biol. 2017, 62, 1707–1720. [Google Scholar] [CrossRef]
- Gálvez, Á.; Aguilar-Alberola, J.A.; Armengol, X.; Bonilla, F.; Iepure, S.; Monrós, J.S.; Olmo, C.; Rojo, C.; Rueda, J.; Rueda, R.; et al. Environment and space rule, but time also matters for the organization of tropical pond metacommunities. Front. Ecol. Evol. 2020, 8, 353. [Google Scholar] [CrossRef]
- Castillo-Escrivà, A.; Valls, L.; Rochera, C.; Camacho, A.; Mesquita-Joanes, F. Metacommunity dynamics of Ostracoda in temporary lakes: Overall strong niche effects except at the onset of the flooding period. Limnologica 2017, 62, 104–110. [Google Scholar] [CrossRef]
- Csercsa, A.; Krasznai-K, E.Á.; Várbíró, G.; Szivák, I.; Tóth, M.; Árva, D.; Bódis, E.; Deák, C.; Mauchart, P.; Móra, A.; et al. Seasonal changes in relative contribution of environmental control and spatial structuring on different dispersal groups of stream macroinvertebrates. Hydrobiologia 2019, 828, 101–115. [Google Scholar] [CrossRef] [Green Version]
- Sarremejane, R.; Cañedo-Argüelles, M.; Prat, N.; Mykrä, H.; Muotka, T.; Bonada, N. Do metacommunities vary through time? Intermittent rivers as model systems. J. Biogeogr. 2017, 44, 2752–2763. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.; Wu, Y.; Du, Y.; Xia, L.; Ge, D.; Yang, Q.; Chen, L. Seasonal change of species diversity patterns of non-volant small mammals along three subtropical elevational gradients. Biotropica 2014, 46, 479–488. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, Q.; Wen, Z.; Xia, L.; Zhang, Q.; Zhou, H. What drives the species richness patterns of non-volant small mammals along a subtropical elevational gradient? Ecography 2013, 36, 185–196. [Google Scholar] [CrossRef]
- Dudgeon, D. Tropical Asian Streams: Zoobenthos, Ecology and Conservation; Hong Kong University Press: Hongkong, China, 1999. [Google Scholar]
- Morse, J.C.; Yang, L.; Tian, L. Aquatic Insects of China Useful for Monitoring Water Quality; Hohai University Press: Nanjing, China, 1994. [Google Scholar]
- Jacobus, L.M. Revision of Ephemerellidae Genera (Ephemeroptera). Trans. Am. Entomol. Soc. 2008, 185, 185–274. [Google Scholar] [CrossRef]
- Jacobus, L.M. Two new species of Asian Serratella Edmunds (Ephemeroptera: Ephemerellidae). Zootaxa 2009, 58, 52–58. [Google Scholar] [CrossRef]
- Shi, W.; Tong, X. Taxonomic notes on the genus Baetiella Uéno from China, with the descriptions of three new species (Ephemeroptera: Baetidae). Zootaxa 2015, 4012, 553–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heino, J.; Alahuhta, J.; Ala-Hulkko, T.; Antikainen, H.; Bini, L.M.; Bonada, N.; Datry, T.; Erős, T.; Hjort, J.; Kotavaara, O.; et al. Integrating dispersal proxies in ecological and environmental research in the freshwater realm. Environ. Rev. 2017, 25, 334–349. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Gilbert, B.; Bennett, J.R. Partitioning variation in ecological communities: Do the numbers add up? J. Appl. Ecol. 2010, 47, 1071–1082. [Google Scholar] [CrossRef]
- Dray, S.; Legendre, P. Testing the species traits–environment relationships: The fourth-corner problem revisited. Ecology 2008, 89, 3400–3412. [Google Scholar] [CrossRef]
- Legendre, P.; Borcard, D.; Blanchet, F.G.; Dray, S. PCNM: MEM Spatial Eigenfunction and Principal Coordinate Analyses. R Package Version 2.1-2/r109. 2013. Available online: https://r-forge.r-project.org/projects/sedar/ (accessed on 17 August 2021).
- Beck, M.W. ggord: Ordination Plots with ggplot2. R Package Version 1.0.0. 2017. Available online: https://zenodo.org/badge/latestdoi/35334615 (accessed on 17 August 2021).
- Dray, S.; Blanchet, G.; Borcard, D.; Guenard, G.; Jombart, T.; Larocque, G.; Larocque, G.; Legendre, P.; Madi, N.; Wagner, H.H. adespatial: Multivariate Multiscale Spatial Analysis. R Package Version 0.0-9. 2017. Available online: https://CRAN.R-project.org/package=adespatial (accessed on 17 August 2021).
- Peres-Neto, P.R.; Legendre, P.; Dray, S.; Borcard, D. Variation partitioning of species data matrices: Estimation and comparison of fractions. Ecology 2006, 87, 2614–2625. [Google Scholar] [CrossRef]
- R Core Team. R Foundation for Statistical Computing, Vienna. Available online: www.r-project.org (accessed on 17 August 2021).
- Heino, J.; Grönroos, M.; Soininen, J.; Virtanen, R.; Muotka, T. Context dependency and metacommunity structuring in boreal headwater streams. Oikos 2012, 121, 537–544. [Google Scholar] [CrossRef]
- Kärnä, O.-M.; Grönroos, M.; Antikainen, H.; Hjort, J.; Ilmonen, J.; Paasivirta, L.; Heino, J. Data from: Inferring the effects of potential dispersal routes on the metacommunity structure of stream insects: As the crow flies, as the fish swims or as the fox runs? J. Anim. Ecol. 2015, 84, 1342–1353. [Google Scholar] [CrossRef] [PubMed]
- Reinhardt, K.; Köhler, G.; Maas, S.; Detzel, P. Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: The Orthoptera of Germany. Ecography 2005, 28, 593–602. [Google Scholar] [CrossRef]
- Tsang, T.P.N.; Bonebrake, T.C. Contrasting roles of environmental and spatial processes for common and rare urban butterfly species compositions. Landsc. Ecol. 2017, 32, 47–57. [Google Scholar] [CrossRef]
- Vidal, T.; Santos, J.I.; Marques, C.R.; Pereira, J.L.; Claro, M.T.; Pereira, R.; Castro, B.B.; Soares, A.; Gonçalves, F. Resilience of the macroinvertebrate community of a small mountain river (Mau River, Portugal) subject to multiple stresses. Mar. Freshw. Res. 2014, 65, 633–644. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Peng, C.; Huang, S.; Wang, J.; Xiong, X.; Li, D. The relative role of spatial and environmental processes on seasonal variations of phytoplankton beta diversity along different anthropogenic disturbances of subtropical rivers in China. Environ. Sci. Pollut. Res. 2019, 26, 1422–1434. [Google Scholar] [CrossRef]
- Vilmi, A.; Karjalainen, S.M.; Hellsten, S.; Heino, J. Bioassessment in a metacommunity context: Are diatom communities structured solely by species sorting? Ecol. Indic. 2016, 62, 86–94. [Google Scholar] [CrossRef]
- Heino, J.; Mykrä, H. Control of stream insect assemblages: Roles of spatial configuration and local environmental factors. Ecol. Entomol. 2008, 33, 614–622. [Google Scholar] [CrossRef]
- Vanschoenwinkel, B.; De Vries, C.; Seaman, M.; Brendonck, L. The role of metacommunity processes in shaping inverte-brate rock pool communities along a dispersal gradient. Oikos 2007, 116, 1255–1266. [Google Scholar] [CrossRef]
- Wang, J.; Hu, J.; Tang, T.; Heino, J.; Jiang, X.; Li, Z.; Xie, Z. Seasonal shifts in the assembly dynamics of benthic macroinvertebrate and diatom communities in a subtropical river. Ecol. Evol. 2020, 10, 692–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanschoenwinkel, B.; Waterkeyn, A.; Jocque, M.; Boven, L.; Seaman, M.; Brendonck, L. Species sorting in space and time—the impact of disturbance regime on community assembly in a temporary pool metacommunity. J. North Am. Benthol. Soc. 2010, 29, 1267–1278. [Google Scholar] [CrossRef]
- Cottenie, K. Integrating environmental and spatial processes in ecological community dynamics. Ecol. Lett. 2005, 8, 1175–1182. [Google Scholar] [CrossRef] [PubMed]
- Grönroos, M.; Heino, J.; Siqueira, T.; Landeiro, V.L.; Kotanen, J.; Bini, L.M. Metacommunity structuring in stream networks: Roles of dispersal mode, distance type, and regional environmental context. Ecol. Evol. 2013, 3, 4473–4487. [Google Scholar] [CrossRef] [PubMed]
- Raitif, J.; Plantegenest, M.; Agator, O.; Piscart, C.; Roussel, J.-M. Seasonal and spatial variations of stream insect emergence in an intensive agricultural landscape. Sci. Total Environ. 2018, 644, 594–601. [Google Scholar] [CrossRef]
- Datry, T.; Bonada, N.; Heino, J. Towards understanding the organisation of metacommunities in highly dynamic ecological systems. Oikos 2016, 125, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Zhou, T.; Cui, Y.; Li, Z.; Wang, W.; Chen, Y.; Xie, Z. Environmental filtering and spatial processes equally contributed to macroinvertebrate metacommunity dynamics in the highly urbanized river networks in Shenzhen, South China. Ecol. Process. 2021, 10, 1–12. [Google Scholar] [CrossRef]
- Cid, N.; Bonada, N.; Heino, J.; Cañedo-Argüelles, M.; Crabot, J.; Sarremejane, R.; Soininen, J.; Stubbington, R.; Datry, T. A Metacommunity Approach to Improve Biological Assessments in Highly Dynamic Freshwater Ecosystems. Biosciences 2020, 70, 427–438. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2017 | 2018 | 2019 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | |
| ALT(m) ns | 2417 | 3769 | 3041 | 2200 | 3800 | 3000 | 2158 | 3773 | 3005 |
| COND (µs/cm) ns | 172 | 286.4 | 246.54 | 159.7 | 259 | 226.94 | 176 | 298.2 | 252.72 |
| Sal (ppt) ns | 0.12 | 0.19 | 0.16 | 0.12 | 0.18 | 0.15 | 0.14 | 0.2 | 0.17 |
| TDS (mg/L) ns | 167.7 | 250.9 | 217.89 | 163.15 | 239.85 | 214.35 | 193.7 | 265.2 | 240.35 |
| WT (°C) ns | 7.5 | 13.7 | 10.88 | 6 | 13 | 8.63 | 3.6 | 13.4 | 8.26 |
| DO (mg/L) *** | 6.94 | 7.87 | 7.4 | 7.26 | 8.12 | 7.59 | 7.86 | 8.81 | 8.22 |
| pH *** | 8.62 | 8.96 | 8.86 | 8.26 | 8.56 | 8.48 | 8.02 | 8.31 | 8.18 |
| ORP (mV) *** | 80.6 | 138.1 | 110.96 | 125.3 | 236.9 | 195.13 | 25.1 | 63.1 | 46.88 |
| 2017 | 2018 | 2019 | ||||
|---|---|---|---|---|---|---|
| F-Value | p-Value | F-Value | p-Value | F-Value | p-Value | |
| ALT(m) | 3.712 | >0.05 | 2.001 | >0.05 | 2.800 | >0.05 |
| COND (µs/cm) | 2.224 | 0.04 | 2.172 | >0.05 | 2.038 | >0.05 |
| Sal (ppt) | 1.489 | >0.05 | 2.201 | 0.03 | 1.692 | >0.05 |
| TDS (mg/L) | 1.590 | >0.05 | 2.262 | 0.02 | 1.692 | >0.05 |
| WT (°C) | 3.584 | 0.05 | 1.635 | >0.05 | 2.643 | 0.01 |
| DO (mg/L) | 0.663 | >0.05 | 1.701 | 0.09 | 0.755 | >0.05 |
| pH | 1.443 | >0.05 | 2.427 | 0.01 | 2.549 | <0.01 |
| ORP (mV) | 4.128 | 0.01 | 1.315 | >0.05 | 2.927 | 0.01 |
| PCNM1 | 8.489 | <0.01 | 2.326 | 0.02 | 3.145 | 0.03 |
| PCNM2 | 3.957 | 0.01 | 2.282 | 0.02 | 3.108 | 0.01 |
| PCNM3 | 3.273 | 0.02 | 2.282 | >0.05 | 1.883 | >0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farooq, M.; Li, X.; Li, Z.; Yang, R.; Tian, Z.; Tan, L.; Fornacca, D.; Li, Y.; Cili, N.; Ciren, Z.; et al. The Joint Contributions of Environmental Filtering and Spatial Processes to Macroinvertebrate Metacommunity Dynamics in the Alpine Stream Environment of Baima Snow Mountain, Southwest China. Diversity 2022, 14, 28. https://doi.org/10.3390/d14010028
Farooq M, Li X, Li Z, Yang R, Tian Z, Tan L, Fornacca D, Li Y, Cili N, Ciren Z, et al. The Joint Contributions of Environmental Filtering and Spatial Processes to Macroinvertebrate Metacommunity Dynamics in the Alpine Stream Environment of Baima Snow Mountain, Southwest China. Diversity. 2022; 14(1):28. https://doi.org/10.3390/d14010028
Chicago/Turabian StyleFarooq, Muhammad, Xianfu Li, Zhengfei Li, Ronglong Yang, Zhen Tian, Lu Tan, Davide Fornacca, Yanpeng Li, Nima Cili, Zhongyong Ciren, and et al. 2022. "The Joint Contributions of Environmental Filtering and Spatial Processes to Macroinvertebrate Metacommunity Dynamics in the Alpine Stream Environment of Baima Snow Mountain, Southwest China" Diversity 14, no. 1: 28. https://doi.org/10.3390/d14010028
APA StyleFarooq, M., Li, X., Li, Z., Yang, R., Tian, Z., Tan, L., Fornacca, D., Li, Y., Cili, N., Ciren, Z., Liu, S., & Xiao, W. (2022). The Joint Contributions of Environmental Filtering and Spatial Processes to Macroinvertebrate Metacommunity Dynamics in the Alpine Stream Environment of Baima Snow Mountain, Southwest China. Diversity, 14(1), 28. https://doi.org/10.3390/d14010028