
Eciton Army Ants—Umbrella Species for Conservation in Neotropical Forests

Abstract
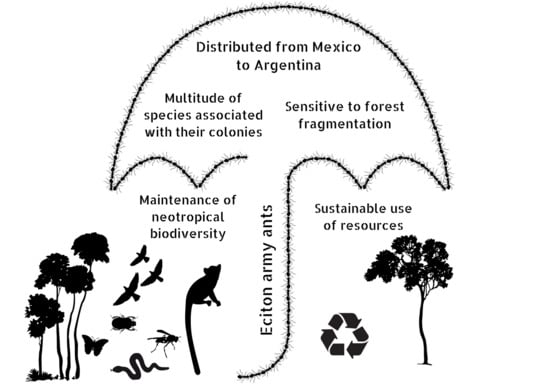
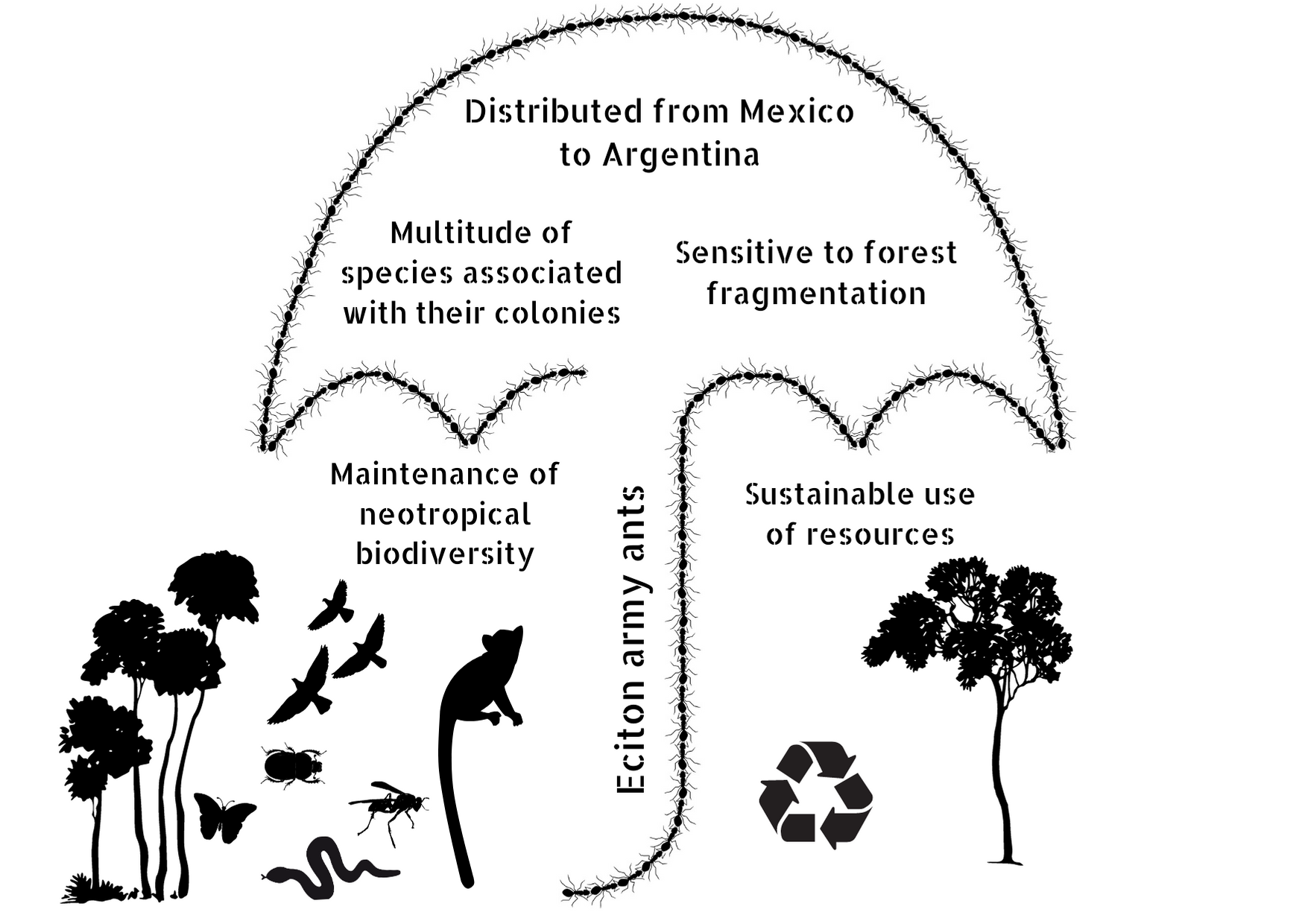{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Eciton Army Ants
3. Associates of Eciton Army Ants
3.1. Birds
3.2. Other Vertebrates
3.3. Mites
3.4. Flies
3.5. Beetles
3.6. Other Arthropods
3.7. Microbes
3.8. Further Research on Eciton Associates
4. Deforestation and Eciton Army Ants
5. Eciton Army Ants as Umbrella Species in Neo-Tropical Forests
Supplementary Materials
Funding
Acknowledgments
Conflicts of Interest
References
- Sala, O.E.; Chapin, F.S.; Armesto, J.J.; Berlow, E.; Bloomfield, J.; Dirzo, R.; Huber-Sanwald, E.; Huenneke, L.F.; Jackson, R.B.; Kinzig, A.; et al. Global biodiversity scenarios for the year 2100. Science 2000, 287, 1770–1774. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of Habitat Fragmentation on Biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Fischer, J.; Lindenmayer, D.B. Landscape modification and habitat fragmentation: A synthesis. Glob. Ecol. Biogeogr. 2007, 16, 265–280. [Google Scholar] [CrossRef]
- de Chazal, J.; Rounsevell, M.D.A. Land-use and climate change within assessments of biodiversity change: A review. Glob. Environ. Chang. 2009, 19, 306–315. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Mooney, H.A. Ecosystem Function of Biodiversity: A Summary; Schulze, E.-D., Mooney, H.A., Eds.; Springer: Berlin, Germany; London, UK, 1994. [Google Scholar]
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; MacE, G.M.; Tilman, D.; Wardle, D.A.; et al. Biodiversity loss and its impact on humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Mitchell, M.G.E.; Bennett, E.M.; González, A. Strong and nonlinear effects of fragmentation on ecosystem service provision at multiple scales. Environ. Res. Lett. 2015, 10. [Google Scholar] [CrossRef]
- Keenan, R.J.; Reams, G.A.; Achard, F.; de Freitas, J.V.; Grainger, A.; Lindquist, E. Dynamics of global forest area: Results from the FAO Global Forest Resources Assessment 2015. For. Ecol. Manage. 2015, 352, 9–20. [Google Scholar] [CrossRef]
- Tyukavina, A.; Hansen, M.C.; Potapov, P.V.; Stehman, S.V.; Smith-Rodriguez, K.; Okpa, C.; Aguilar, R. Types and rates of forest disturbance in Brazilian Legal Amazon, 2000–2013. Sci. Adv. 2017, 3, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dossa, G.G.O.; Paudel, E.; Schaefer, D.; Zhang, J.L.; Cao, K.F.; Xu, J.C.; Harrison, R.D. Quantifying the factors affecting wood decomposition across a tropical forest disturbance gradient. For. Ecol. Manage. 2020, 468. [Google Scholar] [CrossRef]
- Alroy, J. Effects of habitat disturbance on tropical forest biodiversity. Proc. Natl. Acad. Sci. USA 2017, 114, 6056–6061. [Google Scholar] [CrossRef]
- CBD. Secretariat of the Convention on Biological Diversity (2020). In Global Biodiversity Outlook 5; CBD: Montreal, QC, USA; ISBN 9292255398.
- FAO; UNEP. The State of the World’s Forests 2020. Forests, Biodiversity and People; FAO and UNEP: Rome, Italy, 2020. [Google Scholar]
- Stork, N.E.; Srivastava, D.S.; Eggleton, P.; Hodda, M.; Lawson, G.; Leakey, R.R.B.; Watt, A.D. Consistency of effects of tropical-forest disturbance on species composition and richness relative to use of indicator taxa. Conserv. Biol. 2017, 31, 924–933. [Google Scholar] [CrossRef] [PubMed]
- Gardner, T.A.; Barlow, J.; Araujo, I.S.; Ávila-Pires, T.C.; Bonaldo, A.B.; Costa, J.E.; Esposito, M.C.; Ferreira, L.V.; Hawes, J.; Hernandez, M.I.M.; et al. The cost-effectiveness of biodiversity surveys in tropical forests. Ecol. Lett. 2008, 11, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Noss, R.F. Indicators for monitoring biodiversity: A hierarchical approach. Conserv. Biol. 1990, 4, 355–364. [Google Scholar] [CrossRef]
- Lawton, J.H.; Bignell, D.E.; Bolton, B.; Bloemers, G.F.; Eggleton, P.; Hammond, P.M.; Hodda, M.; Holt, R.D.; Larsen, T.B.; Mawdsley, N.A.; et al. Biodiversity inventories, indicator taxa and effects of habitat modification in tropical forest. Nature 1998, 391, 72–76. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Westgate, M.J. Are flagship, umbrella and keystone species useful surrogates to understand the consequences of landscape change? Curr. Landsc. Ecol. Reports 2020, 5, 76–84. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Margules, C.R.; Botkin, D.B. Indicators of biodiversity for ecologically sustainable forest management. Conserv. Biol. 2000, 14, 941–950. [Google Scholar] [CrossRef]
- Landres, P.B.; Verner, J.; Thomas, J.W. Ecological uses of vertebrate indicator species: A critique. Conserv. Biol. 1988, 2, 316–328. [Google Scholar] [CrossRef]
- McGeogh, M.A. The selection, testing and application of terrestrial insects as bioindicators. Biol. Rev. 1998, 73, 181–201. [Google Scholar] [CrossRef]
- Caro, T.M.; O’Doherty, G. On the use of surrogate species in conservation biology. Conserv. Biol. 1999, 13, 805–814. [Google Scholar] [CrossRef]
- Lewandowski, A.S.; Noss, R.F.; Parsons, D.R. The effectiveness of surrogate taxa for the representation of biodiversity. Conserv. Biol. 2010, 24, 1367–1377. [Google Scholar] [CrossRef] [PubMed]
- Didham, R.K.; Stork, N.E.; Davis, A.J. Insects in fragmented forests: A functional approach. Trends Ecol. Evol. 1996, 11, 255–260. [Google Scholar] [CrossRef]
- Rands, M.R.W.; Adams, W.M.; Bennun, L.; Butchart, S.H.M.; Clements, A.; Coomes, D.; Entwistle, A.; Hodge, I.; Kapos, V.; Scharlemann, J.P.W.; et al. Biodiversity conservation: Challenges beyond 2010. Science 2010, 329, 1298–1303. [Google Scholar] [CrossRef]
- Kremen, C.; Colwell, R.K.; Erwin, T.L.; Murphy, D.D.; Noss, R.F.; Sanjayan, M.A. Terrestrial arthropod assemblages: Their use in conservation planning. Conserv. Biol. 1993, 7, 796–808. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Basset, Y.; Benke, K.K.; Grimbacher, P.S.; Miller, S.E.; Novotný, V.; Samuelson, G.A.; Stork, N.E.; Weiblen, G.D.; Yen, J.D.L. Quantifying uncertainty in estimation of tropical arthropod species richness. Am. Nat. 2010, 176, 90–95. [Google Scholar] [CrossRef]
- Brown, K.S. Diversity, disturbance, and sustainable use of Neotropical forests: Insects as indicators for conservation monitoring. J. Insect Conserv. 1997, 1, 25–42. [Google Scholar] [CrossRef]
- Andersen, A.N. Using ants as bioindicators: Multiscales issues in ant community ecology. Conserv. Ecol. 1997, 1. [Google Scholar] [CrossRef]
- Andersen, A.N.; Hoffmann, B.D.; Müller, W.J.; Griffiths, A.D. Using ants as bioindicators in land management: Simplifying assessment of ant community responses. J. Appl. Ecol. 2002, 39, 8–17. [Google Scholar] [CrossRef]
- Majer, J.D.; Orabi, G.; Bisevac, L. Ants (Hymenoptera: Formicidae) pass the bioindicator scorecard. Myrmecol. News 2007, 10, 69–76. [Google Scholar]
- Bihn, J.H.; Gebauer, G.; Brandl, R. Loss of functional diversity of ant assemblages in secondary tropical forests. Ecology 2010, 91, 782–792. [Google Scholar] [CrossRef]
- Groc, S.; Delabie, J.H.C.; Fernandez, F.; Petitclerc, F.; Corbara, B.; Leponce, M.; Céréghino, R.; Dejean, A. Litter-dwelling ants as bioindicators to gauge the sustainability of small arboreal monocultures embedded in the Amazonian rainforest. Ecol. Indic. 2017, 82, 43–49. [Google Scholar] [CrossRef]
- Boswell, G.P.; Britton, N.F.; Franks, N.R. Habitat fragmentation, percolation theory and the conservation of a keystone species. Proc. R. Soc. B Biol. Sci. 1998, 265, 1921–1925. [Google Scholar] [CrossRef]
- Kaspari, M.; O’Donnell, S. High rates of army ant raids in the Neotropics and implications for ant colony and community structure. Evol. Ecol. Res. 2003, 5, 933–939. [Google Scholar]
- Schneirla, T. Army Ants: A Study in Social Organization; Topoff, H.R., Ed.; Freeman, WH: San Francisco, CA, USA, 1971. [Google Scholar]
- Rettenmeyer, C.W. Arthropods associated with neotropical army ants with a review of the behaviour of these ants (Arthropoda; Formicidae; Dorylinae). Ph.D. Thesis, The University of Kansas, Lawrence, KS, USA, 1961. [Google Scholar]
- Gotwald, W.H., Jr. Army Ants: The Biology of Social Predation; Cornell University Press: Ithaca, NY, USA, 1995. [Google Scholar]
- Partridge, L.W.; Britton, N.F.; Franks, N.R. Army ant population dynamics: The effects of habitat quality and reserve size on population size and time to extinction. Proc. R. Soc. B Biol. Sci. 1996, 263, 735–741. [Google Scholar] [CrossRef]
- Secretariat of the Convention on Biological Diversity. Assessment, Conservation and Sustainable Use of Forest Biodiversity; CBD Technical Series no. 3; SCBD: Montreal, QC, USA, 2001; 130p. [Google Scholar]
- Alonso, L.E.; Persaud, J.; Williams, A. Biodiversity Assessment Survey of the South Rupununi Savannah, Guyana. BAT Survey Report No 1.; WWF-Guianas, Guyana Office: Georgetown, Guyana, 2016. [Google Scholar]
- Kistner, D.H. The social insects’ bestiary. In Social Insects; Hermann, H.R., Ed.; Academic Press: London, UK, 1982; Volume 3, pp. 1–244. ISBN 0323148964, 9780323148962. [Google Scholar]
- Brady, S.G.; Fisher, B.L.; Schultz, T.R.; Ward, P.S. The rise of army ants and their relatives: Diversification of specialized predatory doryline ants. BMC Evol. Biol. 2014, 14, 1–14. [Google Scholar] [CrossRef]
- Borowiec, M.L. Convergent evolution of the army ant syndrome and congruence in big-data phylogenetics. Syst. Biol. 2019, 68, 642–656. [Google Scholar] [CrossRef] [PubMed]
- Gotwald, W.H., Jr. Army Ants; Academic Press, Inc.: London, UK, 1982. [Google Scholar]
- Brady, S.G. Evolution of the army ant syndrome: The origin and long-term evolutionary stasis of a complex of behavioral and reproductive adaptations. Proc. Natl. Acad. Sci. USA 2003, 100, 6575–6579. [Google Scholar] [CrossRef] [PubMed]
- Kronauer, D.J.C. Army Ants: Nature’s Ultimate Social Hunters; Harvard University Press: Cambridge, MA, USA, 2020; ISBN 9780674241558. [Google Scholar]
- Franks, N.R.; Hölldobler, B. Sexual competition during colony reproduction in army ants. Biol. J. Linn. Soc. 1987, 30, 229–243. [Google Scholar] [CrossRef]
- Watkins, J.F. The Identification and Distribution of New World Army Ants; Baylor University Press: Waco, TX, USA, 1976. [Google Scholar]
- Winston, M.E. Bridging Micro- and Macroevolution in Neotropical Army Ants; University of Chicago: Chicago, IL, USA, 2017. [Google Scholar]
- Pérez-Espona, S.; Goodall-Copestake, W.P.; Berghoff, S.M.; Edwards, K.J.; Franks, N.R. Army imposters: Diversification of army ant-mimicking beetles with their Eciton hosts. Insectes Soc. 2018, 65, 59–75. [Google Scholar] [CrossRef]
- Rettenmeyer, C.W. Behavioral studies of army ants. Univ. Kansas Sci. Bull. 1963, 44, 281–465. [Google Scholar]
- Powell, S.; Baker, B. The hidden big predators of the Neotropics: The behaviour, diet, and impact of New World army ants (Ecitoninae). Insetos Sociais Biol. Apl. 2008, 18–37. [Google Scholar]
- Schneirla, T.C. Raiding and other outstanding ohenomena in the behavior of army ants. Proc. Natl. Acad. Sci. USA 1934, 20, 316–321. [Google Scholar] [CrossRef]
- Hoenle, P.O.; Blüthgen, N.; Brückner, A.; Kronauer, D.J.C.; Fiala, B.; Donoso, D.A.; Smith, M.A.; Ospina Jara, B.; von Beeren, C. Species-level predation network uncovers high prey specificity in a Neotropical army ant community. Mol. Ecol. 2019, 28, 2423–2440. [Google Scholar] [CrossRef]
- Franks, N.R.; Fletcher, C.R. Spatial patterns in army ant foraging and migration: Eciton burchelli on Barro Colorado Island, Panama. Behav. Ecol. Sociobiol. 1983, 12, 261–270. [Google Scholar] [CrossRef]
- O’Donnell, S.; Jeanne, R.L. Notes on an army ant (Eciton burchelli) raid on a social wasp colony (Agelaia yepocapa) in Costa Rica. J. Trop. Ecol. 1990, 6, 507–509. [Google Scholar] [CrossRef]
- Silva Vieira, S.; Höfer, H. Prey spectrum of two army ant species in central Amazonia, with special attention on their effect on spider populations. Andrias 1994, 13, 189–198. [Google Scholar]
- Powell, S.; Franks, N.R. Ecology and the evolution of worker morphological diversity: A comparative analysis with Eciton army ants. Funct. Ecol. 2006, 20, 1105–1114. [Google Scholar] [CrossRef]
- O’Donnell, S.; Lattke, J.; Powell, S.; Kaspari, M. Army ants in four forests: Geographic variation in raid rates and species composition. J. Anim. Ecol. 2007, 76, 580–589. [Google Scholar] [CrossRef]
- Manubay, J.A.; Powell, S. Detection of prey odours underpins dietary specialization in a Neotropical top-predator: How army ants find their ant prey. J. Anim. Ecol. 2020, 89, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Hou, C.; Kaspari, M.; Vander Zanden, H.B.; Gillooly, J.F. Energetic basis of colonial living in social insects. Proc. Natl. Acad. Sci. USA 2010, 107, 3634–3638. [Google Scholar] [CrossRef]
- Schneirla, T.; Brown, R.Z. Army-ant life and behavior under dry-season. 4. Further investigation of cyclic processess in behavioral and reproductive functions. Bull. Am. Museum Nat. Hist. 1950, 95, 263–354. [Google Scholar]
- Franks, N.R.; Sendova-Franks, A.B. Army Ants: A Collective Intelligence: A neural network Seems an apt analogy as a colony of army ants navigates the tropical rain forest. Am. Sci. 1989, 77, 138–145. [Google Scholar]
- Kaspari, M.; Powell, S.; Lattke, J.; O’Donnell, S. Predation and patchiness in the tropical litter: Do swarm-raiding army ants skim the cream or drain the bottle? J. Anim. Ecol. 2011, 80, 818–823. [Google Scholar] [CrossRef] [PubMed]
- Franks, N.R. Reproduction, foraging efficiency and worker polymorphism in army ants. In Experimental Behavioral Ecology and Sociobiology: In memoriam Karl von Frisch, 1886–1982; Hölldobler, B., Lindauer, M., Franks, N.R., Eds.; Wiley Online Library: Hoboken, NJ, USA, 1985; Volume Fortschrit, pp. 91–107. ISBN 087893460X. [Google Scholar]
- Franks, N.R.; Bossert, W.H. The influence of swarm raiding army ants on the patchiness and diversity of a tropical leaf litter ant community. In Tropical Rain Forest: Ecology and Management. Special Publication No. 2 of the British Ecological Society; Sutton, S.L., Whitmore, T.C., Chadwick, A.C., Eds.; Blackwell: Oxford, UK, 1983; pp. 151–163. [Google Scholar]
- Willis, E.O.; Oniki, Y. Birds and Army Ants. Annu. Rev. Ecol. Syst. 1978, 9, 243–263. [Google Scholar] [CrossRef]
- Rettenmeyer, C.W.; Rettenmeyer, M.E.; Joseph, J.; Berghoff, S.M. The largest animal association centered on one species: The army ant Eciton burchellii and its more than 300 associates. Insectes Soc. 2011, 58, 281–292. [Google Scholar] [CrossRef]
- Johnson, R.A. The behavior of birds attending army ant raids on Barro Colorado Island, Panama Canal Zone. Proc. Linn. Soc. New York 1954, 63–65, 41–70. [Google Scholar]
- Brumfield, R.T.; Tello, J.G.; Cheviron, Z.A.; Carling, M.D.; Crochet, N.; Rosenberg, K.V. Phylogenetic conservatism and antiquity of a tropical specialization: Army-ant-following in the typical antbirds (Thamnophilidae). Mol. Phylogenet. Evol. 2007, 45, 1–13. [Google Scholar] [CrossRef]
- Willis, E. the Behavior of Spotted Antbirds. Ornithol. Monogr. 1972, 10, 1–162. [Google Scholar] [CrossRef]
- Willis, E.O. Studies of the behavior of Lunulated and Salvin’s Antbirds. Condor 1968, 70, 128–148. [Google Scholar] [CrossRef]
- Faria, C.M.A.; Rodrigues, M. Birds and army ants in a fragment of the Atlantic Forest of Brazil. J. F. Ornithol. 2009, 80, 328–335. [Google Scholar] [CrossRef]
- Howell, T.R. Birds of a second-growth rain forest area of Nicaragua. Condor 1957, 59, 73–111. [Google Scholar] [CrossRef]
- Driver, R.J.; DeLeon, S.; O’Donnell, S. Novel observation of a raptor, collared forest-falcon (Micrastur semitorquatus) depredating a fleeing snake at an army ant (Eciton burchellii parvispinum) raid front. Wilson J. Ornithol. 2018, 130, 792–796. [Google Scholar] [CrossRef]
- Chaves-Campos, J.; DeWoody, J.A. The spatial distribution of avian relatives: Do obligate army-ant-following birds roost and feed near family members? Mol. Ecol. 2008, 17, 2963–2974. [Google Scholar] [CrossRef]
- Kumar, A.; O’Donnell, S. Fragmentation and elevation effects on bird–army ant interactions in neotropical montane forest of Costa Rica. J. Trop. Ecol. 2007, 23, 581. [Google Scholar] [CrossRef]
- Willson, S.K. Obligate army-ant following birds: A study of ecology, spatial movement patterns, and behavior in Amazonian Peru. Ornithol. Monogr. 2004, 1, 1–67. [Google Scholar] [CrossRef]
- Swartz, M.B. Bivouac checking, a novel behavior distinguishing obligate from opportunistic species of army-ant-following birds. Condor 2001, 103, 629–633. [Google Scholar] [CrossRef]
- Wrege, P.H.; Wikelski, M.; Mandel, J.T.; Rassweiler, T.; Couzin, I.D. Antbirds parasitizes foraging army ants. Ecology 2005, 86, 555–559. [Google Scholar] [CrossRef]
- Von Ihering, H. Biologie und Verbreitung der brasilianischen Arten von Eciton. Entomol. Mitteilungen 1912, 1, 226–335. [Google Scholar]
- Bequaert, J. The predaceous enemies of ants. Auk 1923, 40, 162. [Google Scholar] [CrossRef]
- Chesser, R.T. Comparative diets of obligate ant-following birds at a site in Northern Bolivia. Biotropica 1995, 27, 382–390. [Google Scholar] [CrossRef]
- Rylands, A.B.; Da Cruz, M.A.O.M.; Ferrari, S.F. An association between marmosets and army ants in Brazil. J. Trop. Ecol. 1989, 5, 113–116. [Google Scholar] [CrossRef]
- Martins, M.M. Foraging over army ants by Callithrix aurita (Primates: Callitrichidae): Seasonal occurrence? Rev. Biol. Trop. 2000, 48, 261–262. [Google Scholar]
- Eickwort, G.C. Associations of mites with social insects. Annu. Rev. Entomol. 1990, 35, 469–488. [Google Scholar] [CrossRef]
- Gotwald, W.H. Mites that live with army ants: A natural history of some myrmecophilous hitch-hikers, browsers, and parasites. J. Kansas Entomol. Soc. 1996, 69. [Google Scholar]
- Elzinga, R.J. Holdfast mechanisms in certain Uropodine mites (Acarina: Uropodina). Ann. Entomol. Soc. Am. 1978, 71, 896–900. [Google Scholar] [CrossRef]
- Elzinga, R.J.; Rettenmeyer, C.W. Seven new species of Circocylliba (Acarina: Uropodina) found on army ants. Acarologia 1975, 16, 595–611. [Google Scholar]
- Elzinga, R.J. Two new species of Planodiscus (Acari: Uropodine), range extensions and a synonomy within the genus. Acarologia 1990, 31, 229–233. [Google Scholar]
- Rettenmeyer, C.W. Notes on host specificity and behavior of myrmecophilous macrochelid mites. J. Kansas Entomol. Soc. 1962, 35, 358–360. [Google Scholar]
- Berghoff, S.M.; Wurst, E.; Ebermann, E.; Sendova-Franks, A.B.; Rettenmeyer, C.W.; Franks, N.R. Symbionts of societies that fission: Mites as guests or parasites of army ants. Ecol. Entomol. 2009, 34, 684–695. [Google Scholar] [CrossRef]
- Krantz, G.W. A Manual of Acarology, 2nd ed.; Oregon State University Book Stores: Corvallis, OR, USA, 1978. [Google Scholar]
- Brown, B.V.; Feener, D.H. Parasitic phorid flies (Diptera: Phoridae) associated with army ants (Hymenoptera: Formicidae: Ecitoninae, Dorylinae) and their conservation biology. Biotropica 1998, 30, 482–487. [Google Scholar] [CrossRef]
- Rettenmeyer, C.W. Observations on the biology and taxonomy of flies found over swarm raids of army ants (Diptera: Tachinidae, Conopidae). Univ. Kans. Sci. Bull. 1961, 42, 993–1066. [Google Scholar]
- Feener, D.H.; Jacobs, L.F.; Schmidt, J.O. Specialized parasitoid attracted to a pheromone of ants. Anim. Behav. 1996, 51, 61–66. [Google Scholar] [CrossRef][Green Version]
- Santiago-Jiménez, Q.J.; Espinosa De Los Monteros, A. Exploring myrmecophily based on the phylogenetic interrelationships of Falagonia Sharp, 1883 (Coleoptera: Staphylinidae: Aleocharinae) and allied genera. Syst. Entomol. 2016, 41, 794–807. [Google Scholar] [CrossRef]
- Von Beeren, C.; Maruyama, M.; Kronauer, D.J.C. Community sampling and integrative taxonomy reveal new species and host specificity in the army ant-associated beetle genus Tetradonia (Coleoptera, Staphylinidae, Aleocharinae). PLoS ONE 2016, 11, 1–19. [Google Scholar] [CrossRef]
- Seevers, C.H. A revision of the Vatesini, a tribe of neotropical myrmecophiles (Coleoptera, Staphylinidae). Rev. Bras. Entomol. 1958, 8, 181–202. [Google Scholar]
- Akre, R.D.; Rettenmeyer, C.W. Trail-following by guests of army ants (Hymenoptera: Formicidae: Ecitonini). J. Kansas Entomol. Soc. 1968, 41, 165–174. [Google Scholar]
- Akre, R. The behaviour of Euxenister and Pulvinister, histerid beetles associated with army ants. (Coleoptera: Histeridae; Hymenoptera: Formicidae: Dorylinae.). Pan Pacific Entomol. 1968, 44, 87–101. [Google Scholar]
- Mann, W.M. Guests of Eciton Hamatum (Fab.) Collected by Professor W. M. Wheeler. Psyche (New York) 1925, 32, 166–177. [Google Scholar] [CrossRef]
- Seevers, C.H. The systematics, evolution and zoogeography of staphylinid beetles associated with army ants (Coleoptera, Staphylinidae). Fieldiana Zool. 1965, 47, 137–351. [Google Scholar]
- Akre, R.D.; Rettenmeyer, C.W. Behavior of Staphylinidae associated with army ants (Formicidae: Ecitonini). J. Kansas Entomol. Soc. 1966, 39, 745–782. [Google Scholar]
- Kistner, D.H.; Jacobson, H.R. Cladistic analysis and taxonomic revision of the ecitophilous tribe Ecitocharini with studies of their behavior and evolution (Coleoptera, Staphylinidae, Aleocharinae). Sociobiology 1990, 17, 333–480. [Google Scholar]
- Reichensperger, A. Ecitophilen aus Costa Rica (II), Brasilien und Peru (Staph. Hist. Clavig.). Rev. Entomol. 1933, 3, 179–194. [Google Scholar]
- Reichensperger, A. Beitrag zur Kenntnis der Myrmecophilenfauna Brasiliens und Costa Ricas III. (Col. Staphyl. Hist.). Arb. iiber Morphol. Taxon. Entomol. Berlin Dahlem 1935, 2, 188–218. [Google Scholar]
- von Beeren, C.; Brückner, A.; Maruyama, M.; Burke, G.; Wieschollek, J.; Kronauer, D.J.C. Chemical and behavioral integration of army ant-associated rove beetles—A comparison between specialists and generalists. Front. Zool. 2018, 15, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Akre, R.; Torgerson, R. Behavior of Vatesus beetles associated with army ants (Coleoptera: Staphylinidae). Pan Pacific Entomol. 1969, 45, 269–281. [Google Scholar]
- Von Beeren, C.; Maruyama, M.; Kronauer, D.J.C. Cryptic diversity, high host specificity and reproductive synchronization in army ant-associated Vatesus beetles. Mol. Ecol. 2016, 25, 990–1005. [Google Scholar] [CrossRef] [PubMed]
- Loomis, H.F. New myrmecophilous millipeds from Barro Colorado Island, Canal Zone, and Mexico. Source J. Kansas Entomol. Soc. Kansas Entomol. Soc. 1959, 32, 1–7. [Google Scholar]
- Rettenmeyer, C.W. The behavior of millipeds found with neotropical army ants. J. Kansas Entomol. Soc. 1962, 35, 377–384. [Google Scholar]
- Ferrière, C. Nouveaux Diapriides du Brésil, hôtes des Eciton. Zool. Anz. 1929, 82, 156–171. [Google Scholar]
- Borgmeier, T. Sobre alguns Diapriideos myrmecophilos, principalmente do Brasil (Hym. Diapriidae). Rev. Entomol. 1939, 10, 530–535. [Google Scholar]
- Zikán, J.F. Myrmekophilie bei Hesperiden? Entomol. Rundschau 1929, 46, 27–28. [Google Scholar]
- Drummond, B.A. Buttterflies associated with an army ant swarm raid in Honduras. J. Lepid. Soc. 1976, 30, 237–238. [Google Scholar]
- Haber, W.A. Evolutionary Ecology of Tropical Mimetic Butterflies (Lepidoptera: Ithomiiae). Ph.D. Thesis, University of Minnesota, Minneapolis, MN, USA, 1978. [Google Scholar]
- Young, A.M. Butterflies associated with an army ant swarm raid in Honduras: The “feeding hypothesis” as an alternate explanation. J. Lepid. Soc. 1977, 31, 190. [Google Scholar]
- Ray, T.S.; Andrews, C.C. Antbutterflies: Butterflies that follow army ants to feed on antbird droppings. Science 1980, 210, 1147–1148. [Google Scholar] [CrossRef]
- Austin, G.T.; Brock, J.P.; Mielke, O.H.H. Ants, birds, and skippers. Trop. Lepidop. 1993, 4, 1–11. [Google Scholar]
- Funaro, C.F.; Kronauer, D.J.C.; Moreau, C.S.; Goldman-Huertas, B.; Pierce, N.E.; Russell, J.A. Army ants harbor a host-specific clade of Entomoplasmatales bacteria. Appl. Environ. Microbiol. 2011, 77, 346–350. [Google Scholar] [CrossRef]
- Łukasik, P.; Newton, J.A.; Sanders, J.G.; Hu, Y.; Moreau, C.S.; Kronauer, D.J.C.; O’Donnell, S.; Koga, R.; Russell, J.A. The structured diversity of specialized gut symbionts of the New World army ants. Mol. Ecol. 2017, 26, 3808–3825. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011. [Google Scholar] [CrossRef] [PubMed]
- Tiede, J.; Scherber, C.; Mutschler, J.; McMahon, K.D.; Gratton, C. Gut microbiomes of mobile predators vary with landscape context and species identity. Ecol. Evol. 2017, 7, 8545–8557. [Google Scholar] [CrossRef]
- Paula, D.P.; Linard, B.; Crampton-Platt, A.; Srivathsan, A.; Timmermans, M.J.T.N.; Sujii, E.R.; Pires, C.S.S.; Souza, L.M.; Andow, D.A.; Vogler, A.P. Uncovering trophic interactions in arthropod predators through DNA shotgun-sequencing of gut contents. PLoS ONE 2016, 11, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Padial, J.M.; Miralles, A.; De la Riva, I.; Vences, M. The integrative future of taxonomy. Front. Zool. 2010, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Laurance, W.F. Theory meets reality: How habitat fragmentation research has transcended island biogeographic theory. Biol. Conserv. 2008. [CrossRef]
- Offerman, H.L.; Dale, V.H.; Pearson, S.M.; Bierregaard, R.O.; O’Neill, R.V. Effects of forest fragmentation on neotropical fauna: Current research and data availability. Environ. Rev. 1995, 3, 191–211. [Google Scholar] [CrossRef]
- Levings, S.C. Seasonal, annual, and among-site variation in the ground ant community of a deciduous tropical forest: Some causes of patchy species distributions. Ecol. Monogr. 1983. [Google Scholar] [CrossRef]
- Meisel, J.E. Thermal ecology of the neotropical army ant Eciton burchellii. Ecol. Appl. 2006, 16, 913–922. [Google Scholar] [CrossRef]
- Kumar, A.; Longino, J.T.; Colwell, R.K.; Donnell, S.O. Elevational patterns of diversity and abundance of eusocial paper wasps (Vespidae) in Costa Rica. Biotropica 2009, 41, 338–346. [Google Scholar] [CrossRef]
- Willson, S.K.; Sharp, R.; Ramler, I.P.; Sen, A. Spatial movement optimization in amazonian Eciton burchellii army ants. Insectes Soc. 2011, 58, 325–334. [Google Scholar] [CrossRef]
- Lozano-Zambrano, F.H.; Ulloa-Chacón, P.; Armbrecht, I. Ants: Species-Area relationship in tropical dry forest fragments. Neotrop. Entomol. 2009, 38, 44–54. [Google Scholar] [CrossRef] [PubMed]
- Perfecto, I.; Vandermeer, J. Biodiversity conservation in tropical agroecosystems: A new conservation paradigm. Ann. N. Y. Acad. Sci. 2008, 1134, 173–200. [Google Scholar] [CrossRef]
- Perfecto, I.; Snelling, R. Biodiversity and the transformation of a tropical agroecosystem: Ants in coffee plantations. Ecol. Appl. 1995, 5, 1084–1097. [Google Scholar] [CrossRef]
- Perfecto, I.; Rice, R.A.; Greenberg, R.; van der Voort, M.E. Shade coffee: Update on a disappearing refuge for biodiversity. Bioscience 1996, 46, 598–608. [Google Scholar] [CrossRef]
- Roberts, D.L.; Cooper, R.J.; Petit, L.J. Use of premontane moist forest and shade coffee agroecosystems by army ants in western Panama. Conserv. Biol. 2000, 14, 192–199. [Google Scholar] [CrossRef]
- Delabie, J.H.C.; Jahyny, B.; Do Nascimento, I.C.; Mariano, C.S.F.; Lacau, S.; Campiolo, S.; Philpott, S.M.; Leponce, M. Contribution of cocoa plantations to the conservation of native ants (Insecta: Hymenoptera: Formicidae) with a special emphasis on the Atlantic Forest fauna of southern Bahia, Brazil. Biodivers. Conserv. 2007, 16, 2359–2384. [Google Scholar] [CrossRef]
- Stouffer, P.C. Survival of the ant followers. Nat. Hist. Hist. 1998, 107, 40–43. [Google Scholar]
- Pérez-Espona, S.; McLeod, J.E.; Franks, N.R. Landscape genetics of a top neotropical predator. Mol. Ecol. 2012, 21. [Google Scholar] [CrossRef] [PubMed]
- Soare, T.W.; Kumar, A.; Naish, K.A.; O’Donnell, S. Genetic evidence for landscape effects on dispersal in the army ant Eciton burchellii. Mol. Ecol. 2014, 23, 96–109. [Google Scholar] [CrossRef]
- Berghoff, S.M.; Kronauer, D.J.C.; Edwards, K.J.; Franks, N.R. Dispersal and population structure of a New World predator, the army ant Eciton burchellii. J. Evol. Biol. 2008, 21, 1125–1132. [Google Scholar] [CrossRef]
- Jaffé, R.; Moritz, R.F.A.; Kraus, F.B. Gene flow is maintained by polyandry and male dispersal in the army ant Eciton burchellii. Popul. Ecol. 2009, 51, 227–236. [Google Scholar] [CrossRef]
- Soare, T.W.; Kumar, A.; Naish, K.A.; O’Donnell, S. Multi-year genetic sampling indicates maternal gene flow via colony emigrations in the army ant Eciton burchellii parvispinum. Insectes Soc. 2020, 67, 155–166. [Google Scholar] [CrossRef]
- Schneirla, T. Army-ant life and behavior under dry-season conditions with special reference to reproductive functions. II. The appearance and fate of the males. Zoologica 1948, 33, 89–112. [Google Scholar]
- Baldridge, R.S.; Rettenmeyer, C.W.; Watkins, J.F., II. Seasonal, nocturnal and diurnal flight periodicities of Nearctic army ant males (Hymenoptera: Formicidae). J. Kansas Entomol. Soc. 1980. [Google Scholar]
- Stouffer, P.C.; Bierregaard, R.O. Use of Amazonian forest fragments by understory insectivorous birds. Ecology 1995, 76, 2429–2445. [Google Scholar] [CrossRef]
- Hooper, D.U.; Chapin, F.S.; Ewel, J.J.; Hector, A.; Inchausti, P.; Lavorel, S.; Lawton, J.H.; Lodge, D.M.; Loreau, M.; Naeem, S.; et al. Effects of biodiversity on ecosystem functioning: A consensus of current knowledge. Ecol. Monogr. 2005, 75, 3–35. [Google Scholar] [CrossRef]
- Hooper, D.U.; Adair, E.C.; Cardinale, B.J.; Byrnes, J.E.K.; Hungate, B.A.; Matulich, K.L.; Gonzalez, A.; Duffy, J.E.; Gamfeldt, L.; Connor, M.I. A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 2012, 486, 105–108. [Google Scholar] [CrossRef]
- Newbold, T.; Hudson, L.N.; Hill, S.L.L.; Contu, S.; Lysenko, I.; Senior, R.A.; Börger, L.; Bennett, D.J.; Choimes, A.; Collen, B.; et al. Global effects of land use on local terrestrial biodiversity. Nature 2015, 520, 45–50. [Google Scholar] [CrossRef]
- Stratford, J.A.; Robinson, W.D. Gulliver travels to the fragmented tropics: Geographic variation in mechanisms of avian extinction. Front. Ecol. Environ. 2005, 3, 91–98. [Google Scholar] [CrossRef]
- Lovejoy, T.E.; Bierregaard, R.O.; Rylands, A.B. Edge and other effects of isolation on Amazon forest fragments. In Conservation Biology: The Science of Scarcity and Diversity; Soulé, M.E., Ed.; Sinauer Associates: Sunderland, MA, USA, 1986; pp. 257–285. [Google Scholar]
- Harper, L.H. The persistence of ant-following birds in small forest fragments. Acta Amaz. 1989, 19, 249–283. [Google Scholar] [CrossRef]
- Sekercioglu, Ç.H.; Ehrlich, P.R.; Daily, G.C.; Aygen, D.; Goehring, D.; Sandí, R.F. Disappearance of insectivorous birds from tropical forest fragments. Proc. Natl. Acad. Sci. USA 2002, 99, 263–267. [Google Scholar] [CrossRef] [PubMed]
- Koh, L.P.; Dunn, R.R.; Sodhi, N.S.; Colwell, R.K.; Proctor, H.C.; Smith, V.S. Species coextinctions and the biodiversity crisis. Science 2004, 305, 1632–1634. [Google Scholar] [CrossRef] [PubMed]
- Lees, A.C.; Peres, C.A. Habitat and life history determinants of antbird occurrence in variable-sized Amazonian forest fragments. Biotropica 2010, 42, 614–621. [Google Scholar] [CrossRef]
- Margules, C.R.; Pressey, R.L.; Willias, P.H. Representing biodiversity: Data and procedures for identifying priority areas for conservation. J. Biosci. 2002, 27, 309–326. [Google Scholar] [CrossRef] [PubMed]
- Simberloff, D. Flagships, umbrellas, and keystones: Is single-species management passé in the landscape area? Biol. Conserv. 1998, 83, 247–257. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Dinerstein, E.; Olson, D.M.; Loucks, C. Who’s where in North? Patterns of species richness and the utility of indicator taxa. Bioscience 1999, 49, 369–381. [Google Scholar] [CrossRef]
- Pearson, D.L.; Cassola, F. World-wide species richness patterns of Tiger beetles (Coleoptera: Cicindelidae): Indicator taxon for biodiversity and conservation studies. Conserv. Biol. 1992, 6, 376–391. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Reid, W.V. Biodiversity hotspots. Trends Ecol. Evol. 1998, 13, 275–280. [Google Scholar] [CrossRef]
- Van Jaarsveld, A.S.; Freitag, S.; Chown, S.L.; Muller, C.; Koch, S.; Hull, H.; Bellamy, C.; Krüger, M.; Endrödy-Younga, S.; Mansell, M.W.; et al. Biodiversity assessment and conservation strategies. Science 1998, 279, 2106–2108. [Google Scholar] [CrossRef] [PubMed]
- Frankel, O.H.; Soulé, M.E. Conservation and Evolution; Cambridge University Press: Cambridge, UK, 1981; ISBN 0521232759. [Google Scholar]
- Martikainen, P.; Kaila, L.; Haila, Y. Threatened Beetles in White-Backed Woodpecker Habitats. Conserv. Biol. 1998, 12, 293–301. [Google Scholar] [CrossRef]
- Swengel, S.R.; Swengel, A.B. Correlations in abundance of grassland songbirds and prairie butterflies. Biol. Conserv. 1999, 90, 1–11. [Google Scholar] [CrossRef]
- Launer, A.E.; Murphy, D.D. Umbrella species and the conservation of habitat fragments: A case of threatened butterfly and a vanishing grassland ecosystem. Biol. Conserv. 1994, 69, 145–153. [Google Scholar] [CrossRef]
- Berger, J. Population constraints associated with the use of Black rhinos as an umbrella species for desert herbivores. Conserv. Biol. 1997, 11, 69–78. [Google Scholar] [CrossRef]
- Caro, T.M. Umbrella species: Critique and lessons from East Africa. Anim. Conserv. 2003, 6, 171–181. [Google Scholar] [CrossRef]
- Zhang, C.; Zhu, R.; Sui, X.; Chen, K.; Li, B.; Chen, Y. Ecological use of vertebrate surrogate species in ecosystem conservation. Glob. Ecol. Conserv. 2020, 24, e01344. [Google Scholar] [CrossRef]
- Sergio, F.; Caro, T.; Brown, D.; Clucas, B.; Hunter, J.; Ketchum, J.; McHugh, K.; Hiraldo, F. Top predators as conservation tools: Ecological rationale, assumptions, and efficacy. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 1–19. [Google Scholar] [CrossRef]
- Whiteman, N.K.; Sites, R.W. Aquatic insects as umbrella species for ecosystem protection in Death Valley National Park. J. Insect Conserv. 2008, 12, 499–509. [Google Scholar] [CrossRef]
- Lambeck, R.J. Focal species: A multi-species umbrella for nature conservation. Conserv. Biol. 1997, 11, 849–856. [Google Scholar] [CrossRef]
- Kumar, A.; O’Donnell, S. Elevation and forest clearing effects on foraging differ between surface—And subterranean—Foraging army ants (Formicidae: Ecitoninae). J. Anim. Ecol. 2009, 78, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Kaspari, M. A primer on ant ecology. In Ants: Standard Methods for Measuring and Monitoring Biodiversity; Agosti, D., Majer, J.D., Alonso, L.E., Schultz, T.R., Eds.; Smithsonian Institution Press: Washington, DC, USA, 2000; pp. 9–24. [Google Scholar]
- Barbour, W.; Schlesinger, C. Who’s the boss? Post-colonialism, ecological research and conservation management on Australian Indigenous lands. Ecol. Manag. Restor. 2012, 13, 36–41. [Google Scholar] [CrossRef]
- Domínguez, L.; Luoma, C. Decolonising conservation policy: How colonial land and conservation ideologies persist and perpetuate indigenous injustices at the expense of the environment. Land 2020, 9, 65. [Google Scholar] [CrossRef]
- Baker, K.; Eichhorn, M.P.; Griffiths, M. Decolonizing field ecology. Biotropica 2019, 51, 288–292. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pérez-Espona, S. Eciton Army Ants—Umbrella Species for Conservation in Neotropical Forests. Diversity 2021, 13, 136. https://doi.org/10.3390/d13030136
Pérez-Espona S. Eciton Army Ants—Umbrella Species for Conservation in Neotropical Forests. Diversity. 2021; 13(3):136. https://doi.org/10.3390/d13030136
Chicago/Turabian StylePérez-Espona, Sílvia. 2021. "Eciton Army Ants—Umbrella Species for Conservation in Neotropical Forests" Diversity 13, no. 3: 136. https://doi.org/10.3390/d13030136
APA StylePérez-Espona, S. (2021). Eciton Army Ants—Umbrella Species for Conservation in Neotropical Forests. Diversity, 13(3), 136. https://doi.org/10.3390/d13030136