
Orchid Extinction over the Last 150 Years in the Czech Republic



Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. The Database
2.3. Nomenclature
2.4. Data Analysis
3. Results and Discussion
3.1. Extinct Species
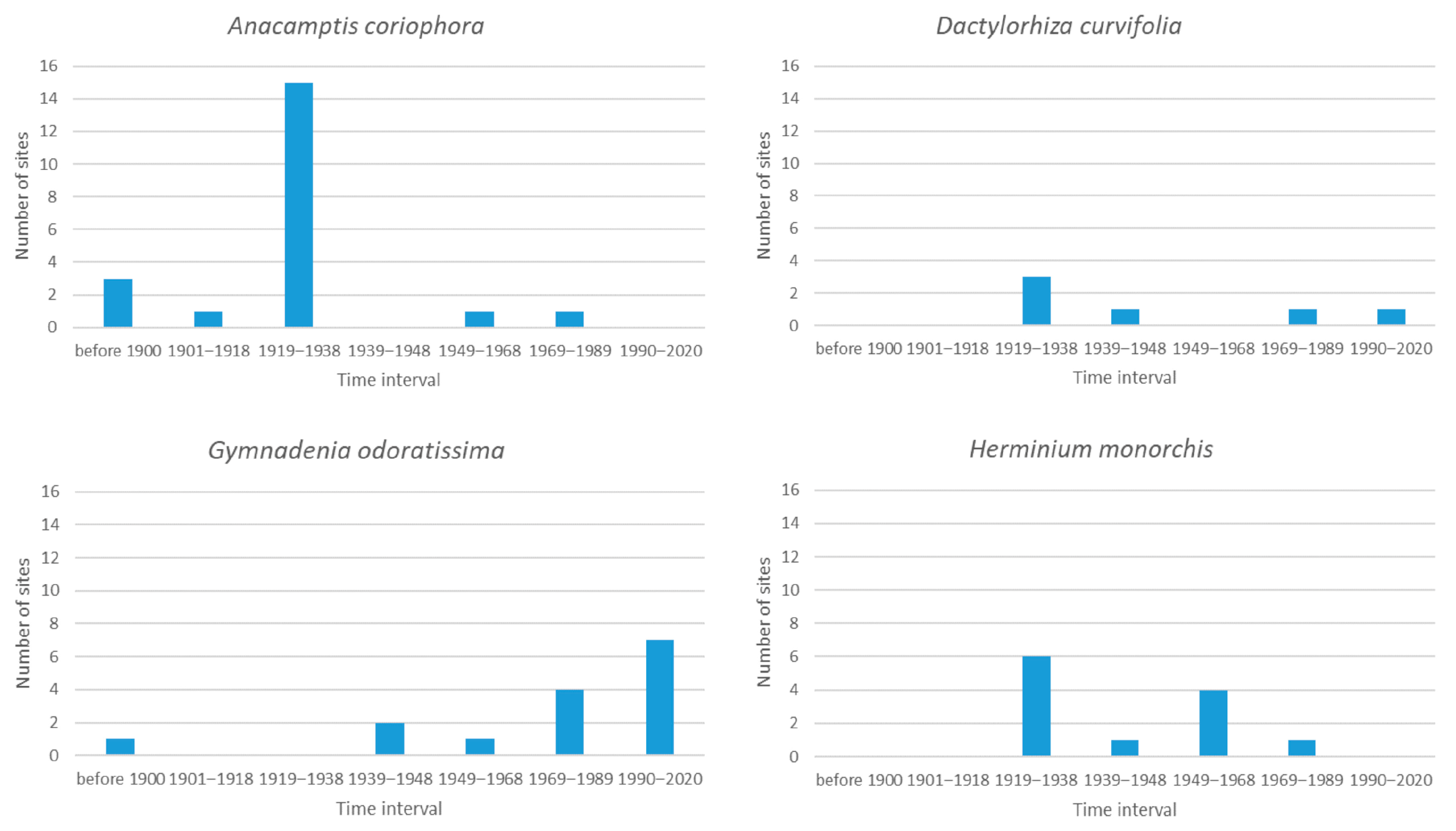
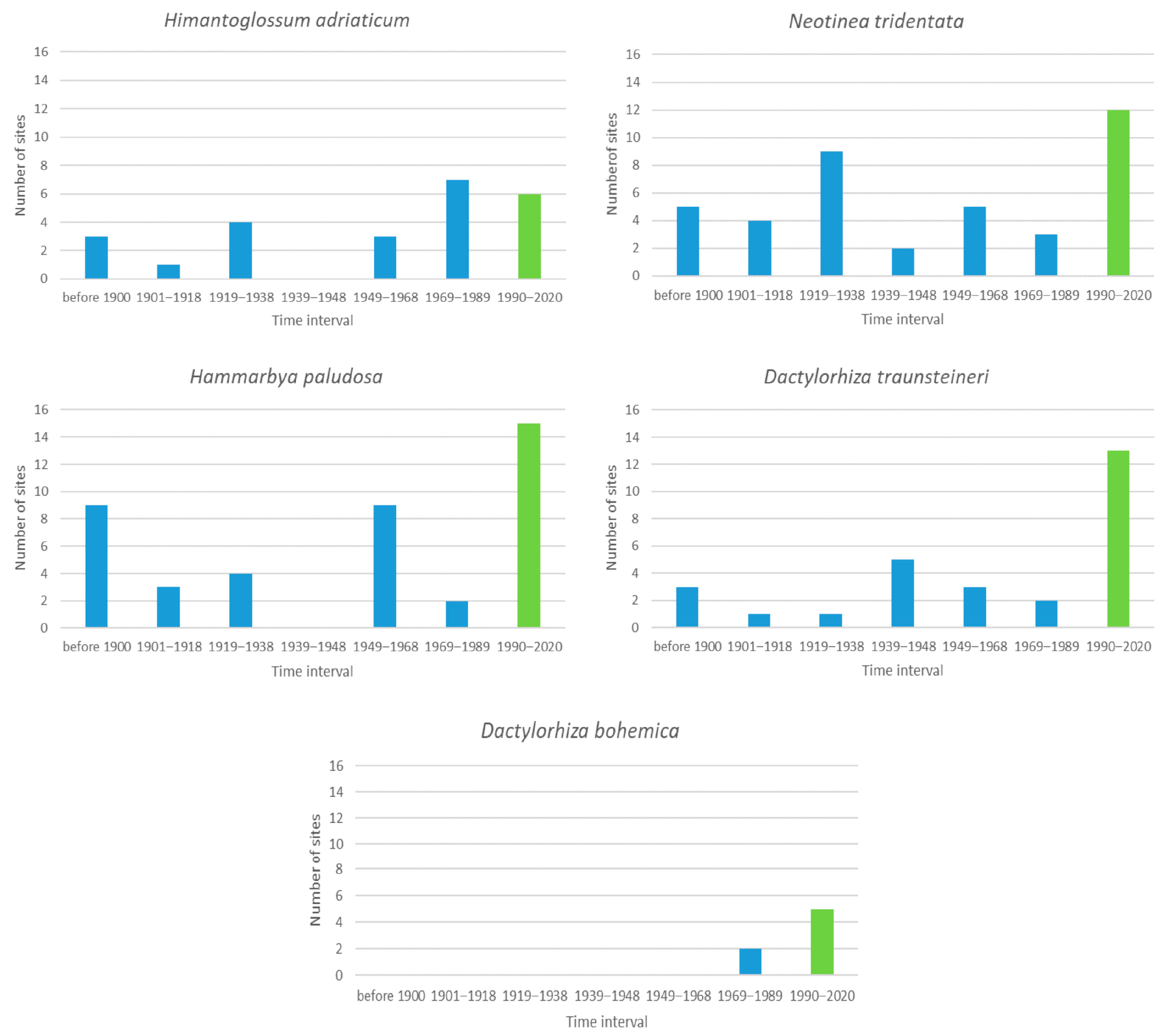
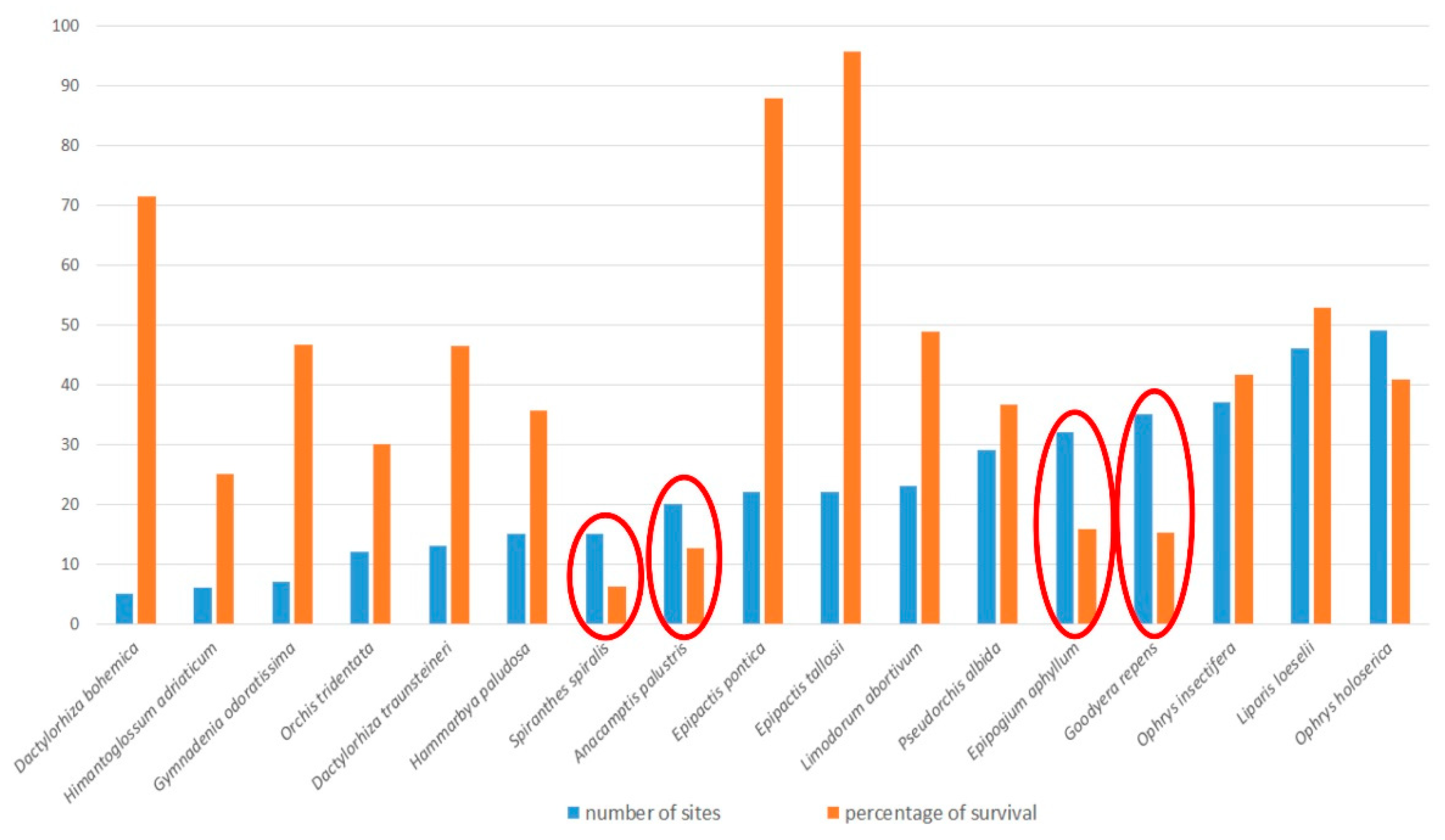
3.2. Species with 1 to 20 Sites
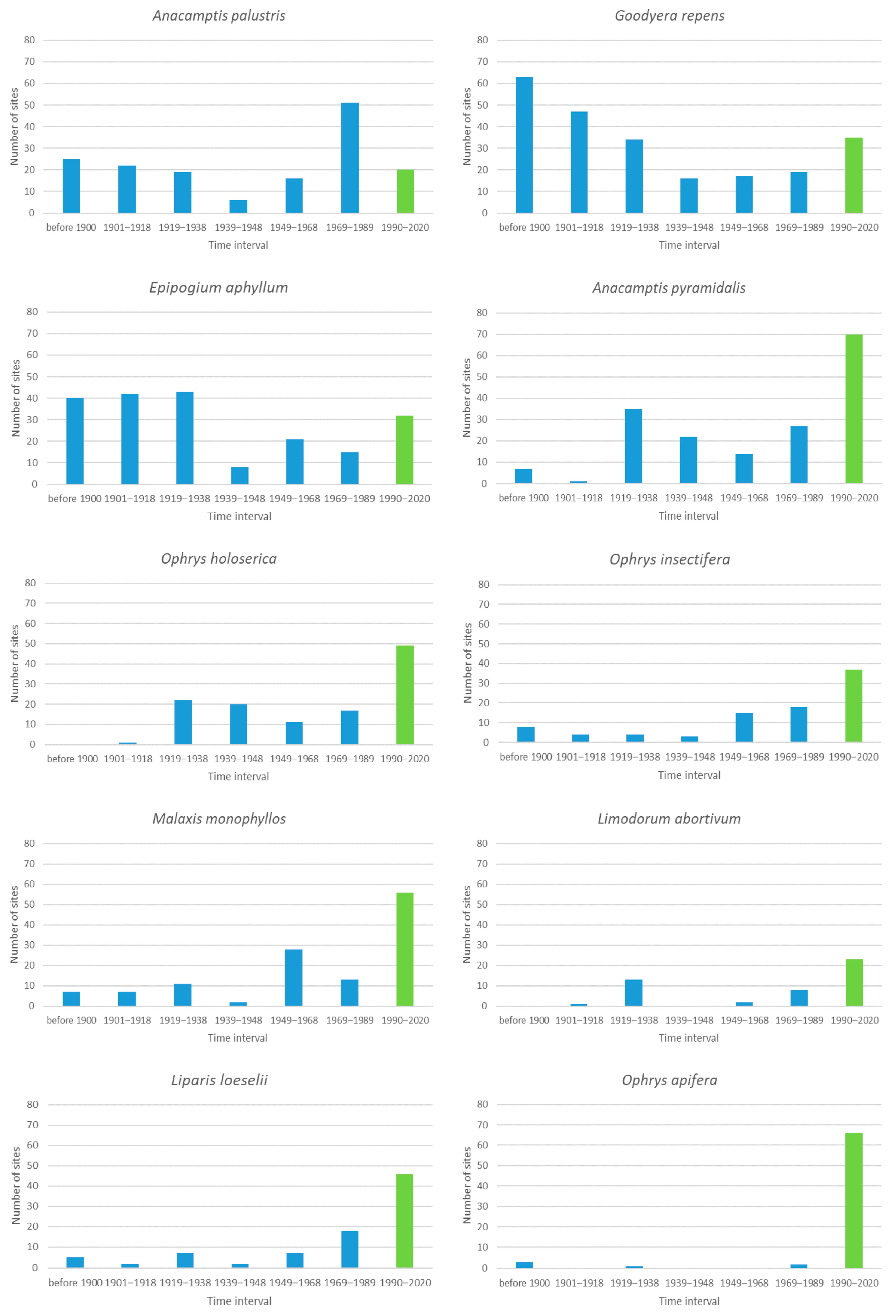
3.3. Species with 21 to 100 Sites
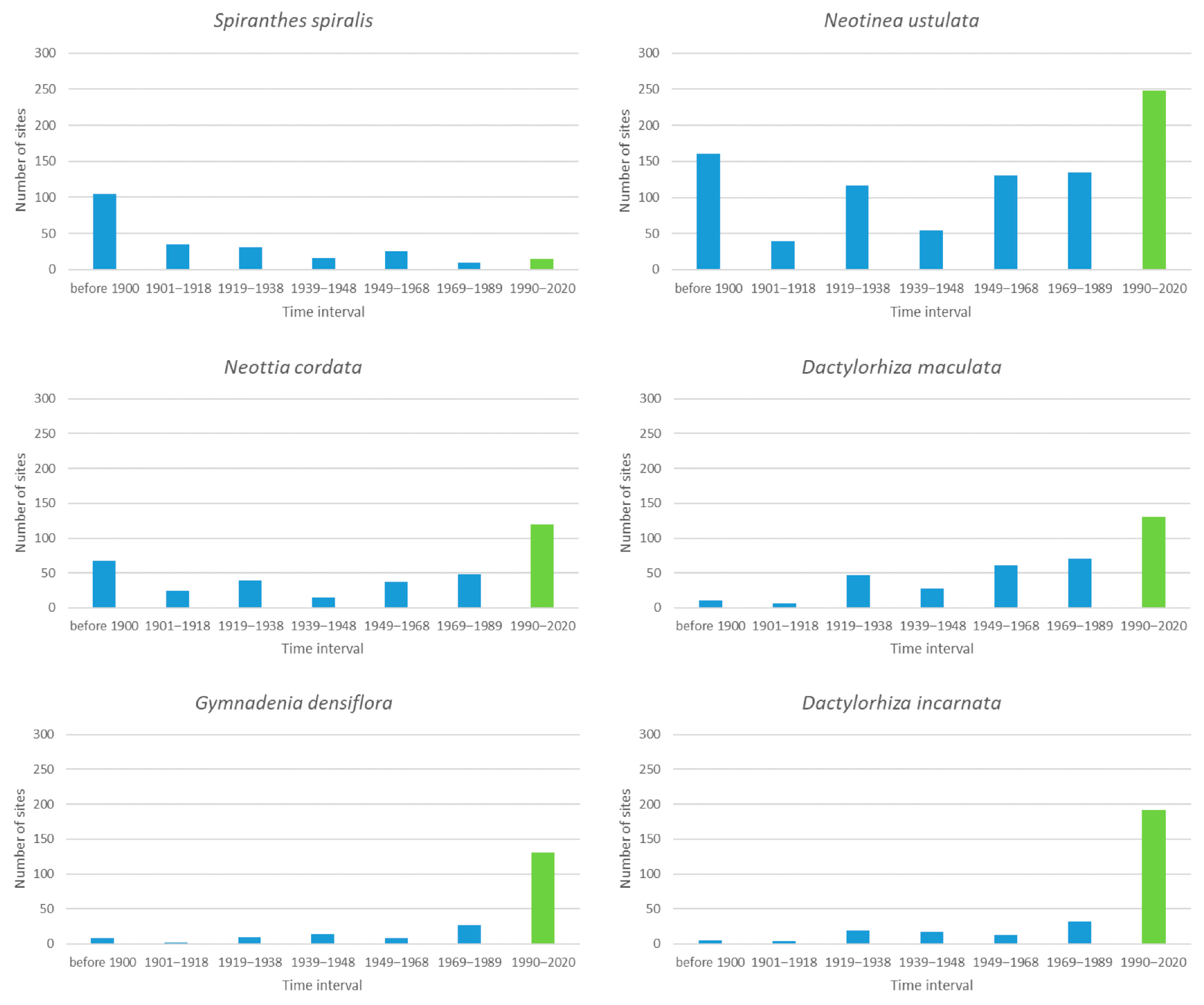
3.4. Species with 101 to 300 Sites
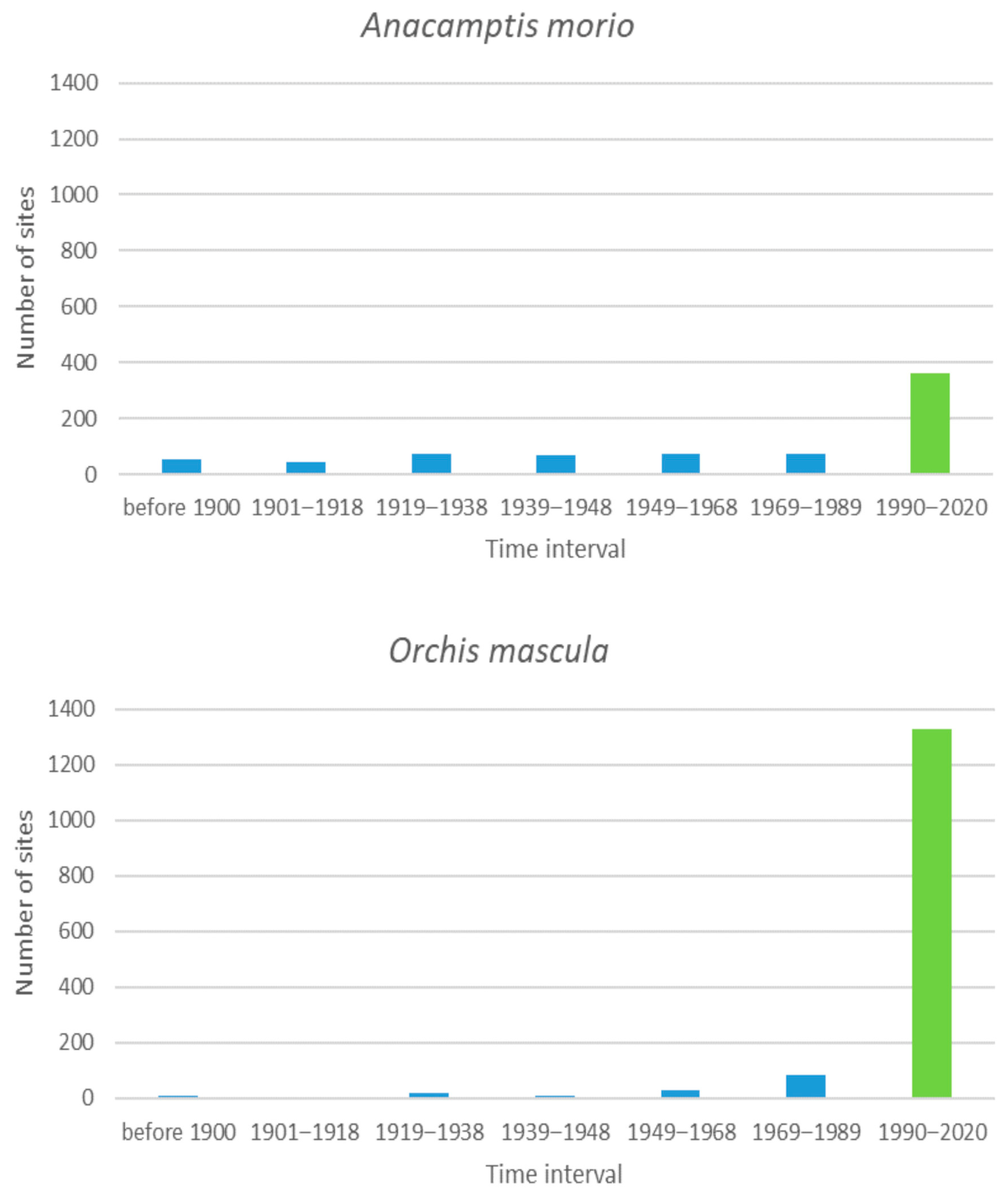
3.5. Species with More Than 300 Sites
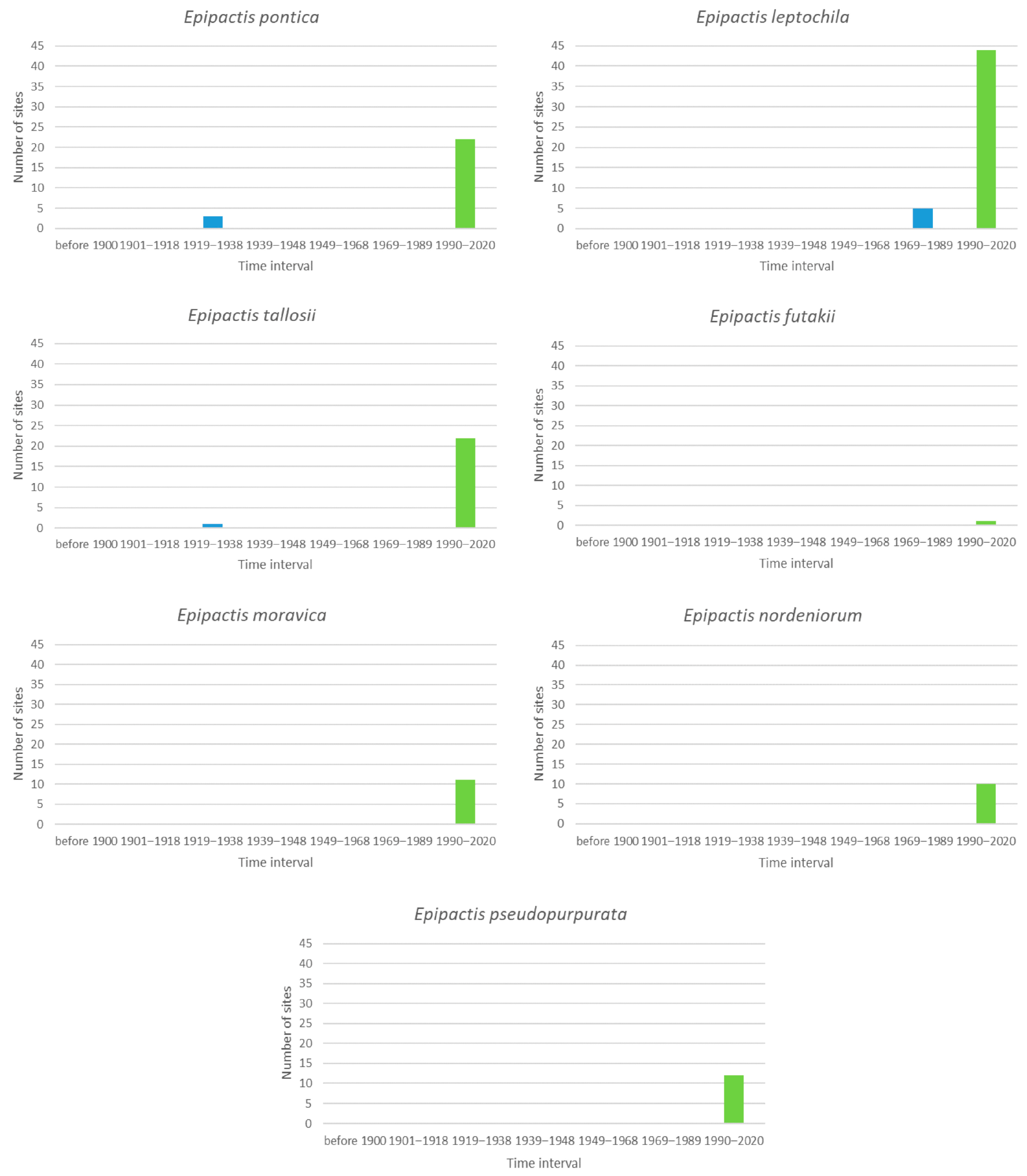
3.6. New Species of Epipactis
3.7. General Analysis
4. Other Aspects That May Cause Decline in Number of Orchid Sites
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, K.J.; Blackburn, T.M. Patterns and Process in Macroecology; Blackwell Science Ltd.: Cambridge, UK, 2000. [Google Scholar]
- Fischer, M.; Stöcklin, J. Local extinction of plants in remnants of extensively used calcareous grasslands 1950–1985. Conserv. Biol. 1997, 11, 727–737. [Google Scholar] [CrossRef]
- Kull, T.; Kukk, T.; Leht, M.; Krall, H.; Kukk, Ü.; Kull, K.; Kuusk, V. Distribution trends of rare vascular plant species in Estonia. Biodivers. Conserv. 2002, 11, 171–196. [Google Scholar] [CrossRef]
- Bianchi, F.J.J.A.; Honěk, A.; van der Werf, W. Changes in agricultural land use can explain population decline in a ladybeetle species in the Czech Republic: Evidence from a process-based spatially explicit model. Landsc. Ecol. 2007, 22, 1541–1554. [Google Scholar] [CrossRef]
- Le Roux, J.J.; Hui, C.; Castillo, M.L.; Iriondo, J.M.; Keet, J.-H.; Khapugin, A.A.; Médail, F.; Rejmánek, M.; Theron, G.; Yannelli, F.A.; et al. Recent anthropogenic plant extinctions differ in biodiversity hotspots and coldspots. Curr. Biol. 2019, 29, 2912–2918. [Google Scholar] [CrossRef] [PubMed]
- Khapugin, A.A.; Kuzmin, I.V.; Silaeva, T.B. Anthropogenic drivers leading to reagional extinction of threatened plants: Insights from regional Red Data Books of Russia. Biodivers. Conserv. 2020, 29, 2765–2777. [Google Scholar] [CrossRef]
- Knapp, W.M.; Frances, A.; Noss, R.; Naczi, R.F.C.; Weakley, A.; Gann, G.D.; Baldwin, B.G.; Miller, J.; McIntyre, P.; Mishler, B.D.; et al. Vascular plant extinction in the continental United States and Canada. Conserv. Biol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Bilz, M.; Kell, S.P.; Maxted, N.; Lansdown, R.V. European Red List of Vascular Plants; Publication Office of the European Union: Luxembourg, 2011. [Google Scholar]
- Henle, K.; Alard, D.; Clitherow, J.; Cobb, P.; Firbank, L.; Kull, T.; McCracken, D.; Moritz, R.F.A.; Niemela, J.; Rebane, M.; et al. Identifying and managing the conflicts between agriculture and biodiversity conservation in Europe—A review. Agr. Ecosyst. Environ. 2008, 124, 60–71. [Google Scholar] [CrossRef]
- Stewart, J. The Conservation of European Orchids. Nature and Environment, No. 57; Council of Europe Press: Strasbourg, France, 1992. [Google Scholar]
- Tsiftsis, S.; Tsiripidis, I.; Trigas, P. Identifying important areas for orchid conservation in Crete. Eur. J. Environ. Sci. 2011, 1, 28–37. [Google Scholar] [CrossRef]
- Kull, T.; Selgis, U.; Pecina, M.V.; Metsare, M.; Ilves, A.; Tali, K.; Sepp, K.; Kull, K.; Shefferson, R.P. Factors influencing IUCN threat levels to orchids across Europe on the basis of national Red Lists. Ecol. Evol. 2016, 6, 6245–6265. [Google Scholar] [CrossRef]
- Wädekin, K.E. Agrarian Policies in Communist Europe. A Critical Introduction; Allanheld, Osmun Publishers: Totowa, NJ, USA, 1982. [Google Scholar]
- Adams, A.E.; Adams, J.S. Men Versus Systems. Agriculture in the USSR, Poland, and Czechoslovakia; Free Press: New York, NY, USA, 1971. [Google Scholar]
- Krčmářová, J.; Jeleček, L. Czech traditional agroforestry: Historic and current status. Agroforest Syst. 2017, 91, 1087–1100. [Google Scholar] [CrossRef]
- Skaloš, J.; Wber, M.; Lipský, Z.; Trpáková, I.; Šnatrůčková, M.; Uhlířová, L.; Kukla, P. Using old military survey maps and orthophotograph maps to analyse long-term land cover changes—Case study (Czech Republic). Appl. Geogr. 2011, 31, 426–438. [Google Scholar] [CrossRef]
- Reif, J.; Voříšek, P.; Šťastný, K.; Bejček, V.; Petr, J. Agricultural intensification and farmland birds: New insight from a central European country. Ibis 2008, 150, 596–605. [Google Scholar] [CrossRef]
- Swarts, D.N.; Dixon, K.W. Terrestrial orchid conservation in the age of extinction. Ann. Bot. 2009, 104, 543–556. [Google Scholar] [CrossRef]
- Djordjević, V.; Lukašić, D.; Jovanović, S.; Stevanović, V. Distribution and conservation status of some rare and threatened orchid taxa in the central Balkans and the southern part of the Pannonian Plain. Wulfenia 2017, 24, 143–162. [Google Scholar]
- Khapugin, A.A. A global systematic review on orchid data in protected areas. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 19–33. [Google Scholar] [CrossRef]
- Govaerts, R. World Checklist of Orchidaceae; The Royal Botanic Gardens: Kew, UK, 2020; Available online: http://wcsp.science.kew.org/ (accessed on 31 January 2020).
- Cribb, P.J.; Kell, S.P.; Dixon, K.W.; Barrett, R.L. Orchid conservation: A global perspective. In Orchid Conservation; Dixon, K.W., Kell, S.P., Barrett, R.L., Cribb, P.J., Eds.; Natural History Publications: Kota Kinabalu, Sabah, 2003; pp. 1–2. [Google Scholar]
- Kull, T.; Hutchings, M.J. A comparative analysis of decline in the distribution ranges of orchid species in Estonia and the United Kingdom. Biol. Conserv. 2006, 129, 31–39. [Google Scholar] [CrossRef]
- Wagensommer, R.P.; Medagli, P.; Turco, A.; Perrino, E.V. IUCN Red List evaluation of the Orchidaceae endemic to Apulia (Italy) and considerations of the application of the IUCN protocol to rare species. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 90–101. [Google Scholar] [CrossRef]
- Jersáková, J.; Kindlmann, P.; Stříteský, M. Population dynamics of Orchis morio in the Czech Republic under human influence. In Trends and Fluctuations and Underlying Mechanisms in Terrestrial Orchid Populations; Kindlmann, P., Willems, J.H., Whigham, D.F., Eds.; Backhuys Publishers: Leiden, The Netherlands, 2002; pp. 209–224. [Google Scholar]
- Wotavová, K.; Balounová, Z.; Kindlmann, P. Factors affecting persistence of terrestrial orchids in wet meadows and implications for their conservation in a changing agricultural landscape. Biol. Conserv. 2004, 118, 271–279. [Google Scholar] [CrossRef]
- Janečková, P.; Wotavová, K.; Schödelbauerová, I.; Jersáková, J.; Kindlmann, P. Relative effects of management and environmental conditions on performance and survival of population of a terrestrial orchid, Dactylorhiza majalis. Biol. Conserv. 2006, 129, 40–49. [Google Scholar] [CrossRef]
- Štípková, Z.; Kindlmann, P. Extent and reasons for meadows in South Bohemia becoming unsuitable for orchids. Eur. J. Environ. Sci. 2015, 5, 124–147. [Google Scholar] [CrossRef]
- Darwin, C. The Various Contrivances by which Orchids are Fertilized by Orchids; John Murray: London, UK, 1862. [Google Scholar]
- Neiland, M.R.M.; Wilcock, C.C. Fruit set, nectar reward, and rarity in the Orchidaceae. Am. J. Bot. 1998, 85, 1657–1671. [Google Scholar] [CrossRef] [PubMed]
- Charitonidou, M.; Halley, J.M. What goes up must come down—Why high fecundity orchids challenge conserve beliefs. Biol. Conserv. 2020, 252, 108835. [Google Scholar] [CrossRef]
- Kull, T.; Kindlmann, P.; Hutchings, M.; Primack, R. Conservation biology of orchids: Introduction to the special issue. Biol. Conserv. 2006, 129, 1–3. [Google Scholar] [CrossRef]
- Zhang, Z.; Yan, Y.; Tian, Y.; Li, J.; He, J.-S.; Tang, Z. Distribution and conservation of orchid species richness in China. Biol. Conserv. 2015, 181, 64–72. [Google Scholar] [CrossRef]
- Zheleznaya, E. Results of a study of Cypripedium in several regions of Siberia (Russia). Eur. J. Environ. Sci. 2015, 5, 134–141. [Google Scholar] [CrossRef][Green Version]
- Bhattacharyya, P.; Van Staden, J. Ansellia africana (Leopard orchid): A medicinal orchid species with untapped reserves of important biomolecules—A mini review. S. Afr. J. Bot. 2016, 106, 181–185. [Google Scholar] [CrossRef]
- Kolanowska, M.; Baranow, P.; Rykaczewski, M. Elleanthus ortizianus—A new orchid species from southern Colombia. Bot. Lett. 2017, 164, 155–158. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I. Temporal and spatial patterns of orchid species distribution on Greece: Implications for conservation. Biodivers. Conserv. 2020, 29, 3461–3489. [Google Scholar] [CrossRef]
- Tatarenko, I.; Dodd, M.; Wallace, H.; Bellamy, G.; Fleckney, A. Protecting small populations of rare species. Case study on Dactylorhiza viridis (Orchidaceae) in Fancott Woods and Meadows SSSI, Bedfordshire, UK. Nat. Conserv. Res. 2020, 5 (Suppl. 1), 165–171. [Google Scholar] [CrossRef]
- Zizka, A.; Silvestro, D.; Vitt, P.; Knight, T.M. Automated conservation assessment of the orchid family with deep learning. Conserv. Biol. 2020. [Google Scholar] [CrossRef]
- Danihelka, J.; Chrtek, J., Jr.; Kaplan, Z. Checklist of vascular plants of the Czech Republic. Preslia 2012, 84, 647–811. [Google Scholar]
- Grulich, V. Red list of vascular plants of the Czech Republic: 3rd edition. Preslia 2012, 84, 631–645. [Google Scholar]
- Hoffmann, M.; Brooks, T.M.; da Fonseca, G.A.; Gascon, C.; Hawkins, A.F.A.; James, R.E.; Langhammer, P.; Mittermeier, R.A.; Pilgrim, J.D.; Rodrigues, A.S.L.; et al. Conservation planning and the IUCN Red List. Endanger. Species Res. 2008, 6, 113–125. [Google Scholar] [CrossRef]
- Zamin, T.; Baillie, J.E.M.; Miller, M.; Rodriguez, J.; Ardid, A.; Collen, B. National Red Listing beyond the 2010 Target. Conserv. Biol. 2010, 24, 1012–1020. [Google Scholar] [CrossRef]
- Corlett, R.T. Safeguarding our future by protecting biodiversity. Plant Divers. 2020, 42, 221–228. [Google Scholar] [CrossRef] [PubMed]
- Maes, D.; Isaac, N.J.B.; Harrower, C.A.; Collen, B.; van Strien, A.J.; Roy, D.B. The use of opportunistic data for IUCN Red List assessment. Biol. J. Lin. Soc. 2015, 115, 690–706. [Google Scholar] [CrossRef]
- Sharrock, S.; Jones, M. Conserving Europe´s Threatened Plants: Progress towards Target 8 of the Global Strategy for Plant Conservation; Botanic Gardens Conservation International: Richmond, UK, 2009. [Google Scholar]
- Bachman, S.P.; Field, R.; Reader, T.; Raimondo, D.; Donaldson, J.; Schatz, G.E.; Lughadha, E.N. Progress, challenges and opportunities for Red Listing. Biol. Conserv. 2019, 234, 45–55. [Google Scholar] [CrossRef]
- Grulich, V. The Red List of vascular plants of the Czech Republic. Příroda 2017, 35, 75–132. [Google Scholar]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1km spatial resolution climate surface for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2012. [Google Scholar]
- Climatic Conditions of the Czech Republic. Available online: https://geography.upol.cz/soubory/lide/smolova/GCZ/GCZ_Klima.pdf (accessed on 6 January 2020).
- Štípková, Z.; Tsiftsis, S.; Kindlmann, P. Pollination mechanisms are driving orchid distribution in space. Sci. Rep. 2020, 10, 850. [Google Scholar] [CrossRef]
- Botany.cz. Available online: https://botany.cz/cs/ (accessed on 15 December 2020).
- Průša, D. Orchideje České Republiky; Computer Press: Brno, Czech Republic, 2005. [Google Scholar]
- Jacquemyn, H.; Brys, R.; Hermy, M.; Willems, J. Does nectar reward affect rarity and extinction probabilities of orchid species? As assessment using historical records from Belgium and the Netherlands. Biol. Conserv. 2005, 12, 257–263. [Google Scholar] [CrossRef]
- Vogt-Schilb, H.; Munoz, F.; Richard, F.; Schatz, B. Recent declines and range changes of orchids in Western Europe (France, Belgium and Luxembourg). Biol. Conserv. 2015, 190, 133–141. [Google Scholar] [CrossRef]
- Gustafsson, S.; Sjogren-Gulve, P. Genetic diversity in the rare orchid, Gymnadenia odoratissima and a comparison with the more common congener, G-conopsea. Conserv. Genet. 2002, 3, 225–234. [Google Scholar] [CrossRef]
- Kaplan, Z.; Koutecký, P.; Danihelka, J.; Šumberová, K.; Ducháček, M.; Štěpánková, J.; Ekrt, L.; Grulich, V.; Řepka, R.; Kubát, K.; et al. Distributions of vascular plants in the Czech Republic. Part 6. Preslia 2018, 90, 235–346. [Google Scholar] [CrossRef]
- Bódis, J.; Biró, E.; Nagy, T.; Takács, A.; Sramkó, G.; Bateman, R.M.; Gilian, L.; Illyes, Z.; Tokolyi, J.; Lukacs, B.A.; et al. Biological flora of Central Europe Himantoglossum adriaticum H. Baumann. Perspect. Plant Ecol. 2019, 40, 125461. [Google Scholar] [CrossRef]
- Eccarius, W. Die Orchideengattung Dactylorhiza: Phylogenie, Taxonomie, Morphologie, Biologie, Verbreitung, Ökologie und Hybridisation; Wolfgang Eccarius: Eisenach, Germany, 2016. [Google Scholar]
- Kaplan, Z.; Danihelka, J.; Šumberová, K.; Chrtek, J., Jr.; Rotreklová, O.; Ekrt, L.; Štěpánková, J.; Taraška, V.; Trávníček, B.; Prančl, J.; et al. Distributions of vascular plants in the Czech Republic. Part 5. Preslia 2017, 89, 333–439. [Google Scholar] [CrossRef]
- Tsiftsis, S.; Tsiripidis, I.; Papaioannou, A. Ecology of the orchid Goodyera repens in its southern distribution limits. Plant Biosyst. 2012, 146, 857–866. [Google Scholar] [CrossRef]
- Melnyk, V.I. Geographical distribution, habitats and modern state of populations of Goodyera repens (Orchidaceae) in Ukraine. Ukrayins´kyi Bot. Zhurnal 2015, 72, 364–373. [Google Scholar] [CrossRef]
- Bjørndalen, J.E. Protection of Norwegian orchids—A review of achievements and challenges. Eur. J. Environ. Sci. 2015, 5, 121–133. [Google Scholar] [CrossRef][Green Version]
- Nagy, T.; Notari, K.; Takacs, A.; Malkocs, T.; Tokolyi, J.; Molnar, A.V. Precipitation and timing of flowering in Ghost orchids (Epipogium aphyllum Sw.). Acta Bot. Hung. 2018, 60, 223–230. [Google Scholar] [CrossRef]
- Taylor, L.; Roberts, D.L. Biological Flora of the British Isles: Epipogium aphyllum Sw. J. Ecol. 2011, 99, 878–890. [Google Scholar] [CrossRef]
- Bufková, I. Náprava narušeného vodního režimu rašelinišť v národním parku Šumava. Ochr. Přírody 2013, 2, 17–19. [Google Scholar]
- Nilsson, S. Ophrys apifera, a new orchid for Sweden. Bot. Not. 2014, 147, 1–4. [Google Scholar]
- Pignatti, S. Flora d´Italia. III; Edagricole: Bologna, Italy, 1982. [Google Scholar]
- Jacquemyn, H.; Hutchings, M.J. Biological flora of the British Isles: Spiranthes spiralis (L.) Chevall. J. Ecol. 2010, 98, 1253–1267. [Google Scholar] [CrossRef]
- Feldmann, P.; Prat, D. Conservation recommendations from a large survey of French orchids. Eur. J. Environ. Sci. 2011, 1, 18–27. [Google Scholar] [CrossRef][Green Version]
- Vogt-Schilb, H.; Těšitelová, T.; Kotilínek, M.; Sucháček, P.; Kohout, P.; Jersáková, J. Altered rhizoctonia assemblages in grasslands on ex-arable land support germination of mycorrhizal generalist, not specialist orchids. New Phytol. 2020, 227, 1200–1212. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time Period | Explanation |
|---|---|
| Before 1900 | Very old data from the 19th century |
| 1901–1918 | Prior to the end of the 1st World War and the founding of Czechoslovakia |
| 1919–1938 | During the existence of the “1st” Czechoslovak Republic |
| 1939–1948 | 2nd World War and the period before the Communist Putsch in 1948 |
| 1949–1968 | Czechoslovakia under the Communist regime, before the Prague Spring |
| 1969–1989 | The period after the Soviet invasion and before the Velvet Revolution |
| 1990–2020 | The period after the Velvet Revolution and the free market economy in Czechoslovakia |
| Species | Total Number of Sites | Sites Occupied (%) | Species | Total Number of Sites | Sites Occupied (%) |
|---|---|---|---|---|---|
| Anacamptis coriophora | 21 | 0 | Goodyera repens | 231 | 15 |
| Anacamptis morio | 750 | 49 | Gymnadenia densiflora | 199 | 65 |
| Anacamptis palustris | 159 | 13 | Gymnadenia odoratissima | 15 | 47 |
| Anacamptis pyramidalis | 176 | 40 | Hammarbya paludosa | 42 | 36 |
| Dactylorhiza bohemica | 7 | 71 | Herminium monorchis | 12 | 0 |
| Dactylorhiza curvifolia | 6 | 17 | Himantglossum adriaticum | 24 | 25 |
| Dactylorhiza incarnata | 282 | 68 | Limodorum abortivum | 47 | 49 |
| Dactylorhiza maculata | 355 | 37 | Liparis loeselii | 87 | 53 |
| Dactylorhiza traunsteineri | 28 | 46 | Malaxis monophyllos | 124 | 45 |
| Epipactis futakii | 1 | 100 | Neotinea tridentata | 40 | 30 |
| Epipactis leptochila | 49 | 90 | Neotinea ustulata | 885 | 28 |
| Epipactis moravica | 11 | 100 | Neottia cordata | 350 | 34 |
| Epipactis nordeniorum | 10 | 100 | Ophrys apifera | 72 | 92 |
| Epipactis pontica | 25 | 88 | Ophrys holoserica | 120 | 41 |
| Epipactis pseudopurpurata | 12 | 100 | Ophrys insectifera | 89 | 42 |
| Epipactis talosii | 23 | 96 | Orchis mascula | 1483 | 89 |
| Epipogium aphyllum | 201 | 16 | Spiranthes spiralis | 237 | 6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štípková, Z.; Kindlmann, P. Orchid Extinction over the Last 150 Years in the Czech Republic. Diversity 2021, 13, 78. https://doi.org/10.3390/d13020078
Štípková Z, Kindlmann P. Orchid Extinction over the Last 150 Years in the Czech Republic. Diversity. 2021; 13(2):78. https://doi.org/10.3390/d13020078
Chicago/Turabian StyleŠtípková, Zuzana, and Pavel Kindlmann. 2021. "Orchid Extinction over the Last 150 Years in the Czech Republic" Diversity 13, no. 2: 78. https://doi.org/10.3390/d13020078
APA StyleŠtípková, Z., & Kindlmann, P. (2021). Orchid Extinction over the Last 150 Years in the Czech Republic. Diversity, 13(2), 78. https://doi.org/10.3390/d13020078