Abstract
In this study, we analyze the current state of knowledge on extant Eunicida systematics, morphology, feeding, life history, habitat, ecology, distribution patterns, local diversity and exploitation. Eunicida is an order of Errantia annelids characterized by the presence of ventral mandibles and dorsal maxillae in a ventral muscularized pharynx. The origin of Eunicida dates back to the late Cambrian, and the peaks of jaw morphology diversity and number of families are in the Ordovician. Species richness is heterogeneous among the seven recent families, with more than half of the valid species belonging to the Eunicidae + Onuphidae clade, one of the latest clades to diverge. Eunicidans inhabit soft and hard substrates from intertidal to deep waters in all oceans. The few freshwater species are restricted to Histriobdellidae, a family exclusively commensal/parasite of crustaceans. The reproductive biology, development and ecology of most families are poorly known and the information available suggests low dispersal ability. However, all families have records of widely distributed species. Scrutiny of these wide distributions has often revealed the presence of exotic species or more than one species. The exploration of the deep-sea and of new habitats has led to recent descriptions of new species. Furthermore, the revision of type specimens, the examination of new morphological features and the use of molecular data have revealed hidden biodiversity under unjustified synonyms, poor understanding of morphological features and incomplete descriptions. Molecular studies are still very few or nonexistent for the families Histriobdellidae, Hartmaniellidae, Lumbrineridae and Oenonidae. The integration of new methodologies for morphological and molecular study, along with information on biological and ecological traits appears to be the path to improve the knowledge on the diversity of Eunicida.
1. Introduction
Eunicida was raised to order level relatively recently (Uschacov [1] as Eunicimorpha, Dales [2] as Eunicida) despite being recognized as a taxon much earlier (superfamily Eunicea [3] or family Eunicidae [4]). The taxon identity is supported by the presence of eversible jaws, which comprise dorsal maxillae and ventral mandibles, in a ventral muscularized pharynx. The composition of Eunicida has been relatively stable including seven recent (Dorvilleidae Chamberlin, 1919; Eunicidae Berthold, 1827; Hartmaniellidae Imajima, 1977; Histriobdellidae Vaillant, 1890; Lumbrineridae Schmarda, 1861; Oenonidae Kinberg, 1865; and Onuphidae Kinberg, 1865) (Figure 1 and Figures 2–10) and 17 extinct families [5].
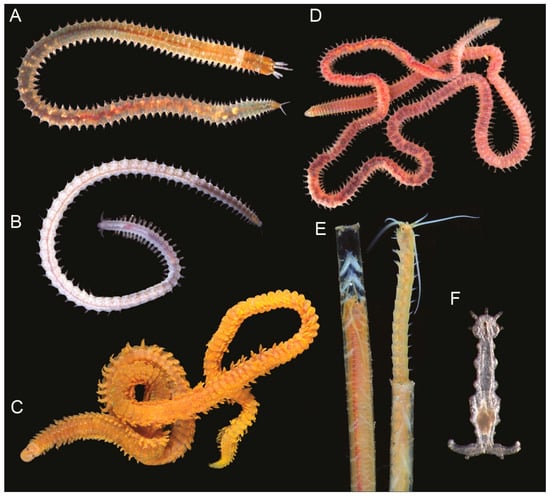
Figure 1.
Live specimens representing eunicidan families: (A) Nicidion sp., Eunicidae, © A. Sememov; (B) Dorvilleidae gen. sp., © A. Sememov; (C) Oenone fulgida (Lamarck, 1818), Oenonidae, © A. Sememov; (D) Lumbrineridae gen. sp., © A. Sememov; (E) Hyalinoecia tubicola (O.F. Müller, 1776), Onuphidae, © N. Budaeva; (F) Histriobdella homari Van Beneden, 1858, Histriobdellidae, © C. Helm.
Eunicida is sister to Phyllodocida within the Errantia clade [6,7]. Phylogenetic analyses using multiple genes or genomic data recover monophyletic Eunicida and families, with exception of Dorvilleidae in analyses including the genus Pettibonea [6,8,9,10,11,12]. Eunicidae and Onuphidae are consistently recovered as sister groups in analyses with only recent taxa and molecular data but formed a polytomy with the extinct genus Esconites Thompson and Johnson, 1977 in analyses with extant and extinct taxa based on morphological data [9]. The relationship among the other families is inconsistent. Oenonidae, Dorvilleidae, Oenonidae + Dorvilleidae or Oenonidae+Lumbrineridae have resulted as sister groups of the Eunicidae + Onuphidae clade. Lumbrineridae and Dorvilleidae have also been recovered as a sister family to all other Eunicida [6,8,9]. The relationships recovered with phylogenomic analyses [6] agree, at least in part, with hypotheses drawn from Orensanz’s [13] and Paxton´s [14] comprehension of the evolution of jaws. All of them recover the clade Eunicidae + Onuphidae and have Dorvilleidae as the sister group to all other Eunicida. These hypotheses contrast in the placement of Lumbrineridae as a sister group to Oenonidae [6,13] or in a clade with Hartmaniellidae, sister group to (Dorvilleidae (Oenonidae (Eunicidae, Onuphidae)) [14]. Orensanz [13] suggests the clade Lumbrineridae+Oenonidae is the sister to (Hartmaniellidae, (Eunicidae, Onuphidae)). Hartmaniellidae and Histriobdellidae have not been included in any formal phylogenetic analyses. Their monophyletic status and placement within Eunicida remain to be tested.
Species richness is heterogeneous among families. Eunicidae (12 genera, 453 spp.) bears the highest number of species followed by Onuphidae (22 genera, 340 spp.), Lumbrineridae (19 genera, 279 spp.), Dorvilleidae (32 genera, 200 spp.), Oenonidae (12 genera, 109 spp.), Histriobdellidae (3 genera, 13 spp.) and Hartmaniellidae (1 genus, 3 spp.) [15]. Despite being among one of the latest clades to diverge, the Eunicidae + Onuphidae clade comprises more than half of the recent species within Eunicida (1397 spp.). This could indicate a higher diversification rate, a lower extinction rate or just an underestimation of species diversity in other families, such as Dorvilleidae, for which 73% of the valid species were described in the last 50 years (Figure 2). In the families Eunicidae and Onuphidae, the species described in the last 50 years correspond, respectively, to 30% and 53% of the total of valid species. Many descriptions of new species in Eunicida are due to the discovery of new habitats and increase in exploration of the deep-sea (e.g., [16,17,18,19]) as well as the study of meiofaunal polychaetes (e.g., [20,21,22,23]). However, the revision of type specimens, the examination of new morphological features and the use of molecular data are revealing a hidden biodiversity under unjustified synonyms, poor understanding of morphological features and incomplete descriptions [24,25,26]. The lack of correspondence between the number of genera and the number of species per family (e.g., Eunicidae) illustrates more homogeneity in the external morphology of some families or just more conservative taxonomic decisions.
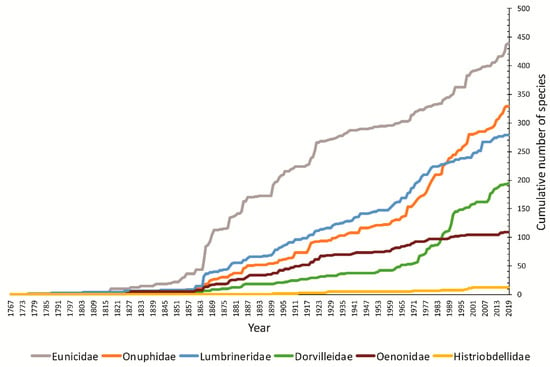
Figure 2.
Cumulative number of species described in six eunicidan families, data on valid species only [15] (Hartmaniellidae not shown).
The origin and diversification of Eunicida date back to the late Cambrian, the age of the earliest fossil maxilla [27]. Maxillae compose the main fossil record of Eunicida and consist of a series of mostly paired sclerotized plates. Maxillae vary among families in relation to the number, morphology, symmetry and arrangement of the plates, and, when present, the shape of the carriers (Figure 3). The diversity of jaw morphology, which translates into the number of families, is more than twice as high in extinct than in extant taxa and is peaked in the Ordovician [27]. The general morphological patterns of maxillae are grouped in jaw-types fine-tuned along different studies considering both extinct and extant diversity [3,13,14,28,29]. Of the six jaw-types currently considered [14], two are exclusive of ten extinct families and all the other four jaw-types are shared among extinct and extant families [14]. Dorvilleidae is the recent family with the highest jaw diversity, motivating the proposition of its division into two different families (Dorvilleidae and Iphitimidae) [13,14,19] or into six informal groups [19], none supported by the only phylogenetic analyses including representatives of the dorvilleid jaw diversity [30].
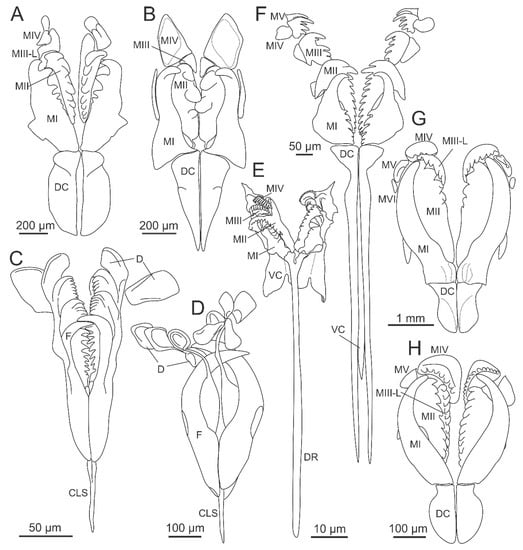
Figure 3.
Diversity of maxillae in Eunicida. (A) Hartmaniella sp., Hartmaniellidae; (B) Augeneria sp., Lumbrineridae; (C) Ophryotrocha mammillata Ravara et al., 2015, Dorvilleidae, P-type maxillae; (D) the same, K-type maxillae (E) Histriobdella homari Van Beneden, 1858, Histriobdellidae; (F) Arabella mutans (Chamberlin, 1919), Oenonidae; (G) Eunice roussaei Quatrefages, 1866, Eunicidae; (H) Paradiopatra fragosa (Ehlers, 1887), Onuphidae. CLS—carrier-like structure, D—free denticles, DC—dorsal carrier, DR—dorsal rod, F—forceps, MIII-L—left maxillae III, MI–MV—maxillae I–V, VC—ventral carrier. (C–D) modified from Ravara et al. [31]; (H) modified from Budaeva and Fauchald [32].
The different jaw morphologies also have different biomechanics with implications for food capturing and shredding [33]. In maxillae bearing carriers of at least Lumbrineridae and Onuphidae (Figure 3B,H), those plates attached to the carriers are primary maxillae (maxillae I, II in both families and unpaired III in Onuphidae) responsible for catching and shredding (exclusive of serrated plates) food items. The unpaired serrated primary maxillae III in onuphid improves the breaking of food with different properties. The interlocking mechanism, present at the posterior end of maxillae I in both families, prevents slipping of the plates and it is an important mechanism in capturing moving food [33]. Feeding habits and food quality consumed are variable within recent and, probably, extinct Eunicida, including grazing, deposit-feeding, and predation, detritivory, herbivory, carnivory and omnivory [34]. Giant raptorial eunicidans were already present 400 million years ago [35].
Recent giant eunicidan species are among the longest annelids with up to 6 m in length (Eunicidae, Eunice aphroditois (Pallas, 1788) [36]). Length variation is great within Eunicida, since it also includes species among the shortest annelids with 255 μm (Dorvilleidae, Neotenotrocha sterreri Eibye-Jacobsen and Kristensen, 1994 [30]). The prostomium is well developed followed by a double (Dorvilleidae, Eunicidae, Hartmaniellidae, Lumbrineridae, and Oenonidae) or single (Onuphidae) ringed peristomium in all families, except for Histriobdellidae in which these structures are fused. Prostomial appendages, one to three antennae and a pair of palps, may be present on the prostomium of Dorvilleidae, Eunicidae, Histriobdellidae and Onuphidae (Figures 4, 5, 7 and 10). Lumbrineridae and Oenonidae generally have conical appendage-free prostomium with the exception of few genera bearing tiny dorsal appendages covered by the anterior fold of the peristomium or visible due to a peristomium dorsal incision (Figures 8 and 9). Buccal lips are conspicuous structures in the prostomium of Onuphidae (Figure 10E) and insconspicuous in Dorvilleidae, Eunicidae (Figure 5), Lumbrineridae, and Oenonidae [37]. The muscularized pharynx holding the maxillae contains at least homologous dorsolateral and pharyngeal folds in Dorvilleidae, Eunicidae, Lumbrineridae, Oenonidae and Onuphidae, and a dorsolateral fold anterior extension in the last four families [37,38]. Metamerism is conspicuous and the number of chaetigers is variable in all families but Histriobdellidae. Parapodia are reduced, uniramous or subbiramous with notopodia reduced to dorsal (notopodial) cirri. Capillary chaetae are present in all families, except Histriobdellidae, which lacks chaetae. Chaetae diversity increases along the evolution of Eunicida being highest in Onuphidae and Eunicidae [39]. The pygidium and two or four pygidial (anal) cirri, which may be absent, bear several nerve cell endings suggesting they have sensory function [40].
Eunicidans are mostly free-living and can be found in the majority of marine habitats in soft and hard substrates from the intertidal zone to the deep waters in all oceans. Hartmaniellidae is the only family that appears to be solely present in fine soft bottoms, sometimes with shell fragments. No other free-living family is exclusively present in one type of habitat or substrate. However, most members of some families, such as Lumbrineridae, Oenonidae and Onuphidae, prefer soft bottoms, while Eunicidae is most common in hard biogenic substrates [41,42]. Histriobdellidae is the only eunicidan family occurring in both marine and continental waters, with highest richness in the latter, and being exclusively commensals/parasites of crustaceans. Parasitic and symbiotic species are also present in the families Dorvilleidae, Eunicidae, and Oenonidae.
All families have records of widely distributed species, which deserve further investigation. Most widespread species studied in detail ended up as more than one species [43,44,45] or exotic species dispersed by human mediated transport [46,47]. However, some species appear to have naturally widespread distributions [48,49] despite the poor natural dispersal capability attributed to eunicidan species. Reproductive modes vary from viviparity, brooding of young in tubes or gelatinous egg masses to broadcast spawning. However, even in the latter the planktonic life span is expected to be relatively short, since all known eunicidan larvae are lecitotrophic [50]. Asexual reproduction happens at least by scissiparity (Oenonidae, [51]) and architomy (Dorvilleidae, [52]).
Eunicida includes species of economic and cultural importance. Eunicidae, Lumbrineridae, Oenonidae and Onuphidae comprise species used as baits for commercial and recreational fishing in diverse localities in Atlantic, Pacific, Adriatic, Mediterranean and Suez Canal coasts [53,54,55,56,57,58], and are also farmed to be used as bait and food supply in aquaculture (e.g., Diopatra aciculata Knox and Cameron, 1971, Onuphidae; [59]). Ripe swarming forms of Palola (Eunicidae) species are a traditional food and the source of festivals in South Pacific communities [49,60].
In this review, we analyze the current state of knowledge on extant Eunicida systematics, morphology, feeding, life history, habitat, ecology, distribution patterns, local diversity and exploitation, pointing out gaps of knowledge and future perspectives. The great diversity present in Eunicida prevents the presentation of a complete overview without considering each family separately. Thus, in the next sections, we present the current state of knowledge on the different aspects of the recent eunicidan families. Extinct families are not further considered because of the lack of information on most of the aspects considered here.
2. Dorvilleidae Chamberlin, 1919
2.1. Systematics and Phylogeny
The family Dorvilleidae comprises 32 genera and around 200 species. It includes the majority of the smaller forms of the eunicemorph polychaetes, with around 10% of the species described hitherto belonging to the interstitial fauna. Although the family name Dorvilleidae has been in common use since it was erected by Chamberlin [61], its validity remained questionable as its type genus name Dorvillea Parfitt, 1866 is a junior homonym of the mollusk genus name Dorvillea Leach in Gray, 1852. The alternative name Stauronereididae Verrill, 1900 was occasionally used (e.g., [62]), regardless the previous synonymy of its type genus Stauronereis Verrill, 1900 with the priority name Dorvillea [61]. Only recently, the genus name Dorvillea Parfitt, 1866 was validated as nomen protectum, based on its use in over 25 publications by more than 10 authors in the last 50 years (Article 23.9.2 of the ICZN code), while Dorvillea Leach, 1852 was deemed as nomen oblitum, since it has not been used as valid since 1899 [63]. This nomenclatural act also protects the family name Dorvilleidae Chamberlin, 1919.
The first dorvilleid species was described by Delle Chiaje [64] as Nereis rudolphi (posteriorly moved to the genus Schistomeringos Jumars, 1974). When Chamberlin erected the family, 21 of the currently valid dorvilleid species were already described. Two genera were included in the family: Dorvillea and Ophryotrocha Claparéde and Mecznikow, 1869. Although already described, the genus Iphitime Marenzeller, 1902 was initially ascribed to a different family (first Lysaretidae Kinberg, 1865 and later Iphitimidae Fauchald, 1970, both currently unaccepted). Not until about four decades later were other genera described for the family. The remaining 29 genera were described following important revisions [13,19,21,30,65,66,67,68], among other works. An updated key and detailed diagnoses to all dorvilleid genera is available in Wiklund et al. [69]. Over two-thirds of the dorvilleid genera include only one or a couple of species, while the largest genera, Ophryotrocha, Dorvillea and Schistomeringos, encompass more than half of the existing species.
Most molecular studies within the family have dealt mainly with Ophryotrocha [70,71], often in combination with species descriptions (e.g., [31,72,73,74]). In several of these restricted molecular analyses, species from different dorvilleid genera fall within Ophryotrocha. This is the case for Exallopus Jumars, 1974, Iphitime and Pseudophryotrocha Hilbig and Blake, 1991 (e.g., [31,72,73,74]). However, since the respective type species for these genera have not been sequenced yet, the authors have chosen to await a formal family revision rather than renaming and moving only those species that have been used in those studies. The genus Mammiphitime Orensanz, 1990 might also belong to Ophryotrocha although no molecular data is available for it yet. The erection of Mammiphitime was justified by the large dorsal lobes in the median and posterior segments [13], but since then, another species with similar lobes has been described, Ophryotrocha mammillata Ravara, Marçal, Wiklund and Hilário, 2015, and molecular data placed it within Ophryotrocha [31]. Due to these, and possibly other doubtful genera, the family Dorvilleidae is in need of a comprehensive revision using a combination of morphological characteristics and molecular data.
The mono- or paraphyletic condition of the family Dorvilleidae has been a matter of discussion depending on whether some taxa such as Dinophilidae Macalister, 1876, Diurodrilidae Kristensen and Niilonen, 1982 and Pettiboneia Orensanz, 1973 are included or not. In their phylogenetic study using morphological characteristics, Eibye-Jacobsen and Kristensen [30] refer to Iphitimidae and Dinophilidae as belonging to Dorvilleidae. However, subsequent studies using molecular methods or ultrastructure analyses have suggested that although Iphitimidae does indeed fall within Dorvilleidae, Dinophilidae and Diurodrilidae are not closely related to Dorvilleidae (e.g., [6,75,76,77]). The monophyly of Dorvilleidae is achieved when excluding the genus Pettiboneia [6,8,39]. In fact, Paxton [14] reported Pettibonea as lacking base dental plates and having two rows of free denticles, corroborating the unique morphology of the genus. However, in the molecular analysis performed by Struck et al. [8] the genus was represented only by two genetic markers (18S and 16S rDNA) of a single species, Pettiboneia urcinensis Campoy and San Martín, 1980, that was recovered as genetically closer to Lumbrineridae. Furthermore, there are very few sequences available for species in the other dorvilleid genera preventing a large-scale family analysis. Thus, phylogenetic analyses with better taxon coverage are required to test the monophyly of the family.
2.2. Morphology
Dorvilleids share with the other eunicidan families the outstanding jaw apparatus. It is mainly the jaw morphology that allows identifying the family and is also useful for delineation of genera and species. The jaws in Dorvilleidae consist of ctenognath maxillae (upper comb-like jaws), formed by two to four rows of symmetrical or subsymmetrical dentate elements and a posterior unpaired element (carrier-like structure) (Figure 3C,D), and mandibles (lower jaws). However, species in some genera lack jaws, e.g., Ikosipodoides Westheide, 2000 and Parapodrilus Westheide, 1965. Furthermore, the shape of the prostomium and its appendages as well as the shape of parapodia and chaetae is of importance to separate genera and species within the family (see [69]). Prostomial appendages, when present, are paired. Lateral antennae can be articulated or unarticulated while palps are simple or biarticulated (Figure 4A–D). Parapodia are missing, sub-biramous or uniramous, with or without dorsal and ventral cirri (Figure 4E). Some genera lack chaetae, but most species have simple supra-acicular and simple and/or compound subacicular chaetae (Figure 4F). A detailed description of the external and internal morphology of Dorvilleidae is available in Wiklund et al. [69]. Due to their small size and frequently minor morphological differences, dorvilleids can be difficult to identify correctly, and different species are often only detected following molecular analysis (e.g., [44]).
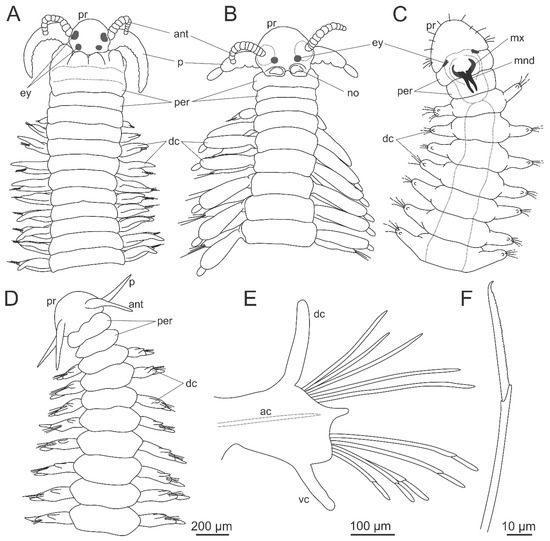
Figure 4.
Morphology of Dorvilleidae. (A), (B) Dorvilleidade gen. sp. anterior end, dorsal view; (C) Ophryotrocha sp. Anterior end, dorsal view; (D) Ophryotrocha langstrumpae Wiklund, Altamira, Glover, Smith, Baco and Dahlgren, 2012, anterior end, dorsal view; (E) the same, midbody parapodium; (F) the same, compound chaeta. ac—acicula, ant—antenna, dc—dorsal cirrus, ey—eye, mnd—mandibles, mx—maxillae, no—nuchal organ, p—palp, per—peristomium, pr—prostomium, vc—ventral cirrus. (D–F) modified from Wiklund et al. [74].
2.3. Feeding and Life History
The feeding habits vary within the family, with some species being reported as carnivores and others as omnivorous, grazing on algae or bacterial mats or living off detritus [34,78].
In most dorvilleid genera, the sexes are separated and the species lack sexual dimorphism. However, in the genus Ophryotrocha, some species exhibit sexual dimorphism and many species are consecutive or simultaneous hermaphrodites (e.g., [79,80]). Epitokal modifications may occur in some species, expressed by enlarged eyes and longer chaetae [81,82,83]. Some dorvilleid species are broadcast spawners with swarming phases [83], other species have pseudocopulation and possibly internal fertilization [84], and a few species have been reported to have asexual reproduction [52]. In Ophryotrocha, pseudocopulation and brooding of the offspring is common, and viviparity occurs in a few cases [79,84]. While some dorvilleids, e.g., Schistomeringos rudolphi (delle Chiaje, 1828), have planktonic larvae that can spend at least a week in the water column before settling [83], others brood their young, expressing direct development with the juveniles emerging from egg cases already resembling miniature adult worms [79].
2.4. Habitats and Ecology
Some of the smallest dorvilleid species are interstitial, while larger species are found on soft or hard substrates, and some are symbiotic/parasitic in e.g., crab gill chambers (species in Iphitime and two species in Ophryotrocha) or in the coelom of other polychaetes (Veneriserva Rossi, 1984). Free-living species can surround themselves with mucus, and some species construct tubes out of mucus [34].
Several studies indicate limited tolerance of some dorvilleid species to environmental factors such as temperature and salinity (e.g., [85,86]) although there are also records of euryhaline species [87,88]. Nevertheless, apart from a few records of estuarine occurrence, dorvilleids seem to be predominantly fully marine organisms. Species of the genera Ophryotrocha and Parougia Wolf, 1986 are particularly common and frequently highly abundant in organically enriched habitats, such as harbors, fish farms, sewage discharges and organic-falls (wood or the carcasses of large animals sunken to the seafloor) [16,31,44,72,73,74,89,90,91,92], being considered by some authors as nonspecialized opportunists (e.g., [48]). These two genera, along with Exallopus, include the majority of the species occurring in deeper waters [19,67,93,94,95,96,97], often in extreme habitats such as hydrothermal vents and cold seeps [18,97,98,99,100,101,102].
2.5. Distribution Patterns and Diversity
Dorvilleid worms are widely distributed and occur from the intertidal zone to the abyss. Significant family and genera revisions covering regional geographic areas include Fauchald [103], Orensanz [13,66], Oug [81], Wolf [68,104,105], Hilbig [95,106], Paiva and Nonato [107], Imajima [108], Núñez et al. [109] and Núñez [62] (as Stauronereididae). However, the majority of these works focus mainly on the Northern Hemisphere benthic fauna, while southern geographic areas such as the South Atlantic, Southeast Pacific and Indian Ocean are very poorly documented.
The exploration of organically enriched areas led to a great increase in the number of Ophryotrocha and Parougia species (e.g., [16,18,31,44,72,73,74,92,96,97,98,99,100,101,102,110]). The occurrence of several sympatric congeners is also frequently reported for both genera (e.g., [16,18,31,44,72,73,74,111,112,113]). Furthermore, with the upswing in deep-sea exploration, more species of these and other genera have been described also from areas that seemingly were not organically enriched (e.g., [19,93,94,114,115,116]). It is likely that there are many more species from these and other dorvilleid genera awaiting discovery, some still unknown in the ocean while others are already collected and stored, but not properly examined and described yet.
Despite the lack of planktonic larvae, some Ophryotrocha species have been shown to have quite a broad distribution range. For instance, Ophryotrocha orensanzi Taboada, Wiklund, Glover, Dahlgren, Cristobo and Ávila, 2013, Ophryotrocha scutellus Wiklund, Glover and Dahlgren, 2009 and Ophryotrocha cyclops Salvo, Wiklund, Dufour, Hamoutene, Pohle and Worsaae, 2014 have been reported from sites with hundreds to thousands kilometers between them, with species identities confirmed by molecular data [31,48,72,90]. Some species with wide distribution ranges are suspected to have been transferred with human activities such as in ballast waters (e.g., [78]). However, many dorvilleids are small and can be difficult to identify properly to species level, and thus the possibility of them having a continuous distribution between distant localities cannot be ruled out. More rigorous taxonomic investigations, preferably in combination with molecular information, are needed before we can make any assumptions about dorvilleid dispersal capacities and species real distribution.
2.6. Exploitation
Given their small size, Dorvilleids have never been an important commercial group. However, a recent study suggested the beneficial use of Ophryotrocha species in pisciculture by feeding on fish feces and in turn being fed to the fish [117]. In addition, because they have a short generation time and are easily kept in laboratory conditions, Ophryotrocha species may be suitable candidates as test model organisms for various kinds of studies [70,79,87,118].
3. Eunicidae Berthold, 1827
3.1. Systematics and Phylogeny
The family Eunicidae comprises 12 genera and approximately 453 species. It was first recognized as a taxon by Lamarck [119] under the name of Eunicea. However, until 1944 it was more inclusive and almost equivalent to the order Eunicida with variations depending on the author. The current definition of the family was established in Hartman [28] including four of the current valid genera, Eunice Cuvier, 1817 (242 species), Lysidice Lamarck, 1818 (30 species), Marphysa Quatrefages, 1865 (74 species), Palola Gray in Stair, 1847 (14 species). The composition of the family has been stable since then. However, currently it includes eight additional genera, one extinct, Esconites Thompson and Johnson, 1977 (one species), and seven extant, Aciculomarphysa Hartmann-Schröder, 1998 in Hartmann-Schröder and Zibrowius [120] (one species), Euniphysa Wesenberg-Lund, 1949 (11 species), Fauchaldius Carrera-Parra and Salazar-Vallejo, 1998 (two species), Leodice Lamarck, 1818 (33 species), Nicidion Kinberg, 1865 (14 species), Paucibranchia Molina-Acevedo, 2018 (19 species), Treadwellphysa Molina-Acevedo and Carrera-Parra, 2017 (eight species).
Phylogenetic analyses based solely on morphological data could not recover monophyletic Eunicidae and Eunice [121]. Combined molecular and morphological phylogenetic analyses recovered a monophyletic Eunicidae and nonmonophyletic Eunice, Lysidice, and Marphysa [10,11]. Based on these results, Leodice and Nicidion were resurrected to include species previously classified as Eunice or Marphysa that made both nonmonophyletic and Nematonereis Schmarda, 1861 was synonymized to Lysidice. It was still not possible to delimit a monophyletic Eunice because of the inconsistent placement of several of its species, including the type species Eunice aphroditois. Eunice species, which bear branchiae starting late, inflated base of the ventral cirri as thick ventral ridges, left MxIV with teeth restricted to the dorsal portion of the plate, and bidentate compound falcigerous chaetae with inconspicuous teeth in anteriormost chaetigers, consistently formed a clade which may represent another monophyletic genus. The further division of Leodice in two monophyletic taxa may also be justified in future analyses including more species. The genus has two main sister clades, one comprising species with dark subacicular hooks and the other species with light subacicular hooks. Thus, further analyses including more representatives of the diversity and other molecular markers are necessary.
A taxonomic key for the identification of genera valid at the time and considering the definitions based on eunicid current phylogenetic hypothesis is provided in Zanol et al. [11]. Marphysa is currently more restricted because some species have been transferred to Paucibranchia and Treadwellphysa. The key to genera in Molina-Acevedo and Carrera-Parra [122] includes Treadwellphysa. However, it considers Nematonereis valid and has more restricted morphological definitions for Eunice, Leodice, Lysidice, Nicidion, and Marphysa, not following genera definitions based on the phylogenetic hypothesis in Zanol et al. [11].
Taxonomic revisions for species are available for Eunice [36], Euniphysa [123], Lysidice from the Western Caribbean [124], Marphysa [25,122,125,126], Palola [127], Paucibranchia [128], and Treadwellphysa [129]. These revisions comprise more detailed descriptions, discussions on taxonomic acts and taxonomic keys, improving the knowledge of diagnostic morphological features, undoing unjustified synonymizations and facilitating identification. The revision of Eunice also includes species of Leodice and Nicidion, which were classified as Eunice at the time. Some of these have not had their genus officially changed yet. Thus, the number of Leodice and Nicidion species is probably much higher than the current figures and that of Eunice is much lower. Palola diversity is discussed in Schulze [49] and Schulze and Timm [116]. Despite the revisions, the identification of species of the genera Eunice, Leodice, Lysidice, Nicidion and Palola may still be confusing, thus, further revisions including more detailed descriptions with intraspecific variation would be of great value.
3.2. Morphology
Eunicidae is defined by two synapomorphies, dorsal buccal lip fused to the dorsal side of the prostomium and dorsolateral fold anterior extensions medially connected [11] (Figure 5A–E). Additional diagnostic features are present at prostomium, peristomium and jaws. These are ventral buccal lips with median transverse groove, one (single median antenna) to five prostomial appendages (a pair of palps, a pair of lateral antennae and a single median antenna), double ringed peristomium (Figure 1A and Figure 5A–E), and asymmetric eulabidognath maxillae (Figure 3G). The family encompasses great length variation, bearing species from few millimeters to few meters long. All species have a variable number of chaetigers.
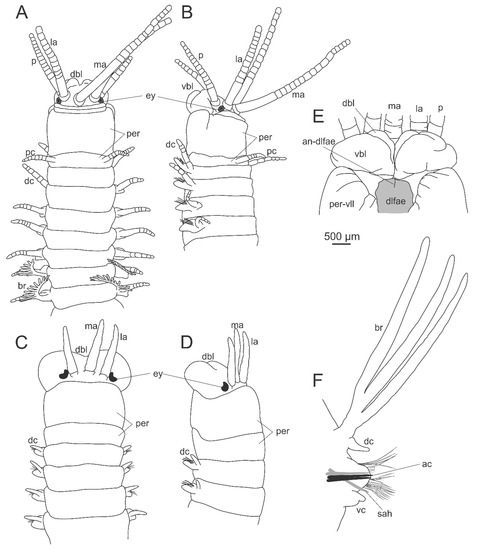
Figure 5.
Morphology of Eunicidae. (A) Leodice sp., anterior end, dorsal view; (B) the same, lateral view; (C) Lysidice sp., anterior end, dorsal view; (D) the same, lateral view; (E) Eunice violaceomaculata Ehlers, 1887, prostomium, ventral view, dissected; (F) Marphysa sp., midbody parapodium. ac—acicula, an-dlfae—anterior notch of the dorsolateral fold anterior extension, br—branchia, dbl—dorsal buccal lip, dc—dorsal cirrus, dlfae—dorsolateral fold anterior extension, la—lateral antenna, ma—median antenna, p—palp, pc—peristomial cirrus, per—peristomium, per-vll—peristomial ventrolateral lips, sah—subacicular hook, vbl—ventral buccal lip, vc—ventral cirrus.
Diagnoses of extant genera take into account presence of lateral antennae and palps, articulation of prostomial appendages, presence of peristomial cirri, flat or curved mandibles, shape of maxillary plates, shape and distribution of branchiae, and presence and shape of pectinate chaetae and subacicular hooks (Figure 5F). Features present in chaetigers may vary in distribution and shape along the body. The complete set of chaetae in most genera includes limbate, pectinate, compound, subacicular hooks and aciculae. Palola is the only genus missing pectinate chaetae and subacicular hooks. In most genera, pectinate chaetae shafts are always thinner than limbate chaetae from the same parapodium (thin pectinate). However, in Marphysa and Treadwellphysa, they may have thick shafts (thicker than limbate chaetae in the same parapodium) and wide blades in mid and posterior chaetigers (thick and wide pectinate chaetae). The outer teeth of pectinate chaetae may be: both as long as inner teeth (anodont), just one longer (heterodont) or both longer than inner teeth (isodont sensu [130]).
In addition to these, features considered in the identification of species are the shape and size of prostomium, peristomium, prostomial appendages and peristomial cirri, dorsal and ventral cirri, prechaetal, chaetal, postchaetal lobes, pygidial cirri, the distribution and shape of branchiae, pectinate and compound chaetae, and the color, distribution, number and shape of aciculae and subacicular hooks. The detailed morphology of the chaetae varies along the body and is an important taxonomic feature, as in the case of pectinate chaetae in Marphysa species [131,132]. Compound chaetae are present in all species at least in part of the body with the exception of some species of Marphysa, which lack them completely. Only some species of Euniphysa bear pseudocompound chaetae in the anterior chaetigers.
Challenges in understanding the morphology of species are the difficulty in sampling complete specimens and the poor knowledge on intraspecific variation, known only for few species (e.g., Leodice rubra (Grube, 1856) in [133]; Leodice vittata (Chiaje, 1929) in [36,134]). Intraspecific variation, sometimes due to size variation, occurs at least in features such as the number of articles in the prostomial appendages, the presence/absence and distribution of subacicular hooks, the number of branchial filaments, the branchial distribution, and the end of inflated base of ventral cirri (e.g., [47,134,135,136,137]). In incomplete specimens, size is commonly estimated as length through chaetiger 10, and width at this chaetiger, however, at least in a few species the peristomium width is a better estimator [138]. An index of the product of the length through chaetiger 10 by the largest width at this region has also been used a size estimator [13]. Detailed descriptions of general morphological features are available in Ehlers [3], Treadwell [139], Fauchald [36], Zanol et al. ([11] in supplementary material, [121]), Carrera-Parra [140], Zanol and Budaeva [141], of branchial distribution patterns in Miura [135] and of maxillary features in Paxton [14], Molina-Acevedo and Carrera-Parra [122] and Molina-Acevedo [128].
3.3. Feeding and Life History
Eunicids have been reported as macrophagous or microphagous omnivorous, however, some species have more restricted diets, being herbivorous or carnivorous [34]. At least one Eunice species is an ambush predator. It captures the preys with the maxillae, which are kept open near the substrate while waiting for the prey [142].
All the species studied have separate sexes and sexual reproduction. Sexual dimorphism of unripe individuals, hermaphroditism and asexual reproduction have not been described for the family. The reproduction biology is known for less than 10% of the species [141]. Thus, it might be more variable than our current knowledge. Free spawning by benthic or epitokous forms followed by fertilization and larval development in the water column is the most common mode of reproduction. Some epitokous forms swarm. Palola swarmings in the South Pacific are the most well-known because they have cultural importance to local communities [60,143,144]. Exceptions to fertilizations in the water column happen in some Leodice, which fertilize eggs within their burrows, and estuarine Marphysa species, which produce jelly egg masses attached to the substrate [145,146,147,148,149,150,151].
All clearly described larvae are lecitotrophic and oocyte diameters also suggest lecitotrophic development [50]. Juvenile settlement may take 10–20 days from the beginning of the development [149,150,151,152], and eunicids are expected to have poor natural dispersal capabilities. However, they may disperse via rafting in drift plastic and wood (e.g., Eunice and Marphysa; [153,154]) and in association with oysters in human-mediated transport (e.g., Marphysa victori Lavesque, Daffe, Bonifácio and Hutchings, 2017; [47]). Commercial oysters can also host species of Leodice, Lysidice, Nicidion and Marphysa as epibionts [155].
3.4. Habitats and Ecology
Species of eunicids are present in soft and hard substrates in estuarine and marine habitats. They also occur associated with biogenic substrates, such as coralline algae, cnidarians, oyster shells, seagrass and sponges [139,155,156,157,158], and authigenic carbonate crusts formed in cold seeps [159]. Species live in burrows or tubes; some crawl outside, usually at night. Tube building appears to be restricted to some Eunice and Leodice species. They are most commonly composed of parchment-like material—soft or stiff. Some tube building species are symbionts with cnidarian species such as Lophelia pertusa (Linnaeus, 1758), Madrepora oculata Linnaeus, 1758 and Epizoanthus spp [160,161,162]. Species associated with corals act in their bioerosion, in assembling coral reefs and in determining the reef framework through tube building, which facilitates coral growth [162,163,164]. The mean density of eunicids in dead coral may be as high as 315 ind/m2, while a single dominant species such as Nicidion cariboea (Grube, 1856) may reach a density of 1219 ind/m2 [165]
3.5. Distribution Patterns and Diversity
Eunicids inhabit all oceans from the intertidal zone to around 3500 m in the deep-sea, being more common and diverse in shallow tropical and subtropical hard substrates. In the Antarctic and Artic regions, a maximum of four species has been recorded and only Eunice and Leodice are present [13,166,167]. Most genera are widespread; the exceptions are Aciculomarphysa, Fauchaldius and Euniphysa. Aciculomarphysa and Fauchaldius have not been recorded since their original description, thus their distribution is restricted to the depths they have been described from, respectively, 460–490 m and 150–250 m [120,130,168]. Euniphysa is mainly restricted to the shallow warm waters of the Northern Hemisphere, with most of the richness concentrated in the South China Sea and adjacencies [123]. The only known records for the South Atlantic Ocean are from the south and northeast of Brazil ([169], personal observation), which suggests that Euniphysa is widespread but not abundant.
Some local fauna studies focusing on eunicid taxa are available for the Antarctic and Subantarctic Seas [13], Argentina [170], Australia [24,131,171,172,173,174,175], Brazil [133,176,177,178,179,180], British Isles [181], the Canary Islands [182], the Caribbean Sea [25,42,122,124,130,136,139,183,184,185,186], China [187,188], the eastern Pacific [28], the El Salvador Pacific Coast [189], Fiji and Samoa [190], the French Atlantic Coast [191], the Gulf of Mexico [192,193], Hong Kong [194], the Iberian Peninsula [132,195], India [196,197], Japan [135,198,199,200,201,202], Korea [203], Malaysia [204], the Mediterranean Sea [205,206,207,208,209,210], the Mexican Pacific Coast [103,211,212], New Zealand [213], the Philippines [214,215], the Scandinavian and North Sea Coasts [167], South Africa [126,216,217], Taiwan [218,219], the USA’s Atlantic Coast [148,220,221], the USA’s Pacific Coast [222], and Venezuela [223,224]. Carrera-Parra [140] includes a taxonomic key for species registered to the warm temperate Northeast Pacific, Tropical Eastern Pacific Ocean and Tropical Northwestern Atlantic Ocean. The eunicid fauna are most poorly known for the Southeast Atlantic, Southeast Pacific and Indian Ocean.
These studies on local fauna are initial guides to the eunicid diversity. However, some of the species included should be considered with care. Several species thought to be widespread at the time of publications are currently known [e.g., Lysidice ninetta Audouin and Milne Edwards, 1833a [124,225], Marphysa sanguinea (Montagu, 1813) [45], Palola viridis Gray in Stair, 1847 [116]] or suspected to have restricted distribution (e.g., Eunice afra Peters, 1854, Eunice aphroditois, Leodice antennata Lamarck, 1818, Leodice pennata (Muller, 1776), Leodice vittata, Lysidice collaris Grube, 1870, Lysidice hebes (Verrill, 1900), Lysidice unicornis (Grube, 1870), Nicidion cariboea, Palola siciliensis (Grube, 1840), Paucibranchia bellii (Audouin and Milne Edwards, 1833)). Eunice aphroditois had several large (giant) species considered to be its junior synonyms, leading to the identification of most giant specimens as E. aphroditois [184]. The identity of most large specimens remains to be solved, as well as the diversity and evolution of giant species in the family [184,226].
The widespread status of most eunicid species is usually due to poor original descriptions, unjustified generalized synonymizations, and the use of local fauna monographs in the identification of the fauna of distinct regions as described for other polychaete groups [227]. Palola sp. (in [49,116,174]) and Marphysa victori are the only species with widespread distribution confirmed by both molecular and morphological data. Palola sp. is present in both Micronesia and the eastern Pacific and its means of dispersion are not fully understood [49,116,174]. Meanwhile, M. victori is an exotic species in France introduced in association with commercial oysters imported from Japan [47].
Taxonomic revisions of species based on type material (e.g., [36,123,127,128,129]) as well as designations of neotypes (e.g., M. sanguinea in [45]) provided the initial framework to improve the knowledge on the diversity (Figure 2) and distribution of eunicid species and to question records of species reported all over the world. Furthermore, more detailed examination of morphological features, such as jaws (e.g., [25,128]), chaetae variation along the body (e.g., [24,131]), and molecular data improved findings of misidentifications and morphologically similar species. These provide tools and encourage other authors to handle similar taxonomic issues generating a virtuous cycle as observed in Marphysa, eunicid genus with the highest number of species described since 2003 when the M. sanguinea neotype was designated in Hutchings and Karageorgopoulos [45].
Lewis and Karageorgopoulos [217] were the first to follow the lead to tackle the widespread issue in Marphysa sanguinea combining morphological and molecular data. They revealed high molecular diversity among species traditionally identified as M. sanguinea. The same pattern has also been found in Palola and Lysidice. Palola includes several deeply separated molecular lineages of mostly morphologically indistinct species and Palola viridis appears to have more restricted distribution than its records in the literature [49,116,174]. In Lysidice, molecular analyses [225,228] supported previously reported morphological differences between Lysidice collaris and Lysidice ninetta [229]. These are the only eunicid species subject to phylogeographic and population studies and depict different evolutionary histories. L. collaris has panmitic populations, while L. ninetta includes two genetic lineages corresponding to different color morphs separated at around 3.7–3.4 million years ago, which may be different species [225,228]. Molecular analyses using nucleotide sequences of cytochrome oxidase I (COI) and 16S rDNA have become common in papers on eunicid species [202,230]. These have aided in understanding intraspecific variation, such as the lack of a subacicular hook in large specimens of some Marphysa species [47], in improving species recognition and in revealing a hidden biodiversity even in well-studied places.
3.6. Exploitation
Species of Eunice and Marphysa are commonly used as bait worms. However, the species identity of many of these has just become clear in the last few years [24,132]. The clear understanding of this diversity is essential to avoid the overexploitation of species as well as the introduction of exotic species due to commercialization or the establishment of farming [24].
4. Hartmaniellidae Imajima 1977
4.1. Systematics and Phylogeny
Hartmaniellidae has only one genus and three species. The first species, Hartmaniella erecta Imajima, 1977, was described from Suruga Bay, Japan. A year later, Amoureux [168] described a new genus and species from Tuléar, Madagascar, Pseudoninoe tulearensis, which he placed within Lumbrineridae. However, as was suspected by Pettibone [231] and Orensanz [13], this species belongs to Hartmaniella [232]. The third species, Hartmaniella fujianensis He and Wu, 1986, was described from the Taiwan Strait. Three other potential new species were described but not formally named from the Gulf of Mexico [232,233], the Andaman Sea [234], and Natuna Island, Indonesia [235].
Hartmaniellids superficially resemble Orbiniidae by having parapodia displaced dorsally (Figure 6A,C). However, they have well-developed maxillae and mandibles (Figure 3A) which unambiguously place them within Eunicida. However, several authors have debated on the classification of hartmaniellid maxillae. They have diverging views on the maxillary apparatus symmetry, homology among maxillary plates, carrier connection to maxillae I, and, thus, on hartmaniellids’ closest families. Imajima [236] mistakenly described a symmetric maxillary apparatus with four plates on both sides and illustrated the carriers as being poorly connected to maxillae I. Following Imajima’s description, Orensanz [13] suggested Hartmaniellidae being closely related to the fossils Synclinophora synclinalis Eisenack, 1975 and the Delosites-like scolecodonts, and with extant Eunicidae and Onuphidae. Later, based on the poor connection between the carriers and maxillae I illustrated in Imajima’s description, Fauchald and Rouse [237] considered the hartmaniellids maxillary apparatus more similar to the prionognath type and the family sister group of Oenonidae and Lumbrineridae. However, illustrations of Hartmaniella erecta by Szaniawski and Imajima [238] and the descriptions and illustrations of Hartmaniella tuleransis and Hartmaniella sp. by Carrera-Parra [232] show that the carriers are completely attached to the base of maxillae I. Szaniawski and Imajima [238] established a remarkable similarity of Hartmaniella jaws with fossil of Palurites Kozur, 1967 by the peculiar shape of maxillae I and II. They also proposed a close relationship between Hartmaniellidae extant Lumbrineridae and extinct Kielanoprionidae (asymmetric jaws), considering that the differences in structure (symmetric vs. asymmetric) of their maxillary apparatus are diagnostic at the family level. Eibye-Jacobsen and Oug [234] described an asymmetric maxillary apparatus in specimens from the Andaman Sea, and reported the right maxillae III and IV to be fused in contrast to the left separate plates. They hypothesized that asymmetry in maxillae could have evolved independently several times within Eunicida and supported the close relationships of hartmaniellids with Oenonidae or Lumbrineridae rather than with Eunicidae and Onuphidae. Carrera-Parra [232] stated that hartmaniellids have an asymmetrical maxillary apparatus with four maxillae on the left side and only three on the right side, reinforcing the earlier idea of close relationship with the asymmetrical fossils of Kielanoprionidae and with Eunicidae/Onuphidae. The latter was also supported by the presence of dorsal and ventral cirri in the three families. Lu [239], likewise based on the revision of Japanese specimens, described an asymmetric maxillary apparatus for H. erecta. Paxton [14] considered hartmaniellid jaws “subsymmetrical” and closest to the extant Lumbrineridae. For both families she proposed a new type of maxillae—symmetrognatha; however, the maxillary apparatus of hartmaniellids is asymmetric with four maxillae on the left side and only three on the right side [232,234,239] (Figure 3A).
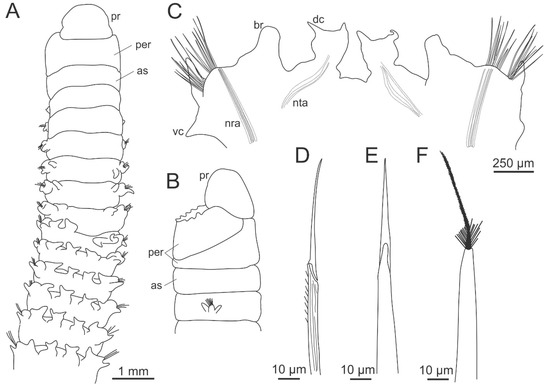
Figure 6.
Morphology of Hartmaniellidae. (A) Hartmaniella sp., anterior end, dorsal view; (B) the same, anterior end, lateral view; (C) the same, parapodia from midbody region shifted dorsally; (D) Hartmaniella tulearensis (Amoureux, 1978), composite spiniger; (E) the same, acicular spine; (F) the same, acicula. as–achaetous segment, br–branchia, dc–dorsal cirrus, nra–neuropodal aciculae, nta–notopodial aciculae, per–peristomium, pr–prostomium, vc–ventral cirrus. (B), (D–F) modified from Carrera-Parra [232].
A combined analysis of morphological characteristics and molecular data will help to clarify the placement of hartmaniellids within Eunicida, which, to date, has been hampered by the extremely scarce material and the lack of specimens suitable for molecular analyses.
No taxonomic revisions, key and phylogenetic analyses are available for the family.
4.2. Morphology
Hartmaniellids are small worms up to 2.5 cm long; however, most of the few collected specimens are incomplete. The prostomium lacks eyes and appendages (Figure 6A). The peristomium is laterally projected over the prostomium, the first ring is dorsally reduced, and the second one is longer dorsally than ventrally (Figure 6B). The absence or presence of papillae in the peristomium may aid to species identification. The first segment is apodous. The body is divided into three regions by the morphology of parapodia. In the first region, the parapodia are lateral; the second region is characterized by having parapodia displaced over the dorsum with bifid dorsal cirri and foliose parapodial branchiae dorsal to prechaetal lobes; while in the third region, the dorsal cirri are digitiform, basally swollen and the branchiae are tapering. The relative length of the body regions might be of taxonomical value at species level; however, it should be taken with caution, as these characteristics may be size-dependent. From chaetiger 6–7, all segments are biannulated. The chaetae of hartmaniellids are limbate with a serrated margin, compound spinigers with distally spinulose shaft, internally with granulated base becoming canaliculated from the median to the distal end; acicular spines with a long mucro appear from about chaetigers 6 (Figure 6D,E). The aciculae are aristate, subdistally hirsute with a long mucro finely hairy (Figure 6F). The number of chaetae and aciculae per parapodium may be of diagnostic value at species level.
Hartmaniellids have an asymmetric maxillary apparatus with four maxillae on the left side and three on the right (Figure 3A). Maxillae I are mainly characterized by long internal teeth and a lateral spur located on the outer posterior margin, while maxillae II are characterized by being a narrow plate with a lateral projection on the anterior outer edge. The number of teeth in maxillae I and maxillae II are important to distinguish the species.
4.3. Feeding and Life History
Jumars et al. [34] suggested hartmaniellids could be predators of meiofauna based on their size and the presence of maxillary apparatus. The only information about reproduction in hartmaniellids is the presence of deep yellow oocytes with large amounts of yolk, 150–200 µm in diameter in median parapodia, suggesting the development via lecithotrophic larvae [234].
4.4. Habitat, Distribution and Diversity
Hartmaniellids have been found in clayey and sandy silt, soft mud, sandy mud, and sandy mud with shell fragments, from depths of 40 to 210 m in six regions of the Pacific Ocean (Japan, China, Indonesia, Thailand, Madagascar, and La Réunion), while in the Atlantic Ocean, they have only been reported in the Gulf of Mexico in clayey and sandy silt at depths of 175 to 189 m. They are considered as being “living fossils”, abundant during the Mesozoic, but currently poorly represented [238].
There is no information about the ecology of the family.
5. Histriobdellidae Claus and Moquin-Tandon, 1884
5.1. Systematics and Phylogeny
The family Histriobdellidae is composed of highly specialized ectosymbionts known as “clown worms”. The prefix “histrio” (from the Etruscan word “hister”), which means actor or dancer, alludes to the performative way in which these worms move, quite similar to the actor Charlie Chaplin, in his character “Carlitos”. The species names Stratiodrilus arreliai Amaral and Morgado, 1997 and Stratiodrilus circensis Steiner and Amaral, 1999 also refer to the circus and its artists.
Histriobdellidae was first included in Hirudinea, which justifies the suffix “bdella” [240]. It has also been included among Rotifera and Archiannelida [241,242,243,244]. Mesnil and Caullery [245] have first suggested that maxillae of histriobdellids were homologous to other Eunicida, as a prionognath type. Tzetlin [246] considered the histriobdellid maxillae as of the ctenognath type. His views were recently corroborated by an extensive study of jaw morphology and ultrastructure in Histriobdella homari Van Beneden, 1858 showing the ultrastructure similarity of the maxillae with Dorvilleidae [247]. The inclusion of the family in Eunicida was followed by latter studies that considered histriobdellids as highly specialized commensals, lacking external resemblance to the traditional Errantia body plan [13,237,248,249,250].
This epizoic group is currently composed of three genera and 13 species that live within the branchial chambers (on branchiae, epipodites, or inner surfaces of carapace), in the egg masses, or, occasionally, on the general body surface (head, thorax, abdomen, or appendages) of marine and freshwater isopod and decapod crustaceans. Histriobdella Van Beneden, 1858 and Steineridrilus Zhang, 2014 are monospecific and exclusively associated with marine hosts, while Stratiodrilus Haswell, 1900 includes 11 species associated with freshwater hosts.
There is no phylogenetic hypothesis that deals with relationships between the genera and species within the family, only hypotheses regarding their marine or freshwater origin [251,252,253,254,255]. To date, most taxonomic and systematic studies on histriobdellids were based on external morphology with no molecular data available. A single study using SEM, TEM and confocal laser scanning microscopy described details of the jaw apparatus [247]. The only review was carried out by Steiner and Amaral [256] who proposed a nomenclature for some morphological structures, redescribed all species based on literature and proposed a new genus, Dayus, which, being preoccupied, was renamed for Steineridrilus [257]. Rosa et al. [255] updated the list of Stratiodrilus hosts based on published data and specialized databases discussing its distribution. An identification key for all described Stratiodrilus was published by Amato et al. [258].
5.2. Morphology
The worm-like translucent and delicate body is small, with the maximum length of adults varying between 0.5 to 1.5 mm, indistinctly and irregularly annulated, with constrictions more visible laterally, and divided into head, trunk and posterior region (Figure 1F and Figure 7A,B,D,E). Histriobdellids have no true segments, parapodia, chaetae or aciculae, but the ventral dark sclerotized jaws can be easily seen by transparency (Figure 7D–G). Despite the absence of metamerism, internally histriobdellids have 9 to 11 ganglia (depending on the genus) in the ventral nerve cord [248,259].
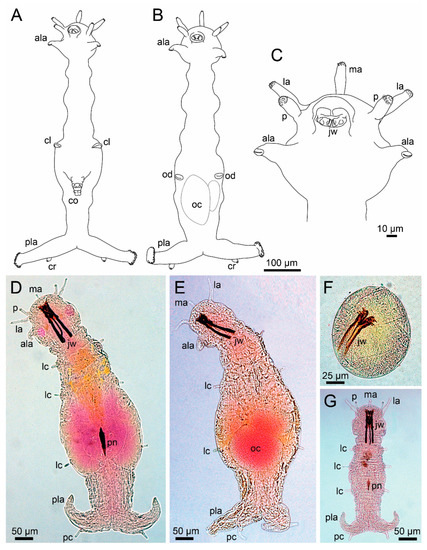
Figure 7.
Morphology of Histriobdellidae. (A) Histriobdella homari, male, ventral view; (B) the same, female, ventral view; (C) the same, anterior end, ventral view; (D) Stratiodrilus arreliai Amaral and Morgado, 1997, male, ventral view; (E) the same, female, ventral view; (F) the same, early embryo; (G) the same, juvenile, ventral view. ala–anterior locomotory appendage, cl–clasper opening, co–copulatory organ, pc–posterior cirrus, jw–jaws, la–lateral antenna, lc–lateral cirrus, ma–median antenna, oc–oocyte, od–oviduct opening, p–palp, pla–posterior locomotory appendage, pn–penis. (D–G), Copyright © 2016, Editora Guanabara Koogan (Fransozo, A. & Negreiros-Fransozo, M.L., Zoologia dos Invertebrados, 1st edition, page 366), reproduced with permission of the publisher.
The head is composed of prostomium fused to peristomium, and the trunk has five segments, but only the second, third and fifth may have lateral cirri, which are probably homologous to dorsal and/or ventral cirri of other Eunicida [260] (Figure 7A–E). Nuchal organs were reported in some species [242,259]. The prostomium bears three antennae dorsally and two sensory palps frontally (Figure 7C,D). One pair of locomotory appendages with adhesive glands on the distal end is located on the lateral of the head, probably the peristomium (Figure 7A–E), retractable in most species. In the fourth trunk segment, males have one pair of lateral retractable claspers for holding females during copulation, and a penis on the ventral side of body (Figure 7A,D,G). The posterior region of the body, after the fifth segment, is composed of several fused segments without cirri or any kind of appendages and two lateral lobes, the posterior locomotory appendages, with the anus located between them (Figure 7A,B,D,E,G). These appendages have a duo-gland adhesive system [261] on their distal ends, in addition to cirri, lobes, and/or tubercles. The jaw apparatus is very complex, enclosed in the ventral pharyngeal organ. The paired mandibles are fused along the mandibular plates bearing about 10 denticles and attached to each other in the median part of the mandibular shafts, which are flattened and elongated. The maxillae consist of a single dorsal rod, a massive ventral carrier and four pairs of dentate plates–maxillae I–IV [247,249] (Figure 3E).
All known species are dioecious and sexually dimorphic when mature, one of the few examples in polychaetes. The chitinous black penis of Stratiodrilus is an eversible spine with a bevelled tip [260] like a hypodermic needle [262]. In Histriobdella, the penis is composed of two hard structures with a central duct [260], but not chinitized. Detailed descriptions of general morphological features are available in Shearer [242], Haswell [262,263], Roubaud [244], Jennings and Gelder [249], and Steiner and Amaral [256].
Histriobdella is characterized mainly by the presence of short and papillae-like antennae and palps and the absence of lateral cirri on trunk (Figure 7A–C). Steineridrilus and Stratiodrilus have lateral cirri and longer and narrower antennae and palps. In Steineridrilus the three antennae are biarticulated, while in Stratiodrilus only the lateral antennae bear two articles (Figure 7D,E). The shape of the lateral cirri and the presence or absence of cirri, lobes and tubercles in posterior locomotory appendages determine Stratiodrilus species, especially those from the Neotropical region. Studies that address intraspecific variations are only known for Stratiodrilus circensis [264] that has been widely recorded in the Neotropical region. These worms are relatively easy to collect, just by finding their hosts, which carry in their body, especially in the branchial chamber, a good amount of complete specimens.
The morphological adaptations to the commensal life habit are diverse, when compared to the Eunicida pattern. They range from the reduction in body size, absence of metamerism, parapodia, chaetae and aciculae, to the presence of a duo-gland adhesive system in the posterior region, which allows its attachment to different parts of the host’s body. Sexual dimorphism and the penis also appear to be adaptations to the specific habitat of these worms.
5.3. Feeding and Life History
Histriobdellids find shelter in the host and feed on the microflora (bacteria, cyanophytic algae, diatoms, among others) that grows abundantly in the chamber and branchial filaments, bringing the benefit of always keeping these areas clean and free, so that gas exchange is carried out more efficiently. Jennings and Gelder [249] found that the relationship between Histriobdella homari and its crustacean host has a firm nutritional basis.
Regarding reproduction, the male hypodermically inserts its penis into the female and the sperm goes directly into the body cavity [265]. Whitish eggs in different stages of maturation are visible in the female’s transparent body (Figure 7E). In all known species, there is a suppression of the larval stage, with egg-laying occurring in branchial chamber or in egg masses of the female host. The development, therefore, is direct, and immature miniatures of adult worms hatch from the eggs. Histriobdellid females attach their eggs to the host (branchial chamber, between eggs of host female, pleopods and uropods), which allows the young worms to continue their life cycle on the host. Shearer [242] and Simon [266] stated that Histriobdella can migrate from the branchial chamber to the egg mass of female host and, after attaching the eggs, return to chamber. Histriobdella homari also has the ability to migrate from one host to another [266].
5.4. Habitats and Ecology
Histriobdellids have been treated as parasites [267,268] or commensals [260,269]. According to Brattey and Campbell [270], in most infestations Histriobdella homari benefits the lobster. However, if the number of individuals is very high, the benefit of microfloral grazing may decrease, as worms can reduce the oxygen flow on the branchiae, also reducing success in hatching host’s eggs. On the other hand, Lerch and Uglem [271] reported success in the hatching of female hosts’ eggs, even with high numbers of H. homari in the branchial chamber. According to Martin and Britayev [269], a symbiotic species that completes the entire life cycle in a single host has a monoxenous pattern, and they consider that H. homari behaves more as a cleaning symbiont rather than as a parasite. Jennings and Gelder [249] classify the species as epizoic microphagous cleaning symbiont.
Studies on species biology are still few; the mechanisms of dispersion and selectivity in host choice are still unknown, as well as the real nature or extent of the association with their hosts.
5.5. Distribution Patterns and Diversity
The differentiation between the histriobdellid genera are also reflected in their geographical distribution. The first described genus, Histriobdella, occurs exclusively on lobsters (Homarus, Nephrops) in the Northern Hemisphere, on both sides of the Atlantic Ocean, in Europe (Norway, Ireland, England, the Netherlands, the Irish Sea, the Clyde Sea), Canada, and the United States (mid-Atlantic Bight). Steineridrilus is the marine group from the Southern Hemisphere, having an intertidal isopod (Cirolana) as its host. It is only known from South Africa, where it occurs from the Lagebaan Lagoon to East London [272]. Stratiodrilus is the most speciose genus and encompasses species living on freshwater decapods from the Southern Hemisphere. It has been identified on different hosts (Astacopsis, Astacoides, Aegla, Cherax, Parastacus, Samastacus, and Trichodactylus) that occur in diverse freshwater environments, such as streams, brooks, flowing rivers, lagoons and caves from Australia (including Tasmania), Madagascar, Chile, Argentina, Uruguay, and Brazil.
The distribution of histriobdellids reveals interesting information that contributes to the continental drift theory, with Histriobdella having a Laurasian distribution, while Stratiodrilus and Steineridrilus show a Gondwanan distribution, occupying the Neotropical, Afrotropical and Australasian zoogeographic regions [254,255]. Harrison [251] suggested that Stratiodrilus distribution demands former land connections between Australia, Madagascar and South America, and proposed a marine ancestor for the family. The co-occurrence of Stratiodrilus in parastacid crayfish suggests a single colonization in continental waters, prior to the breakup of Gondwana [254,260]. Thus, Stratiodrilus appears to have successfully invaded continental waters either in Palaeozoic or in Mesozoic [254]. Jennings and Gelder [249] suggested that the suppression of a free-swimming larval stage is an adaptation to the freshwater environment and that Histriobdella and Steineridrilus have become secondarily adapted to a marine habitat. On the other hand, one of the strategies of some commensal species is simplification of their life cycles, either reducing or eliminating the free-living larval stage, with the adult phase being responsible for dispersion [269]. Thus, the direct development could be an adaptation to the symbiotic lifestyle and not to the freshwater environment, which would support the marine origin theory for the family.
Up to now, 44 species of nine genera of four Decapoda families have been found to host Histriobdella and Stratiodrilus species. Only one species of Isopoda hosts Steineridrilus, which is known only from its original description [272], including some biological information, in addition to a photographic record made by Rouse [268], from an unknown locality. Some histriobdellids do not demonstrate specificity to a single host. Histriobdella homari, for example, is found in three different species of lobsters of the genera Nephrops and Homarus, on both sides of the Atlantic. Stratiodrilus circensis is also widely distributed, occurring in at least 10 species of Aeglidae and in some Parastacidae, in Chile, Argentina and Brazil [255]. Crabs from the genus Aegla, the hosts for several Stratiodrilus species, are widespread in the South America with 72 known species [273] of which only 20 have been studied for the histriobdellid symbionts [255]. Furthermore, other genera and species of decapods and isopods can potentially host histriobdellids. Thus, the diversity of Histriobdellidae could be highly underestimated due to the absence of systematic study covering the wide diversity of potential hosts.
It is not known how common cryptic speciation is within Histriobdellidae. Rosa et al. [255] proposed studies using molecular tools to clarify the taxonomic status of S. circensis, which may represent a complex of cryptic species, considering its wide distribution in four countries. Briggs et al. [274] also considered the possibility that H. homari from two localities of Europe are different species or subspecies. Considering that the species is reported from both sides of the Atlantic, it may represent a species complex.
Stratiodrilus gondwanian distribution suggests it may have speciated with host parastacid crayfish, which have similar distribution. However, despite early suggestions of this relationship [251,254,255] and of the connection between diversity of histriobdellids and their hosts, no study has tested these hypotheses yet. Biogeographic studies incorporating molecular analyses are necessary to study the coevolution of hosts and histriobdellid ectocommensals.
Histriobdellidae is a relatively poorly studied family with knowledge gaps in practically all areas, despite being relatively easy to collect and to observe. Until 1962, studies were limited to descriptions of new species based on external morphology [240,243,251,262,263] and investigations of the internal anatomy [241,242,244,245,259,275]. More recently, the biology and behavior of several histriobdellid species [249,258,264,265,266,276,277], as well as relationships with their hosts and other ectosymbionts [252,271,278] received more attention. Few studies have been carried out on the ecology of the family [249,266,267,270,271,278,279] as well as species distribution records [255,280,281]. Studies that focus on the life cycle, molecular techniques, ecology, and taxonomy will certainly help in reconstructing phylogenetic relationships among the species of the family, as well as their relationships with their hosts.
6. Lumbrineridae Schmarda, 1861
6.1. Systematics and Phylogeny
Lumbrineridae comprises 19 genera and about 279 species. The first species of Lumbrineridae was described by Müller in 1776 as Lumbricus fragilis (now Scoletoma fragilis). Previously, some authors considered the group as a subfamily of Eunicidae, but today it is regarded as an independent family [282].
Lumbrinerids have a very simple body shape, with a marked reduction of external morphological characteristics (Figure 1D). Thus, classifications based on external morphology only resulted in a simplified generic system that lumped all known species in only three or four genera [283]. Hartman [28] and Fauchald [103] treated genera Augeneria Monro, 1930, Cenogenus Chamberlin, 1919, Eranno Kinberg, 1865, and Scoletoma de Blainville, 1828 as synonyms of Lumbrineris de Blainville, 1828. The taxonomy of this family has changed after employing characteristics of the maxillary apparatus and mandibles, which allowed the reestablishment of all above-mentioned genera [13,283,284,285] and the description of 13 new genera [13,284,285,286]. A key to genera valid at the time is available in Carrera-Parra [286] and Carrera-Parra [282].
To date, there is only one phylogenetic study, which is based on morphological data, regarding the intergeneric relationships within Lumbrineridae. The family is divided into four main clades supported by the characteristics of the maxillary apparatus [286]. This study corroborated the inclusion of Lysarete Kinberg, 1865 into Lumbrineridae proposed by Orensanz [13], as well as the recognition of Augeneria, Cenogenus, Eranno, and Scoletoma genera previously considered as junior synonyms of Lumbrineris. Furthermore, there is a phylogenetic study on interspecific relationships of Lumbricalus Frame, 1992 [287]. Only three genera of lumbrinerids underwent complete systematic revision: Kuwaita Mohammad, 1973 [288], Lumbricalus [289], and Lumbrineris [290]. The rest of the genera await revisions based on detailed study of the architecture of the maxillary apparatus and, possibly, molecular information.
6.2. Morphology
Lumbrinerids are worms of small to medium size, ranging from a few to over 60 centimeters [291,292]. Their prostomium can vary from round to conical; in most lumbrinerids, it lacks appendages (Figure 8A,B), except for Augeneria, Lysarete, and Kuwaita, which have three antennae (Figure 8C), as well as Cenogenus and Sergioneris Carrera-Parra, 2006, bearing a single antenna. Although Augeneria typically has three antennae, at least two species have multiple small antennae (Augeneria polytentaculata Imajima and Higuchi, 1975 and Augeneria riojai Aguirrezabalaga and Carrera-Parra 2007). Most lumbrinerids lack eyes, except Lysarete, which possess two pairs (Figure 8C). A pair of ventral pads of the dorsolateral folds anterior extension (sensu [37,38]; buccal lips sensu [284,293]), that may be well-developed, is present at the ventral posterior margin of the prostomium. The peristomium is composed of two rings and lacks appendages; dorsally, it is complete (Figure 8A,B) except in Lysarete, which have a dorsal incision reaching the first chaetiger (Figure 8C). Ventrally, the first peristomial ring is incomplete, and the second ring is projected forward. The parapodia are subbiramous with notopodium reduced to a small knob with notoaciculae in most lumbrinerids (Figure 8A), but well-developed dorsal cirri in Kuwaita and Lysarete (Figure 8C). Neuropodia are well developed in all lumbrinerids, constituted by prechaetal, chaetal, and postchaetal lobes, without ventral cirri (Figure 8D–F). Branchiae are absent in most genera; when present, they are associated with parapodial lobes of anterior chaetigers (Figure 8D); Cenogenus and Ninoe Kinberg, 1865) or over dorsum in posterior chaetigers (Kuwaita). All genera have limbate chaetae. The presence of other chaetae varies among genera: simple and compound multidentate hooded hooks (Figure 8G,H) and compound spinigers (Lumbricalus); simple and compound multidentate hooded hooks (Augeneria, Gallardoneris Carrera-Parra, 2006, Hilbigneris Carrera-Parra, 2006, Loboneris Carrera-Parra, 2006, Lumbrineris, and Sergioneris Carrera-Parra, 2006); simple multidentate hooded hooks and limbate simple multidentate hooded hooks (Abyssoninoe Orensanz, 1990); simple multidentate hooded hooks (Cenogenus, Eranno, Gesaneris Carrera-Parra, 2006, Helmutneris Carrera-Parra, 2006, Kuwaita, Ninoe, and Scoletoma); and simple bidentate hooded hooks (Lumbrinerides Orensanz, 1990 and Lumbrineriopsis Orensanz, 1990). Arabellonereis Hartmann-Schröder, 1979 and Lysarete lack hooks. The pygidium of lumbrinerids has two pairs of pygidial cirri, except in Gallardoneris, Lumbrinerides, and Lumbrineriopsis, which lack pygidial cirri. Until now, the pygidium shape of Gesaneris is unknown.
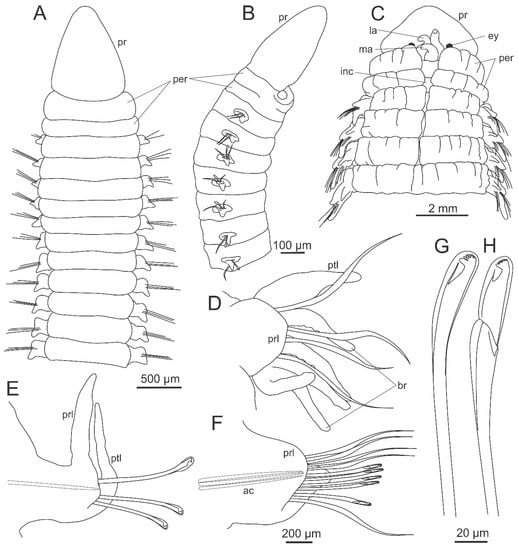
Figure 8.
Morphology of Lumbrineridae. (A) Lumbrineris mixochaeta Oug, 1998, anterior end, dorsal view; (B) Lumbrineriopsis sp., anterior end, lateral view; (C) Lysarete sp., anterior end, dorsal view; (D) Ninoe vargasi Carrera-Parra, 2001, parapodium; (E) Eranno lagunae (Fauchald, 1970), parapodium; (F) Lumbrineris sp., parapodium; (G) Scoletoma sp., simple hook; (H) Lumbrineris sp., compound hook. ac–acicula, br–branchia, ey–eye, ins–dorsal incision, la–lateral antenna, ma–median antenna, per–peristomium, pr–prostomium, prl–prechaetal lobe, ptl–postchaetal lobe.
The study of lumbrinerids requires careful observation of the maxillary apparatus since the recognition of genera is based mainly on the differences in this complex structure. The maxillary apparatus is symmetrical of labidognath type (sensu [286]) or symmetrognath type (sensu [14]); the later was suggested to distinguish the lumbrinerid symmetrical labidognath maxillae from the asymmetrical labidognath maxillae of Eunicidae and Onuphidae (Figure 3). The number of maxillary plates varies among the genera as follows: six pairs in Lysarete; five pairs in Arabelloneris, Eranno, Hilbigneris, Kuwaita, Ninoe, Lumbricalus, Lumbrineris, and Scoletoma; and four pairs in Abyssoninoe, Augeneria (Figure 3B), Cenogenus, Gallardoneris, Gesaneris, Helmutneris, Loboneris, Lumbrinerides, Lumbrineriopsis, and Sergioneris. All maxillae can have either an accessory lamella, a connecting plate, or, in some cases, both structures. As a consequence, the maxillary apparatus of lumbrinerids has a great variety of shapes specific to each genus [286]. The mandibles are also an important characteristic for helping to recognize some genera such as Gallardoneris and Lumbrinerides, which have completely fused mandibles. In contrast, all other genera have mandibles fused up to 3/4 of their length [286].
6.3. Feeding and Life History
In their feeding behavior, lumbrinerids were considered carnivores, feeding on various invertebrates, questionably herbivores [294,295,296] and deposit-feeders [297]. Based on the analysis of stable isotopes (δ15N) in species of the genera Hilbigneris, Lumbrineris, Lumbrinerides, and Scoletoma, most lumbrinerids are carnivores with a few herbivorous [34].
There is little information on the reproductive aspects of this group. However, all species with known reproduction are gonochoric without sexual dimorphism. Mature specimens of Lumbrineris latreilli Audouin and Milne Edwards, 1834 have been reported swimming [298]. In the few existing studies [299,300,301], the morphology of the sperm is described as of the ect-aquasperm type (sensu [302]) suggesting broadcast spawning [84]. Osman et al. [57] described three maturity stages according to yolk deposition during the oogenesis of Scoletoma funchalensis (Kinberg, 1865), the first previtellogenic stage and the other two within the vitellogenic phase. Oocyte diameter is known from four species. It varies from around 100 to 300 μm in diameter, S. funchalensis (140 μm to 280 μm, [57]), Gallardoneris iberica Martins, Carrera-Parra, Quintino and Rodrigues, 2012 (129 μm to 188 μm, [303]), Lumbrineris lusitanica Martins, Carrera-Parra, Quintino and Rodrigues, 2012 (181 μm to 191 μm [303]) and Lumbrineris pinaster Martins, Carrera-Parra, Quintino and Rodrigues, 2012 (98 μm to 147 μm [303]).
Messina et al. [300] studied the first stages of the development of Scoletoma impatiens (Claparède, 1868). They observed the appearance of protrochophore larva in the 24 hours after fertilization. Three days later, it transforms into a metatrochophore with a duration of about seven days, in which time the first two segments are formed. By the eighth day, the postlarva benthic phase begins. Some species lack pelagic larvae; their gametes are deposited into the parental burrow or gelatinous masses attached to the substrate where the first development stages occur and benthic larvae emerge at different times, not allowing for a high dispersal capability [284,304,305,306,307].
6.4. Habitats and Ecology
Lumbrinerids are motile worms dwelling in soft sediment such as mud and sand; however, some species prefer hard substrates such as coral rocks. They can construct very fragile temporary tubes of sediment and mucus. Some species live in association with other invertebrates such as sponges, bryozoans, sea urchins [176,286,308,309]. The species Helmutneris flabellicola (Fage, 1936) and Helmutneris corallicola Carrera-Parra, 2006 are only found forming membranous transparent tubes in ahermatypic corals of the genera Caryophyllia, Balanophyllia, Dendrophyllia, and Flabellum [286,310]. Fossil corals with tube scars created by lumbrinerids were reported from the Miocene and Pliocene deposits in central Europe and in the Western Basin of the Mediterranean [310].
Very little is known about the ecology and population biology of the Lumbrineridae. It has been shown that some species of Lumbrineris and Gallardoneris prefer areas enriched with organic matter [303,311,312]. Kuwaita hanneloreae Arias and Carrera-Parra, 2014 was reported from a fine to medium sand with a mean organic matter content of 1.68%, reaching densities over 4–5 ind/m2 and occurring together with other lumbrinerids, Scoletoma impatiens and Lumbrineris latreilli [313].
6.5. Distribution Patterns and Diversity
Lumbrinerids have a worldwide distribution, from intertidal to abyssal zones. They are more common in temperate and tropical regions, where all genera have been reported [282,286], while in the Antarctic and Artic regions only six genera have been found [13,290,308,314,315]. However, the taxonomic studies of lumbrinerids in the Antarctic and Arctic regions and the deep-sea habitats are scarce.
Some contributions to the regional faunas of lumbrinerid species are available for the Antarctic and Subantarctic seas [13], Argentina [284], Australia [316]; Brazil [176,317,318], the British Isles [181]; the Canary Islands [319]; the Caribbean region [139,220,308]; China [320,321,322]; Costa Rica [323]; the Eastern Pacific [28]; East Africa and the Red Sea [324]; El Salvador [325]; Fiji and Samoa [190]; the Gulf of Mexico [308,326,327,328,329]; the Iberian Peninsula [303]; India [330]; Japan [290,331,332,333]; the Mediterranean Sea [4,334,335,336,337]; New Zealand [338]; South California [291]; Thailand [286,339]; the USA East coast [285]; northern Europe [313,335,340,341]; western Mexico [94,103,290,342]; the Scandinavian and North Sea coasts [315,343,344,345,346]; South Africa [216]; the southeastern Pacific [290,347,348,349,350]; Vietnam [286,351]; and West Africa [289,352,353,354].
Due to recent changes in the group’s taxonomy, it is necessary to re-evaluate the lumbrinerid species worldwide. A number of new genera or species have been described, or other species have been re-established from their supposed synonyms even in well-studied areas [283,286,290,303,336,342]. Several lumbrinerid species have been regarded as having wide distribution (v. gr. Scoletoma tetraura (Schmarda, 1861), Scoletoma impatiens, Lumbrineris latreilli, Lumbrineris inflata Moore, 1911), most probably as a result of brief original descriptions and unjustified synonymies [283,290]. The revision of type materials with a detailed examination of the maxillary apparatus, chaetal morphology and shape of parapodia along the body is imperative to clarify these cases. Recently, some species have been re-established, and others were described as new species with restricted distributions [283,290,303,321,336].
The use of combined morphological and molecular data may help to improve the identification. Until now, only an Antarctic species identified as Lumbrineris kerguelensis-cingulata has been the focus of a molecular analysis [355]. The examined specimens exhibit a high range of genetic distance among 16S rDNA sequences (0–16%), suggesting the presence of more than one species. However, the poor morphological condition of the specimens and the lack of additional genetic markers in the analyses did not allow further conclusions and formal description of the obtained clades.
6.6. Exploitation
Lumbrinerids are used as bait for recreational or subsistence fishing. In the Suez Canal, Scoletoma funchalensis is under intensive harvesting due to its high price and local demand [57]. In the Gulf of Naples, Scoletoma impatiens is harvested from shallow sandy areas: this species is locally named “Esca rossa” or “Tremolina” [54]. In southwest Spain (Cádiz), sold specimens of S. impatiens are harvested in northwest Spain [356]; in the Canary Islands, Lumbrineris latreilli is a highly appreciated bait used by experienced fishers [357]. In northern Spain, Kuwaita hanneloreae, S. impatiens, and L. latreilli are highly appreciated as fishing bait and constitute an important natural resource from this area [313]; however, dermatitis problems have been documented when handling the latter species [358]. In Japan, Kuwaita heteropoda (Marenzeller, 1879) is commonly used as bait by Japanese fishers who observed that certain carnivorous insects die when they settle upon the worm. The insect death occurs by having contact with a neurotoxin called nereistoxin that is localized in the integument of the lumbrinerid [359].
7. Oenonidae Kinberg, 1865
7.1. Systematics and Phylogeny
Oenonidae comprises 12 genera and about 109 species. The first species of oenonids was described in 1804 by Montagu as Nereis iricolor (now Arabella iricolor) [360]. To date since the original family description, Oenonidae has undergone many changes in its composition, giving rise to a series of modifications and synonyms at the family level. Kinberg [361] proposed the family including four genera (Aglaura Savigny in Lamarck, 1818, Andromache Kinberg, 1865, Danymene Kinberg, 1865, and Oenone Savigny in Lamarck, 1818). Hartman [28] considered Oenonidae the synonym of Lysaretidae, another family proposed by Kinberg [361], which is currently included into Lumbrineridae. However, Colbath [362], based on the morphology and mineral composition of the maxillary apparatus, recognized Oenonidae as including the genera Oenone, Halla Costa, 1844, and Tainoka Knox and Green, 1972. One of the most notable changes in the delimitation of Oenonidae was the inclusion of the family Arabellidae Hartman, 1944 as its junior synonym, proposed by Orensanz [13]. Therefore, the family was expanded to 11 genera (Arabella Grube, 1850, Biborin Chamberlin, 1919, Drilognathus Day, 1960, Drilonereis Claparède, 1870, Haematocleptes Wirén, 1886, Halla, Labrorostratus de Saint-Joseph, 1888, Notocirrus Schmarda, 1861, Oenone, Oligognathus Spengel, 1882, and Tainoka). Two years later, Dean [363] proposed one more genus, Pholadiphila, bringing the total number of currently accepted genera to 12. A key to valid genera is available in Carrera-Parra [364].
Two genera, Arabella and Drilonereis, comprise 76% of the oenonid species; three genera are monospecific (Biborin, Drilognathus, and Pholadiphila), all parasites. Five genera have two species (Haematocleptes, Halla, Oenone, Oligognathus, and Tainokia), but Oenone includes another six species regarded as junior synonyms of the supposed cosmopolitan species Oenone fulgida Savigny in Lamarck, 1818 (Figure 1C). Labrorostratus and Notocirrus have six and nine species, respectively.
Phylogenetic studies based on molecular data supported the inclusion of Arabellidae into Oenonidae, finding a monophyletic group containing Arabella, Drilonereis, and Oenone [8]. No phylogenetic analyses focusing on the family have been carried yet.
7.2. Morphology
Oenonids are worms of variable length ranging from few centimeters to almost a meter—all of them are usually very thin. The prostomium usually lacks appendages (Figure 9A,B), except in Tainokia, which has one antenna, and Halla and Oenone with three antennae (Figure 9G). The peristomium is entire (Figure 9A,B,G) but can be dorsally incomplete with a median anterior incision poorly developed as in Tainokia or well-developed as in Halla. Usually, the parapodia have very small globular dorsal cirri (Figure 9C,D), but foliaceous in Oenone and Halla (Figure 9H). All oenonids lack ventral cirri. Drilognathus is the only genus lacking chaetae; in the other genera, all chaetae are simple, including smooth or denticulated limbate chaetae (Figure 9E,F). In some genera, there are acicular spines with or without hoods. Only Oenone species and one species of Tainoika (Tainokia logachevae Ravara and Cunha, 2018) have bidentate hooded hooks [364].
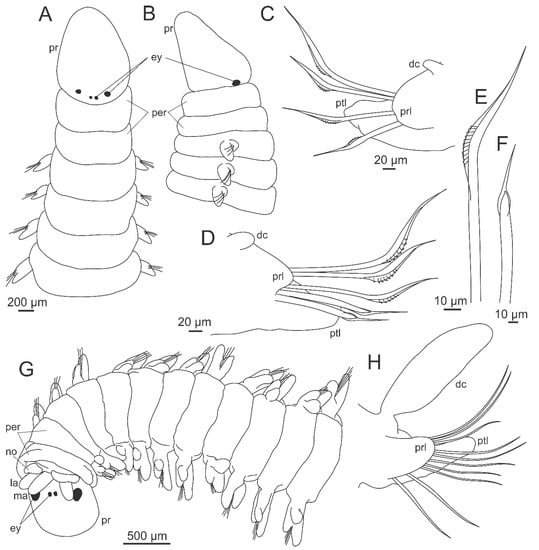
Figure 9.
Morphology of Oenonidae. (A) Arabella mutans (Chamberlin, 1919), anterior end, dorsal view; (B) the same, lateral view; (C) the same, parapodium 5; (D) the same, parapodium 26; (E) the same, limbate chaeta; (F) the same, chaetae tapering abruptly to guards (G) Oenone fulgida, anterior end, dorsal view; (H) the same, parapodium from midbody region. dc– dorsal cirrus, ey–eye, la–lateral antenna, ma–median antenna, no–nuchal organ, per–peristomium, pr–prostomium, prl–prechaetal lobe, ptl–postchaetal lobe. (B–F) modified from Zanol and Ruta [365].
In addition to these important external characteristics, the study of this group of worms requires careful observation of the maxillary apparatus since its taxonomy is based mainly on the differences in this structure. The maxillary apparatus is of prionognath type, characterized by the presence of paired slender dorsal and unpaired ventral maxillary carriers, which are much longer than the maxillae themselves (Figure 3F). Due to the existence of free-living and parasitic species, the architecture of the maxillary apparatus in oenonids is highly variable. It is well developed in free-living species where several pairs of maxillae can be present, such as in Oenone or Arabella, or can be extremely reduced to only maxillary carriers and, sometimes, a pair of maxillae, as in parasitic Drilognathus and Labrorostratus.
The maxillary apparatus of oenonids is symmetrical, with the same number of maxillae on both left and right sides. In some taxa, there is a condition called asymmetric, which refers to the difference in the shape and size of maxillary elements, but not to the number of plates as in Eunicidae and Onuphidae. This type of asymmetry in the shape of the maxillary plates is usually observed in the first two maxillae. In some species, both maxillae I have similar size and falcate shape with a denticulated base, and they are similar in size to maxillae II (Figure 3F). In contrast, other species have the left maxilla I falcate with a denticulated base and the right maxilla I as a denticulated rectangular plate much shorter than maxillae II [14,364]. The mandibles of oenonids are not fused, and without calcified distal plates, lacking growth lines [13], except for Oligognathus (whose mandibles are fused in a single horseshoe-shaped piece), Drilonereis (where the mandibles may be absent in some species), or Arabella aracaensis Steiner and Amaral, 2009 (with growth lines on its mandibles).
7.3. Feeding and Life History
Oenonids were regarded as selective carnivores or surface deposit-feeders [294]. Carnivorous activity in addition to the ability to feed on their hosts has been recently corroborated based on the results obtained with stable isotopes (δ15N) [34]. Studies in species of the genus Halla, which have a marked preference for their prey, bivalves [58,366], indicate that these worms produce mucus that contains paralyzing and digestive substances that force the bivalve to open its valves, to later ingest the soft tissue [367]. Under stress conditions, Halla species secrete another type of purple mucus, which, when in contact with human skin, is difficult to eliminate [368]. This mucus is produced by glandular structures of the epidermis [369], and it is considered a chemical defense mechanism [368]. Purple mucus secretion also occurs in species of Oenone, when they are under handling stress [370].
Very little is known about reproductive biology in oenonids. Most of the studied species are gonochoric without sexual dimorphism. Amaral [371] and Steiner and Amaral [51] reported adult individuals of the parasitic species Labrorotratus prolificus Amaral, 1977 reproducing asexually by stolons. The study of the reproductive biology of Halla parthenopeia (delle Chiaje, 1828) showed that the appearance of mature sperm occurs in two periods, during late April and in late December; while spawning occurs in May and January [372]. The sperm of this species was described as of the ect-aquasperm type (sensu [302,372]). Osman et al. [58] described the oogenesis in this species with three maturity stages; the first one is a previtellogenic phase, with small racemelike oocytes, while the other two stages are vitellogenic phases, with oocytes free in the coelomic cavity.
7.4. Habitats and Ecology
Oenonidae includes free-living species, endoparasitic species, and species that, during their ontogenetic development, have an endoparasitic phase but are free-living as adults [51,371,373,374].
Free-living species are typically soft-sediment dwellers, and although they do not construct a tube, they secrete mucus to prevent their burrow from collapsing [375]. The parasitic forms mainly infest the polychaetes of different families, for example, Terebellidae, Nereididae, Onuphidae, Eunicidae, Arenicolidae, but they have also been found in spoon worms and bivalves [51,269,363,364,365,376,377]. Most of the parasitic species apparently have a high degree of specificity infecting a single host [269]. They can be found from one specimen to more than 50 infecting a single host, sometimes reaching larger sizes than their host [371,373,374,376]. Pettibone [373] suggested that infections occur during the early developmental stages of the parasite, but the mechanisms of infection and the stage at which it occurs are unknown.
Information about oenonid ecology is scarce. They are not frequently recorded, and usually occur in low abundance in mud, sand, coralline rock, among algae, or as parasites.
7.5. Distribution Patterns and Diversity
Oenonids have a worldwide distribution from the intertidal zone to the abyssal depths. They are more common in tropical and temperate regions. In the Arctic and Antarctic regions, no more than eight species of Arabella, Driloneries, and Notocirrus have been recorded [13,378]. Some contributions to regional faunas of oenonid species are available for the Antarctic and Subantarctic seas [13], Argentina [379], Australia [365,380]; Brazil [51,371,381], the British Isles [181]; the Caribbean region [139,220,377,382,383]; Chile [349,350]; Costa Rica, Pacific side [323,384]; the Eastern Pacific [28]; East Africa and the Red Sea [324]; Fiji and Samoa [190]; the Gulf of Cádiz [385]; the Gulf of Mexico [326,386,387,388]; India [330]; Japan [389,390,391,392,393]; the Mediterranean Sea [4,394,395,396,397]; New Zealand [398]; Norway [399]; South California [375]; Thailand [339]; western Mexico [103,376]; South Africa [216]; Vietnam [351]; and West Africa [353,354]. Some records of deep-sea oenonids in various areas of the world were reported by Chamberlin [61], Hartman [400], Hartman and Fauchald [401], and Kirkegaard [402].
Many regions of the world lack studies on oenonids, suggesting that oenonid diversity is still underestimated. Despite the low abundance of oenonids, new species have recently been found both in shallow-water and deep-sea habitats (v gr. [365,385]).
Some oenonid species have been regarded as widely distributed, but this distribution is usually the result of poor original description and unjustified synonymies. Oenone fulgida, originally described from the Red Sea, currently includes six species as junior synonyms: Oenone diphyllidia Schmarda, 1861 from Jamaica, Oenone haswelli Benham, 1915 from Australia, Oenone pacifica Fischli, 1900 from Ternate, Oenone symmetrica Fauvel, 1914 from the Gulf of Guinea, Oenone telura Chamberlin, 1919 from Marshall islands, and Oenone vitiensis Grube, 1871 from Fiji. Arabella iricolor (Montagu, 1804) originally described from England has four junior synonyms: Arabella lagunae Chamberlin, 1919 from California, Arabella maculosa Verrill 1900 from the Bermudas, Arabella multidentata (Ehlers, 1887) from the Gulf of Mexico, and Arabella setosa Treadwell, 1921 from Tobago. Based on the revision of the type material, Perkins [326] found morphological differences to consider A. multidentata and A. maculosa as valid species, rejecting the synonymy with A. iricolor. The revision of type materials, including a detailed examination of maxillary apparatuses and chaetae, is critical to clarify the taxonomic status and distribution of oenonid species.
The use of molecular data may help to improve identification. However, until now, only seven species (A. iricolor, Arabella semimaculata (Moore, 1911), Drilonereis longa Webster, 1879, Drilonereis filum (Claparède, 1868), Halla okudai Imajima, 1967, O. fulgida, and Tainokia logachevae) have available molecular information in public databases. Other molecular information is available, but the specimens were identified just at family or genus level.
7.6. Exploitation
At least two species of the family are used as bait. Halla parthenopeia is commonly utilized for sport fishing in the Mediterranean Sea [58,372,403]. This species is locally named “cuc llobarrer” on the Catalan Maresme coast; fishers capture an average of 2.17 individuals per boat per day [403]. The second species is Halla okudai. It is locally named Tai-mushi in Japan, where considered an effective bait worm, reaching the highest prices among bait worms [404]. This species is also appreciated as bait in Malaysia; the specimens are harvested during low tide [405].
8. Onuphidae Kinberg, 1865
8.1. Systematics and Phylogeny
Onuphidae comprises 22 genera and approximately 340 species. Over their history, onuphids were recognized as a subfamily [4,216] or as a family [28,61] constituting 8–10 genera, with most of species belonging to Onuphis Audouin and Milne Edwards, 1833 and Nothria Malmgren, 1867. The revision of the family started from the work of Pettibone [406] who redefined Nothria and Paradiopatra Ehlers, 1887. Kucheruk [407] revised the generic diagnoses of 11 genera and described three new genera, one of which, Aponuphis Kucheruk, 1978, remains valid in the current system of the family. Fauchald [408] re-examined the type materials for all species of former Onuphis, Nothria, and Paradiopatra and described three new genera, of which Mooreonuphis Fauchald, 1982 and Kinbergonuphis Fauchald, 1982 are currently accepted. Paxton [409] provided the comprehensive revision of the whole family, redefining all accepted genera and describing five new genera. She subdivided onuphids into two monophyletic subfamilies: Onuphinae Kinberg, 1865 and Hyalinoeciinae Paxton, 1986 possessing morphological synapomorphies such as the position of subacicular hooks in the posterior parapodia and arrangement of the nurse cells in the oocytes (Figure 10D,H). Paxton’s [409] system remained largely valid to date with few minor changes: Neonuphis Kucheruk, 1978 was synonymized with Leptoecia Chamberlin, 1919 [13,17]; monotypic Fauchaldonuphis Paxton, 2005 was described [410]; Epidiopatra Augener, 1918 and Notonuphis Kucheruk, 1978 were synonymized with Diopatra Audouin and Milne Edwards, 1833 and Paradioptra, respectively, and two more monotypic genera, Paxtonia Budaeva and Fauchald 2011 and Protodiopatra Budaeva and Fauchald 2011, were described [32]. A key to genera valid at the time is available in Paxton [409] and Carrera-Parra [411].
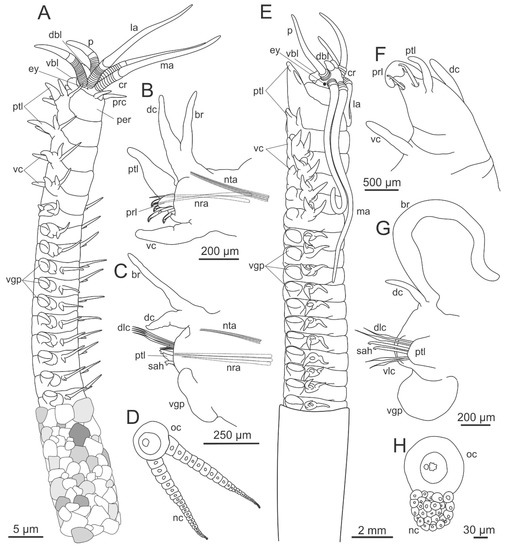
Figure 10.
Morphology of Onuphidae. (A) Onuphis sp. (Onuphinae), anterior end, lateral view; (B) the same, anterior modified parapodium; (C) the same, posterior parapodium; (D) Onuphinae, oocyte; (E) Hyalinoecia sp. (Hyalinoeciinae), anterior end, lateral view; (F) the same, anterior modified parapodium, aciculae not shown; (G) the same, posterior parapodium, aciculae not shown; (H) Leptoecia vivipara Orensanz, 1990, oocyte. br–branchia, cr–ceratophore, dbl–dorsal buccal lip, dc–dorsal cirrus, dlc–dorsal limbate chaetae, ey–eye, la–lateral antenna, ma–median antenna, nc–nurse cells, nra–neuroaciculae, nta–notoaciculae, oc–oocyte, p, palp, per–peristomium, prc–peristomial cirrus, prl–prechaetal lobe, ptl–postchaetal lobe, sah–subacicular hook, vbl–ventral buccal lip, vc–ventral cirrus, vgp–ventral glandular pad, vlc–ventral limbate chaetae.
A number of onuphid genera have been revised and their taxonomy and identification are rather well resolved: Australonuphis Paxton, 1979 [412]; Hirsutonuphis Paxton, 1979 [413,414]; Brevibrachium Paxton, 1986, Longibrachium Paxton, 1986, and Rhamphobrachium Ehlers, 1887 [415,416]; and Paradiopatra [32,417]. However, several species-rich genera, such as Anchinothria Paxton, 1986, Diopatra, Hyalinoecia Malmgren, 1867, Kinbergonuphis, Nothria, and Onuphis await detailed revision.
Phylogenetic hypotheses of Onuphidae were proposed based on morphological [409] and molecular [12] data. Both studies agreed in recognizing the two subfamilies; 10 of 22 genera described based on morphology were tested with molecular tools and were recovered as monophyletic, largely supporting the accepted system of the family provided by Paxton [409]. The hypotheses on the relationships between the genera and recognition of generic groups and complexes sensu Paxton [409] were not corroborated by the molecular data: nevertheless, taxon sampling was incomplete and only one nuclear (18S rDNA) and a fragment of a mitochondrial marker (16S rDNA) were used in the reconstruction [12]. Further detailed studies are required to reveal better resolution at the basal part of the onuphid tree and to assess interrelationships between the genera.
8.2. Morphology
Onuphids vary in size and number of chaetigers from 2 cm and about 100 chaetigers (e.g., Leptoecia, Nothria, and Anchinothria) to 70–300 cm and over 1000 chaetigers in beach worms Australonuphis and Americonuphis. The majority of species have about 200 chaetigers and are 10 cm in length [409].
Onuphids are defined by two main synapomorphies: the presence of well-delimited dorsal buccal lips (frontal lips) attached to the anterior margin of the prostomium and several pairs of modified anterior parapodia (Figure 10A,B,E,F). They have a rounded prostomium with dorsal and ventral buccal lips (frontal and upper lips), a pair of palps, a pair of lateral antennae and a single median antenna (Figure 10A,E). Both antennae and palps are placed on multiringed ceratophores consisting of 2–70 rings (Figure 10A,E). Many species possess eyes and dorsal nuchal organs in the shape of straight to semicircular ciliated grooves. The peristomium is a single ring, with (Figure 10A) or without (Figure 10E) peristomial cirri, ventrally forming a wide lower lip. The anterior 2–8 pairs of parapodia are modified, usually directed slightly anteriorly bearing special kind of chaetae: simple or pseudocompound falcigers (Figure 10B,F). Modified parapodia are used in locomotion and feeding and are highly adaptive to different lifestyles. In Hyalinoeciinae, the modified parapodia are enhanced and enlarged with auricular prechaetal lobes and short and stout falcigers (Figure 10E,F); they are used for crawling on the surface of the sea floor [418]. Species of Rhamphobrachium, Longibrachium and Brevibrachium have elongated modified parapodia with extremely long spiked and distally curved falcigers that can protrude anteriorly and are used in catching prey in an ambush-predator feeding strategy [419]. Australian beach worms (Australonuphis, Hartmanonuphis Paxton, 1986 and Hirsutonuphis) have massive anterior parapodia used for actively moving inside sandy sediments and making temporary borrows [53]. The modified parapodia may not be enlarged but always possess elongated ventral cirri (Figure 10B,F), while in following parapodia the ventral cirri are modified into glandular pads producing the organic secretion used in tube building (Figure 10C,G).
The majority of onuphid worms (except beach worms) are tubicolous dwellers and the shape and the composition of their tubes is quite diverse. Quill worms (Hyalonoecia and Leptoecia) build completely organic transparent light and firm tubes that they are able to carry along as epibenthic crawlers (Figure 1E and Figure 10E; [418]). Anchinothria and Nothria have tubes made of a parchment-like inner layer covered by mud, sand of shell fragments; these tubes are dorsoventrally flattened, suggesting an epibenthic lifestyle for these two genera. Species of Diopatra construct vertical tubes with so-called “tube caps” protruding above the sediment surface and incrusted by fragments of shells, plant debris and other foreign objects. Many other species build cylindrical tubes covered by sand or mud particles, which usually are permanent and significantly exceed their inhabitants in length (Figure 10A).
The number and morphology (i.e., size and shape of various parapodia lobes) of the anterior modified parapodia, the arrangement of branchial filaments, number of rings on the ceratophores, and the presence or absence of specific chaetae are the main characteristics used in the identification of onuphid genera. The species level diagnostic characteristics are mainly the distribution of different types of chaetae and branchiae along the body, the number of denticles in the anterior falcigers, and the number of branchial filaments. Intraspecific variation in morphological characteristics is very common in onuphids, leading to difficulties in species identification and delimitation based on morphology. This is especially prominent in species-rich genera such as Diopatra, Onuphis, Kinbergonuphis and Hyalinoecia lacking complete revisions to date. It is probably impossible to revise these genera utilizing morphology only; a complete revision would require the inclusion of molecular data.
8.3. Feeding and Life History
Omnivorous or scavenging feeding modes have been documented in onuphids with some species feeding primarily on kelp [294]. Stable isotope analysis [418] and analysis of gut content [420] suggest that quill worms are secondary consumers. Being motile predators and/or scavengers, large-bodied quill worms are able to move around in search of sources of food on the sea floor and were many times recorded on underwater photographs and video attracted by fish bait or whale falls [421]. Large-bodied beach worms (Australonuphis spp.) were reported to aggregate in high numbers and feed on large carrion such as washed up whale or sheep’s head [53]. Ambush predator behavior was described in detail for Longibrachium arariensis Nishi and Kato, 2009 utilizing its extremely modified, prolonged and retractable anterior parapodia and falcigers in catching small prey [419]. Diopatra species were shown to feed on foraminiferans [422] or minute invertebrates inhabiting their complex tubes protruding vertically from the sediment [423,424]. Little is known about the feeding habits of the small-bodied and largely sessile onuphids dwelling in permanent tubes inside the sediment.
Onuphids display a great variety of reproductive strategies. Several cases of protandric simultaneous [425,426] or sequential [427] hermaphroditism have been observed in onuphids as well as cases of sperm transfer via spermatophores [428] and sperm storage by females [428] or hermaphrodites [427]. All species investigated so far have lecitotrophic development but many provide parental care for their young by laying eggs inside their tubes or cocoons or egg masses attached to the tubes. The broadcast spawners are also known in the family; however, they are less common, possibly due to sampling bias. Few species were shown to have short-living pelagic stages [429,430] while the majority have direct development inside parental tubes or cocoons [430,431,432,433,434,435]. At least two cases of viviparity were observed in miniature deep-sea Leptoecia species, with juveniles developing inside the parental body cavity up to an advanced stage [13,436].
Large-bodied species (i.e., Diopatra ornata Moore, 1911) were shown to serve as hosts for oenonids, another eunicidan family of bristle worms [376]. Portuguese populations of Diopatra neapolitana Delle Chiaje, 1841 were reported to be infected by digeneid worms [437] and myxozoans [438]. Symbiosis between onuphids and ciliates attached to their parapodia was described by Arias et al. [439] and Arias and Paxton [440].
8.4. Habitats and Ecology
All known onuphid species are free-living. They are predominantly soft-bottom dwellers, either epibenthic or infaunal, commonly found in muddy or sandy sediments. Several species of Diopatra can form large aggregations and were described as ecosystem engineers—species physically creating or modifying the habitat [441]. Having vertical robust and complex tubes sticking above the sediment surface, the species act as sediment stabilizers and increase the abundance and richness of the local benthic fauna [442,443]. While Diopatra cuprea (Bosc, 1802) was shown to reach up to 250 ind/m2 at the Atlantic coast of the US [443], the Hawaiian species, Diopatra dexiognatha Paxton and Bailey-Brock, 1986, is able to form intertidal mounds with extreme densities of up to 21800 ind/m2. These mounds enhance organic matter accumulation and provide a complex tridimensional habitat for small invertebrate species [444]. No onuphids have been reported from hydrothermal vent fields; however, in cold seeps of the US Atlantic continental margin, Hyalinoecia artifex Verrill, 1880 occurs in high abundancies (up to 70 ind/m2) [418].
8.5. Distribution Patterns and Diversity
Onuphids are widely distributed around the world, found in all oceans from the intertidal zone down to over 6000 m deep [445,446]. The diversity of Onuphidae in the Arctic is rather poor with only two species, Nothria conchylega (Sars, 1835) and Paradiopatra pauli (Annenkova, 1952), reported from the shelf areas [447], while the Antarctic onuphid fauna is diverse with many species found both in deep and shallow waters [13,166,448].
Regional onuphid faunas were described from Central America and adjacent waters of the East Pacific and West Atlantic [94,189,449,450,451,452], Antarctic and Subantarctic Seas [13,448]; Japan [453,454,455,456], Australia [53,413,416,417,430,457,458,459], South and West Africa [46,216,460,461,462]; South America [43,463,464] and the eastern Atlantic including Norway [343], southern Europe, the Mediterranean [440,465,466,467,468] and the Macaronesian region [469]. Large parts of the world’s onuphid fauna remain poorly studied; for example, from the Indian Ocean and southeastern Asia as well as from most of the deep-sea habitats. Nonetheless, even relatively well-studied regions hold many undescribed species, which is demonstrated by the recent increase in the description of new species from Australia and the East Atlantic (Figure 2).
Deep-water onuphids were mostly described by McIntosh [445] after the HMS Challenger expedition, followed by the report from the USS Albatross Pacific expeditions by Chamberlin [61] and the USNS Eltanin Antarctic deep-sea expeditions by Hartman [448]. A series of works was published by Kucheruk [407,446,470,471] based on the material form the deep-sea Soviet expeditions. Deep-sea onuphids mostly belong to the genera Anchinothria, Hyalinoecia, Leptoecia, and Paradiopatra, and represent the fourth most diverse annelid family below 2000 m depth [472].
The most species-rich genera, Onuphis, Nothria, Hyalinoecia, and Paradiopatra display cosmopolitan distribution, while some others are restricted to more specific regions. Species of Diopatra are mostly found in intertidal and shelf tropical and subtropical areas with very few deep and temperate records. Leptoecia is almost exclusively a bathyal and abyssal genus distributed mainly in the Southern Hemisphere with few records in the North Atlantic and northeastern Pacific [13,17,446]. Australonuphis, a genus comprising seven species, is also known mostly form the Southern Hemisphere, being reported from Australia and South America and with one species known from western Mexico [412]. Until very recently, Mooreonuphis was restricted to the American waters; nonetheless, the detailed investigation of small-bodied onuphids from relatively well-known areas led to the description of two new species from the eastern Atlantic [461,465] and two new species from Australia [458]. Similarly, three new species of Aponuphis, the genus for long time known only from the North and East Atlantic, have been recently described from Australia [457].
Most onuphid species are known to have rather restricted geographical ranges; however, several examples of widely distributed species confirmed with genetic data were reported recently. Diopatra neapolitana originally described from the Mediterranean and widespread in Iberian Atlantic was subsequently reported from the Suez Canal, Egypt, the Brazilian Atlantic, and India [473,474]. Its sister species, Diopatra aciculata Knox and Cameron, 1971, common in Australian east coastal waters, has been recently reported from the South Africa estuaries and also from Egypt. Both species are broadcast spawners with larvae capable of spending some time in the water column. Thus, potential transport with ballast waters of via aquaculture could result in their recent spread due to human activities [46]. Another example of a species with a wide geographical range, but possibly not due to human mediation, is Nothria conchylega, a species commonly found in shelf and upper slope areas in the Arctic Ocean, the North Atlantic and the North Pacific [447]. Recent molecular investigations revealed very little genetic variation in mitochondrial markers in specimens collected across the whole range, indicating the presence of a large panmictic population in the Northern Hemisphere [475].
The vast majority of onuphid species are described based on morphological data and only a few recent studies have provided molecular support for species identification and description. This was almost exclusively done in Diopatra [43,476,477], the most specious genus of onuphids with high morphological variability in diagnostic characteristics. Diopatra neapolitana and D. aciculata are two very similar species, both genetically and morphologically, which display wide but also partly sympatric distribution. Analysis of 22 morphological characteristics and five genetic markers resulted in conclusion that these two species were in the grey speciation zone. Their validity was retained for the stability of the taxonomical system [474]. A similar case has been reported in newly described Diopatra marinae Steiner and Amaral, in Seixas et al., 2020 and Diopatra victoriae Steiner and Amaral, in Seixas et al., 2020 from Brazil [43] confirming previous observations in very rapid and recent speciation within Diopatra (Hektoen 2020 pers. comm.). Diopatra cryptornata Fauchald, Berke and Woodin, 2012 was synonymized with D. neapolitana based on molecular data. The species was originally described due to the presence of dorsal papillae, which were later shown to be sperm storage organs in simultaneous hermaphrodite individuals of D. neapolitana from the northern Spanish coast [427]. Similar papillae were later described in the Brazilian population of D. neapolitana [473].
Cryptic species diversity, although very common in some annelids [478], has not been described as a widespread phenomenon in Onuphidae, possibly due to their complex external morphology allowing distinguishing between closely related species. None of the studies provided clear evidence of cryptic species in onuphids with detailed morphological and genetic assessment of diversity; however, few cases of species complexes are known in Aponuphis [26], Diopatra [479], Hyalinoecia [480] and Nothria [481].
Population level studies are very scarce in onuphids. A study on the deep-sea Hyalinoecia longibranchiata McIntosh, 1885 based on two mitochondrial markers revealed significant population structure between the worms inhabiting several deep-sea sites west of New Zealand, which was hypothesized to be a result of complex current regimes that operate in the area as well as the potential limited ability for larval dispersal [482]. In contrast, the study of Single Nucleotide Polymorphism (SNP) in Australian beach worms, Australonuphis teres, showed no population structure along the eastern coast of Australia, possibly due natural disturbance events, changes in currents and temperature regimes, and/or anthropogenic disturbance [483].
8.6. Exploitation
Large-bodied onuphids are widely used as bait and thus harvested by professional and recreational fishermen from the intertidal zone habitats in Australia (Australonuphis spp. and Hirsutonuphis spp., [53]), South Africa (D. aciculata, [46]), the Mediterranean and Portuguese coasts (D. neapolitana, [54,56,484]). One species, D. aciculata, is commercially produced in aquaculture in New South Wales, Australia and sold as food in the conditioning of prawn broodstock and as dry bait for recreational fishing [59].
9. Future Perspectives
The phylogenetic relationships between eunicidan taxa and the evolution of jaw morphology are still open questions due to the varying placement of most families in different phylogenetic hypotheses, the absence of phylogenetic analyses including both extinct and extant taxa and the highly modified families Histriobdellidae and Hartmaniellidae, as well as lack of phylogenetic hypotheses for most families. More robust phylogenetic hypotheses for Eunicida could allow estimates of time of divergence, providing better understanding of eunicidan evolution as well as calibration points to time estimates within Annelida.
Homology between the different parts of the maxillae is one of the challenges to overcome in order to understand the evolution of jaw morphology and to allow the inclusion of the extinct taxa in the analyses. Paxton [14] undertook a very thorough analysis of morphology and proposed hypotheses of homology for carriers, maxillae I and basal plate, but homologies for other parts are still unclear and in some cases controversial (e.g., Hartmaniellidae). Information on the biomechanics of extant jaw types may also provide another source of information to understand homology among plates and the feeding biology of different species.
The reproductive biology, development and ecology of most families are poorly known despite being key information for understanding the dispersal and distribution of species. Generalizations are made based on knowledge built on few species, which sometimes makes explaining observed distribution patterns a challenge (e.g., [49]). Other factors that hinder the understanding of distribution patterns are the poor information available about the diversity of some regions, such as the South Atlantic, South Pacific and Indian Ocean, and the difficulty in species identification, which may hide continuous distributions or lead to flawed wide distributions. No information is available on the initial infection of the host by eunicidan parasitic and commensal species. Such information would also be of great importance in understanding the evolution of such life modes.
Despite the existence of poorly studied areas, well-studied regions also need to be better explored regarding synonymizations and widely distributed species. Reviews of type material have uncovered diversity hidden under unjustified synonymizations (e.g., [36,122]). Moreover, detailed analyses of morphology and molecular data, especially from type specimens or specimens collected from type localities, improve the understanding of species delimitation, correct use of names and thus of diversity. However, the most important factor of all in order to understand and to reveal the diversity and distribution patterns of taxa is the presence of specialists working on the groups, which generate large leaps in the number of known species (Figure 2). Therefore, the continuous formation of specialists coupled with new views of the morphology, knowledge on the biology and ecology and the use of molecular data appears to be the path to improve the knowledge on the diversity of Eunicida.
Author Contributions
Conceptualization, J.Z., N.B.; writing—sections Introduction, Eunicidae and future perspectives, J.Z.; Dorvilleidae, H.W., A.R.; Hartmaniellidae, Lumbrineridae and Oenonidae, L.F.C.-P.; Histriobdellidae, T.M.S., A.C.Z.A.; Onuphidae, N.B.; original draft preparation, review and editing, J.Z., N.B., L.F.C.-P., T.M.S., A.C.Z.A., H.W., A.R.; and illustrations, N.B. All authors have read and agreed to the published version of the manuscript.
Funding
J.Z. was funded by Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ Proc. E-26/200.071/2019). A.R. was supported by national funds (OE), through FCT (Fundação para a Ciência e a Tecnologia), in the scope of the framework contract foreseen in the Decree-Law 57/2016 (Nrs 4–6, art. 23), changed by Law 57/2017; and through CESAM by FCT/MCTES (UIDP/50017/2020+UIDB/50017/2020). NB was supported by a grant from the Russian Foundation for Basic Research (RFBR project number 19–04-00501).
Acknowledgments
We are thankful to María Capa and Pat Hutchings for organizing this issue and inviting us to contribute, to the three anonymous reviewers who helped to improve the manuscript, and to Alexander Semenov, Conrad Helm and Editora Guanabara Koogan for the permission to use their pictures.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Uschakov, P.V. Phylum Annelida, Class Polychaeta. In Atlas of the Invertebrates of the Far Eastern Seas of the USSR; Pavlovskii, E.N., Ed.; Stony Brook University: Stony Brook, NY, USA, 1955; pp. 109–128. [Google Scholar]
- Dales, R.P. The polychaete stomodeum and the inter-relationships of the families of Polychaeta. Proc. Zool. Soc. Lond. 1962, 139, 389–428. [Google Scholar] [CrossRef]
- Ehlers, E. Die Borstenwürmer (Annelida Chaetopoda) Nach Systematischen und Anatomischen Untersuchungen Dargestellt. Volume II; Wilhelm Engelmann: Leipzig, Germany, 1868; p. 748. [Google Scholar]
- Fauvel, P. Polychètes Errantes. Faune Fr. 1923, 5, 1–488. [Google Scholar]
- Budaeva, N.; Zanol, J. Eunicida Dales, 1962. In Handbook of Zoology Annelida, Volume 3: Sedentaria III, Errantia I; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 353–360. [Google Scholar]
- Struck, T.H.; Golombek, A.; Weigert, A.; Franke, F.A.; Westheide, W.; Purschke, G.; Bleidorn, C.; Halanych, K.M. The Evolution of Annelids Reveals Two Adaptive Routes to the Interstitial Realm. Curr. Biol. 2015, 25, 1993–1999. [Google Scholar] [CrossRef]
- Weigert, A.; Helm, C.; Meyer, M.; Nickel, B.; Arendt, D.; Hausdorf, B.; Santos, S.R.; Halanych, K.M.; Purschke, G.; Bleidorn, C.; et al. Illuminating the base of the Annelid tree using transcriptomics. Mol. Biol. Evol. 2014, 31, 1391–1401. [Google Scholar] [CrossRef] [PubMed]
- Struck, T.H.; Purschke, G.; Halanych, K.M. Phylogeny of Eunicida (Annelida) and exploring data congruence using a partition addition bootstrap alteration (PABA) approach. Syst. Biol. 2006, 55, 1–20. [Google Scholar] [CrossRef]
- Parry, L.A.; Edgecombe, G.D.; Eibye-Jacobsen, D.; Vinther, J. The impact of fossil data on annelid phylogeny inferred from discrete morphological characters. Proc. R. Soc. B Biol. Sci. 2016, 283, 20161378. [Google Scholar] [CrossRef] [PubMed]
- Zanol, J.; Halanych, K.M.; Struck, T.H.; Fauchald, K. Phylogeny of the bristle worm family Eunicidae (Eunicida, Annelida) and the phylogenetic utility of noncongruent 16S, COI and 18S in combined analyses. Mol. Phylogenet. Evol. 2010, 55, 660–676. [Google Scholar] [CrossRef]
- Zanol, J.; Halanych, K.M.; Fauchald, K. Reconciling taxonomy and phylogeny in the bristleworm family Eunicidae (polychaete, Annelida). Zool. Scr. 2014, 43, 79–100. [Google Scholar] [CrossRef]
- Budaeva, N.; Schepetov, D.; Zanol, J.; Neretina, T.; Willassen, E. When molecules support morphology: Phylogenetic reconstruction of the family Onuphidae (Eunicida, Annelida) based on 16S rDNA and 18S rDNA. Mol. Phylogenet. Evol. 2016, 94, 791–801. [Google Scholar] [CrossRef]
- Orensanz, J.M. The Eunicemorph polychaete annelids from Antarctic and Subantarctic Seas. With addenda to the Eunicemorpha of Argentina, Chile, New Zealand, Australia, and the Southern Indian Ocean. Antarct. Res. Ser. 1990, 52, 1–183. [Google Scholar]
- Paxton, H. Phylogeny of Eunicida (Annelida) based on morphology of jaws. Zoosymposia 2009, 264, 241–264. [Google Scholar] [CrossRef]
- Read, G.; Fauchald, K. World Polychaeta Database. Eunicida. Available online: http://www.marinespecies.org/polychaeta/aphia.php?p=taxdetails&id=895 (accessed on 2 December 2020).
- Wiklund, H.; Glover, A.G.; Dahlgren, T.G. Three new species of Ophryotrocha (Annelida: Dorvilleidae) from a whale-fall in the North-East Atlantic. Zootaxa 2009, 2228, 43–56. [Google Scholar] [CrossRef]
- Budaeva, N. Leptoecia midatlantica, a new species of the deep-sea quill-worms (Polychaeta: Onuphidae: Hyalinoeciinae) from the Mid-Atlantic Ridge. Zootaxa 2012, 3176, 45–60. [Google Scholar]
- Yen, N.K.; Rouse, G.W. Phylogeny, biogeography and systematics of Pacific vent, methane seep, and whale-fall Parougia (Dorvilleidae: Annelida), with eight new species. Invertebr. Syst. 2020, 34, 200–233. [Google Scholar]
- Hilbig, B.; Blake, J.A. Dorvilleidae (Annelida: Polychaeta) from the U.S. Atlantic slope and rise. Description of two new genera and 14 new species, with generic revision of Ophryotrocha. Zool. Scr. 1991, 20, 147–183. [Google Scholar] [CrossRef]
- Westheide, W.; Riser, N.W. Morphology and phylogenetic relationships of the neotenic interstitial polychaete Apodotrocha progenerans n.gen., n.sp. (Annelida). Zoomorphology 1983, 103, 67–87. [Google Scholar] [CrossRef]
- Westheide, W.; von Nordheim, H. Interstitial Dorvilleidae (Annelida, Polychaeta) from Europe Australia and New Zealand. Zool. Scr. 1985, 14, 183–199. [Google Scholar] [CrossRef]
- Westheide, W. Ikosipodus carolensis gen. et sp. n., an interstitial neotenic polychaete from North Carolina, U.S.A., and its phylogenetic relationships within Dorvilleidae. Zool. Scr. 1982, 11, 117–126. [Google Scholar] [CrossRef]
- Núñez, J. Marycarmenia lysandrae, a new genus and interstitial species (Polychaeta: Dorvilleidae) from Madeira. Bull. Mar. Sci. 1998, 62, 115–119. [Google Scholar]
- Zanol, J.; da Silva, T.D.S.C.; Hutchings, P. Marphysa (Eunicidae, polychaete, Annelida) species of the Sanguinea group from Australia, with comments on pseudo-cryptic species. Invertebr. Biol. 2016, 135, 328–344. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C.; Carrera-Parra, L.F. Reinstatement of three species of the Marphysa sanguinea complex (Polychaeta: Eunicidae) from the Grand Caribbean Region. Zootaxa 2015, 3925, 37–55. [Google Scholar] [CrossRef] [PubMed]
- Borisova, P.B.; Schepetov, D.M.; Budaeva, N.E. Aponuphis Kucheruk, 1978 (Annelida: Onuphidae) from western African waters. Invertebr. Zool. 2018, 15, 19–41. [Google Scholar] [CrossRef]
- Hints, O.; Eriksson, M.E. Diversification and biogeography of scolecodont-bearing polychaetes in the Ordovician. Palaeogeogr. Palaeoclim. Palaeoecol. 2007, 245, 95–114. [Google Scholar] [CrossRef]
- Hartman, O. Polychaetous Annelids. Part V. Eunicea. Allan Hancock Pac. Exped. 1944, 10, 1–237. [Google Scholar]
- Kielan-Jaworowska, Z. Polychaete jaw apparatuses from the Ordovician and Silurian of Poland and a comparison with modern forms. Palaeontol. Pol. 1966, 16, 9–152. [Google Scholar]
- Eibye-Jacobsen, D.; Kristensen, R.M. A new genus and species of Dorvilleidae (Annelida, Polychaeta) from Bermuda, with a phylogenetic analysis of Dorvilleidae, Iphitimidae and Dinophilidae. Zool. Scr. 1994, 23, 107–131. [Google Scholar] [CrossRef]
- Ravara, A.; Marçal, A.R.; Wiklund, H.; Hilário, A. First account on the diversity of Ophryotrocha (Annelida, Dorvilleidae) from a mammal-fall in the deep-Atlantic Ocean with the description of three new species. Syst. Biodivers. 2015, 13, 875–890. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Phylogeny of the Diopatra generic complex with a revision of Paradiopatra Ehlres, 1887 (Polychaeta: Onuphidae). Zool. J. Linn. Soc. 2011, 163, 319–436. [Google Scholar] [CrossRef]
- Clemo, W.C.; Dorgan, K.M. Functional morphology of eunicidan (Polychaeta) jaws. Biol. Bull. 2017, 233, 227–241. [Google Scholar] [CrossRef]
- Jumars, P.A.; Dorgan, K.M.; Lindsay, S.M. Diet of worms emended: An update of polychaete feeding guilds. Annu. Rev. Mar. Sci. 2015, 7, 497–520. [Google Scholar] [CrossRef]
- Eriksson, M.E.; Parry, L.A.; Rudkin, D.M. Earth’s oldest ‘Bobbit worm’—Gigantism in a Devonian eunicidan polychaete. Sci. Rep. 2017, 7, 43061. [Google Scholar] [CrossRef] [PubMed]
- Fauchald, K. A review of the genus Eunice (Eunicidae: Polychaeta) based upon type material. Smithson. Contrib. Zool. 1992, 523, 1–422. [Google Scholar] [CrossRef]
- Zanol, J. Homology of prostomial and pharyngeal structures in Eunicida (Annelida) based on innervation and morphological similarities. J. Morphol. 2010, 271, 1023–1043. [Google Scholar] [CrossRef] [PubMed]
- Tzetlin, A.; Purschke, G. Pharynx and intestine. Hydrobiologia 2005, 535, 199–225. [Google Scholar]
- Tilic, E.; Bartolomaeus, T.; Rouse, G.W. Chaetal type diversity increases during evolution of Eunicida (Annelida). Org. Divers. Evol. 2016, 16, 105–119. [Google Scholar] [CrossRef]
- Starunov, V. Musculature and innervation of the pygidium in Eunicida (Annelida: Errantia). Biol. Commun. 2019, 64, 31–40. [Google Scholar] [CrossRef]
- Fernández, P.V.; Bone, D.; Rodríguez, C. Cryptofaunal polychaetes from oceanic and continental reefs in Venezuela. Bull. Mar. Sci. 2012, 88, 339–352. [Google Scholar] [CrossRef]
- Hernández-Alcántara, P.; Cruz-Pérez, I.N.; Solís-Weiss, V. Composition and diversity patterns of Eunicida and Amphinomida (Annelida) associated to dead coral in the Chinchorro Bank Biosphere Reserve, Caribbean Sea. J. Mar. Biol. Assoc. UK 2019, 99, 1547–1555. [Google Scholar] [CrossRef]
- Seixas, V.C.; Steiner, T.M.; Solé-Cava, A.M.; Amaral, A.C.Z.; Paiva, P.C. Hidden diversity within the Diopatra cuprea complex (Annelida: Onuphidae): Morphological and genetics analyses reveal four new species in the south-west Atlantic. Zool. J. Linn. Soc. 2020, 1840, 1–35. [Google Scholar] [CrossRef]
- Taboada, S.; Leiva, C.; Bas, M.; Schult, N.; McHugh, D. Cryptic species and colonization processes in Ophryotrocha (Annelida, Dorvilleidae) inhabiting vertebrate remains in the shallow-water Mediterranean. Zool. Scr. 2017, 46, 611–624. [Google Scholar] [CrossRef]
- Hutchings, P.A.; Karageorgopoulos, P. Designation of a neotype of Marphysa sanguinea (Montagu, 1813) and a description of a new species of Marphysa from eastern Australia. Hydrobiologia 2003, 496, 87–94. [Google Scholar] [CrossRef]
- van Rensburg, H.; Matthee, C.A.; Simon, C.A. Moonshine worms Diopatra aciculata (Onuphidae, Annelida) in the Knysna Estuary, South Africa; taxonomy and distribution. J. Mar. Biol. Assoc. UK 2020, 100, 897–907. [Google Scholar] [CrossRef]
- Lavesque, N.; Hutchings, P.; Abe, H.; Daffe, G.; Gunton, L.M.; Glasby, C.J. Confirmation of the exotic status of Marphysa victori Lavesque, Daffe, Bonifácio & Hutchings, 2017 (Annelida) in French waters and synonymy of Marphysa bulla Liu, Hutchings & Kupriyanova, 2018. Aquat. Invasions 2020, 15, 355–366. [Google Scholar] [CrossRef]
- Paxton, H.; Wiklund, H.; Alexander, F.; Taboada, S. Is the Antarctic Ophryotrocha orensanzi (Annelida: Dorvilleidae) a circumpolar non-specialized opportunist? Syst. Biodivers. 2017, 15, 105–114. [Google Scholar] [CrossRef]
- Schulze, A. Phylogeny and genetic diversity of Palolo worms (Palola, Eunicidae) from the tropical North Pacific. Biol. Bull. 2006, 210, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Rouse, G.W. The epitome of hand waving? Larval feeding and hypotheses of metazoan phylogeny. Evol. Dev. 2000, 2, 222–233. [Google Scholar] [CrossRef]
- Steiner, T.M.; Amaral, A.C.Z. Arabella aracaensis, a new species with growth rings on its mandibles, and some remarks on the endoparasitic Labrorostratus prolificus (Polychaeta: Oenonidae) from southeast Brazil. J. Nat. Hist. 2009, 43, 2537–2551. [Google Scholar] [CrossRef]
- Åkesson, B.; Rice, S.A. Two new Dorvillea species (Polychaeta, Dorvilleidae) with obligate asexual reproduction. Zool. Scr. 1992, 21, 351–362. [Google Scholar] [CrossRef]
- Paxton, H. Taxonomy and aspects of the life history of Australian beachworms (Polychaeta: Onuphidae). Aust. J. Mar. Freshw. Res. 1979, 30, 265–294. [Google Scholar] [CrossRef]
- Gambi, M.C. Polychaetes of commercial and applied interest in Italy: An overview. Mémoires Muséum Natl. D’histoire Nat. 1994, 162, 593–601. [Google Scholar]
- Bettoso, N.; Faverio, G.; Orel, G. Note sulla biologia e sulla pesca del verme de rimini (Eunice aphroditois Pallas) in alto Adriatico. Hydrores 1998, 16, 24–34. [Google Scholar]
- Cunha, T.; Hall, A.; Queiroga, H. Estimation of the Diopatra neapolitana annual harvest resulting from digging activity in Canal de Mira, Ria de Aveiro. Fish. Res. 2005, 76, 56–66. [Google Scholar] [CrossRef]
- Osman, I.; Gabr, H.; Saito, H.; El-Etreby, S. Reproductive biology of the highly commercial polychaetes in the Suez Canal. J. Mar. Biol. Assoc. UK 2010, 90, 281–290. [Google Scholar] [CrossRef]
- Osman, I.H.; Gabr, H.R.; El-etreby, S.G. Rearing trials of Halla parthenopeia under laboratory conditions (Polychaeta: Oenonidae). J. Exp. Mar. Biol. Ecol. 2010, 383, 1–7. [Google Scholar] [CrossRef]
- Safarik, M.; Redden, A.M.; Schreider, M.J. Density-dependent growth of the polychaete Diopatra aciculata. Sci. Mar. 2006, 70, 337–341. [Google Scholar]
- Mondragón, C. Of Winds, Worms and Mana: The Traditional Calendar of the Torres Islands, Vanuatu. Oceania 2004, 74, 289–308. [Google Scholar] [CrossRef]
- Chamberlin, R.V. The Annelida Polychaeta. Mem. Mus. Comp. Zool. Harv. Coll. 1919, 48, 1–514. [Google Scholar]
- Núñez, J. Annelida Polychaeta: Staurocephalidae Verrill, 1900. In Annelida Polychaeta V, Fauna Ibérica, Vol. 45; Etal, P.J., Ed.; Museo Nacional de Ciencias Naturales. Consejo Superior de Investigaciones Cientificas: Madrid, Spain, 2018. [Google Scholar]
- Salazar-Vallejo, S.; Carrera-Parra, L.F. Reversal of precedence for Dorvillea Parfitt, 1866 (Annelida, Errantia) over its senior homonym Dorvillea Leach, 1852 (Mollusca, Bivalvia): Maintenance of prevailing usage for Dorvillea Parfitt, 1866 and Dorvilleidae Chamberlin, 1919. Bull. Zool. Nomencl. 2020, 77, 105–111. [Google Scholar] [CrossRef]
- delle Chiaje, S. Memorie Sulla Storia e Notomia Degli Animali Senza Vertebre del Regno di Napoli. Volume 3; Stamperia della Societá Tipografica: Naples, Italy, 1828; p. 232. [Google Scholar]
- Pettibone, M.H. New species of polychaete worms from the Atlantic Ocean, with a revision of the Dorvilleidae. Proc. Biol. Soc. Wash. 1961, 74, 167–186. [Google Scholar]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. III. Dorvilleidae. Physis 1973, 32, 325–342. [Google Scholar]
- Jumars, P.A. A generic revision of the Dorvilleidae (Polychaeta), with six new species from the deep North Pacific. Zool. J. Linn. Soc. 1974, 54, 101–135. [Google Scholar] [CrossRef]
- Wolf, P.S. Four new genera of Dorvilleidae (Annelida: Polychaeta) from the Gulf of Mexico. Proc. Biol. Soc. Wash. 1986, 99, 616–626. [Google Scholar]
- Wiklund, H.; Purschke, G.; Ravara, A. Dorvilleidae Chamberlin, 1919. In Handbook of Zoology Annelida, Volume 3: Sedentaria III, Errantia I; Purschke, G., Böggemann, M., Westheide, W., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 361–382. ISBN 978-3-11-029148-3. [Google Scholar]
- Dahlgren, T.G.; Åkesson, B.; Schander, C.; Halanych, K.M.; Sundberg, P. Molecular Phylogeny of the Model Annelid Ophryotrocha. Biol. Bull. 2001, 201, 193–203. [Google Scholar] [CrossRef][Green Version]
- Heggoy, K.K.; Schander, C.; Åkesson, B. The phylogeny of the annelid genus Ophryotrocha (Dorvilleidae). Mar. Biol. Res. 2007, 3, 412–420. [Google Scholar] [CrossRef]
- Ravara, A.; Wiklund, H.; Cunha, M.R. New species and records of Dorvilleidae (Annelida, Polychaeta) from deep-sea organic substrata, NE Atlantic. Eur. J. Taxon. 2020, in press. [Google Scholar]
- Taboada, S.; Wiklund, H.; Glover, A.G.; Dahlgren, T.G.; Cristobo, J.; Ávila, C. Two new Antarctic Ophryotrocha (Annelida: Dorvilleidae) described from shallow-water whale bones. Polar Biol. 2013, 36, 1031–1045. [Google Scholar] [CrossRef]
- Wiklund, H.; Iris, V.; Glover, A.G.; Smith, C.R.; Baco, A.M.Y.R.; Dahlgren, T.G. Systematics and biodiversity of Ophryotrocha (Annelida, Dorvilleidae) with descriptions of six new species from deep-sea whale-fall and wood-fall habitats in the north-east Pacific. Syst. Biodivers. 2012, 10, 243–259. [Google Scholar] [CrossRef]
- Struck, T.H.; Halanych, K.M.; Purschke, G. Dinophilidae (Annelida) is most likely not a progenetic Eunicida: Evidence from 18S and 28S rDNA. Mol. Phylogenet. Evol. 2005, 37, 619–623. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, R.M.; Eibye-Jacobsen, D. Ultrastructure of spermiogenesis and spermatozoa in Diurodrilus subterraneus (Polychaeta, Diurodrilidae). Zoomorphology 1995, 115, 117–132. [Google Scholar] [CrossRef]
- Laumer, C.E.; Bekkouche, N.; Kerbl, A.; Goetz, F.; Neves, R.C.; Sørensen, M.V.; Kristensen, R.M.; Hejnol, A.; Dunn, C.W.; Giribet, G.; et al. Spiralian Phylogeny Informs the Evolution of Microscopic Lineages. Curr. Biol. 2015, 25, 2000–2006. [Google Scholar] [CrossRef]
- Simonini, R.; Massamba-N’Siala, G.; Grandi, V.; Prevedelli, D. Distribution of the genus Ophryotrocha (Polychaeta) in Italy: New records and comments on the biogeography of Mediterranean species. Vie Milieu-Life Environ. 2009, 59, 79–88. [Google Scholar]
- Åkesson, B. Reproduction and larval morphology of five Ophryotrocha species (Polychaeta, Dorvilleidae). Zool. Scr. 1973, 2, 145–155. [Google Scholar] [CrossRef]
- Åkesson, B. Reproduction in the genus Ophryotrocha (Polychaeta, Dorvilleidae). Pubbl. della Stn. Zool. di Napoli 1975, 39, 377–398. [Google Scholar]
- Oug, E. New and lesser known Dorvilleidae (Annelida, Polychaeta) from Scandinavian and northeast American waters. Sarsia 1978, 63, 285–303. [Google Scholar]
- Purschke, G.; Nowak, K. Ultrastructure of pigmented eyes in Dorvilleidae (Annelida, Errantia, Eunicida) and their importance for understanding the evolution of eyes in polychaetes. Acta Zool. 2015, 96, 67–81. [Google Scholar] [CrossRef]
- Richards, T.L. Reproduction and development of the polychaete Stauronereis rudolphi, including a summary of development in the superfamily Eunicea. Mar. Biol. 1967, 1, 124–133. [Google Scholar] [CrossRef]
- Jamieson, B.G.M.; Rouse, G.W. The spermatozoa of the Polychaeta (Annelida): An ultrastructural review. Biol. Rev. 1989, 64, 93–157. [Google Scholar] [CrossRef]
- Åkesson, B. A new Ophryotrocha species of the Labronica group (Polychaeta, Dorvilleidae) revealed in crossbreeding experiments. In Proceedings of the NATO Conference Series (Marine Science). 2 Marine Organisms, Venice, Italy, 24 March–4 April 1977; Battaglia, B., Beardmore, J., Eds.; Plenum Publishing: New York, NY, USA, 1978; pp. 573–590. [Google Scholar]
- Prevedelli, D.; Simonini, R. Effect of temperature on demography of Ophryotrocha labronica (Polychaeta: Dorvilleidae). Vie Milieu-Life Environ. 2001, 51, 173–180. [Google Scholar]
- Åkesson, B. Ophryotrocha labronica as test animal for the study of marine pollution. Helgoländer Wiss. Meeresunters. 1970, 20, 293–303. [Google Scholar] [CrossRef]
- Simonini, R. Distribution and ecology of the genus Ophryotrocha (Polychaeta: Dorvilleidae) in Italian harbors and lagoons. Vie Milieu-Life Environ. 2002, 2, 59–65. [Google Scholar]
- Paxton, H.; Davey, A. A new species of Ophryotrocha (Annelida: Dorvilleidae) associated with fish farming at Macquarie Harbour, Tasmania, Australia. Zootaxa 2010, 2509, 53–61. [Google Scholar] [CrossRef]
- Salvo, F.; Wiklund, H.; Dufour, S.D.; Hamoutene, D.; Pohle, G.; Worsaae, K. A new annelid species from whalebones in Greenland and aquaculture sites in Newfoundland: Ophryotrocha cyclops, sp. nov. (Eunicida: Dorvilleidae). Zootaxa 2014, 3887, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Simonini, R.; Grandi, V.; Massamba-N’Siala, G.; Pia Martino, M.; Castelli, A.; Prevedelli, D. Diversity, habitat affinities and diet of Ophryotrocha species (Polychaeta, Dorvilleidae) living in Mediterranean harbour habitats. Vie Milieu-Life Environ. 2010, 60, 27–38. [Google Scholar]
- Taboada, S.; Bas, M.; Ávila, C. A new Parougia species (Annelida, Dorvilleidae) associated with eutrophic marine habitats in Antarctica. Polar Biol. 2015, 38, 517–527. [Google Scholar] [CrossRef]
- Detinova, N.N. Polychaetous worms from the Reykjanes Ridge (the North Atlantic) Bottom Fauna from Mid-Ocean Rises in the North Atlantic Donnaya fauna. Otkryto-Okeanicheskikh Pod. Sev. Atl. Tr. Institua Okeanol. 1985, 120, 96–136. [Google Scholar]
- Fauchald, K. Benthic polychaetous annelids from deep water off western Mexico and adjacent areas in the Eastern Pacific Ocean. Allan Hancock Monogr. Mar. Biol. 1972, 7, 1–575. [Google Scholar]
- Hilbig, B. Reconsideration of the genus Exallopus Jumars, 1974 (Polychaeta: Dorvilleidae) with descriptions of two new species from the western North Atlantic. Ophelia 1991, S5, 101–110. [Google Scholar]
- Thornhill, D.J.; Struck, T.H.; Ebbe, B.; Lee, R.W.; Mendoza, G.F.; Levin, L.A.; Halanych, K.M. Adaptive radiation in extremophilic Dorvilleidae (Annelida): Diversification of a single colonizer or multiple independent lineages? Ecol. Evol. 2012, 2, 1958–1970. [Google Scholar] [CrossRef]
- Zhang, D.-S.; Zhou, Y.-D.; Wang, C.-S.; Rouse, G. A new species of Ophryotrocha (Annelida, Eunicida, Dorvilleidae) from hydrothermal vents on the Southwest Indian Ridge. Zookeys 2017, 687, 1–9. [Google Scholar]
- Blake, J.A. Polychaeta from the vicinity of deep-sea geothermal vents in the eastern Pacific. I: Euphrosinidae, Phyllodocidae, Hesionidae, Nereididae, Glyceridae, Dorvilleidae, Orbiniidae and Maldanidae. Bull. Biol. Soc. Wash. 1985, 6, 67–101. [Google Scholar]
- Blake, J.A.; Hilbig, B. Polychaeta from the vicinity of deep-sea hydrothermal vents in the Eastern Pacific Ocean. II. New species and records from the Juan de Fuca and Explorer Ridge systems. Pac. Sci. 1990, 44, 219–253. [Google Scholar]
- Hilbig, B.; Fiege, D. A new species of Dorvilleidae (Annelida: Polychaeta) from a cold seep site in the northeast Pacific. Proc. Biol. Soc. Wash. 2001, 114, 396–402. [Google Scholar]
- Paxton, H.; Morineaux, M. Three species of Dorvilleidae (Annelida: Polychaeta) associated with Atlantic deep-sea reducing habitats, with the description of Ophryotrocha fabriae, new species. Proc. Biol. Soc. Wash. 2009, 122, 14–25. [Google Scholar] [CrossRef]
- Solís-Weiss, V.; Hilbig, B. Redescription of Ophryotrocha platykephale Blake (Polychaeta, Dorvilleidae) from the Guaymas Basin hydrothermal vents. Bull. South. Calif. Acad. Sci. 1992, 91, 92–96. [Google Scholar]
- Fauchald, K. Polychaetous annelids of the families Eunicidae, Lumbrineridae, Iphitimidae, Arabellidae, Lysaretidae and Dorvilleidae from western Mexico. Allan Hancock Monogr. Mar. Biol. 1970, 5, 1–335. [Google Scholar]
- Wolf, P.S. Family Dorvilleidae Chamberlin, 1919. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; Barry A. Vittor & Associates: Mobile, AL, USA, 1984; pp. 44.1–44.37. [Google Scholar]
- Wolf, P.S. Three new species of Dorvilleidae (Annelida: Polychaeta) from Puerto Rico and Florida and a new genus for dorvillleids from Scandinavia and North America. Proc. Biol. Soc. Wash. 1986, 99, 627–638. [Google Scholar]
- Hilbig, B. Family Dorvilleidae Chamberlin, 1919. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and the Western Santa Barbara Channel. Vol. 5. The Annelida Part 2, Polychaeta: Phyllodocida (Syllidae and Scale-Bearing Families), Amphinomida, and Eunicida; Lake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History Publications: Santa Barbara, CA, USA, 1995; pp. 341–364. [Google Scholar]
- Paiva, P.C.; Nonato, E.F. On the genus Iphitime (Polychaeta: Iphitimidae) and description of Iphitime sartorae sp. nov. a commensal of brachyuran crabs. Ophelia 1991, 34, 209–215. [Google Scholar] [CrossRef]
- Imajima, M. Dorvilleidae (Annelida, Polychaeta) from Japan. I. The genus Dorvillea (Dorvillea). Bull. Natl. Sci. Mus. Tokyo 1992, 18, 131–147. [Google Scholar]
- Núñez, J.; Pascual, M.; Brito, M.C. Contribución al estudio de los Dorvilleidos (Annelida, Polychaeta) de Canarias. Rev. Acad. Canar. Cienc. 1996, 8, 139–151. [Google Scholar]
- Hartmann-Schröder, G.; Rosenfeldt, P. Die Polychaeten der ’Polarstern’-Reise ANT V/1 in die Antarktis 1986. Teil 1: Euphrosinidae bis Iphitimidae. Mitt. Hambg. Zool. Mus. Inst. 1992, 89, 85–124. [Google Scholar]
- Cunha, M.R.; Matos, F.L.; Génio, L.; Hilário, A.; Moura, C.J.; Ravara, A.; Rodrigues, C.F. Are organic falls bridging reduced environments in the deep sea?—Results from colonization experiments in the Gulf of Cádiz. PLoS ONE 2013, 8, e76688. [Google Scholar] [CrossRef] [PubMed]
- Levin, L.A.; Ziebis, W.; Mendoza, G.F.; Growney, V.A.; Tryon, M.D.; Brown, K.M.; Mahn, C.; Gieskes, J.M.; Rathburn, A.E. Spatial heterogeneity of macrofauna at northern California methane seeps: Influence of sulfide concentration and fluid flow. Mar. Ecol. Prog. Ser. 2003, 265, 123–139. [Google Scholar] [CrossRef]
- Prevedelli, D.; Massamba-N’Siala, G.; Simonini, R. The seasonal dynamics of six species of Dorvilleidae (Polychaeta) in the harbour of La Spezia (Italy). Mar. Ecol. Evol. Perspect. 2005, 26, 286–293. [Google Scholar] [CrossRef]
- Fauchald, K.; Hancock, D.R. Deep-water polychaetes from a transect off central Oregon. Allan Hancock Monogr. Mar. Biol. 1981, 11, 1–73. [Google Scholar]
- Hartmann-Schröder, G. Die polychaeten der Fahrten 11, 19, 21 und 26 (1967-1972) von F.S. “Meteor” in das Gebiet zwischen dem Golf von Biscaya und dem Auftriebsgebiet vor Westafrika. Meteor Forsch.-Ergeb. 1981, 33, 23–36. [Google Scholar]
- Schulze, A.; Timm, L.E. Palolo and un: Distinct clades in the genus Palola (Eunicidae: Polychaeta). Mar. Biodivers. 2011, 42, 161–171. [Google Scholar] [CrossRef]
- Nederlof, M.A.J.; Jansen, H.M.; Dahlgren, T.G.; Fang, J.; Meier, S.; Strand, Ø.; Sveier, H.; Verdegem, M.C.J.; Smaal, A.C. Application of polychaetes in (de)coupled integrated aquaculture: Production of a high-quality marine resource. Aquac. Environ. Interact. 2019, 11, 221–237. [Google Scholar] [CrossRef]
- Åkesson, B. The use of certain polychaetes in bioassay studies. Rapp. Proces Verbaux Reun. 1980, 179, 315–321. [Google Scholar]
- Lamarck, J.B. Histoire Naturelle des Animaux Sans Vertèbres, Préséntant les Caractères Généraux et Particuliers de ces Animaux, Leur Distribution, Leurs Classes, Leurs Familles, Leurs Genres, et la Citation des Principales Espèces qui s’y Rapportent; Precedes d’une Introduction Offrant la Determination des Caracteres Essentiels de L’animal, sa Distinction du Vegetal et Desautres Corps Naturels, Enfin, l’Exposition des Principes Fondamentaux de la Zoologie. Vol. 5; Deterville Libraire & Verdiere Libraire: Paris, France, 1818; p. 612. [Google Scholar]
- Hartmann-Schröder, G.; Zibrowius, H. Polychaeta associated with Antipatharia (Cnidaria: Anthozoa): Description of Polynoidae and Eunicidae. Mitt. Hambg. Zool. Mus. Inst. 1998, 95, 29–44. [Google Scholar]
- Zanol, J.; Fauchald, K.; Paiva, P.C. A phylogenetic analysis of the genus Eunice (Eunicidae, polychaete, Annelida). Zool. J. Linn. Soc. 2007, 150, 413–434. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C.; Carrera-Parra, L.F. Revision of Marphysa de Quatrefages, 1865 and some species of Nicidion Kinberg, 1865 with the erection of a new genus (Polychaeta: Eunicidae) from the Grand Caribbean. Zootaxa 2017, 4241, 1–62. [Google Scholar] [CrossRef]
- Lu, H.; Fauchald, K. A phylogenetic and biogeographic study of Euniphysa (Eunicidae, Polychaeta). J. Nat. Hist. 2000, 34, 997–1044. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F.; Fauchald, K.; Gambi, M.C. Revision of the taxonomic status of Lysidice (Polychaeta, Eunicidae) in the Western Caribbean Sea with observation on species reproductive features and habitat preference. Ital. J. Zool. 2011, 78, 27–40. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C.; Idris, I. Reinstatement of species belonging Marphysa sanguinea complex (Annelida: Eunicidae) and description of new species from the mid-Pacific Ocean and the Adriatic Sea. Zootaxa 2020, 4816, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Kara, J.; Molina-Acevedo, I.C.; Zanol, J.; Simon, C.; Idris, I. Morphological and molecular systematic review of Marphysa Quatrefages, 1865 (Annelida: Eunicidae) species from South Africa. PeerJ 2020, 8, e10076. [Google Scholar] [CrossRef] [PubMed]
- Fauchald, K. Review of the types of Palola (Eunicidae: Polychaeta). J. Nat. Hist. 1992, 26, 1177–1225. [Google Scholar] [CrossRef]
- Molina-Acevedo, I.C. Morphological revision of the Subgroup 1 Fauchald, 1970 of Marphysa de Quatrefages, 1865 (Eunicidae: Polychaeta). Zootaxa 2018, 4480, 1–125. [Google Scholar] [CrossRef] [PubMed]
- Molina-Acevedo, I.C. Redescription of two species previously regarded as Marphysa de Quatrefages, 1865, with the description of new species of Treadwellphysa Molina-Acevedo and Carrera-Parra, 2017. J. Nat. Hist. 2019, 53, 517–540. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F.; Salazar-Vallejo, S.I. A new genus and 12 new species of Eunicidae (Polychaeta) from the Caribbean Sea. J. Mar. Biol. Assoc. UK 1998, 78, 145–182. [Google Scholar] [CrossRef]
- Glasby, C.J.; Hutchings, P.A. A new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) including the genus Nauphanta Kinberg, 1865. Zootaxa 2010, 45, 29–45. [Google Scholar] [CrossRef]
- Martin, D.; Gil, J.; Zanol, J.; Meca, M.A.; Portela, R.P. Digging the diversity of Iberian bait worms Marphysa (Annelida, Eunicidae). PLoS ONE 2020, 15, e0226749. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.M.; Nogueira, J.M.D.M.; Amaral, A.C.Z. Eunice rubra Grube (Annelida, Polychaeta, Eunicidae), a redescription based on the holotype and Brazilian specimens. Rev. Bras. Zool. 2002, 19, 243–259. [Google Scholar] [CrossRef]
- Parapar, J.; Freire, J.; Urgorri, V.; Besteiro, C. Morphological variability in Eunice vittata (Chiaje, 1828) (Polychaeta, Eunicidae) in the Ría de Ferrol (Galicia, NW Spain). Ophelia 1993, 37, 117–125. [Google Scholar] [CrossRef]
- Miura, T. Japanese polychaetes of the genera Eunice and Euniphysa: Taxonomy and branchial distribution patterns. Publ. Seto Mar. Biol. Lab. 1986, 31, 269–325. [Google Scholar] [CrossRef][Green Version]
- Salazar-Vallejo, S.I.; Carrera-Parra, L.F. Eunícidos (Polychaeta) del caribe mexicano con claves para las especies del Gran Caribe: Fauchaldius, Lysidice, Marphysa, Nematonereis y Palola. Rev. Biol. Trop. 1998, 45, 1481–1498. [Google Scholar]
- Parapar, J.; Harto, I. On the presence in the Iberian waters and morphological variability in Eunice roussaei Quatrefages, 1866 (Polychaeta, Eunicidae). Bol. Real Soc. Esp. Hist. Nat. Secc. Biol. 2001, 96, 165–174. [Google Scholar]
- Costa-Paiva, E.M.; Paiva, P.C. A morphometric analysis of Eunice Cuvier (Annelida, Polychaeta) species. Rev. Bras. Zool. 2007, 24, 353–358. [Google Scholar] [CrossRef]
- Treadwell, A.L. Leodicidae of the West Indian region. Carnegie Inst. Wash. Publ. 1921, 293, 1–131. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Eunicidae Berthold, 1827. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Parra, L.F.C., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 165–181. [Google Scholar]
- Zanol, J.; Budaeva, N. Eunicidae. In Handbook of Zoology Annelida, Volume 3: Sedentaria III, Errantia I; Purschke, G., Westheide, W., Böggemann, M., Eds.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2021; pp. 414–452. [Google Scholar]
- Lachat, J.; Haag-Wackernagel, D. Novel mobbing strategies of a fish population against a sessile annelid predator. Sci. Rep. 2016, 6, 33187. [Google Scholar] [CrossRef]
- Hofmann, D.K. Nachweise der Epitokie als Fortpflanzungsmodus des Polychaeten Eunice siciliensis (Polychaeta: Annelida). Mar. Biol. 1972, 11, 341–344. [Google Scholar] [CrossRef]
- Pamungkas, J. Species richness and macronutrient content of wawo worms (Polychaeta, Annelida) from Ambonese waters, Maluku, Indonesia. Biodiveristy Data J. 2015, 3, e4251. [Google Scholar] [CrossRef] [PubMed]
- Southern, R. Polychaeta of the Chilka Lake and also of fresh and brackish water in other parts of India. Mem. Indian Mus. 1921, 4, 563–659. [Google Scholar]
- Aiyar, R.G. An account of the development and breeding habits of a brackish water Polychaete worm of the genus Marphysa. J. Linn. Soc. Lond. 1931, 37, 387–403. [Google Scholar] [CrossRef]
- Pillai, T.G. Studies on a brackish-water polychaetous annelid, Marphysa borradailei, sp. n. from Ceylon. Ceylon J. Sci. (Biol. Sci.) 1958, 1, 94–106. [Google Scholar]
- Pettibone, M.H. Marine polychate worms of the New England region Part 1. Aphroditidae through Trochochetidae. United States Natl. Mus. Bull. 1963, 227, 1–356. [Google Scholar]
- Åkesson, B. The embryology of the Polychaete Eunice kobiensis. Acta Zool. 1967, 48, 141–192. [Google Scholar] [CrossRef]
- Prevedelli, D.; N’siala, G.M.; Ansaloni, I.; Simonini, R. Life cycle of Marphysa sanguinea (Polychaeta: Eunicidae) in the Venice Lagoon (Italy). Mar. Ecol. 2007, 28, 384–393. [Google Scholar] [CrossRef]
- Malathi, E.; Sunder Raj, S.K.; Mercy Bai, P. Larval development of Marphysa gravelyi (Polychaeta: Eunicidae) from Pulicat Lake, India. Ital. J. Zool. 2011, 78, 249–254. [Google Scholar] [CrossRef]
- Herpin, R. Recherches biologiques sur la reproduction et le développement de quelques Annélides polychètes. Bull. Société Sci. Nat. L’ouest Fr. 1925, 5, 1–250. [Google Scholar]
- Goldstein, M.C.; Carson, H.S.; Eriksen, M. Relationship of diversity and habitat area in North Pacific plastic-associated rafting communities. Mar. Biol. 2014, 161, 1441–1453. [Google Scholar] [CrossRef]
- Jang, M.; Shim, W.J.; Han, G.M.; Song, Y.K.; Hong, S.H. Formation of microplastics by polychaetes (Marphysa sanguinea) inhabiting expanded polystyrene marine debris. Mar. Pollut. Bull. 2018, 131, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Díaz, O.; Liñero-Arana, I. Epifaunal polychaetes on Pinctada imbricata Roding, 1798 (Bivalvia: Pteriidae) from the Gulf of Cariaco Venezuela. Interciencia 2003, 28, 298–301. [Google Scholar]
- Hartman, O. Descriptions of new species and new generic records of polychaetous annelids from California of the families Glyceridae, Eunicidae, Stauronereidae, and Opheliidae. Univ. Calif. Publ. Zool. 1938, 43, 93–111. [Google Scholar]
- Gambi, M.C. Spatio-temporal distribution and ecological role of polychaete borers of Posidonia oceanica (L.) Delile scales. Bull. Mar. Sci. 2002, 71, 1323–1331. [Google Scholar]
- Molodtsova, T.; Budaeva, N. Modifications of corallum morphology in black corals as an effect of associated fauna. Bull. Mar. Sci. 2007, 81, 469–479. [Google Scholar]
- Quiroga, E.; Levin, L.A. Eunice pennata (Polychaeta: Eunicidae) From Active and Passive Cold Seep Sites in Central and Southern Chile (36–46° S). An. Inst. Patagon. 2010, 38, 31–37. [Google Scholar] [CrossRef]
- Mueller, C.E.; Lundälv, T.; Middelburg, J.J.; van Oevelen, D. The Symbiosis between Lophelia pertusa and Eunice norvegica stimulates coral calcification and worm assimilation. PLoS ONE 2013, 8, e58660. [Google Scholar] [CrossRef]
- Kise, H.; Reimer, J.D. Unexpected diversity and a new species of Epizoanthus (Anthozoa, hexacorallia) attached to eunicid worm tubes from the Pacific Ocean. Zookeys 2016, 2016, 49–71. [Google Scholar] [CrossRef]
- Roberts, J.M. Reef-aggregating behaviour by symbiotic eunicid polychaetes from cold-water corals: Do worms assemble reefs? J. Mar. Biol. Assoc. UK 2005, 85, 813–819. [Google Scholar] [CrossRef]
- Hutchings, P.A. Biological destruction of coral reefs. Coral Reefs 1986, 4, 239–252. [Google Scholar] [CrossRef]
- Chisholm, J.R.M.; Kelley, R. Worms start the reef-building process. Nature 2001, 409, 152. [Google Scholar] [CrossRef]
- Ochoa-Rivera, V.; Granados-Barba, A.; Solís-Weiss, V. The polychaete cryptofauna from Cozumel Island, Mexican Caribbean. Bull. Mar. Sci. 2000, 67, 137–146. [Google Scholar]
- Hartman, O. Polychaeta Errantia of Antarctica. Antarct. Res. Ser. 1964, 3, 1–131. [Google Scholar]
- Winsnes, I.M. Eunicid polychaetes (Annelida) from Scandinavian and adjacent waters. Family Eunicidae. Zool. Scr. 1989, 18, 483–500. [Google Scholar] [CrossRef]
- Amoureux, L. Annélides Polychètes profondes de Madagascar Description de deux nouvelles espèces (Collections Crosnier et Jouannie). Bull. Muséum Natl. d’Histoire Nat. 1978, 344, 1093–1108. [Google Scholar]
- Nonato, E.F. Contribuição ao Conhecimento dos Anelídeos Poliquetas Bentônicos da Plataforma Continental Brasileira, Entre Cabo Frio e o Arroio Chuí. Livre Docência Thesis, Universidade de São Paulo, São Paulo, Brazil, 1981. [Google Scholar]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. VII. Eunicidae y Lysaretidae. Physis 1975, 34, 85–111. [Google Scholar]
- Fauchald, K. Review of the types and key to the species of Eunice (Eunicidae: Polychaeta) from the Australian region. Rec. Aust. Mus. 1986, 38, 241–262. [Google Scholar] [CrossRef]
- Zanol, J.; da Silva, T.S.C.; Hutchings, P. One new species and two redescriptions of Marphysa (Eunicidae, Annelida) species of the Aenea-group from Australia. Zootaxa 2017, 4268, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Zanol, J.; Hutchings, P.A.; Fauchald, K. Eunice sensu latu (Annelida: Eunicidae) from Australia: Description of seven new species and comments on previously reported species of the genera Eunice, Leodice and Nicidion. Zootaxa 2020, 4748, 1–43. [Google Scholar] [CrossRef]
- Schulze, A. Six genetically distinct clades of Palola (Eunicidae, Annelida) from Lizard Island, Great Barrier Reef, Australia. Zootaxa 2015, 4019, 695–706. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kott, P. Nereidae and Eunicidae of south Western Australia; also notes on the ecology of Western Australian limestone reefs. J. R. Soc. West. Aust. 1951, 35, 85–130. [Google Scholar]
- Morgado, E.H.; Amaral, A.C.Z. Anelidos poliquetos associados a um briozoario I. Eunicidae, Lumbrineridae, Lysaretidae e Dorvilleidae. Iheringia 1981, 60, 33–54. [Google Scholar]
- Steiner, T.; Amaral, A.C.Z. Two New Species of Marphysa Quatrefages, 1865 (Eunicidae, Polychaeta) From Intertidal Sandy Beaches of the São Sebastião Channel, State of São Paulo (Brazil). Bull. Mar. Sci. 2000, 67, 479–489. [Google Scholar]
- Zanol, J.; Paiva, P.C.; Attolini, F.S. Eunice and Palola (Eunicidae: Polychaeta) from the eastern Brazilian Coast (13°00′–22°30′ S). Bull. Mar. Sci. 2000, 67, 449–463. [Google Scholar]
- Nogueira, J.M.M.; Steiner, T.M.; Amaral, A.C.Z. Descriptions of two new species of Eunice Cuvier, 1817 (Polychaeta: Eunicidae) from coastal islands of the State of São Paulo, Brazil. Sci. Mar. 2001, 65, 47–57. [Google Scholar] [CrossRef]
- Bergamo, G.; Carrerette, O.; Zanol, J.; Nogueira, J.M.M. Species of Eunicidae (Annelida, Errantia, Eunicida) from rhodolith beds off the states of Paraíba and Pernambuco, northeastern Brazilian coast. Zootaxa 2018, 4521, 376–390. [Google Scholar] [CrossRef] [PubMed]
- George, J.D.; Hartmann-Schröder, G. Polychaetes: British Amphinomida, Spintherida and Eunicida. Keys and notes for the identification of the species. Synop. Br. Fauna 1985, 32, 1–221. [Google Scholar]
- Núñez, J.; Pascual, M.; Delgado, J.D.; Brito, M.C. Anélidos poliquetos de Canarias: Familia Eunicidae. Vieraea 1997, 26, 47–75. [Google Scholar]
- Carrera-Parra, L.F.; Salazar-Vallejo, S.I. Eunícidos (Polychaeta) del Caribe mexicano con claves para las especies del Gran Caribe: Eunice. Rev. Biol. Trop. 1998, 45, 1499–1521. [Google Scholar]
- Salazar-Vallejo, S.I.; Carrera-Parra, L.F.; de León-González, J.A. Giant Eunicid Polychaetes (Annelida) in shallow tropical and temperate seas. Rev. Biol. Trop. 2011, 59, 1463–1474. [Google Scholar] [CrossRef][Green Version]
- Carrera-Parra, L.F.; Salazar-Vallejo, S.I. Redescriptions of Eunice filamentosa and E. denticulata and description of E. tovarae n. sp. (Polychaeta: Eunicidae), highlighted with morphological and molecular data. Zootaxa 2011, 2880, 51–64. [Google Scholar] [CrossRef]
- de Léon-González, J.A. A new eunicid polychaete from the Caribbean Coast of Mexico. Rev. Biol. Trop. 1988, 36, 75–79. [Google Scholar]
- Liu, Y.; Hutchings, P.; Sun, S. Three new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) from the south coast of China and redescription of Marphysa sinensis Monro, 1934. Zootaxa 2017, 4263, 228–250. [Google Scholar] [PubMed]
- Liu, Y.; Hutchings, P.; Kupriyanova, E. Two new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicida: Eunicidae) from northern coast of China and redescription for Marphysa orientalis Treadwell, 1936. Zootaxa 2018, 4377, 191. [Google Scholar] [CrossRef] [PubMed]
- de León-González, J.A.; Rivera, C.G.; Romero, M.Y. Sublittoral Eunicidae and Onuphidae (Polychaeta) from softbottom of El Salvador, eastern Pacific. J. Mar. Biol. Assoc. UK 2004, 84, 93–101. [Google Scholar]
- Treadwell, A.L. Leodicidae from Fiji and Samoa. Carnegie Inst. Washingt. Publ. 1922, 312, 127–170. [Google Scholar]
- Lavesque, N.; Daffe, G.; Bonifácio, P.; Hutchings, P. A new species of the Marphysa sanguinea complex from French waters (Bay of Biscay, NE Atlantic) (Annelida, Eunicidae). Zookeys 2017, 716, 1–17. [Google Scholar] [CrossRef]
- Gathof, J.M. Family Eunicidae Savigny, 1818. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; pp. 40.1–40.31. [Google Scholar]
- de León-González, J.Á.; Díaz-Castañeda, V. Eunicidae (Annelida: Polychaeta) associated with Phragmathopoma caudata Morch, 1863 and some coral reefs from Veracruz, Gulf of Mexico. Sci. Mar. 2006, 70, 91–99. [Google Scholar] [CrossRef][Green Version]
- Wang, Z.; Zhang, Y.; Qiu, J. A New Species in the Marphysa sanguinea Complex (Annelida, Eunicidae) from Hong Kong. Zool. Stud. 2018, 48, 1–13. [Google Scholar] [CrossRef]
- Arias, A.; Fernández-Álvarez, F.Á.; Martins, R.; Anadón, N. Rediscovery and redescription of Leodice laurillardi (Quatrefages, 1866) comb. nov. (Annelida: Eunicidae)—A rare European polychaete or just an overlooked species? Zootaxa 2015, 3964, 475–481. [Google Scholar] [CrossRef][Green Version]
- Hutchings, P.; Lavesque, N.; Priscilla, L.; Daffe, G.; Malathi, E.; Glasby, C.J. A new species of Marphysa (Annelida: Eunicida: Eunicidae) from India, with notes on previously described or reported species from the region. Zootaxa 2020, 4852, 285–308. [Google Scholar] [CrossRef] [PubMed]
- Veeramuthu, S.; Ramadoss, R.; Subramaniyan, B.; Jeyaram, S.; Fernando, O.J. Abundance of the Boring Polychaetes of Eunicidae (Annelida) in Great Nicobar Islands. Our Nat. 2013, 10, 76–88. [Google Scholar] [CrossRef]
- Miura, T. Eunicid polychaetous annelids from Japan—I. La Mer (Bulletin la Société Fr. D’océanographie) 1977, 15, 1–20. [Google Scholar]
- Miura, T. Eunicid polychaetous annelids from Japan—II. La Mer (Bulletin la Société Fr. D’océanographie) 1977, 15, 61–81. [Google Scholar]
- Miura, T. Eunicid polychaetous annelids from Japan—III. La Mer (Bulletin la Société Fr. D’océanographie) 1979, 17, 33–42. [Google Scholar]
- Miura, T. New or little known species of the Family Eunicidae (Annelida, Polychaeta) from Japan. Proc. Jpn. Soc. Syst. Zool. 1987, 36, 1–9. [Google Scholar]
- Abe, H.; Tanaka, M.; Taru, M.; Abe, S.; Nishigaki, A. Molecular evidence for the existence of five cryptic species within the Japanese species of Marphysa (Annelida: Eunicidae) known as “iwa-mushi”. Plankt. Benthos Res. 2019, 14, 303–314. [Google Scholar] [CrossRef]
- Choi, H.K.; Kim, J.G.; Kang, D.W.; Yoon, S.M. A new species of Leodice from Korean waters (Annelida, Polychaeta, Eunicidae). Zookeys 2017, 2017, 53–67. [Google Scholar] [CrossRef][Green Version]
- Idris, I.; Hutchings, P.; Arshad, A. Description of a new species of Marphysa Quatrefages, 1865 (Polychaeta: Eunicidae) from the west coast of Peninsular Malaysia and comparisons with species from Marphysa Group A from the Indo-West Pacific and Indian Ocean. Mem. Mus. Vic. 2014, 71, 109–121. [Google Scholar] [CrossRef]
- Katsiaras, N.; Simboura, N.; Koutsoubas, D. The rare subgroup C1 of Marphysa (Polychaeta, Eunicidae): Re-description of species and first records in the Mediterranean Sea. Zootaxa 2014, 3873, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Kurt-Sahin, G.; Çinar, M.E. Eunicidae (Polychaeta) species in and around Iskenderun Bay (Levantine Sea, Eastern Mediterranean) with a new alien species for the Mediterranean Sea and a re-description of Lysidice collaris. Turk. J. Zool. 2009, 33, 331–347. [Google Scholar] [CrossRef]
- Kurt-Sahin, G.; Çinar, M.E. Presence of Marphysa disjuncta (Polychaeta: Eunicidae) in the Mediterranean Sea. Mediterr. Mar. Sci. 2009, 10, 145–150. [Google Scholar] [CrossRef][Green Version]
- Kurt-Sahin, G. Marphysa cinari, a new species of Eunicidae (Polychaeta) from the coasts of Turkey (eastern Mediterranean) and re-descriptions of Marphysa kinbergi McIntosh, 1910 and Marphysa disjuncta Hartman, 1961. J. Nat. Hist. 2014, 48, 1989–2006. [Google Scholar] [CrossRef]
- Kurt-Sahin, G.; Çinar, M.E. Distribution of Eunicidae (Annelida: Polychaeta) along the Levantine coast of Turkey, with special emphasis on alien species. Mar. Biodivers. 2017, 47, 421–431. [Google Scholar] [CrossRef]
- Cantone, G. Censimento dei policheti dei mari italiani: Eunicidae Berthold, 1827. Atti Soc. Toscana Sci. Nat. Resid. Pisa Mem. Ser. B 1993, 100, 229–243. [Google Scholar]
- de León-González, J.A. Eunice orensanzi n.sp. from the western coast of Baja California Sur, México and key to the Mexican Eunice (Polychaeta: Eunicidae). Rev. Biol. Trop. 1990, 38, 259–266. [Google Scholar]
- Ruiz-Cancino, G.; Carrera-Parra, L.F.; Bastida-Zavala, R. Eunícidos (Polychaeta: Eunicidae) del Pacífico sur de México. Ciencia y Mar 2010, 14, 27–60. [Google Scholar]
- Knox, G.A.; Green, K.M. The polychaetous annelids of New Zealand Part 4. Eunicidae. J. R. Soc. N. Z. 1972, 2, 459–470. [Google Scholar] [CrossRef]
- Glasby, C.J.; Mandario, M.A.E.; Burghardt, I.; Kupriyanova, E.; Gunton, L.M.; Hutchings, P.A. A new species of the sanguinea-group Quatrefages, 1866 (Annelida: Eunicidae: Marphysa) from the Philippines. Zootaxa 2019, 4674, 264–282. [Google Scholar] [CrossRef]
- Díaz, A.; López, E. Four New Records of Eunicidae (Annelida: Polychaeta) from the Western Philippines Islands. Thalassas 2020, 36, 321–332. [Google Scholar] [CrossRef]
- Day, J.H. A Monograph on the Polychaeta of Southern Africa. Part I. Errantia; Trustees of the British Museum (Natural History): London, UK, 1967; p. 458. [Google Scholar]
- Lewis, C.; Karageorgopoulos, P. A new species of Marphysa (Eunicidae) from the western Cape of South Africa. J. Mar. Biol. Assoc. UK 2008, 88, 277–287. [Google Scholar] [CrossRef]
- Wu, Q.Q.; He, M.H. A new genus and new species of Eunicidae from Taiwan Strait. Acta Zootaxon. Sin. 1988, 13, 123–126. [Google Scholar]
- Hsueh, P.-W.; Li, Y.-H. New species and new records of eunicids (Polychaeta, Eunicidae) from Taiwan. Zootaxa 2014, 3802, 151–172. [Google Scholar] [CrossRef] [PubMed]
- Ehlers, E. Reports on the results of dredging, under the direction of L. F. Pourtalès, during the years 1868-1870, and of Alexander Agassiz, in the Gulf of Mexico (1877-78), and in the Caribbean Sea (1878-79), in the U.S. Coast Survey steamer “Blake”, Lieut-Com. C. D. Sigsbee, U.S.N. and Commander J. R. Bartlett, U.S.N., commanding. XXXI. Report on the Annelids. Mem. Mus. Comp. Zool. Harv. Coll. 1887, 15, 1–335. [Google Scholar]
- Ebbs, N.K. The coral-inhabiting polychaetes of the northern Florida reef tract. Part I. Aphroditidae, Polynoidae, Amphinomidae, Eunicidae and Lysaretidae. Bull. Mar. Sci. 1966, 16, 485–555. [Google Scholar]
- Hartman, O. Polychaetous annelids from California, including the descriptions of two new genera and nine new species. Allan Hancock Pac. Exped. 1944, 10, 239–307. [Google Scholar]
- Liñero-Arana, I. Poliquetos errantes bentónicos de la platforma continental nor-oriental de Venezuela. II. Eunicidae. Bol. Inst. Oceanogr. Venez. Univ. Oriente 1990, 24, 91–103. [Google Scholar]
- Díaz-Díaz, O.F.; Bone, D.; Rodríguez, C.T.; López, A. Polychaetes associated to calcareous sediments, Venezuela: Eunicidae. Panam. J. Aquat. Sci. 2016, 11, 18–25. [Google Scholar]
- Iannotta, M.A.; Gambi, M.C.; Patti, F.P. Molecular evidence of intraspecific variability in Lysidice ninetta (Polychaeta: Eunicidae) in the Mediterranean Sea. Aquat. Biol. 2009, 6, 121–132. [Google Scholar] [CrossRef]
- Schulze, A. The Bobbit worm dilemma: A case for DNA. Rev. Biol. Trop. 2011, 59, 1475–1477. [Google Scholar]
- Hutchings, P.; Kupriyanova, E. Cosmopolitan polychaetes—Fact or fiction? Personal and historical perspectives. Invertebr. Syst. 2017, 32, 1–9. [Google Scholar] [CrossRef]
- Iannotta, M.A.; Patti, F.P.; Ambrosino, M.; Procaccini, G.; Gambi, M. Phylogeography of two species of Lysidice (Polychaeta, Eunicidae) associated to the seagrass Posidonia oceanica in the Mediterranean Sea. Mar. Biol. 2007, 150, 1115–1126. [Google Scholar] [CrossRef]
- Martin, D. Anélidos poliquetos asociados a las concreciones de algas calcáreas del litoral Catalán. Miscellània Zoològica Barc. 1987, 11, 61–75. [Google Scholar]
- Lavesque, N.; Daffe, G.; Grall, J.; Zanol, J.; Gouillieux, B.; Hutchings, P. Guess who? On the importance of using appropriate name: Case study of Marphysa sanguinea (Montagu, 1813). Zookeys 2019, 859, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Pettibone, M.H. Annelida. In Synopsis and Classification of Living Organisms. 2; Parker, S.P., Ed.; McGraw-Hill Book Co.: New York, NY, USA, 1982; pp. 1–43. [Google Scholar]
- Carrera-Parra, L.F. Redescription of Hartmaniella tulearensis n. comb. (Amoureux, 1978) with comments on Hartmaniella sp and affinities of the family (Polychaeta: Hartmaniellidae). J. Nat. Hist. 2003, 37, 49–55. [Google Scholar] [CrossRef]
- Gathof, J.M. Family Hartmaniellidae Imajima, 1977. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VII; Uebelacker, J.M., Johnson, P.G., Eds.; Barry A. Vittor & Associates Inc.: Mobile, AL, USA, 1984; pp. 57.1–57.3. [Google Scholar]
- Eibye-Jacobsen, D.; Oug, E. A report of Hartmaniellidae (Annelida: Polychaeta) from the Andaman Sea, Indian Ocean. Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 101–114. [Google Scholar]
- Al-Hakim, I.; Glasby, C.J. Polychaeta (Annelida) of the Natuna Islands, South China Sea. Raffles Bull. Zool. 2004, 11, 25–45. [Google Scholar]
- Imajima, M. A new polychaete family, Hartmaniellidae, from Japan. In Essays on Polychaetous Annelids in Memory of Dr. Olga Hartman; Reish, D.J., Fauchald, K., Eds.; The Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1977; pp. 211–216. [Google Scholar]
- Fauchald, K.; Rouse, G.W. Polychaete systematics: Past and present. Zool. Scr. 1997, 26, 71–138. [Google Scholar] [CrossRef]
- Szaniawski, H.; Imajima, M. Hartmaniellidae—Living fossils among polychaetes. Acta Palaeontol. Pol. 1996, 41, 111–125. [Google Scholar]
- Lu, H. Systematics of Eunicida (Polychaeta: Annelida) Homology, Ontogeny, Morphometrics, Fossils and Phylogeny. Ph.D. Thesis, George Washington University, Washington, DC, USA, 2006. [Google Scholar]
- Van Beneden, P.J. Histoire naturelle d’un animal nouveau designe sous le nom d’Histriobdella. Bull. l’Académie R. Belg. Brux. 1858, 2, 270–303. [Google Scholar]
- Foettinger, A. Recherches sur l’organisation de Histriobdella homari, P.-J. Van Beneden rapportée aux Archiannélides. Arch. Biol. (Liege) 1884, 5, 435–516. [Google Scholar]
- Shearer, C. On the anatomy of Histriobdella homari. Q. J. Microsc. Sci. 1910, 55, 17–20. [Google Scholar]
- Cordero, E.H. Un nuevo Arquianélido, Stratiodrilus platensis, n. sp., que habita sobre Aegla laevis (Latr.). Nota preliminar. Physis 1927, 8, 574–578. [Google Scholar]
- Roubaud, G. Recherches sur les Stratiodrilus platensis Cordero, Archiannelides parasites des Aegla des lacs de Patagonie. In Biologie de l’Amerique Australe II. Études sur les Faune du sol; Deboutteville, C.D., Rapport, E., Eds.; Éditions du Centre National de la Recherche Scientifique: Paris, France, 1962; pp. 31–54. [Google Scholar]
- Mesnil, F.; Caullery, M. L’appareil maxillaire d’Histriobdella homari; affinités des Hitriobdellides avec les Euniciens. Comptes Rendus l’Académie des Sci. 1922, 174, 913–917. [Google Scholar]
- Tzetlin, A.B. Ophryotrocha schubravyi sp.n. and the problem of evolution of the mouthparts in the Eunicemorpha (Polychaeta). Zool. Zhurnal 1980, 59, 666–676. (In Russian) [Google Scholar]
- Tzetlin, A.; Budaeva, N.; Vortsepneva, E.; Helm, C. New insights into the morphology and evolution of the ventral pharynx and jaws in Histriobdellidae (Eunicida, Annelida). Zool. Lett. 2020, 6, 1–19. [Google Scholar] [CrossRef]
- Gelder, S.R.; Jennings, J.B. The nervous system of the aberrant symbiotic polychaete Histriobdella homari and its implications for the taxonomic position of the Histriobdellidae. Zool. Anz. 1975, 194, 293–304. [Google Scholar]
- Jennings, J.B.; Gelder, S.R. Observations on the Feeding Mechanism, Diet and Digestive Physiology of Histriobdella homari Van Beneden 1858: An Aberrant Polychaete Symbiotic with North American and European Lobsters. Biol. Bull. 1976, 151, 489–517. [Google Scholar] [CrossRef]
- Fauchald, K. The polychaete worms. Definitions and keys to the orders, families and genera. Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1977, 28, 1–188. [Google Scholar]
- Harrison, L. On the genus Stratiodrilus (Archiannelida: Histriobdella) with a description of a new species from Madagascar. Rec. Aust. Mus. 1928, 16, 116–121. [Google Scholar] [CrossRef]
- Dioni, W. Didymorchis, Temnocephala (Platyhelminthes) y Stratiodrilus (Annelida) vermes epizoicos sobre Aegla y Parastacus (Crustacea, Decapoda) de lagos Andino-Patagonicos: Notas taxonomicas y biogeograficas. Acta Zool. Lilloana 1972, 29, 167–179. [Google Scholar]
- Rouse, G.W.; Pleijel, F. Polychaetes; Oxford University Press: New York, NY, USA, 2001; p. 354. [Google Scholar]
- Glasby, C.J.; Timm, T. Global diversity of polychaetes (Polychaeta; Annelida) in freshwater. Hydrobiologia 2008, 595, 107–115. [Google Scholar] [CrossRef]
- Rosa, J.J.D.S.; Marçal, I.C.; Teixeira, G.M.; Aguiar, A. Checklist of species of Stratiodrilus Haswell, 1900 (Annelida: Histriobdellidae), and new host records from Southern Brazil. Zootaxa 2018, 4399, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Steiner, T.M.; Amaral, A.C.Z. The family Histriobdellidae (Annelida, Polychaet) including descriptions of two new species from Brazil and a new genus. Contrib. Zool. 1999, 68, 1–17. [Google Scholar] [CrossRef]
- Zhang, Y.T. New substitute name for the genus Dayus Steiner & Amaral, 1999 (Annelida: Polychaeta: Histriobdellidae). Zootaxa 2014, 3802, 400. [Google Scholar]
- Amato, J.F.R.; Daudt, L.C.C.; Amato, S.B. New species of Stratiodrilus (Polychaeta, Histriobdellidae) from freshwater anomuran crustaceans of Southern Brazil. Biociências 2004, 12, 121–127. [Google Scholar]
- Lang, K. A contribution to the morphology of Stratiodrilus platensis Cordero (Histriobdellidae). Ark. Zool. 1949, 42, 1–30. [Google Scholar]
- Rouse, G.W. Histriobdellidae Vaillant, 1890. In Polychaetes; Rouse, G., Pleijel, F., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 160–163. [Google Scholar]
- Gelder, S.R.; Tyler, S. Anatomical and cytochemical studies on the adhesive organs of the ectosymbiont Histriobdella homari (Annelida: Polychaeta). Trans. Am. Microsc. Soc. 1986, 105, 348–356. [Google Scholar] [CrossRef]
- Haswell, W.A. On a new histriobdellid. Q. J. Microsc. Sci. 1900, 43, 299–335. [Google Scholar]
- Haswell, W.A. Notes on the Histriobdellidae. Q. J. Microsc. Sci. 1913, 59, 197–226. [Google Scholar]
- Daudt, L.C.C.; Amato, J.F.R. Morphological variation of Stratiodrius circensis (Polychaeta, Histriobdellidae) from a new host, Aegla leptodactyla (Crustacea, Anomura, Aeglidae) with identification of its type host species. Zootaxa 2007, 1450, 57–62. [Google Scholar] [CrossRef]
- Jamieson, B.G.M.; Afzelius, B.A.; Franzén, Å. Ultrastructure of the acentriolar, aflagellate spermatozoa and the eggs of Histriobdella homari and Stratiodrilus novaehollandiae (Histriobdellidae, Polychaeta). J. Submicrosc. Cytol. 1985, 17, 363–372. [Google Scholar]
- Simon, J.L. Incidence and behavior of Histriobdella homari (Annelida: Polychaeta) a commensal of the American lobster. Bioscience 1968, 18, 35–36. [Google Scholar]
- Boghen, A.D. A parasitological survey of the American lobster Homarus americanus from the Northumberland Strait, southern Gulf of St. Lawrence. Can. J. Zool. 1978, 56, 2460–2462. [Google Scholar] [CrossRef]
- Rouse, G.W. Polychaeta (Bristle Worms)—Chapter 5: Minor groups and fossils. In Marine Parasitology; Rohde, K., Ed.; CSIRO Publishing: Victoria, Australia, 2005; pp. 193–196. [Google Scholar]
- Martin, D.; Britayev, T.A. Symbiotic polychaetes: Review of known species. Oceanogr. Mar. Biol. Annu. Rev. 1998, 36, 217–340. [Google Scholar]
- Brattey, J.; Campbell, A. Occurrence of Histriobdella homari (Annelida: Polychaeta) on the American lobster in the Canadian Maritimes. Can. J. Zool. 1985, 63, 392–395. [Google Scholar] [CrossRef]
- Lerch, F.; Uglem, I. High density of Histriobdella homari van Beneden, 1858 (Annelida, Polychaeta) on ovigerous female european lobsters (Decapoda, Nephropidae). Crustaceana 1996, 69, 916–921. [Google Scholar] [CrossRef]
- Führ, I.M. A new histriobdellid on a marine isopod from South Africa. S. Afr. J. Sci. 1971, 67, 325–326. [Google Scholar]
- Bond-Buckup, G.; Buckup, L. A Família Aeglidae (Crustacea, Decapoda, Anomura). Arq. Zool. 1994, 32, 159–346. [Google Scholar]
- Briggs, R.P.; Atkinson, R.J.A.; McAliskey, M.; Rogerson, A. Histriobdella homari on Nephrops norvegicus from the Irish Sea and Clyde Sea area. J. Mar. Biol. Assoc. UK 1997, 77, 557–559. [Google Scholar] [CrossRef]
- Haswell, W.A. On the embryology of Stratiodrilus (Histriobdellidae). Q. J. Microsc. Sci. 1916, 61, 301–311. [Google Scholar]
- Amato, J.F.R.; Amato, S.B.; Daudt, L.C.C. The eggs of Stratiodrilus sp. (Polychaeta, Histriobdellidae). Biociências 2002, 10, 49–54. [Google Scholar]
- Vila, I.; Véliz, D. Life cycle of a freshwater Gondwanic remnant polychaete Stratiodrilus aeglaphilus (Annelida: Eunicida: Histriobdellidae), commensal with Aegla laevis (Crustacea: Anomura). Guyana 2014, 78, 120–126. [Google Scholar] [CrossRef]
- Cannon, L.R.G.; Jennings, J.B. Occurrence and nutritional relationships of four ectosymbiotes of the freshwater crayfishes Cherax dispar Riek and Cherax punctatus Clark (Crustacea: Decapoda) in Queensland. Aust. J. Mar. Freshw. Res. 1987, 38, 419–427. [Google Scholar] [CrossRef]
- Pardo, R.; Vila, I.; Bustamante, R. Demographic response of Stratiodrilus aeglaphilus (Annelida, Histriobdellidae) to organic enrichment: Experimental assessement. Hydrobiologia 2008, 598, 11–16. [Google Scholar] [CrossRef]
- Uzmann, J.R. Histriobdella homari (Annelida: Polychaeta) in the American lobster, Homarus americanus. J. Parasitol. 1967, 53, 210–211. [Google Scholar] [CrossRef]
- Moyano, H.I.G.; Carrasco, F.D.; Gacitua, S. Sobre las especies chilenas de Stratiodrilus Haswell, 1900, (Polychaeta, Histriobdellidae). Bol. Soc. Biol. Concepc. 1993, 64, 147–157. [Google Scholar]
- Carrera-Parra, L.F. Lumbrineridae Schmarda, 1861. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 263–275. [Google Scholar]
- Carrera-Parra, L.F. Recognition of Cenogenus Chamberlin, 1919 (Polychaeta: Lumbrineridae) based on type material. Proc. Biol. Soc. Wash. 2001, 114, 720–724. [Google Scholar]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. IV. Lumbrineridae. Physis 1973, 32, 343–393. [Google Scholar]
- Frame, A.B. The lumbrinerids (Annelida: Polychaeta) collected in two northwestern Atlantic surveys with descriptions of a new genus and two new species. Proc. Biol. Soc. Wash. 1992, 105, 185–218. [Google Scholar]
- Carrera-Parra, L.F. Phylogenetic analysis of Lumbrineridae Schmarda, 1861 (Annelida: Polychaeta). Zootaxa 2006, 1332, 1–36. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Phylogenetic relationship of Lumbricalus Frame, 1992 (Polychaeta: Lumbrineridae), with ecological and distribution data. Mar. Ecol. 2005, 26, 166–170. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F.; Orensanz, J.M. Revision of Kuwaita Mohammad, 1973 (Annelida, Polychaeta, Lumbrineridae). Zoosystema 2002, 24, 273–281. [Google Scholar]
- Carrera-Parra, L.F. Revision of Lumbricalus (Polychaeta: Lumbrineridae). J. Mar. Biol. Assoc. UK 2004, 84, 81–91. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Revision of Lumbrineris de Blainville, 1828 (Polychaeta: Lumbrineridae). Zootaxa 2006, 1336, 1–64. [Google Scholar] [CrossRef]
- Hilbig, B. Family Lumbrineridae Malmgren, 1867, emended Orensanz, 1990. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel. 5—The Annelida Part 2; Blake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1995; pp. 279–313. [Google Scholar]
- Pleijel, F. Lumbrineridae Schmarda, 1861. In Polychaetes; Rouse, G., Pleijel, F., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 164–165. [Google Scholar]
- Pruvot, G. Recherches anatomiques et morphologiques sur le système nerveux des Annélides polychètes. Arch. Zool. Expérimentale Générale 1885, 3, 211–326. [Google Scholar]
- Fauchald, K.; Jumars, P. The diet of worms: A study of polychaete feeding guilds. Oceanogr. Mar. Biol. Annu. Rev. 1979, 17, 193–284. [Google Scholar] [CrossRef]
- Valderhaug, V.A. Population structure and production of Lumbrineris fragilis (Polychaeta, Lumbrineridae) in the Oslofjord (Norway) with a note on metal content of jaws. Mar. Biol. 1985, 86, 203–211. [Google Scholar] [CrossRef]
- Carrasco, F.D.; Oyarzún, C. Diet of the polychaete Lumbrineris tetraura (Schmarda) (Lumbrineridae) in a polluted soft-bottom environmen. Bull. Mar. Sci. 1988, 42, 358–365. [Google Scholar]
- Petch, D.A. Selective Deposit-Feeding by Lumbrineris cf. latreilli (Polychaeta, Lumbrineridae), with a New Method for Assessing Selectivity by Deposit-Feeding Organisms. Mar. Biol. 1986, 93, 443–448. [Google Scholar]
- Gravier, C.; Dantan, J.L. Péchés nocturnes à la lumière dans la Baie d’Alger. Annélides polychètes. Ann. l’Institut Océanographique 1928, 5, 1–187. [Google Scholar]
- Rouse, G.W. An ultrastructural study of the spermatozoa of Eulalia sp.(Phyllodocidae), Lepidonotus sp. (Polynoidae), Lumbrineris sp.(Lumbrineridae) and Owenia fusiformis (Oweniidae). Helgoländer Wiss. Meeresunters. 1988, 42, 67–78. [Google Scholar] [CrossRef]
- Messina, P.; Di Filippo, M.; Gambi, M.C.; Zupo, V. In vitro fertilisation and larval development of a population of Lumbrineris (Scoletoma) impatiens (Claparede) (Polychaeta, Lumbrineridae) of the Gulf of Naples (Italy) in relation to aquaculture. Invertebr. Reprod. Dev. 2005, 48, 31–40. [Google Scholar] [CrossRef]
- Conti, G.; Loy, F.; Diana, M.; Messina, P. The spermatozoon of Lumbrineris (Scoletoma) impatiens (Claparède, 1868) (Polychaeta, Lumbrineridae). Ital. J. Zool. 2011, 78, 215–218. [Google Scholar] [CrossRef]
- Rouse, G.W.; Jamieson, B.G.M. An ultrastructural study of the sprematozoa of the polychaetes Eurythoe complanata (Amphinomidae), Clymenella sp. and Micromaldane sp. (Maldanidae), with definition of sperm types in relation to reproductive biology. J. Submicrosc. Cytol. 1987, 19, 573–584. [Google Scholar]
- Martins, R.; Carrera-Parra, L.F.; Quintino, V.; Rodrigues, A.M. Lumbrineridae (Polychaeta) from the Portuguese continental shelf (NE Atlantic) with the description of four new species. Zootaxa 2012, 3416, 1–21. [Google Scholar] [CrossRef]
- Hartman, O. New species of polychaetous annelids from southern California. With a discussion on the host of one of them. Allan Hancock Pac. Exped. 1939, 7, 157–172. [Google Scholar]
- Nishihira, M.; Tsuchiya, M.; and Sato, M. Ecological aspect of the breeding of the polychaete, Lumbrineris latreilli (Audouin et Milne-Edwards) at Asamushi, northern Japan. Bull. Mar. Biol. Stn. Asamushi 1980, 16, 201–212. [Google Scholar]
- Nishihira, M.; Tsuchiya, M.; Sato, M. Distribution and spawning site selection of the polychaete Lumbrineris latreilli (Audouin et Milne-Edwards) at Asamushi, northern Japan. Bull. Mar. Biol. Stn. Asamushi 1981, 17, 17–28. [Google Scholar]
- Nishihira, M.; Tsuchiya, M.; Sato, M. Dispersal and recruitment of juveniles of the polychaete Lumbrineris latreilli (Audouin et Milne-Edwards). Bull. Mar. Biol. Stn. Asamushi 1984, 17, 191–203. [Google Scholar]
- Carrera-Parra, L.F. Lumbrineridae (Annelida: Polychaeta) from the Grand Caribbean region with the description of six new species. J. Mar. Biol. Assoc. UK 2001, 81, 599–621. [Google Scholar] [CrossRef]
- Ayyagari, A.; Kondamudi, R. Ecological significance of the association between Stomopneustes variolaris (Echinoidea) and Lumbrineris latreilli (Polychaeta) from Visakhapatnam Coast, India. J. Mar. Biol. 2014, 2014, 640784. [Google Scholar] [CrossRef]
- Zibrowius, H.; Southward, E.C.; Day, J.H. New observations on a little known species of Lumbrineris (Polychaeta) living on various cnidarians, with notes on its recent and fossil scleractinian hosts. J. Mar. Biol. Assoc. UK 1975, 55, 83–108. [Google Scholar] [CrossRef]
- García-Gómez, S.C.; Carrera-Parra, L.F.; Mas, F.A.; Freitas, R.; Martins, R. Novel insights on the diversity and ecology of the family Lumbrineridae (Polychaeta) along the Iberian Peninsula coasts. J. Mar. Biol. Assoc. UK 2016, 96, 1427–1435. [Google Scholar] [CrossRef]
- Katsiaras, N.; Rousou, M.; Carrera-Parra, L.F.; García-Gómez, S.C.; Simboua, N.; Louizidou, P.; Chintiroglou, C.C.; Martins, R. Taxonomy, ecology and geographic distribution of Gallardoneris iberica (Polychaeta, Lumbrineridae) in southern Europe. J. Mar. Biol. Assoc. UK 2018, 98, 1609–1618. [Google Scholar] [CrossRef]
- Arias, A.; Carrera-Parra, L.F. First record of the genus Kuwaita (Annelida: Lumbrineridae) in Europe with the description of a new species and new ultramorphological data for the genus. Zootaxa 2014, 3887, 68–78. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Levenstein, R.Y. A new genus and species of Polychaeta (Family Lumbrineridae) from the deep-water trenches the the North Pacific. In Essays on Polychaetous Annelids in Memory of Dr. Olga Hartman; Reish, D.J., Fauchald, K., Eds.; The Allan Hancock Foundation, University of Southern California: Los Angeles, CA, USA, 1977; pp. 189–198. [Google Scholar]
- Oug, E. A new small species of Lumbrineris from northern Norway and arctic waters, with comments on L. minuta (Theel, 1879) and L. vanhoeffeni (Michaelsen, 1898)—(Polychaeta: Lumbrineridae). Ophelia 1998, 49, 147–162. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Zur Kenntnis des Eulitorals der australischen Küsten unter besonderer Berücksichtigung der Polychaeten und Ostracoden, 13. Die Polychaeten der antiborealen Küste von Victoria (Australien) (Zwischen Warrnambool im Wasten und Port Welshpool im Osten). Mitt. Hambg. Zool. Mus. Inst. 1987, 84, 27–66. [Google Scholar]
- Camargo, M.G.; da Lana, P.C. Lumbrineridae (Polychaeta: Eunicemorpha) da costa sul e sudeste do Brasil. I. Iheringia Série Zool. 1995, 79, 77–91. [Google Scholar]
- Camargo, M.G.; Lana, P. da C. Lumbrineridae (Polychaeta: Eunicemorpha) da costa sul e sudeste do Brasil. II. Lumbrineris. Iheringia 1995, 79, 93–120. [Google Scholar]
- Núñez, J.; Talavera, J.A.; Ocaña, O. Anélidos Poliquetos de Canarias: Familia Lumbrineridae. Vieraea 1991, 20, 89–100. [Google Scholar]
- Paxton, H.; Chou, L.M. Polychaetous annelids from the South China Sea. Raffles Bull. Zool. 2000, 48, 209–232. [Google Scholar]
- Cai, W.; Li, X. Eranno carrerai sp. nov. a new polychaete worm species (Polychaeta: Lumbrineridae) from coastal waters of China. Chin. J. Oceanol. Limnol. 2011, 29, 217–231. [Google Scholar] [CrossRef]
- Cai, W.; Li, X. A new species and new recorded species of Lumbrineridae Schmarda, 1861 (Anellida: Polychaeta) from China. Chin. J. Oceanol. Limnol. 2011, 29, 356–365. [Google Scholar] [CrossRef]
- Dean, K.D. Marine biodiversity of Costa Rica: Class Polychaeta (Annelida). Rev. Biol. Trop. 2004, 52, 131–181. [Google Scholar] [PubMed]
- Crossland, C. Polychaeta of tropical East Africa, the Red Sea, and Cape Verde Islands collected by Cyril Crossland, and of the Maldive Archipelago collected by Professor Stanley Gardiner, M.A., F.R.S. The Lumbriconereidae and Staurocephalidae. Proc. Zool. Soc. Lond. 1924, 1, 1–106. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Zur Ökologie der Polychaeten des Mangrove-Estero-Gebietes von El Salvador. Beiträge zur Neotropischen Fauna 1959, 1, 69–183. [Google Scholar] [CrossRef]
- Perkins, T.H. Lumbrineridae, Arabellidae, and Dorvilleidae (Polychaeta) from Florida, with descriptions of six new species. Proc. Biol. Soc. Wash. 1979, 92, 415–465. [Google Scholar]
- Uebelacker, J.M. Family Lumbrineridae Malmgren, 1867. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; p. 41. [Google Scholar]
- Granados-Barba, A.; Solís-Weiss, V. New records of polychaetous annelids (Order: Eunicida) from the southeastern Gulf of Mexico. Bull. Mar. Sci. 1994, 52, 420–427. [Google Scholar]
- Solís-Weiss, V.; Granados-Barba, A.; Rodríguez-Villanueva, L.V.; Miranda-Vázquez, L.A.; Ochoa-Rivera, V.; Hernández-Alcántara, P. The Lumbrineridae of the continental shelf in mexican portion of the Gulf of Mexico. Mitt. Hambg. Zool. Mus. Inst. 1995, 92, 61–75. [Google Scholar]
- Fauvel, P. Annelida Polychaeta of the Indian Museum, Calcutta. Mem. Indian Mus. 1932, 12, 1–262. [Google Scholar]
- Imajima, M.; Higuchi, M. Lumbrineridae of polychaetous annelids from Japan, with descriptions of six new species. Bull. Natl. Sci. Mus. 1975, 1, 5–37. [Google Scholar]
- Imajima, M. Six species of Lumbrinerides (Polychaeta, Lumbrineridae) from Japan. Bull. Natl. Sci. Mus. 1985, 11, 171–184. [Google Scholar]
- Miura, T. Classification and morphological variations of the Japanese species of Lumbrinerides (Annelida: Lumbrineridae). Species Divers. 2017, 22, 7–27. [Google Scholar] [CrossRef][Green Version]
- Ramos, J.M. Lumbrineridae (Polychètes errantes) de Méditterranée. Ann. l’Institut Océanograph. 1976, 52, 103–137. [Google Scholar]
- Miura, T. Lumbrineridae (Annélides Polychètes) abyssaux récoltés au cours de campagnes du Centre Océanologique de Bretagne dans l’Atlantique et la Mediterranée. Bull. Muséum d’Histoire Nat. 1980, 2, 1019–1057. [Google Scholar]
- Carrera-Parra, L.F.; Çinar, M.E.; Dagli, E. Description of a new species of Lumbrineris (Polychaeta: Lumbrineridae) from the coasts of Turkey (eastern Mediterranean). Mar. Biodivers. 2011, 41, 343–347. [Google Scholar] [CrossRef]
- D’Alessandro, M.; Cosentino, A.; Giacobbe, S.; Andaloro, F.; Romeo, T. Description of a new species of Abyssoninoe (Polychaeta: Lumbrineridae) from the NE Sicily (central Mediterranean Sea). J. Mar. Biol. Assoc. UK 2014, 94, 747–752. [Google Scholar] [CrossRef]
- Knox, G.A.; Green, K.M. The Polychaetous Annelids of New Zealand Part 2: Lumbrineridae. J. R. Soc. N. Z. 1972, 2, 69–81. [Google Scholar]
- Oug, E. Lumbrineridae from the Andaman Sea, Thailand, with notes on Oenonidae and Dorvilleidae (Annelida: Polychaeta). Phuket Mar. Biol. Cent. Spec. Publ. 2002, 24, 117–138. [Google Scholar]
- Parapar, J.; Oconnor, B.; Besteiro, C.; Urgorri, V. Abyssoninoe hibernica (McIntosh) (Polychaeta: Lumbrineridae) a valid species from the Northeast Atlantic. Sarsia 1994, 79, 157–162. [Google Scholar] [CrossRef]
- Aguirrezabalaga, F.; Carrera-Parra, L.F. Lumbrineridae (Polychaeta) from the Capbreton Canyon (Bay of Biscay, NE Atlantic) with the description of two new species. Sci. Mar. 2006, 70, 17–25. [Google Scholar] [CrossRef]
- Hernández-Alcántara, P.; Pérez-Mendoza, A.Y.; Solís-Weiss, V. Description of three new species of Ninoe and Cenogenus (Polychaeta: Lumbrineridae) from the Mexican Pacific. Sci. Mar. 2006, 70, 81–90. [Google Scholar]
- Fauchald, K. Deep-water errant polychaetes from Hardangerfjorden, western Norway. Sarsia 1974, 57, 1–32. [Google Scholar] [CrossRef]
- Winsnes, I.M. A new species of Lumbrineris (Polychaeta) and a new subspecies of Lumbrineris scopa Fauchald from the coast of Norway. Zool. Scr. 1981, 10, 91–94. [Google Scholar] [CrossRef]
- Winsnes, I.M. Augeneria algida (Wiren) comb. n., a deep-sea lumbrinerid from Spitzbergen with aberrant setae (Annelida, Polychaeta): Redescription of holotype. Zool. Scr. 1987, 16, 39–45. [Google Scholar] [CrossRef]
- Oug, E. Guide to identification of Lumbrineridae (Polychaeta) in Norwegian and adjacent waters. Nor. Polych. Forum Guid. 2011, 1–22. [Google Scholar]
- Hartmann-Schröder, G. Zur polychaeten-fauna von Peru. Beiträge zur Neotropischen Fauna 1960, 2, 1–44. [Google Scholar] [CrossRef]
- Hartmann-Schröder, G. Die Polychaeten des Eulitorals. Zur Kenntnis des Eulitorals der chilenischen Pazifikküste und der argentinischen Küste Südpatagoniens unter besonderer Berücksichtigung der Polychaeten und Ostracoden. Mitt. Hambg. Zool. Mus. Inst. 1962, 60, 57–167. [Google Scholar]
- Rozbaczylo, N.; Vásquez-Yáñez, P.; Moreno, R.A.; Díaz-Díaz, O. Poliquetos bentónicos Amphinomida, Phyllodocida y Eunicida (Annelida: Polychaeta) de la región de fiordos y canales australes de Chile recolectados durante los cruceros CIMAR 13 al 20 fiordos. An. Inst. Patagon. 2018, 45, 51–91. [Google Scholar] [CrossRef]
- Rozbaczylo, N.; Moreno, R.A.; Díaz-Díaz, O. Poliquetos bentónicos submareales de fondos blandos de la región de Aysén, Chile: Clados Amphinomida, Eunicida, Spionida, Sabellida y Scolecida (Annelida, Polychaeta). Investig. Mar. 2006, 34, 43–62. [Google Scholar] [CrossRef]
- Gallardo, V.A. Polychaeta from the bay of Nha Trang, South Viet Nam. NAGA Rep. 1968, 4, 35–279. [Google Scholar]
- Fauvel, P.; Rullier, F. Annélides Polychètes. Resultats Scientifiques des Campagnes de Calypso IV. Ann. l’Institut Océanograph. 1959, 37, 143–205. [Google Scholar]
- Amoureux, L. Quelques annélides polychètes de l’Afrique occidentale et équatoriale. Cah. ORSTOM sér Océnnogr 1973, 11, 41–65. [Google Scholar]
- Intes, A.; Le Loeuff, P. Les annélides polychètes de Côte d’Ivoire. 1. Polychètes Errantes—Compte rendu systématique. Cah. ORSTOM sér Océnnogr 1975, 13, 267–321. [Google Scholar]
- Brasier, M.J.; Wiklund, H.; Neal, L.; Jeffreys, R.; Linse, K.; Ruhl, H.; Glover, A.G. DNA barcoding uncovers cryptic diversity in 50% of deep-sea antarctic polychaetes. R. Soc. Open Sci. 2016, 3, 160432. [Google Scholar] [CrossRef] [PubMed]
- Font, T.; Gil, J.; Lloret, J. The commercialization and use of exotic baits in recreational fisheries in the north-western Mediterranean: Environmental and management implications. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 651–661. [Google Scholar] [CrossRef]
- Núñez, J.; Núñez, L.; Maggio, Y. Invertebrados que se Comercializan en Canarias Como cebo vivo para la Pesca Deportiva; Gobierno de Canarias: La Laguna, Spain, 2011. [Google Scholar]
- Usamentiaga, P.; Rodríguez, F.; Martin-Gil, D.; Saiz, N.; Morchon, E.; Alonso, S.; Jerez, J.; Bartolomé, B. Protein contact dermatitis by fishing bait (Lumbrinereis latreilli). Contact Dematitis 2005, 53, 236–237. [Google Scholar] [CrossRef]
- Ruggeri, G.D. Drugs from the Sea. Marine organisms with novel chemical constituents are excellent sources of new drugs. Science 1976, 194, 491–497. [Google Scholar]
- Montagu, G. Description of several marine animals found on the south coast of Devonshire. Trans. Linn. Soc. Lond. 1804, 7, 80–84. [Google Scholar] [CrossRef]
- Kinberg, J.G.H. Annulata nova. Öfversigt af Königlich Vetensk. Förhandlingar Stock 1865, 21, 559–574. [Google Scholar]
- Colbath, G.K. Revision of the family Lysaretidae, and recognition of the family Oenonidae Kinberg, 1865 (Eunicida: Polychaeta). Proc. Biol. Soc. Wash. 1989, 102, 116–123. [Google Scholar]
- Dean, H.K. A new arabellid polychaete living in the mantle cavity of deep-sea wood boring bivalves (family Pholadidae). Proc. Biol. Soc. Wash. 1992, 105, 224–232. [Google Scholar]
- Carrera-Parra, L.F. Oenonidae Kinberg, 1865. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 355–362. [Google Scholar]
- Zanol, J.; Ruta, C. New and previously known species of Oenonidae (Polychaeta: Annelida) from Lizard Island, Great Barrier Reef, Australia. Zootaxa 2015, 4019, 745–772. [Google Scholar] [CrossRef]
- Saito, H.; Imabayashi, H.; Kawai, K. Growth of the bivalve-feeder Halla okudai (Polychaete: Lysaretidae) under wild and rearing conditions, in relation to species and abundance of prey organisms. Fish. Sci. 1999, 65, 230–234. [Google Scholar] [CrossRef]
- Kawai, K.; Kunitake, H.; Saito, H.; Imabayashi, H. Paralytic and digestive activities of jelly-like substances secreted by a lysaretid polychaete, Halla okudai. Benthos Res. 1999, 54, 1–7. [Google Scholar] [CrossRef][Green Version]
- Iori, D.; Forti, L.; Massamba-N’Siala, G.; Prevedelli, D.; Simonini, R. Toxicity of the purple mucus of the polychaete Halla parthenopeia (Oenonidae) revealed by a battery of ecotoxicological bioassays. Sci. Mar. 2014, 78, 589–595. [Google Scholar]
- Bielig, H.J.; Möllinger, H. Über hallochrom, den epithelfarbstoff von Halla parthenopeia Costa. Physiol. Chem. 1960, 321, 276–289. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Oenonidae Kinberg, 1865. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.A., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Salazar-Vallejo, S.I., Solís-Weiss, V., Tovar-Hernández, M.A., Eds.; unpublished.
- Amaral, A.C. Um poliqueto endoparasita, Labrorostratus prolificus sp. nov. em nereídeo. Bol. Inst. Oceanogr. 1977, 26, 285–292. [Google Scholar] [CrossRef]
- Abd-Elnaby, F.A. Observations on Spermatogenesis in Polychaete. Species, Halla parthenopeia (Family: Oenonidae, Polychaeta). World Appl. Sci. J. 2009, 6, 144–153. [Google Scholar]
- Pettibone, M.H. Endoparasitic polychaetous annelids of the family Arabellidae with descriptions of new species. Biol. Bull. Biol. Lab. 1957, 113, 170–187. [Google Scholar] [CrossRef]
- Pleijel, F. Oenonidae Kinberg, 1865. In Polychaetes; Rouse, G., Pleijel, F., Eds.; Oxford University Press: New York, NY, USA, 2001; pp. 166–168. [Google Scholar]
- Hilbig, B. Family Oenonidae Kinberg, 1865, emended Orensanz, 1990. In Taxonomic Atlas of the Benthic Fauna of the Santa Maria Basin and Western Santa Barbara Channel. 5—The Annelida Part 2; Blake, J.A., Hilbig, B., Scott, P.H., Eds.; Santa Barbara Museum of Natural History: Santa Barbara, CA, USA, 1995; pp. 315–339. [Google Scholar]
- Hernández-Alcántara, P.; Sólis-Weiss, V. Parasitism among polychaetes: A rare case illustrated by a new species: Labrorostratus zaragozensis, n. sp. (Oenonidae) found in the Gulf of California, Mexico. J. Parasitol. 1998, 84, 978–982. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Alcántara, P.; Cruz-Pérez, I.N.; Sólis-Weiss, V. Labrorostratus caribensis, a new oenonid polychaete from the Grand Caribbean living in the body cavity of a nereidid, with emendation of the genus. Zootaxa 2015, 4048, 127–139. [Google Scholar] [CrossRef]
- Carr, C.M. Polychaete diversity and distribution patterns in Canadian marine waters. Mar. Biodivers. 2012, 42, 93–107. [Google Scholar] [CrossRef]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. VI. Arabellidae. Physis 1974, 33, 381–408. [Google Scholar]
- Hartmann-Schröder, G. Die Polychaeten der tropischen Nordwestküste Australiens (zwischen Derby im Norden und Port Hedland im Süden). Teil 2. Zur Kenntnis des Eulittorals der australischen Küsten unter besonderer Berücksichtigung der Polychaeten und Ostracoden. Mitteilungen aus dem Hamburgischen Zoologischen Museum und Institut 1979, 76, 75–218. [Google Scholar]
- Rizzo, A.E.; Almeida dos Santos, S.; da Conceição Guerreiro Couto, E. First report of Labrorostratus caribensis (Annelida, Oenonidae) as endoparasite of Haplosyllis rosenalessoae (Annelida, Syllidae) from Brazil. Int. J. Parasitol. Parasites Wildl. 2020, 12, 64–66. [Google Scholar] [CrossRef]
- Ibarzabal, D.R. Poliquetos de Punta del Este, Isla de la Juventud, Cuba. IV Orden Eunicida. Poeyana 1989, 384, 1–28. [Google Scholar]
- Díaz-Díaz, O.; Liñero-Arana, I. Poliquetos asociados a substratos artificiales sumergidos en la costa noroerental de Venezuela 3. Eunicida (Annelida: Polychaeta). Bol. del Inst. Oceanogr. Venez. 2002, 41, 3–14. [Google Scholar]
- Dean, K.D.; Sibaja-Cordero, J.A.; Cortés, J. Polychaetes (Annelida: Polychaeta) of Cocos Island National Park, Pacific Costa Rica. Pac. Sci. 2012, 66, 347–386. [Google Scholar] [CrossRef]
- Ravara, A.; Cunha, M.R. An unusual new species of Oenonidae (Polychaeta, Eunicida) from the North Atlantic Ocean. Mar. Biodivers. 2018, 48, 1037–1043. [Google Scholar] [CrossRef]
- Uebelacker, J.M. A new parasitic polychaetous annelid (Arabellidae) from the Bahamas. J. Parasitol. 1978, 64, 151–154. [Google Scholar] [CrossRef]
- Uebelacker, J.M. Family Arabellidae Hartman, 1944. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; pp. 42.1–42.29. [Google Scholar]
- Gilbert, K. Family Lysaretidae Kinberg, 1865. In Taxonomic Guide to the Polychaetes of the Northern Gulf of Mexico. Final Report to the Minerals Management Service, Contract 14-12-001-29091. Volume VI; Uebelacker, J.M., Johnson, P.G., Eds.; U.S. Department of the Interior Mineral Management Service Gulf of Mexico OCS.: Mobile, Italy, 1984; pp. 43.1–43.7. [Google Scholar]
- Imajima, M. Errant polychaetous annelids from Tsukumo Bay and vicinity of Noto Penisula, Japan. Bull. Natl. Sci. Mus. 1967, 10, 403–441. [Google Scholar]
- Imajima, M. Polychaetous annelids of Suruga Bay, central Japan. Natl. Sci. Mus. Monogr. 1997, 12, 149–228. [Google Scholar]
- Imajima, M. Deep-sea benthic polychaetous annelids of Tosa Bay, southwestern Japan. Natl. Sci. Mus. Monogr. 2001, 20, 31–100. [Google Scholar]
- Imajima, M. Polychaetous annelids from Sagami Bay and the Sagami Sea, central Japan. Mem. Natl. Sci. Mus. 2006, 40, 317–408. [Google Scholar]
- Kobayashi, G.; Mukai, R.; Itoh, H. New record of Halla okudai Imajima, 1967 (Annelida, Eunicida, Oenonidae) from Fukue Island in the Goto Islands, Japan. Check List 2020, 16, 1199–1203. [Google Scholar]
- Ramos, J.M. Arabellidae (Polychètes Errantes) de Méditerranée. Ann. l’Institut Océanograph. 1976, 52, 247–257. [Google Scholar]
- San Martín, G.; Sardá, R. Sobre la presencia de un Arabélido (Polychaeta: Arabellidae) parásito de Sílidos (Polychaeta: Syllidae), del género Labrorostratus en las costas espanolas. Boletín la Real Soc. Española Hist. Nat. 1986, 82, 141–146. [Google Scholar]
- Cantone, G. Censimento de policheti de mari Italiani: Arabellidae Hartman, 1944, Oenonidae Kinberg, 1865. Atti Soc. Toscana Sci. Nat. Mem. Ser. B 1996, 103, 99–103. [Google Scholar]
- Mikac, B. A sea of worms: Polychaete checklist of the Adriatic Sea. Zootaxa 2015, 3943, 1–172. [Google Scholar] [CrossRef][Green Version]
- Knox, G.A.; Green, K.M. The polychaetes of New Zealand. Part 3. Lysaretidae. J. R. Soc. N. Z. 1972, 2, 431–434. [Google Scholar] [CrossRef]
- Fauchald, K. Some Polychaetous Annelids From the Deep Basins in Sognefjorden, Western Norway. Sarsia 1972, 49, 89–106. [Google Scholar] [CrossRef]
- Hartman, O. Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic areas. Occas. Pap. Allan Hancock Found. 1965, 28, 1–378. [Google Scholar]
- Hartman, O.; Fauchald, K. Deep-water benthic polychaetous annelids off New England to Bermuda and other North Atlantic Areas. Part II. Allan Hancock Monogr. Mar. Biol. 1971, 6, 1–327. [Google Scholar]
- Kirkegaard, J.B. Bathyal and abyssal polychaetes (errant species). Galathea Repor 1995, 17, 7–56. [Google Scholar]
- Baeta, M.; Navarrete, A.; Ballesteros, M. First description of a new worm bait fishery in the NW Mediterranean Sea. Mediterr. Mar. Sci. 2019, 20, 302–308. [Google Scholar]
- Saito, H.; Kawai, K.; Umino, T.; Imabayashi, H. Fishing bait worm supplies in Japan in relation to their physiological traits. Mem. Mus. Vic. 2014, 71, 279–287. [Google Scholar] [CrossRef]
- Idris, I.; Arshad, A. Checklist of polychaetous annelids in Malaysia with redescription of two commercially exploited species. Asian J. Anim. Vet. Adv. 2013, 8, 409–436. [Google Scholar] [CrossRef]
- Pettibone, M.H. Polychaeta Errantia of the Siboga Expedition. Part IV. Some additional polychaetes of the Polynoidae, Hesionidae, Nereidae, Goniadidae, Eunicidae, and Onuphidae, selected as new species by the late Dr. Hermann Augener with remarks on other related species. Siboga-Exped. Monogr. 1970, 24, 199–270. [Google Scholar]
- Kucheruk, N.V. Deep-water Onuphidae (Polychaeta) from the collections of 16th cruise of R/V Dmitry Mendeleev (to the generic classification of the family Onuphidae). Tr. Institut. Okeanol. Akad. Nauk SSSR 1978, 113, 88–106. (In Russian) [Google Scholar]
- Fauchald, K. Revision of Onuphis, Nothria, and Paradiopatra (Polychaeta: Onuphidae) based upon type material. Smithson. Contrib. Zool. 1982, 356, 1–109. [Google Scholar] [CrossRef]
- Paxton, H. Generic Revision and Relationships of the Family Onuphidae (Annelida: Polychaeta). Rec. Aust. Mus. 1986, 38, 1–74. [Google Scholar] [CrossRef]
- Paxton, H. Fauchaldonuphis, a new genus for the enigmatic polychaete Diopatra paradoxa Quatrefages, 1866. Mar. Ecol. 2005, 26, 209–215. [Google Scholar] [CrossRef]
- Carrera-Parra, L.F. Onuphidae Kinberg, 1865. In Poliquetos (Annelida: Polychaeta) de México y América Tropical; de León-González, J.Á., Bastida-Zavala, J.R., Carrera-Parra, L.F., García-Garza, M.E., Peña-Rivera, A., Salazar-Vallejo, S.I., Solís-Weiss, V., Eds.; Universidad Autónoma de Nuevo León: Monterrey, Mexico, 2009; pp. 363–378. [Google Scholar]
- de León-González, J.A.; Cornejo-Rodríguez, M.H.; Degraer, S. A new species of Australonuphis (Polychaeta: Onuphidae) from the eastern Pacific. J. Mar. Biol. Assoc. UK 2008, 88, 739–742. [Google Scholar] [CrossRef]
- Paxton, H. Hirsutonuphis (Polychaeta: Onuphidae) from Australia, with a discussion of setal progression in juveniles. Invertebr. Taxon. 1996, 10, 77–96. [Google Scholar] [CrossRef]
- Estrella-Ruiz, A.; Hernández-Alcántara, P.; Solís-Weiss, V. Description of a new species of Hirsutonuphis (Annelida: Onuphidae) from the Mexican Pacific, with notes on the world distribution of the genus. J. Mar. Biol. Assoc. UK 2013, 93, 1777–1783. [Google Scholar] [CrossRef]
- Paxton, H. Revision of the Rhamphobrachium complex (Polychaeta: Onuphidae). Rec. Aust. Mus. 1986, 38, 75–104. [Google Scholar] [CrossRef][Green Version]
- Paxton, H.; Budaeva, N. Minibrachium, a new subgenus of Rhamphobrachium (Annelida: Onuphidae) from Australia with the description of three new species. Zootaxa 2015, 4019, 621–634. [Google Scholar] [CrossRef]
- Paxton, H.; Budaeva, N. Paradiopatra (Annelida: Onuphidae) from eastern Australian waters, with the description of six new species. Zootaxa 2013, 3686, 140–164. [Google Scholar]
- Meyer, K.S.; Wagner, J.K.S.; Ball, B.; Turner, P.J.; Young, C.M.; Van Dover, C.L. Hyalinoecia artifex: Field notes on a charismatic and abundant epifaunal polychaete on the US Atlantic continental margin. Invertebr. Biol. 2016, 135, 211–224. [Google Scholar] [CrossRef]
- Nishi, E.; Kato, T. Longibrachium arariensis, a new species of Onuphidae (Annelida: Polychaeta) from the shallow water of Izu Peninsula, central Japan, with notes on its feeding behavior. Zootaxa 2009, 2081, 46–56. [Google Scholar] [CrossRef]
- Gaston, G.R.G. Benthic Polychaeta of the Middle Atlantic Bight: Feeding and distribution. Mar. Ecol. Prog. Ser. 1987, 36, 251–262. [Google Scholar] [CrossRef]
- Dayton, P.K.; Hessler, R.R. Role of biological disturbance in maintaining diversity in the deep sea. Deep Sea Res. 1972, 19, 199–208. [Google Scholar] [CrossRef]
- Lipps, J.H.; Ronan, T.E. Predation on Foraminifera by the polychete worm Diopatra. J. Foraminifer. Res. 1974, 4, 139–143. [Google Scholar] [CrossRef]
- Mangum, C.P.; Santos, S.L.; Rhodes, W.R., Jr. Distribution and feeding in the onuphid polychaete, Diopatra cuprea (Bosc). Mar. Biol. 1968, 2, 33–40. [Google Scholar] [CrossRef]
- Brenchley, G.A.; Tidball, J.G. Tube-cap orientations of Diopatra cuprea (Bosc) (Polychaeta): The compromise between physiology and foraging. Mar. Behav. Physiol. 1980, 7, 1–13. [Google Scholar] [CrossRef]
- Arias, A.; Richter, A.; Anadón, N.; Paxton, H. Evidence of simultaneous hermaphroditism in the brooding Diopatra marocensis (Annelida: Onuphidae) from northern Spain. J. Mar. Biol. Assoc. UK 2013, 93, 1533–1542. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. The cryptogenic bait worm Diopatra biscayensis Fauchald et al., 2012 (Annelida: Onuphidae)—Revisiting its history, biology and ecology. Estuar. Coast. Shelf Sci. 2015, 163, 22–36. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H.; Budaeva, N. Redescription and biology of Diopatra neapolitana (Annelida: Onuphidae), a protandric hermaphrodite with external spermaducal papillae. Estuar. Coast. Shelf Sci. 2016, 174, 1–17. [Google Scholar] [CrossRef]
- Hsieh, H.L.; Simon, J.L. The sperm transfer system in Kinbergonuphis simoni (Polychaeta: Onuphidae). Biol. Bull. 1990, 178, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Blake, J.A. The larval development of Polychaeta from the nothern California coast. II. Nothria elegans (Family Onuphidae). Ophelia 1975, 13, 43–61. [Google Scholar] [CrossRef]
- Paxton, H. Diopatra Audouin and Milne Edwards (Polychaeta, Onuphidae) from Australia, with a discussion of developmental patterns in the genus. Beagle Rec. North Territ. Mus. Arts Sci. 1993, 10, 115–154. [Google Scholar]
- Carrasco, F.D. Description of adults and larvae of a new deep water species of Hyalinoecia (Polychaeta, Onuphidae) from the southeastern Pacific Ocean. J. Nat. Hist. 1983, 17, 87–93. [Google Scholar] [CrossRef]
- Hsieh, H.L.; Simon, J.L. Larval development of Kinbergonuphis simoni, with a summary of development patterns in the family Onuphidae (Polychaeta). Bull. Biol. Soc. Wash. 1987, 7, 194–210. [Google Scholar]
- Fadlaoui, S.A.; Lechapt, J.P.; Retière, C. Larval development of the onuphid Diopatra marocensis (Annelida: Polychaeta) from the Atlantic coast of Morocco. J. Mar. Biol. Assoc. UK 1995, 75, 957–966. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Diopatra tuberculantennata, a new species of Onuphidae (Polychaeta) from Belize with a key to onuphids from the Caribbean Sea. Zootaxa 2008, 1795, 29–45. [Google Scholar] [CrossRef]
- Budaeva, N.; Fauchald, K. Larval development of Mooreonuphis stigmatis (Treadwell 1922) (Polychaeta: Onuphidae) from the north-east Pacific. Mar. Biol. Res. 2010, 6, 6–24. [Google Scholar] [CrossRef]
- Budaeva, N.; Pyataeva, S.; Meißner, K. Development of the deep-sea viviparous quill worm Leptoecia vivipara (Hyalinoeciinae, Onuphidae, Annelida). Invertebr. Biol. 2014, 133, 242–260. [Google Scholar] [CrossRef]
- Rangel, L.; Santos, M. Diopatra neapolitana (Polychaeta: Onuphidae) as a second intermediate host of Gymnophallus choledochus (Digenea: Gymnophallidae) in the Aveiro Estuary (Portugal): Distribution within the host and histopathology. J. Parasitol. 2009, 95, 1233–1236. [Google Scholar] [CrossRef]
- Rangel, L.F.; Cech, G.; Székely, C.; Santos, M.J. A new actinospore type unicapsulactinomyxon (Myxozoa), infecting the marine polychaete, Diopatra neapolitana (Polychaeta: Onuphidae) in the Aveiro Estuary (Portugal). Parasitology 2011, 138, 698–712. [Google Scholar] [CrossRef] [PubMed]
- Arias, A.; Anadón, N.; Paxton, H. New records of Diopatra marocensis (Annelida: Onuphidae) from northern Spain. Zootaxa 2010, 2691, 67–68. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. Hidden diversity within the polychaete Onuphis eremita sensu lato (Annelida: Onuphidae)—Redescription of O. eremita Audouin & Milne-Edwards, 1833 and reinstatement of Onuphis pancerii Claparède, 1868. Zootaxa 2014, 3861, 145–169. [Google Scholar] [PubMed]
- Jones, C.; Lawton, J.; Shachak, M. Organisms as ecosystem engineers. Oikos 1994, 69, 373–386. [Google Scholar] [CrossRef]
- Berke, S.K.; Mahon, A.R.; Lima, F.P.; Halanych, K.M.; Wethey, D.S.; Woodin, S.A. Range shifts and species diversity in marine ecosystem engineers: Patterns and predictions for European. Glob. Ecol. Biogeogr. 2010, 19, 223–232. [Google Scholar] [CrossRef]
- Berke, S.K. Biogeographic variability in ecosystem engineering: Patterns in the abundance and behavior of the tube-building polychaete Diopatra cuprea. Mar. Ecol. Prog. Ser. 2012, 447, 1–13. [Google Scholar] [CrossRef]
- Bailey-Brock, J. Ecology of the tube-building polychaete Diopatra leuckarti Kinberg, 1865 (Onuphidae) in Hawaii: Community structure, and sediment stabilizing properties. Zool. J. Linn. Soc. Lond. 1984, 80, 191–199. [Google Scholar] [CrossRef]
- McIntosh, W.C. Report on the Annelida Polychaeta collected by H.M.S. Challenger during the years 1873–1876. Rep. Sci. Results Voyag. H.M.S.Chall. Dur. Years 1872–76 1885, 12, 1–554. [Google Scholar]
- Kucheruk, N.V. On the specific composition and distribution of the deep-sea genus Paraonuphis (sic) (Polychaeta, Eunicidae). Tr. Institut. Okeanol. Akad. Nauk SSSR 1977, 108, 44–51. (In Russian) [Google Scholar]
- Jirkov, I.A. Polychaeta of the Arctic Ocean; Yanus-K: Moscow, Russia, 2001. [Google Scholar]
- Hartman, O. Polychaetous annelids collected by the USNS Eltanin and Staten Island cruises, chiefly from Antarctic Seas. Allan Hancock Monogr. Mar. Biol. 1967, 2, 1–387. [Google Scholar]
- Fauchald, K. Onuphidae (Polychaeta) from Western Mexico. Allan Hancock Monogr. Mar. Biol. 1968, 3, 1–82. [Google Scholar]
- Fauchald, K. Polychaetes from Central American sandy beaches. Bull. South Calif. Acad. Sci. 1973, 72, 19–31. [Google Scholar]
- Fauchald, K. Onuphidae (Polychaeta) from Belize, Central America, with notes on related taxa. Biol. Soc. Wash. 1980, 93, 797–829. [Google Scholar]
- de León-González, J.A. de Soft bottom polychaetes from the western coast of Baja California Sur, México. 4. Onuphidae. Cah. Biol. Mar. 1994, 35, 57–67. [Google Scholar]
- Imajima, M. Two New Species of Nothria (Polychaeta, Onuphidae) from off Sanriku, Northeastern Honshu, Japan. Bull. Natl. Sci. Mus. 1989, 15, 1–10. [Google Scholar]
- Imajima, M. Onuphidae (Annelida, Polychaeta) from Japan, excluding the genus Onuphis. Natl. Sci. Mus. Monogr. 1999, 16, 1–115. [Google Scholar]
- Maekawa, N.; Hayashi, I. Onuphid polychaetes from Wakasa Bay, Sea of Japan. Mem. Coll. Agric. 1989, 134, 61–93. [Google Scholar]
- Maekawa, N.; Hayashi, I. Taxonomic study on the genus Onuphis (Polychaeta, Onuphidae) from Japan and adjacent seas, with descriptions of six new species. Bull. Natl. Sci. Mus. 1999, 25, 163–214. [Google Scholar]
- Paxton, H. Three new species of Aponuphis (Annelida: Onuphidae) from eastern Australia. Zootaxa 2017, 4344, 246–260. [Google Scholar] [CrossRef]
- Paxton, H. Two new species of Mooreonuphis (Annelida: Onuphidae) from Australia. Zootaxa 2018, 4486, 589–600. [Google Scholar] [CrossRef]
- Budaeva, N.; Paxton, H. Nothria and Anchinothria (Annelida: Onuphidae) from eastern Australian waters, with a discussion of ontogenetic variation of diagnostic characters. J. Mar. Biol. Assoc. UK 2013, 93, 1487–1502. [Google Scholar] [CrossRef]
- Augener, H. Polychaeta. Beiträge zur Kennt. der Meeresfauna Westafrikas 1918, 2, 67–625. [Google Scholar]
- Arias, A. Onuphis and Mooreonuphis (Annelida: Onuphidae) from West Africa with the description of three new species and the reinstatement of O. landanaensis Augener, 1918. Zootaxa 2016, 4168, 481–511. [Google Scholar] [CrossRef] [PubMed]
- Paxton, H. A new species of Diopatra (Annelida: Onuphidae) from Namibia, southwestern Africa. Mar. Biodivers. 2016, 46, 889–895. [Google Scholar] [CrossRef]
- Orensanz, J.M. Los anelidos poliquetos de la provincia biogeografica Argentina. V. Onuphidae. Physis 1974, 33, 75–122. [Google Scholar]
- Paxton, H. The Diopatra chiliensis confusion—Redescription of D. chiliensis (Polychaeta, Onuphidae) and implicated species. Zool. Scr. 1998, 27, 31–48. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H.; Anadón, N. Mooreonuphis vespa, a new brooding species of Onuphidae (Annelida) from northern Spain (Bay of Biscay). Zootaxa 2013, 3741, 583–592. [Google Scholar] [CrossRef]
- Arias, A.; Núñez, J.; Paxton, H. Onuphid polychaetes associated with the Cymodocea nodosa meadows of La Gomera (Canary Islands, NW Africa)—New species and new records from the eastern North Atlantic. J. Mar. Biol. Assoc. UK 2017, 97, 857–869. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. Onuphis and Aponuphis (Annelida: Onuphidae) from southwestern Europe, with the description of a new species. Zootaxa 2015, 3949, 345–369. [Google Scholar] [CrossRef]
- Arias, A.; Paxton, H. Paradiopatra Ehlers, 1887 (Annelida: Onuphidae) from southwestern Europe with the description of a new species and new ultramorphological data for the genus. Zootaxa 2015, 4040, 149–168. [Google Scholar] [CrossRef]
- Paxton, H.; Arias, A. Unveiling a surprising diversity of the genus Diopatra Audouin & Milne Edwards, 1833 (Annelida: Onuphidae) in the Macaronesian region (eastern North Atlantic) with the description of four new species. Zootaxa 2017, 4300, 505–535. [Google Scholar]
- Kucheruk, N.V. New species of the family Onuphidae (Polychaeta) from the Pacific. Zool. Zhurnal 1979, 58, 1227–1230. (In Russian) [Google Scholar]
- Kucheruk, N.V. A new genus and species of polychaetes (Onuphidae). Zool. Zhurnal 1979, 58, 1585–1587. (In Russian) [Google Scholar]
- Paterson, G.L.J.; Glover, A.G.; Frojan, C.B.; Whitaker, A.; Budaeva, N.; Chimonides, J.; Doner, S. A census of abyssal polychaetes. Deep-Sea Res. II 2009, 56, 1739–1746. [Google Scholar] [CrossRef]
- Bergamo, G.; Carrerette, O.; Nogueira, J.M.M. Continuous and non-seasonal reproductive cycle of the alien species Diopatra neapolitana (Onuphidae, Annelida) in a tropical bay of SW Atlantic. Estuar. Coast. Shelf Sci. 2019, 231, 106479. [Google Scholar] [CrossRef]
- Elgetany, A.H.; van Rensburg, H.; Hektoen, M.; Matthee, C.; Budaeva, N.; Simon, C.A.; Struck, T.H. Species delineation in the speciation grey zone—The case of Diopatra (Annelida, Onuphidae). Zool. Scr. 2020, 49, 516–534. [Google Scholar]
- Budaeva, N.; Borisova, P.; Willassen, E. Nothria conchylega (Onuphidae, Annelida)—An example of a widespread species of polychaetes in the Northern Hemisphere. unpublished.
- Rodrigues, A.M.; Pires, A.; Mendo, S.; Quintino, V. Diopatra neapolitana and Diopatra marocensis from the Portuguese coast: Morphological and genetic comparison. Estuar. Coast. Shelf Sci. 2009, 85, 609–617. [Google Scholar] [CrossRef]
- Pires, A.; Paxton, H.; Quintino, V.; Rodrigues, A.M. Diopatra (Annelida: Onuphidae) diversity in European waters with the description of Diopatra micrura, new species. Zootaxa 2010, 2395, 17–33. [Google Scholar] [CrossRef]
- Nygren, A. Cryptic polychaete diversity: A review. Zool. Scr. 2014, 43, 172–183. [Google Scholar]
- Hektoen, M.M. (Norwegian University of Science and Technology, Trondheim, Norway). Personal communication. 2020.
- Budaeva, N.; Paxton, H.; Ribeiro, P.; Sellanes, J.; Willassen, E. Systematics and diversity of quill worms (Hyalinoeciinae, Onuphidae, Annelida). unpublished.
- Paxton, H.; Budaeva, N. Nothria (Onuphidae, Annelida) from deep Australian waters. unpublished.
- Bors, E.K.; Rowden, A.A.; Maas, E.W.; Clark, M.R.; Shank, T.M. Patterns of Deep-Sea Genetic Connectivity in the New Zealand Region: Implications for Management of Benthic Ecosystems. PLoS ONE 2012, 7, e49474. [Google Scholar] [CrossRef] [PubMed]
- Padovan, A.; Chick, R.C.; Cole, V.J.; Dutoit, L.; Hutchings, P.A.; Jack, C.; Fraser, C.I. Genomic analyses suggest strong population connectivity over large spatial scales of the commercially important baitworm, Australonuphis teres (Onuphidae). Mar. Freshw. Res. 2020, 71, 1549–1556. [Google Scholar] [CrossRef]
- Dağli, E.; Ergen, Z.; Çinar, M.E. One-year observation on the population structure of Diopatra neapolitana Delle Chiaje (Polychaeta, Onuphidae) in Izmir Bay (Aegean Sea, eastern Mediterranean). Mar. Ecol. 2005, 26, 265–272. [Google Scholar] [CrossRef]










Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).