
Bermuda’s Walsingham Caves: A Global Hotspot for Anchialine Stygobionts



Abstract

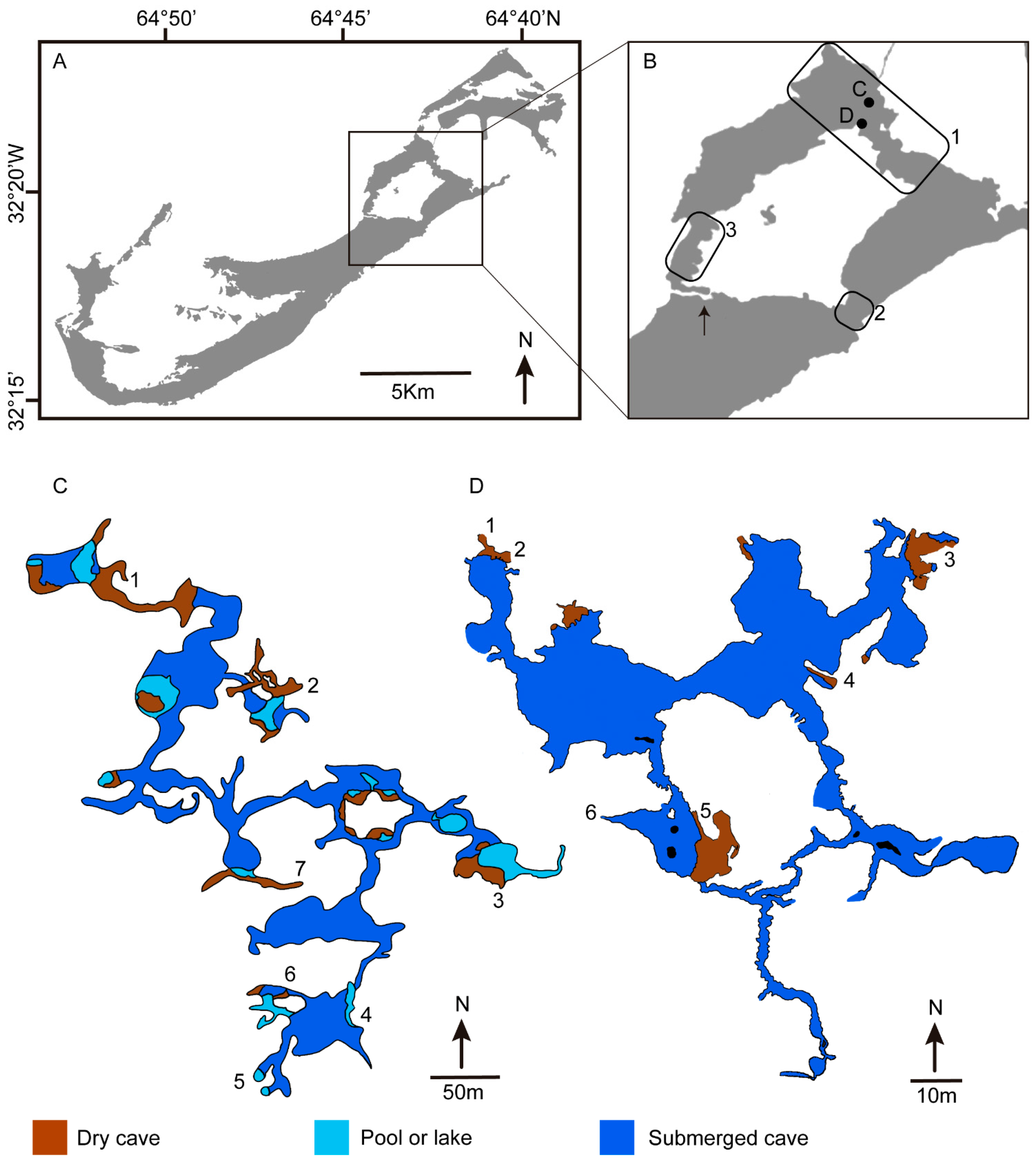
1. Introduction
- “The water flowing to them [Harrington Sound] underground,
- Being most salt, and all along the shore
- There are dark caves, of a miles length or more
- Extending under ground, in which there be
- Deep holes with water, though no one can see
- A passage for it in…”
2. Anchialine
3. Bermuda Geography and Geology
4. Bermuda Caves
5. Cave Biology
6. Threats to Bermuda’s Cave Fauna
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smithsonian.com. Bermuda—History and Heritage. Available online: https://www.smithsonianmag.com/travel/bermuda-history-and-heritage-14340790/?no-ist (accessed on 8 June 2021).
- Iliffe, T.M. Speleological history of Bermuda. Acta Carsologica 1993, 22, 114–135. [Google Scholar]
- Lefroy, L.H. Memorials of the Discovery and Early Settlement of the Bermudas or Somers Islands; Franklin Classics Trade Press: London, UK, 1877; pp. 1511–1687. [Google Scholar]
- Thomson, C.W. The Voyage of the “Challenger”. The Atlantic; A Preliminary Account of the General Results of the Exploring Voyage of H.M.S. “Challenger” during the Year 1873 and the Early Part of the Year 1876; Harper & Brothers: New York, NY, USA, 1878. [Google Scholar]
- Iliffe, T.M.; Hart, C.W.; Manning, R.B. Biogeography and the caves of Bermuda. Nature 1983, 302, 141–142. [Google Scholar] [CrossRef]
- Holthuis, L.B. Caridean shrimps found in Land-locked saltwater pools at four Indo-West Pacific localities (Sinai Peninsula, Funafuti Atoll, Maui and Hawaii Islands), with the description of one new genus and four new species. Zool. Verh. 1973, 128, 3–48. [Google Scholar]
- Stock, J.H.; Iliffe, T.M.; Williams, D. The concept of “anchialine” reconsidered. Stygologia 1986, 2, 90–92. [Google Scholar]
- Bishop, R.E.; Humphreys, W.F.; Cukrov, N.; Žic, V.; Boxshall, G.A.; Cukrov, M.; Iliffe, T.M.; Kršinić, F.; Moore, W.S.; Pohlman, J.W.; et al. ‘Anchialine’ redefined as a subterranean estuary in a crevicular or cavernous geological setting. J. Crustac. Biol. 2015, 35, 511–514. [Google Scholar] [CrossRef]
- van Hengstum, P.J.; Cresswell, J.N.; Milne, G.A.; Iliffe, T.M. Development of anchialine cave habitats and karst subterranean estuaries since the last ice age. Sci. Rep. 2019, 9, 11907. [Google Scholar] [CrossRef] [PubMed]
- Rowe, M.P. The Geology of Bermuda; bermudageology.com: Bermuda, UK, 2020. [Google Scholar]
- Palmer, A.N.; Palmer, M.V.; Queen, J.M. Geology and origin of the caves of Bermuda. In Proceedings of the Seventh International Congress of Speleology, Sheffield, England, 11–16 September 1977; pp. 336–339. [Google Scholar]
- Illife, T.M. The submarine caves of Bermuda. In Proceedings of the Eighth International Congress of Speleology, Bowling Green, KY, USA, 18–24 July 1981; pp. 161–163. [Google Scholar]
- Thomas, M.L.H. The Ecology of Harrington Sound, Bermuda; Bermuda Zoological Society: Bermuda, UK, 2005. [Google Scholar]
- Iliffe, T.M. Anchialine cave ecology. In Ecosystems of the World. 30. Subterranean Ecosystems; Wilkens, H., Culver, D.C., Humphreys, W.F., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; pp. 59–76. [Google Scholar]
- Pohlman, J.W.; Cifuentes, L.A.; Iliffe, T.M. Food Web Dynamics and Biogeochemistry of Anchialine Caves: A Stable Isotope Approach. Ecosyst. World 2000, 30, 351–363. [Google Scholar]
- Sket, B.; Iliffe, T.M. Cave fauna of Bermuda. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1980, 65, 871–882. [Google Scholar] [CrossRef]
- Culver, D.C.; Sket, B. Hotspots of subterranean biodiversity in caves and wells. J. Cave Karst Stud. 2000, 62, 11–17. [Google Scholar]
- Culver, D.C.; Pipan, T. The Biology of Caves and Other Subterranean Habitats; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Hart, C.W.J.; Manning, R.B. The cavernicolous caridean shrimps of Bermuda (Alpheidae, Hippolytidae, and Atyidae). J. Crustac. Biol. 1981, 1, 441–456. [Google Scholar] [CrossRef]
- Anker, A.; Iliffe, T.M. Description of Bermudacaris harti, a new genus, and species (Crustacea: Decapoda: Alpheidae) from anchialine caves of Bermuda. Proc. Biol. Soc. Wash. 2000, 113, 761–775. [Google Scholar]
- Bowman, T.E.; Iliffe, T.M. Mictocaris halope, a new unusual peracaridan crustacean from marine caves on Bermuda. J. Crustac. Biol. 1985, 5, 58–73. [Google Scholar] [CrossRef]
- von Martens, E. Über Cubanische Crustaceen. Arch. Naturgeschich. 1872, 38, 77–147. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Available online: http://www.iucnredlist.org/ (accessed on 30 May 2021).
- WoRMS. Editorial Board World Register of Marine Species. Available online: http://www.marinespecies.org (accessed on 30 May 2021).
- Hill, B.F.; Small, E.B.; Iliffe, T.M. Euplotes iliffei n. sp.: A new species of Euplotes (Ciliophora, Hypotrichida) from the marine caves of Bermuda. J. Wash. Acad. Sci. 1986, 76, 244–249. [Google Scholar]
- Small, E.B.; Heisler, J.; Sniezek, J.; Iliffe, T.M. Glauconema bermudense n. sp. (Scuticociliatida, Oligohymenophorea), a troglobitic ciliophoran from Bermudian marine caves. Stygologia 1986, 2, 167–179. [Google Scholar]
- Sterrer, W.; Iliffe, T.M. Mesonerilla prospera, a new archiannelid from marine caves in Bermuda. Proc. Biol. Soc. Wash. 1982, 95, 509–514. [Google Scholar]
- Worsaae, K.; Sterrer, W.; Iliffe, T.M. Longipalpa saltatrix, a new genus and species of the meiofaunal family Nerillidae (Annelida: Polychaeta) from an anchihaline cave in Bermuda. Proc. Biol. Soc. Wash. 2004, 117, 346–362. [Google Scholar]
- Erséus, C. A new species of Phallodrilus (Oligochaeta, Tubificidae) from a limestone cave on Bermuda. Sarsia 1986, 71, 7–9. [Google Scholar] [CrossRef]
- Moolenbeek, R.; Faber, M.; Iliffe, T.M. Two new species of the genus Caecum (Gastropoda) from marine caves on Bermuda. Stud. Honour Pieter Wagenaar Hummelinck 1988, 123, 209–216. [Google Scholar]
- Bartsch, I.; Iliffe, T.M. The halacarid fauna (Halacaridae, Acari) of Bermuda’s cave. Stygologia 1985, 1, 300–321. [Google Scholar]
- Fosshagen, A.; Boxshall, G.A.; Iliffe, T.M. The Epacteriscidae, a cave-living family of calanoid copepods. Sarsia 2001, 86, 245–318. [Google Scholar] [CrossRef]
- Fosshagen, A. A new genus and species of bottom living calanoid (Copepoda) from Florida and Colombia. Sarsia 1973, 52, 145–154. [Google Scholar] [CrossRef]
- Fosshagen, A.; Iliffe, T.M. Two new genera of Calanoida and a new order of Copepoda, Platycopioida, from marine caves on Bermuda. Sarsia 1985, 70, 345–358. [Google Scholar] [CrossRef]
- Fosshagen, A. Marine Biological Investigations in the Bahamas. 15. Ridgewayia (Copepoda, Calanoida) and two new genera of calanoids from the Bahamas. Sarsia 1970, 44, 25–58. [Google Scholar] [CrossRef]
- Yeatman, H.C. Miostephos leamingtonensis, a new species of copepod from Bermuda. J. Tenn. Acad. Sci. 1980, 55, 21–22. [Google Scholar]
- Esterly, C.O. Calanoid Copepoda from the Bermuda Islands. Proc. Am. Acad. Arts Sci. 1911, 47, 219. [Google Scholar] [CrossRef]
- Rocha, C.E.F.; Iliffe, T.M. New cyclopoids (Copepoda) from anchialine caves in Bermuda. Sarsia 1993, 78, 43–56. [Google Scholar] [CrossRef]
- Lotufo, G.R.; da Rocha, C.E.F. Intertidal interstitial Halicyclops from the Brazilian coast (Copepoda: Cyclopoida). Hydrobiologia 1993, 264, 175–184. [Google Scholar] [CrossRef]
- Huys, R. Superornatiremidae fam. nov. (Copepoda: Harpacticoida): An enigmatic family from North Atlantic anchihaline caves. Sci. Mar. 1996, 60, 497–542. [Google Scholar]
- Boxshall, G.A.; Iliffe, T.M. New cave-dwelling misophrioids (Crustacea: Copepoda) from Bermuda. Sarsia 1986, 71, 55–64. [Google Scholar] [CrossRef]
- Boxshall, G.A.; Iliffe, T.M. Three new species of misophrioid copepods from oceanic islands. J. Nat. Hist. 1990, 24, 595–613. [Google Scholar] [CrossRef]
- Fosshagen, A.; Iliffe, T.M. A new genus of Platycopioida (Copepoda) from a marine cave on Bermuda. In Biology of Copepods; Boxshall, G.A., Schminke, H.K., Eds.; Springer: Dordrecht, The Netherlands, 1988; pp. 357–361. [Google Scholar]
- Maddocks, R.F.; Iliffe, T.M. Podocopid Ostracoda of Bermudian caves. Stygologia 1986, 2, 26–76. [Google Scholar]
- Skogsberg, T. Studies on marine ostracods. Part 1. (Cypridinids, Halocyprids and Polycopids). Zool. Bidr. Fran Upps. 1920, 1–784. [Google Scholar] [CrossRef]
- Angel, M.V.; Iliffe, T.M. Spelaeoecia bermudensis, new genus, new species, a halocyprid ostracod from marine caves in Bermuda. J. Crustac. Biol. 1987, 7, 541–553. [Google Scholar] [CrossRef]
- Kornicker, L.S.; Iliffe, T.M. Ostracoda (Myodocopina, Cladocopina, Halocypridina) from anchialine caves in Bermuda. Smithson. Contrib. Zool. 1989, 475, 1–88. [Google Scholar] [CrossRef]
- Maddocks, R.F. Revision of the family Pontocyprididae (Ostracoda), with new anchialine species and genera from Galapagos Islands. Zool. J. Linn. Soc. 1991, 103, 309–333. [Google Scholar] [CrossRef]
- Hart, C.W.J.; Manning, R.B. Two new shrimps (Procarididae and Agostocarididae, new family) from marine caves of the western North Atlantic. J. Crustac. Biol. 1986, 6, 408–416. [Google Scholar] [CrossRef]
- Sket, B. Atlantasellus cavernicolus n. gen., n. sp. (Isopoda Asellota, Atlantasellidae n. fam.) from Bermuda. Biol. Vestn. Ljubl. 1979, 7, 175–183. [Google Scholar]
- Bowman, T.E.; Iliffe, T.M. Bermudalana aruboides, a new genus and specioes of troglobitic isopoda (Cirolanidae) from marine caves on Bermuda. Proc. Biol. Soc. Wash. 1983, 96, 291–300. [Google Scholar]
- Wägele, J.W.; Brandt, A. New west Atlantic localities for the stygobiont paranthurid Curassanthura (Crustacea, Isopoda, Anthuridea) with description of C. bermudensis n. sp. Bijdr. Dierkd. 1985, 55, 324–330. [Google Scholar] [CrossRef]
- Kensley, B. Records of shallow-water marine isopods from Bermuda with descriptions of four new species. J. Crustac. Biol. 1994, 14, 319–336. [Google Scholar] [CrossRef]
- Karaman, G.S. Revision of the genus Gitanopsis Sars 1895 with description of new genera Afrogitanopsis and Rostrogitanopsis n. gen. (fam. Amphilochidae). Poljopr. Sumar. 1980, 26, 43–69. [Google Scholar]
- Stock, J.H.; Sket, B.; Iliffe, T.M. Two new amphipod crustaceans from anchihaline caves in Bermuda. Crustaceana 1987, 53, 54–66. [Google Scholar] [CrossRef]
- Karaman, G.S. Revision of genus Idunella Sars with description of new species, I. sketi, n. sp. (Fam. Liljeborgiidae). Acta Adriat. 1980, 21, 409–435. [Google Scholar]
- Karaman, G.S. Cocoharpinia iliffei, new genus and species from Bermuda, with remarks to other genera and species (Fam. Phoxocephalidae). (Contribution to the Knowledge of the Amphipoda 103). Stud. Mar. 1980, 9–10, 149–175. [Google Scholar]
- Barnard, J.L.; Clark, J. A new sea-cave amphipod from Bermuda (Dulichiidae). Proc. Biol. Soc. Wash. 1985, 98, 1048–1053. [Google Scholar]
- Stock, J.H.; Holsinger, J.R.; Sket, B.; Iliffe, T.M. Two new species of Pseudoniphargus (Amphipoda), in Bermudian groundwaters. Zool. Scr. 1986, 15, 237–249. [Google Scholar] [CrossRef]
- Gutu, M.; Iliffe, T.M. Apseudes orghidani, a new species of Tanaidacea (Crustacea) from an anchialine cave on Bermuda. Trav. Mus. Natl. d’Hist. Nat. Grigore Antipa 1989, 30, 161–167. [Google Scholar]
- Băcescu, M. Apseudes bermudeus n.sp. from caves around Bermude [sic] Islands [Apseudes bermudeus n.sp. iz pecina oko Bermudskih Otoka]. Acta Adriat. 1980, 21, 401–407. [Google Scholar]
- Bäcescu, M. Deux espèces nouvelles de Cumella (Crustacea, Cumacea) des grottes sous-marines de Bermuda. Trav. Mus. Natl. d’Hist. Nat. Grigore Antipa 1992, 32, 257–262. [Google Scholar]
- Băcescu, M.; Iliffe, T.M. Nouvelles espèces de Cumella des grottes sous-marines de Bermude. Rev. Behav. Almyracuma Prox. 1991, 36, 9–13. [Google Scholar]
- Băcescu, M. New Cumacea from the littoral waters of Florida (Caribbean Sea). Trav. Mus. Natl. d’Hist. Nat. Grigore Antipa 1971, 11, 5–24. [Google Scholar]
- Petrescu, I.; Sterrer, W. Cumacea (Crustacea) from shallow waters of Bermuda. Ann. Nat. Naturhist. Mus. Wien Ser. B Bot. Zool. 2001, 103, 89–128. [Google Scholar]
- Băcescu, M.; Iliffe, T.M. Bermudamysis g.n., Platyops g.n. and other mysids from Bermudian caves. Stygologia 1986, 2, 93–104. [Google Scholar]
- Ditter, R.E.; Mejía-Ortíz, L.M.; Bracken-Grissom, H.D. Anchialine adjustments: An updated phylogeny and classification for the family Barbouriidae Christoffersen, 1987 (Decapoda: Caridea). J. Crustac. Biol. 2020, 40, 401–411. [Google Scholar] [CrossRef]
- Keyser, D.; Schöning, C. Holocene ostracoda (Crustacea) from Bermuda. Senckenberg. Lethaea 2000, 80, 567–591. [Google Scholar] [CrossRef]
- Martín, A.; Díaz Díaz, Y.J. Biodiversidad de crustáceos peracáridos en el delta del Río Orinoco, Venezuela. Rev. Biol. Trop. 2006, 55. [Google Scholar] [CrossRef]
- Stock, J.H.; Vonk, R. Marine interstitial Amphipoda and Isopoda (Crustacea) from Santiago, Cape Verde Islands. Bijdr. Dierkd. 1992, 62, 21–36. [Google Scholar] [CrossRef]
- Winfield, I.; Abarca-Ávila, M.; Ortiz, M.; Cházaro-Olvera, S.; Lozano-Aburto, M.Á. Biodiversidad de los tanaidáceos (Crustacea: Peracarida: Tanaidacea) del Parque Nacional Arrecife Puerto Morelos, Quintana Roo, México. Rev. Mex. Biodivers. 2017, 88, 572–578. [Google Scholar] [CrossRef]
- Suárez-Morales, E.; Cervantes-Martínez, A.; Gutiérrez-Aguirre, M.A.; Iliffe, T.M. A new Speleophria (Copepoda, Misophrioida) from an anchialine cave of the Yucatán Peninsula with comments on the biogeography of the genus. Bull. Mar. Sci. 2017, 93, 1–16. [Google Scholar] [CrossRef]
- Jurado-Rivera, J.A.; Pons, J.; Alvarez, F.; Botello, A.; Humphreys, W.F.; Page, T.J.; Iliffe, T.M.; Willassen, E.; Meland, K.; Juan, C.; et al. Phylogenetic evidence that both ancient vicariance and dispersal have contributed to the biogeographic patterns of anchialine cave shrimps. Sci. Rep. 2017, 7, 2852. [Google Scholar] [CrossRef]
- Stokkan, M.; Jurado-Rivera, J.A.; Oromí, P.; Juan, C.; Jaume, D.; Pons, J. Species delimitation and mitogenome phylogenetics in the subterranean genus Pseudoniphargus (Crustacea: Amphipoda). Mol. Phylogenet. Evol. 2018, 127, 988–999. [Google Scholar] [CrossRef]
- Kornicker, L.S.; Iliffe, T.M.; Harrison-Nelson, E. Ostracoda (Myodocopa) from Anchialine Caves and Ocean Blue Holes. Zootaxa 2007, 1565, 1–151. [Google Scholar] [CrossRef]
- Koenemann, S.; Iliffe, T.M. Class Remipedia Yager, 1981. In Treatise on Zoology—Anatomy, Taxonomy, Biology: The Crustacea; von Vaupel Klein, J.C., Charmantier-Daures, M., Schram, F.R., Eds.; Brill: Leiden, The Netherlands, 2013; pp. 125–177. [Google Scholar]
- Ballou, L.; Iliffe, T.M.; Kakuk, B.; Gonzalez, B.C.; Osborn, K.J.; Worsaae, K.; Meland, K.; Broad, K.; Bracken-Grissom, H.; Olesen, J. Monsters in the dark: Systematics and biogeography of the stygobitic genus Godzillius (Crustacea: Remipedia) from the Lucayan Archipelago. Eur. J. Taxon. 2021, 751, 115–139. [Google Scholar] [CrossRef]
- Moldovan, O.T.; Kováč, L.; Halse, S. Ecological Studies, In Cave Ecology; Springer: Cham, Germany, 2018. [Google Scholar]
- Martínez, A.; Gonzalez, B.C.; Núñez, J.; Wilkens, H.; Oromí, P.; Iliffe, T.M.; Worsaae, K. Guide to the Anchialine Ecosystems of Jameos del Agua and Túnel de la Atlántida; Cabildo de Lanzarote: Lanzorote, Spain, 2016. [Google Scholar]
- Iliffe, T.M. Bermuda’s caves: A non-renewable resource. Environ. Conserv. 1979, 6, 181–186. [Google Scholar] [CrossRef]
- Mammola, S.; Cardoso, P.; Culver, D.C.; Deharveng, L.; Ferreira, R.L.; Fišer, C.; Galassi, D.M.P.; Griebler, C.; Halse, S.; Humphreys, W.F.; et al. Scientists’ warning on the conservation of subterranean ecosystems. Bioscience 2019, 69, 641–650. [Google Scholar] [CrossRef]
- Worldometers.info. Bermuda Population (LIVE). Available online: https://www.worldometers.info/world-population/bermuda-population/ (accessed on 30 June 2021).
- Iliffe, T.M.; Jickells, T.D.; Brewer, M.S. Organic pollution of an inland marine cave from Bermuda. Mar. Environ. Res. 1984, 12, 173–189. [Google Scholar] [CrossRef]
- Parravicini, V.; Guidetti, P.; Morri, C.; Montefalcone, M.; Donato, M.; Bianchi, C.N. Consequences of sea water temperature anomalies on a Mediterranean submarine cave ecosystem. Estuar. Coast. Shelf Sci. 2010, 86, 276–282. [Google Scholar] [CrossRef]
- Glasspool, A. Management Plan for Bermuda’s Critically Endangered Cave Fauna; Government of Bermuda, Department of Conservation Services: Bermuda, UK, 2014.
- Darrell, M. Letter to Bermuda Department of Planning, re: Proposed Demolition of Unstable Mound with Subterranean Void in Wilkinson Quarry, Hamilton Parish. Hamilton: Bermuda. 10 June 2020. Available online: https://www.bnt.bm/images/Education/Wilkinson Quarry Cave Objection.pdf (accessed on 28 June 2021).
- Copeland, A. Walsingham Trust Nature Reserve: Special Features and Potential Projects; Museum and Zoo Report Number BAMZ#3405; Bermuda Aquarium: Bermuda, UK, 2020. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ecology | Species Habitat | # Bermuda Caves | Monospecific Taxa | Species in the Genus | Habitat Relationship | Region Relationship | Conservation Status | Reference | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cave/Interstitial | Deep Sea | Caribbean | Atlantic | Other | ||||||||
| Chromista | ||||||||||||
| Ciliophora, Euplotidae | ||||||||||||
| Euplotes iliffei Hill & Small in Hill, Small & Iliffe, 1986 | B,e | W | 1 | 50 | X | X | X | [25] | ||||
| Ciliophora, Parauronematidae | ||||||||||||
| Glauconema bermudense Small, 1986 | B,e | W | 4 | 2 | X | [26] | ||||||
| Annelida | ||||||||||||
| Polychaeta, Nerillidae | ||||||||||||
| Leptonerilla prospera (Sterrer & Iliffe, 1982) | B,e | S | 8 | 3 | 2 | X | X | Cr | [27] | |||
| Speleonerilla saltatrix (Worsaae, Sterrer & Iliffe, 2004) | B,e | S | 1 | 4 | 3 | X | X | [28] | ||||
| Oligochaeta, Tubificidae | ||||||||||||
| Phallodriloides macmasterae (Erséus, 1986) | B,e | S | 1 | 5 | 2 | X | Cr | [29] | ||||
| Mollusca | ||||||||||||
| Gastropoda, Caecidae | ||||||||||||
| Caecum caverna Moolenbeek, Faber & Iliffe, 1988 | B,e | S | 2 | 218 | 1 | X | X | X | [30] | |||
| Caecum troglodyta Moolenbeek, Faber & Iliffe, 1988 | B,e | S | 2 | 218 | 1 | X | X | X | [30] | |||
| Arthropoda | ||||||||||||
| Arachnida | ||||||||||||
| Acari, Halacaridae | ||||||||||||
| Agauopsis bermudensis Bartsch & Iliffe, 1985 | B,e | S | 2 | 90 | 1 | X | X | X | [31] | |||
| Agauopsis littoralis Bartsch & Iliffe, 1985 | B,e | S | 1 | 90 | 1 | X | X | X | [31] | |||
| Copidognathus bermudensis Bartsch & Iliffe, 1985 | B,e | S | 2 | 380 | 2 | X | X | X | [31] | |||
| Copidognathus longispinus Bartsch & Iliffe, 1985 | B,e | S | 1 | 380 | 2 | X | X | X | [31] | |||
| Copidognathus subterraneus Bartsch & Iliffe, 1985 | B,e | S | 1 | 380 | 2 | X | X | X | [31] | |||
| Crustacea | ||||||||||||
| Copepoda, Calanoida | ||||||||||||
| Enantiosis bermudensis Fosshagen, Boxshall & Iliffe, 2001 | B,e | W | 10 | 7 | 6 | X | X | [32] | ||||
| Epacteriscus rapax Fosshagen, 1973 | P | W | 9 | 3 | 2 | X | X | [33] | ||||
| Erebonectes nesioticus Fosshagen, in Fosshagen & Iliffe, 1985 | B,e | W | 4 | G | 1 | 29 | 1 | X | X | Cr | [34] | |
| Exumella polyarthra Fosshagen, 1970 | P | W | X | 4 | 2 | X | X | [35] | ||||
| Miostephos leamingtonensis Yeatman, 1980 | B,e | W | 1 | 2 | 1 | X | [36] | |||||
| Paracyclopia naessi Fosshagen in Fosshagen & Iliffe, 1985 | B,e | W | 5 | G | 1 | 4 | Cr | [34] | ||||
| Ridgewayia marki (Esterly, 1911) | P | W | 2 | 14 | 4 | X | X | X | [37] | |||
| Copepoda, Cyclopoida | ||||||||||||
| Halicyclops bowmani Rocha & Iliffe, 1993 | B,e | W | 2 | 89 | 21 | X | X | X | [38] | |||
| Halicyclops herbsti Rocha & Iliffe, 1993 | B,e | W | 1 | 89 | 21 | X | X | X | ||||
| Halicyclops ytororoma Lotufo & Rocha, 1993 | P | W | 2 | 89 | 21 | X | X | X | [39] | |||
| Speleoithona bermudensis Rocha & Iliffe, 1993 | B,e | W | 2 | 3 | 2 | X | Cr | [38] | ||||
| Copepoda, Harpacticoida | ||||||||||||
| Intercrusia problematica Huys, 1996 | B,e | W | 1 | 2 | 1 | X | [40] | |||||
| Neoechinophora fosshageni Huys, 1996 | B,e | W | 2 | 5 | 4 | X | X | X | [40] | |||
| Neoechinophora daltonae Huys, 1996 | B,e | W | 3 | 5 | 4 | X | X | X | [40] | |||
| Neoechinophora jaumei Huys, 1996 | B,e | W | 2 | 5 | 4 | X | X | X | [40] | |||
| Superornatiremis mysticus Huys, 1996 | B,e | W | 3 | 2 | 1 | X | [40] | |||||
| Copepoda, Misophrioida | ||||||||||||
| Speleophria bivexilla Boxshall & Iliffe, 1986 | B,e | W | 1 | 6 | 5 | X | X | X | Cr | [41] | ||
| Speleophriopsis scottodicarloi (Boxshall & Iliffe, 1990) | B,e | W | 2 | 5 | 4 | X | X | Cr | [42] | |||
| Copepoda, Platycopioida | ||||||||||||
| Antrisocopia prehensilis Fosshagen in Fosshagen & Iliffe, 1985 | B,e | W | 1 | G | 1 | 1 | X | X | Cr | [34] | ||
| Nanocopia minuta Fosshagen in Fosshagen & Iliffe, 1988 | B,e | W | 1 | G | 1 | 1 | X | X | Cr | [43] | ||
| Ostracoda, Bairdiidae | ||||||||||||
| Aponesidea iliffei Maddocks in Maddocks & Iliffe, 1986 | B,e | S | 10 | 4 | X | [44] | ||||||
| Havanardia keiji Maddocks in Maddocks & Iliffe, 1986 | P,e | S | 9 | 12 | X | X | [44] | |||||
| Neonesidea omnivaga Maddocks in Maddocks & Iliffe, 1986 | P,e | S | 3 | 99 | X | X | X | [44] | ||||
| Paranesidea sterreri Maddocks in Maddocks & Iliffe, 1986 | P,e | S | 2 | 41 | X | X | X | [44] | ||||
| Ostracoda, Candonidae | ||||||||||||
| Dolerocypria bifurca Maddocks in Maddocks & Iliffe, 1986 | B,e | S | 3 | 14 | 2 | X | [44] | |||||
| Paracypris crispa Maddocks in Maddocks & Iliffe, 1986 | P,e | S | 3 | 39 | X | X | X | [44] | ||||
| Ostracoda, Cytherellidae | ||||||||||||
| Cytherella bermudensis Maddocks in Maddocks & Iliffe, 1986 | P,e | S | 2 | 158 | + | X | X | X | [44] | |||
| Cytherella kornickeri Maddocks in Maddocks & Iliffe, 1986 | B,e | S | 6 | 158 | + | X | X | X | [44] | |||
| Ostracoda, Cylindroleberididae | ||||||||||||
| Parasterope muelleri (Skogsberg, 1920) | P | 2 | 46 | X | X | [45] | ||||||
| Ostracoda, Deeveyidae | ||||||||||||
| Spelaeoecia bermudensis Angel & Iliffe, 1987 | B,e | W | 11 | 11 | 10 | X | Cr | [46] | ||||
| Ostracoda, Halocypridae | ||||||||||||
| Metapolycope duplex Kornicker & Iliffe, 1989 | B,e | S | 12 | 10 | 4 | X | [47] | |||||
| Micropolycope eurax Kornicker & Iliffe, 1989 | B,e | S | 3 | 11 | 1 | X | X | [47] | ||||
| Micropolycope styx Kornicker & Iliffe, 1989 | B,e | S | 6 | 11 | 1 | X | X | [47] | ||||
| Ostracoda, Philomedidae | ||||||||||||
| Pseudophilomedes kylix Kornicker & Iliffe, 1989 | B,e | S | 5 | 11 | X | X | [47] | |||||
| Ostracoda, Polycopidae | ||||||||||||
| Polycopissa anax Kornicker & Iliffe, 1989 | B,e | S | 8 | 4 | X | X | [47] | |||||
| Ostracoda, Pontocyprididae | ||||||||||||
| Iliffeoecia iliffei Maddocks, 1991 | B | S | 2 | 3 | X | [48] | ||||||
| Kareloecia minacis (Maddocks in Maddocks & Iliffe, 1986) | B,e | S | 1 | 2 | 1 | X | [44] | |||||
| Thomontocypris lurida (Maddocks in Maddocks & Iliffe, 1986) | B,e | S | 5 | 9 | 6 | X | X | X | [44] | |||
| Ostracoda, Sarsiellidae | ||||||||||||
| Eusarsiella styx Kornicker & Iliffe, 1989 | B,e | S | 6 | 86 | 7 | + | X | X | X | [47] | ||
| Caridea, Atyidae | ||||||||||||
| Typhlatya iliffei Hart & Manning, 1981 | B,e | W | 1 | 17 | 16 | X | X | X | Cr | [19] | ||
| Caridea, Alpheidae | ||||||||||||
| Bermudacaris harti Anker & Iliffe, 2000 | B,e | W | 4 | 3 | X | [20] | ||||||
| Caridea, Barbouriidae | ||||||||||||
| Barbouria cubensis (von Martens, 1872) | B | W | 3 | G | 1 | 8 | X | X | X | Cr | [22] | |
| Parhippolyte sterreri (Hart & Manning, 1981) | B | W | 2 | 6 | 5 | X | X | X | Cr | [19] | ||
| Caridea, Procarididae | ||||||||||||
| Procaris chacei Hart & Manning, 1986 | B,e | W | 1 | 5 | 4 | X | X | X | Cr | [49] | ||
| Isopoda, Atlantasellidae | ||||||||||||
| Atlantasellus cavernicolus Sket, 1979 | B,e | S | 1 | 2 | 1 | 1 | X | Cr | [50] | |||
| Isopoda, Cirolanidae | ||||||||||||
| Arubolana aruboides (Bowman & Iliffe, 1983) | B,e | W | 4 | 4 | 3 | X | X | Cr | [51] | |||
| Isopoda, Leptanthuridae | ||||||||||||
| Curassanthura bermudensis Wägele & Brandt, 1985 | B,e | S | 1 | 5 | 4 | X | X | Cr | [52] | |||
| Isopoda, Stenetriidae | ||||||||||||
| Stenobermuda iliffei Kensley, 1994 | B,e | S | 1 | 6 | X | X | X | [53] | ||||
| Amphipoda, Amphilochidae | ||||||||||||
| Hourstonius petulans (Karaman, 1980) | P | S | 1 | 18 | 1 | X | [54] | |||||
| Amphipoda, Bogidiellidae | ||||||||||||
| Bermudagidiella bermudiensis (Stock, Sket & Iliffe, 1987) | B,e | S | 2 | G | 1 | 115 | 114 | X | X | X | Cr | [55] |
| Amphipoda, Liljeborgiidae | ||||||||||||
| Idunella sketi Karaman, 1980 | P | S | 1 | 44 | 1 | X | X | X | Cr | [56] | ||
| Idunella verrilli Yabut & Sawicki, 2015 | B,e | 1 | 44 | 1 | X | X | X | [56] | ||||
| Amphipoda, Phoxocephalidae | ||||||||||||
| Cocoharpinia iliffei Karaman, 1980 | B,e | 1 | G | 1 | + | X | X | X | Cr | [57] | ||
| Amphipoda, Podoceridae | ||||||||||||
| Podobothrus bermudensis Barnard & Clark, 1985 | B,e | 1 | G | 1 | + | X | X | X | [58] | |||
| Amphipoda, Pseudoniphargidae | ||||||||||||
| Pseudoniphargus grandimanus Stock, Holsinger, Sket & Iliffe, 1986 | B,e | W | 20 | 72 | 71 | X | X | Cr | [59] | |||
| Ingolfiellida, Ingolfiellidae | ||||||||||||
| Ingolfiella (Tethydiella) longipes Stock, Sket & Iliffe, 1987 | B,e | S | 1 | 44 | + | + | X | X | X | Cr | [55] | |
| Tanaidacea, Apseudidae | ||||||||||||
| Apseudes orghidani Gutu & Iliffe, 1989 | B,e | W | 1 | 44 | + | X | X | X | [60] | |||
| Paradoxapseudes bermudeus (Băcescu, 1980) | P | S | 1 | 18 | 3 | 1 | X | X | [61] | |||
| Mictacea, Mictocarididae | ||||||||||||
| Mictocaris halope Bowman & Iliffe, 1985 | B,e | W | 4 | O | 1 | 3 | 3 | X | X | Cr | [21] | |
| Cumacea, Nannastacidae | ||||||||||||
| Cumella (Cumella) iliffei Băcescu, 1992 | B,e | S | 2 | 102 | 15 | X | X | X | [62] | |||
| Cumella (Cumella) ocellata Băcescu, 1992 | B,e | S | 1 | 102 | 15 | X | X | X | [62] | |||
| Cumella (Cumella) spinosa Băcescu & Iliffe, 1991 | S | 1 | 102 | 15 | X | X | X | [63] | ||||
| Schizotrema agglutinanta (Băcescu, 1971) | P | S | 8 | 20 | 1 | X | X | X | [64] | |||
| Schizotrema wittmanni Petrescu & Sterrer, 2001 | P | S | 1 | 20 | 1 | X | X | X | [65] | |||
| Mysida, Mysidae | ||||||||||||
| Bermudamysis speluncola Băcescu & Iliffe, 1986 | B,e | W | 2 | G | 1 | 2 | 1 | X | X | Cr | [66] | |
| Platyops sterreri Băcescu & Iliffe, 1986 | B,e | W | 2 | G | 1 | 2 | 1 | X | X | Cr | [66] | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iliffe, T.M.; Calderón-Gutiérrez, F. Bermuda’s Walsingham Caves: A Global Hotspot for Anchialine Stygobionts. Diversity 2021, 13, 352. https://doi.org/10.3390/d13080352
Iliffe TM, Calderón-Gutiérrez F. Bermuda’s Walsingham Caves: A Global Hotspot for Anchialine Stygobionts. Diversity. 2021; 13(8):352. https://doi.org/10.3390/d13080352
Chicago/Turabian StyleIliffe, Thomas M., and Fernando Calderón-Gutiérrez. 2021. "Bermuda’s Walsingham Caves: A Global Hotspot for Anchialine Stygobionts" Diversity 13, no. 8: 352. https://doi.org/10.3390/d13080352
APA StyleIliffe, T. M., & Calderón-Gutiérrez, F. (2021). Bermuda’s Walsingham Caves: A Global Hotspot for Anchialine Stygobionts. Diversity, 13(8), 352. https://doi.org/10.3390/d13080352