
Defining Conservation Requirements for the Suweon Treefrog (Dryophytes suweonensis) Using Species Distribution Models



Abstract

1. Introduction
2. Materials and Methods
2.1. Species Introduction
2.2. Study Area
2.3. Modeling Method
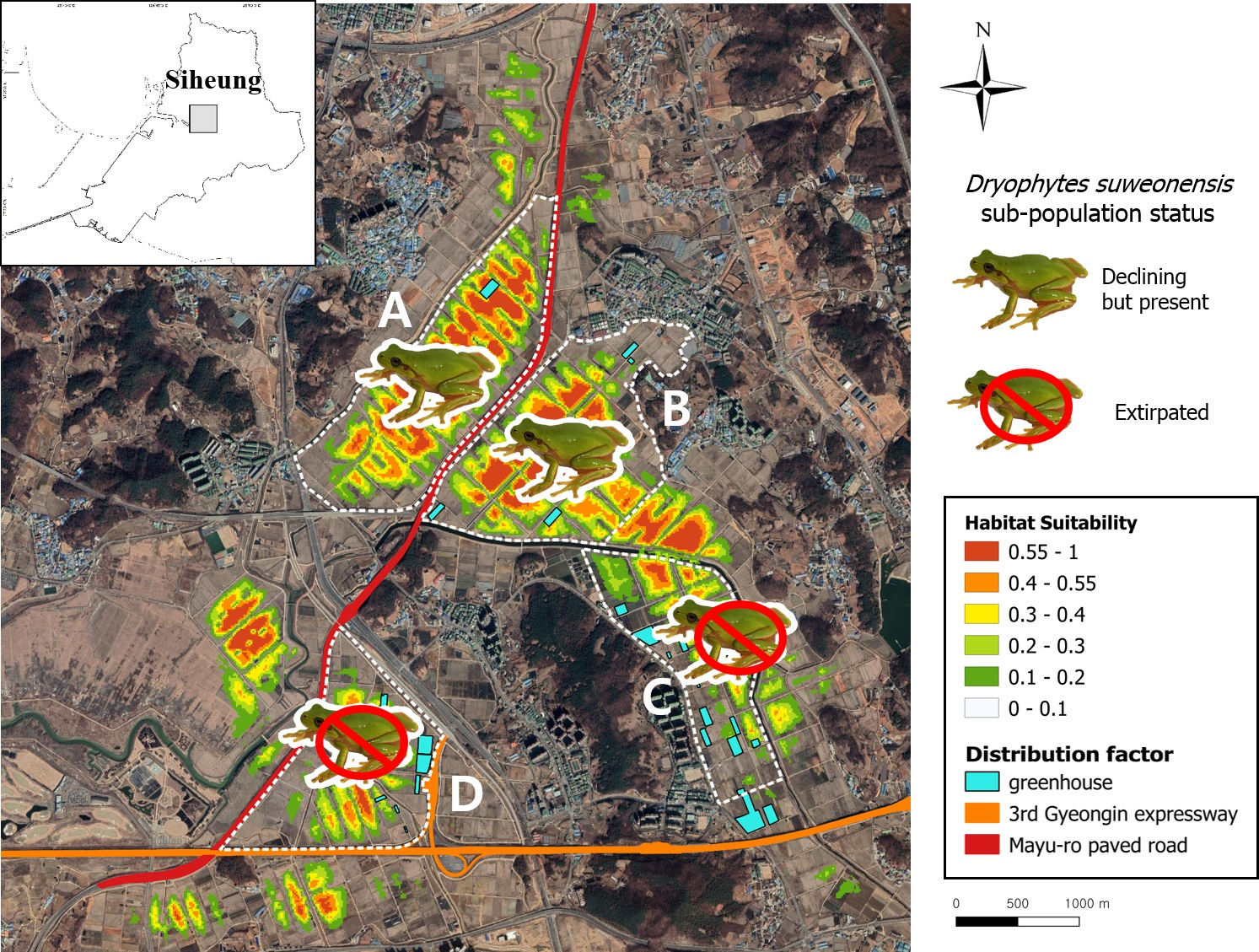
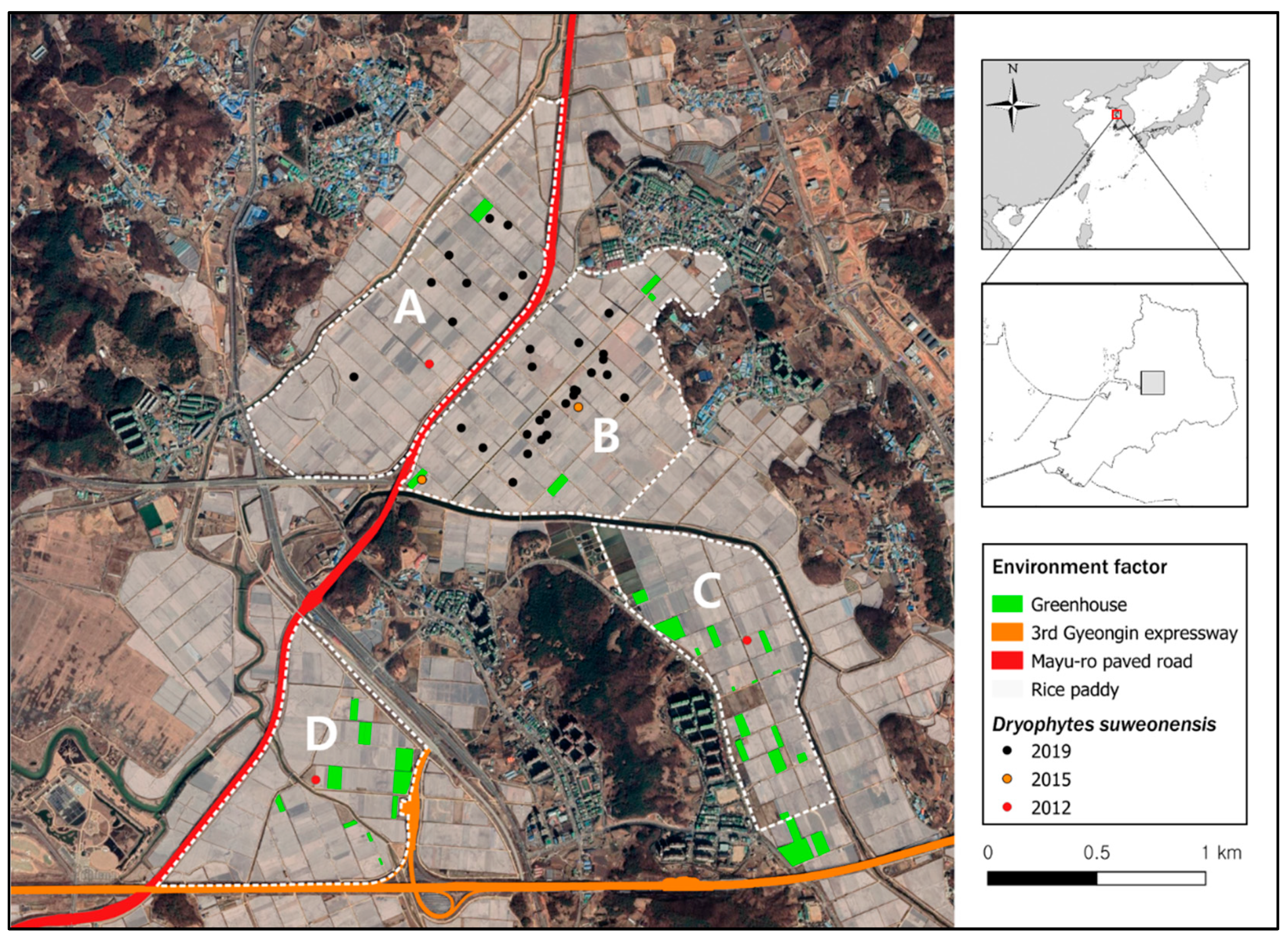
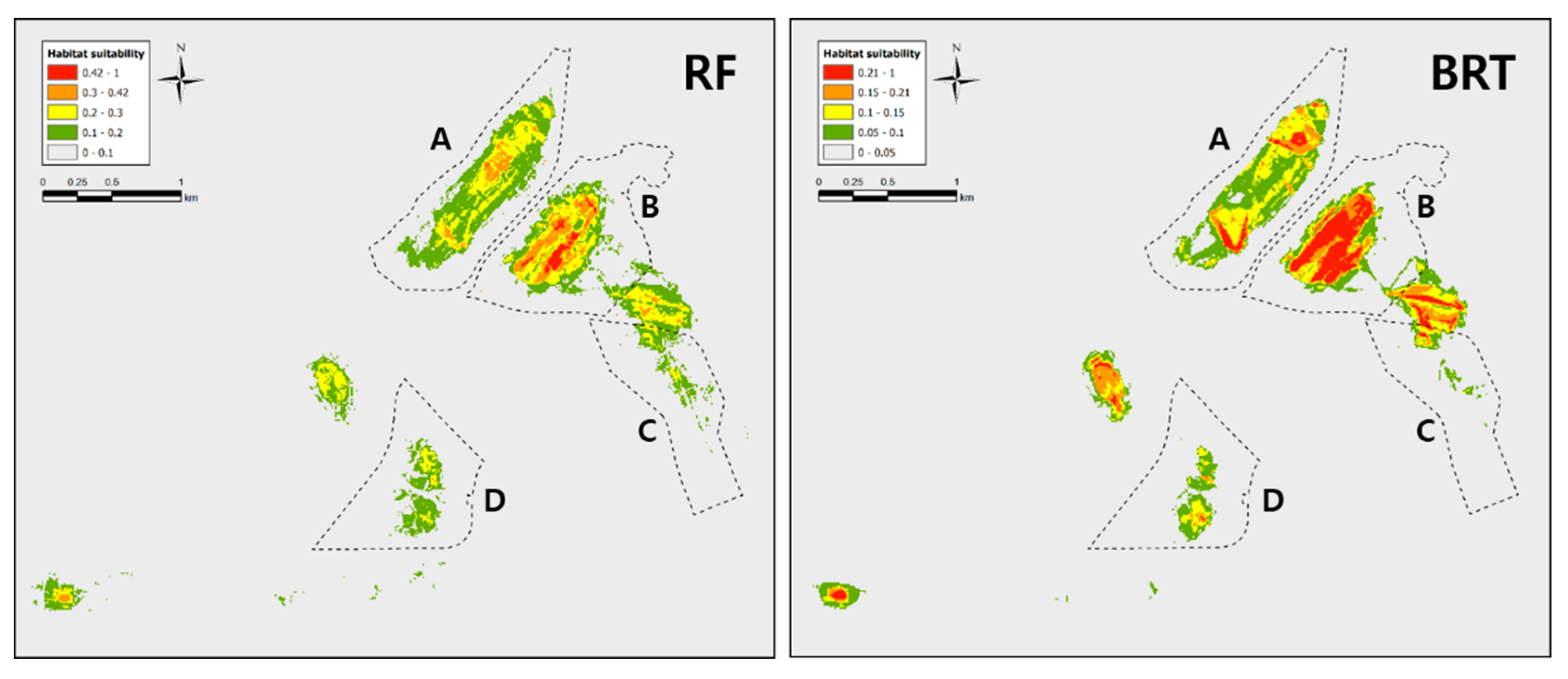
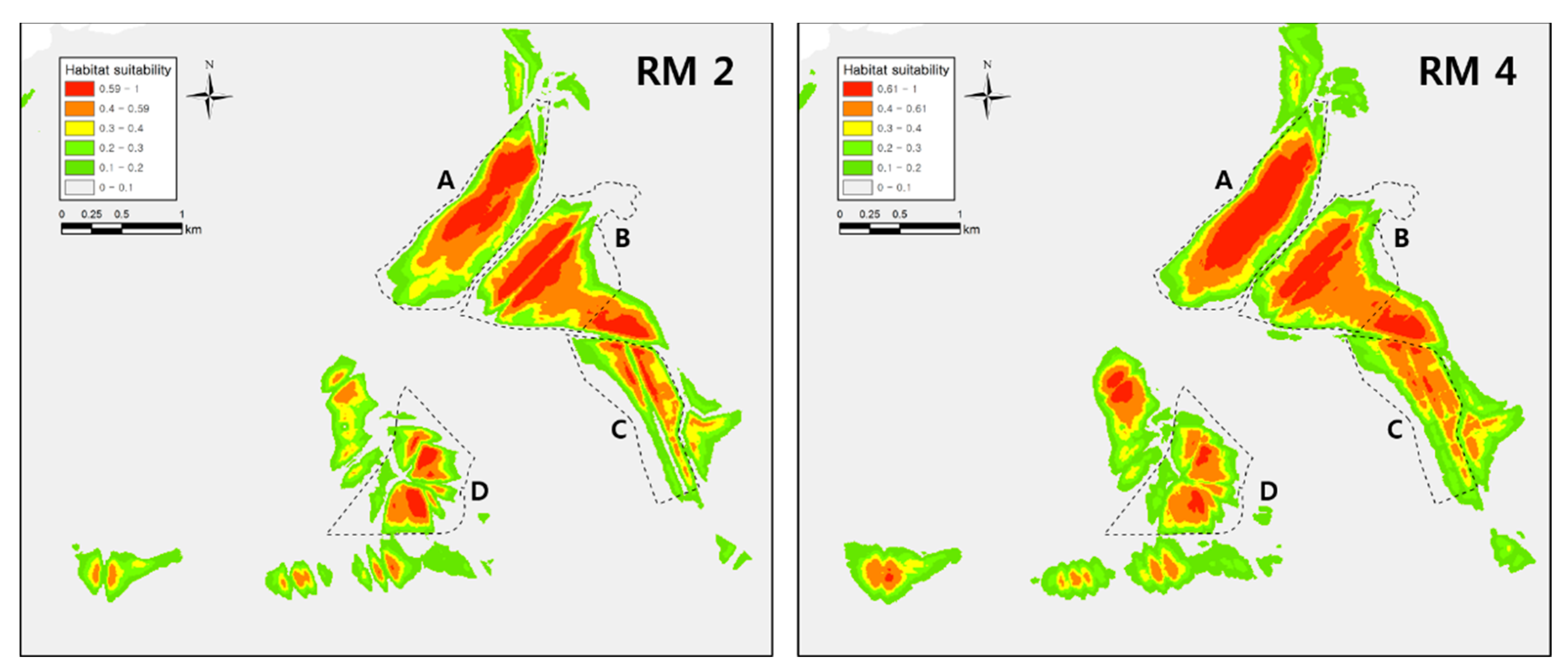
3. Results
3.1. Sampling and Habitat Analysis
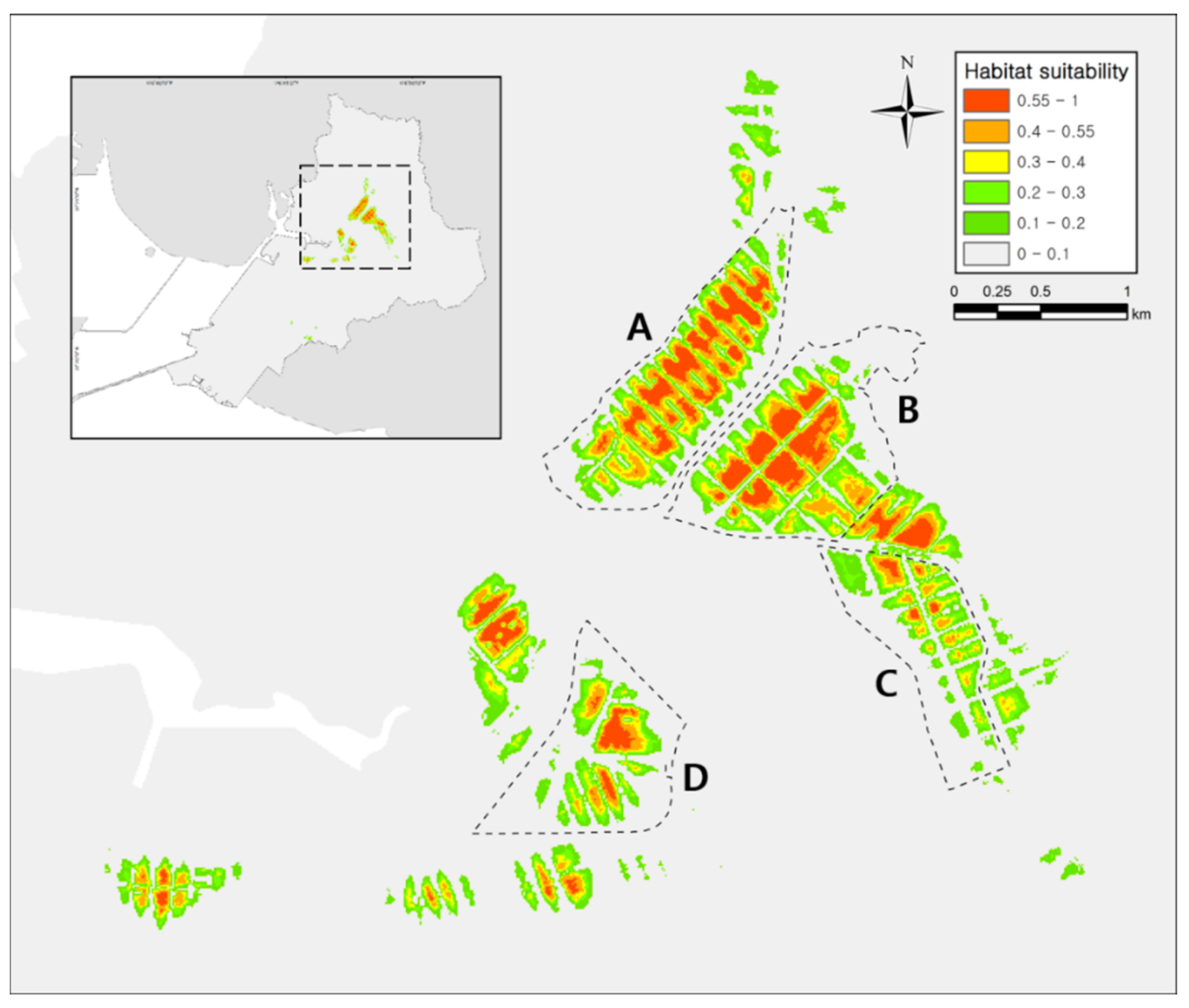
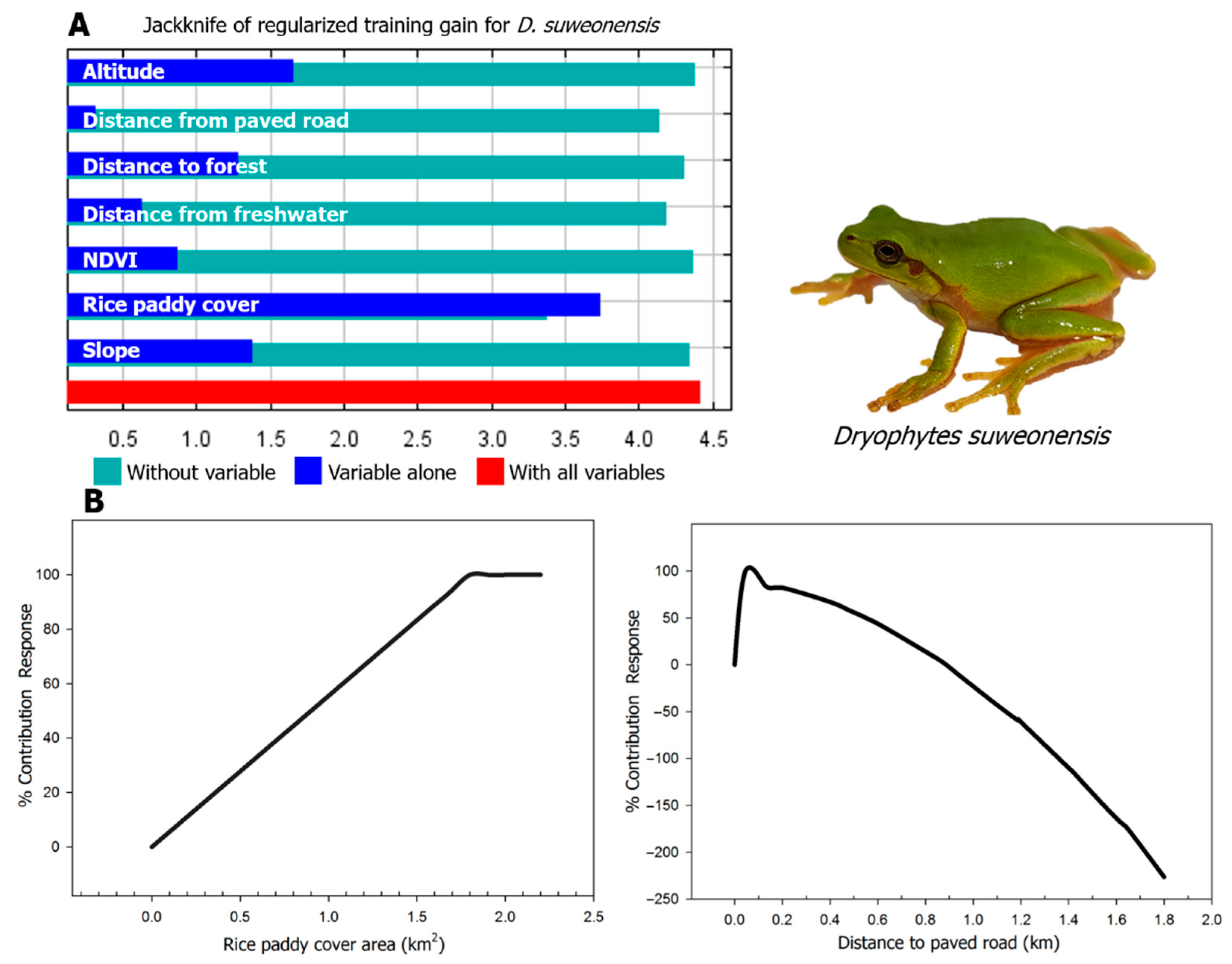
3.2. Modeling Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A


References
- Fisher, R.N.; Shaffer, H.B. The Decline of amphibians in California’s Great Central Valley. Conserv. Biol. 1996, 10, 1387–1397. [Google Scholar] [CrossRef]
- Alford, R.A.; Richards, S.J. Global amphibian declines: A problem in applied ecology. Annu. Rev. Ecol. Syst. 1999, 30, 133–165. [Google Scholar] [CrossRef]
- Gardner, T. Declining amphibian populations: A global phenomenon in conservation of biology. Anim. Biodivers. Conserv. 2001, 24, 25–44. [Google Scholar]
- Collins, J.P. Amphibian decline and extinction: What we know and what we need to learn. Dis. Aquat. Org. 2010, 92, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Tilman, D.; May, R.M.; Lehman, C.L.; Nowak, M.A. Habitat destruction and the extinction debt. Nature 1994, 371, 65–66. [Google Scholar] [CrossRef]
- Marsh, D.M.; Trenham, P.C. Metapopulation dynamics and amphibian conservation. Conserv. Biol. 2001, 15, 40–49. [Google Scholar] [CrossRef]
- Fahrig, L. Effects of habitat fragmentation on biodiversity. Annu. Rev. Ecol. Evol. Syst. 2003, 34, 487–515. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Hobbs, R.J.; Montague-Drake, R.; Alexandra, J.; Bennett, A.; Burgman, M.; Cale, P.; Calhoun, A.; Cramer, V.; Cullen, P.; et al. A checklist for ecological management of landscapes for conservation. Ecol. Lett. 2007, 11, 78–91. [Google Scholar] [CrossRef]
- Olds, A.D.; Connolly, R.M.; Pitt, K.A.; Maxwell, P.S. Habitat connectivity improves reserve performance. Conserv. Lett. 2011, 5, 56–63. [Google Scholar] [CrossRef]
- Hanski, I. Metapopulation dynamics: Does it help to have more of the same? Trends Ecol. Evol. 1989, 4, 113–114. [Google Scholar] [CrossRef]
- Cushman, S.A. Effects of habitat loss and fragmentation on amphibians: A review and prospectus. Biol. Conserv. 2006, 128, 231–240. [Google Scholar] [CrossRef]
- Hamer, A.J.; McDonnell, M.J. Amphibian ecology and conservation in the urbanising world: A review. Biol. Conserv. 2008, 141, 2432–2449. [Google Scholar] [CrossRef]
- Radeloff, V.C.; Hammer, R.B.; Stewart, S.I. Rural and suburban sprawl in the U.S. Midwest from 1940 to 2000 and its relation to forest fragmentation. Conserv. Biol. 2005, 19, 793–805. [Google Scholar] [CrossRef]
- Burkey, T.V. Extinction in Nature Reserves: The effect of fragmentation and the importance of migration between reserve fragments. Oikos 1989, 55, 75–81. [Google Scholar] [CrossRef]
- Feldman, C.R.; Spicer, G.S. Comparative phylogeography of woodland reptiles in California: Repeated patterns of cladogenesis and population expansion. Mol. Ecol. 2006, 15, 2201–2222. [Google Scholar] [CrossRef] [PubMed]
- Castoe, T.A.; Spencer, C.L.; Parkinson, C.L. Phylogeographic structure and historical demography of the western diamondback rattlesnake (Crotalus atrox): A perspective on North American desert biogeography. Mol. Phylogenet. Evol. 2007, 42, 193–212. [Google Scholar] [CrossRef]
- Lemmon, E.M.; Lemmon, A.R.; Collins, J.T.; Lee-Yaw, J.A.; Cannatella, D.C. Phylogeny-based delimitation of species boundaries and contact zones in the trilling chorus frogs (Pseudacris). Mol. Phylogenet. Evol. 2007, 44, 1068–1082. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.T.T. Connecting wildlife populations in fractured landscapes. In Safe Passages: Highways, Wildlife, and Habitat Connectivity; Beckmann, J.P., Clevenger, A.P., Huijser, M.P., Hillty, J.A., Eds.; Island Press: Washington, DC, USA, 2012; pp. 3–16. [Google Scholar]
- Groffen, J.; Borzée, A.; Jang, Y. Preference for natural borders in rice paddies by two treefrog species. Anim. Cells Syst. 2018, 22, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Marvier, M.; Kareiva, P.; Neubert, M.G. Habitat destruction, fragmentation, and disturbance promote invasion by habitat generalists in a multispecies metapopulation. Risk Anal. 2004, 24, 869–878. [Google Scholar] [CrossRef] [PubMed]
- Kurechi, M. Restoring rice paddy wetland environments and the local sustainable society-project for achieving co-existence of rice paddy agriculture with waterbirds at Kabukuri-Numa, Miyagi Prefecture, Japan. Glob. Environ. Res. 2007, 11, 141–152. (In English) [Google Scholar]
- Green, R.E.; Cornell, S.J.; Scharlemann, J.P.W.; Balmford, A. Farming and the fate of wild nature. Science 2005, 307, 550–555. [Google Scholar] [CrossRef]
- Ramsar, COP10. Enhancing biodiversity in rice paddies as wetland systems. In Proceedings of the 10th Meeting of the Conference of the Parties to the Convention on Wetlands (Ramsar, Iran, 1971), Changwon, Korea, 28 October–4 November 2008. [Google Scholar]
- Fuller, R.J.; Norton, L.; Feber, R.; Johnson, P.; Chamberlain, D.; Joys, A.; Mathews, F.; Stuart, R.; Townsend, M.; Manley, W.; et al. Benefits of organic farming to biodiversity vary among taxa. Biol. Lett. 2005, 1, 431–434. [Google Scholar] [CrossRef]
- Butler, S.J.; Brooks, D.; Feber, R.E.; Storkey, J.; Vickery, J.A.; Norris, K. A Cross-Taxonomic index for quantifying the health of farmland biodiversity. J. Appl. Ecol. 2009, 46, 1154–1162. [Google Scholar] [CrossRef]
- Jang, H.J.; Suh, J.H. Distribution of amphibian species in South Korea. Korean J. Herpetol. 2010, 2, 45–51. [Google Scholar]
- Hazell, D.; Hero, J.-M.; Lindenmayer, D.; Cunningham, R. A comparison of constructed and natural habitat for frog conservation in an Australian agricultural landscape. Biol. Conserv. 2004, 119, 61–71. [Google Scholar] [CrossRef]
- Borzée, A.; Jang, Y. Description of a seminatural habitat of the endangered Suweon treefrog Hyla suweonensis. Anim. Cells Syst. 2015, 19, 216–220. [Google Scholar] [CrossRef]
- Borzée, A.; Andersen, D.; Jang, Y. Population trend inferred from aural surveys for calling anurans in Korea. PeerJ 2018, 6, e5568. [Google Scholar] [CrossRef]
- Borzée, A.; Heo, K.; Jang, Y. Relationship between agro-environmental variables and breeding Hylids in rice paddies. Sci. Rep. 2018, 8, 8049. [Google Scholar] [CrossRef]
- Borzée, A.; Kim, K.; Heo, K.; Jablonski, P.G.; Jang, Y. Impact of land reclamation and agricultural water regime on the distribution and conservation status of the endangered Dryophytes suweonensis. PeerJ 2017, 5, e3872. [Google Scholar] [CrossRef]
- Kim, I.H.; Son, S.H.; Kang, S.W.; Kim, J.B. Distribution and habitat characteristics of the endangered Suweon-Tree Frog (Hyla suweonensis). Korean J. Herpetol. 2012, 4, 15–22. [Google Scholar]
- Smith, M.A.; Green, D.M. Dispersal and the metapopulation paradigm in amphibian ecology and conservation: Are all amphibian populations metapopulations? Ecography 2005, 28, 110–128. [Google Scholar] [CrossRef]
- Borzée, A. Why Are Anurans Threatened? The case of Dryophytes suweonensis. Ph.D. Thesis, Seoul National University, Seoul, Korea, February 2018. [Google Scholar]
- Borzée, A.; Kosch, T.A.; Kim, M.; Jang, Y. Introduced bullfrogs are associated with increased Batrachochytrium dendrobatidis prevalence and reduced occurrence of Korean treefrogs. PLoS ONE 2017, 12, e0177860. [Google Scholar]
- Rodríguez, J.P.; Brotons, L.; Bustamante, J.; Seoane, J. The application of predictive modelling of species distribution to biodiversity conservation. Divers. Distrib. 2007, 13, 243–251. [Google Scholar] [CrossRef]
- Franklin, J. Species distribution models in conservation biogeography: Developments and challenges. Divers. Distrib. 2013, 19, 1217–1223. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef]
- Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Elith, J.; Gordon, A.; Kujala, H.; Lentini, P.E.; McCarthy, M.A.; Tingley, R.; Wintle, B.A. Is my species distribution model fit for purpose? Matching data and models to applications. Glob. Ecol. Biogeogr. 2015, 24, 276–292. [Google Scholar] [CrossRef]
- Heikkinen, R.K.; Marmion, M.; Luoto, M. Does the interpolation accuracy of species distribution models come at the expense of transferability? Ecography 2012, 35, 276–288. [Google Scholar] [CrossRef]
- Phillips, S.J. A brief tutorial on Maxent. ATT Res. 2005, 190, 231–259. [Google Scholar]
- Young, N.; Carter, L.; Evangelista, P. A MaxEnt Model v3. 3.3 e Tutorial (ArcGIS v10); Colorado State University: Fort Collins, CO, USA, 2011. [Google Scholar]
- Radosavljevic, A.; Anderson, R.P. Making better Maxent models of species distributions: Complexity, overfitting and evaluation. J. Biogeogr. 2014, 41, 629–643. [Google Scholar] [CrossRef]
- Borzée, A.; Kim, J.Y.; Jang, Y. Asymmetric competition over calling sites in two closely related treefrog species. Sci. Rep. 2016, 6, 32569. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Hanley, J.A.; McNeil, B.J. The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 1982, 143, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Bradley, A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recognit. 1997, 30, 1145–1159. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Beck, P.S.A.; Atzberger, C.; Høgda, K.A.; Johansen, B.; Skidmore, A.K. Improved monitoring of vegetation dynamics at very high latitudes: A new method using MODIS NDVI. Remote Sens. Environ. 2006, 100, 321–334. [Google Scholar] [CrossRef]
- Lee, B.; Kim, E.; Lee, J.; Chung, J.M.; Lim, J.H. Detecting Phenology Using MODIS vegetation indices and Forest Type Msp in South Korea. Korean J. Remote Sens. 2018, 34, 267–282. [Google Scholar]
- Ziegel, E.R.; Neter, J.; Kutner, M.; Nachtsheim, C.; Wasserman, W. Applied Linear Statistical Models. Technometrics 1997, 39, 342. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Roh, G.; Borzée, A.; Jang, Y. Spatiotemporal distributions and habitat characteristics of the endangered treefrog, Hyla suweonensis, in relation to sympatric H. japonica. Ecol. Inform. 2014, 24, 78–84. [Google Scholar] [CrossRef]
- Carr, L.W.; Fahrig, L. Effect of road traffic on two amphibian species of differing vagility. Conserv. Biol. 2001, 15, 1071–1078. [Google Scholar] [CrossRef]
- Karraker, N.E.; Gibbs, J.P. Contrasting road effect signals in reproduction of long-versus short-lived amphibians. Hydrobiologia 2011, 664, 213–218. [Google Scholar] [CrossRef]
- Ministry of Environment. Documentation Guideline for Ecological Nature Map; Technical report No. 645; Ministry of Environment Regulations: Sejong, Korea, 2018. [Google Scholar]
- Borzée, A.; Choi, Y.; Kim, Y.E.; Jablonski, P.G.; Jang, Y. Interspecific variation in seasonal migration and brumation behavior in two closely related species of treefrogs. Front. Ecol. Evol. 2019, 7, 55. [Google Scholar] [CrossRef]
- Shin, Y.; Jeong, D.; Borzée, A. Mass displacement of Korean clawed salamanders (Onychodactylus koreanus) and the threat of Road-Kill. Herpetol. Bull. 2020, 151, 28–31. [Google Scholar] [CrossRef][Green Version]
- Lee, J.H.; Lee, C.W.; Yang, H.S.; Kim, T.S.; Lee, J.H.; Park, S.J.; Yang, B.G. Post-Breeding Dispersal and Movement patterns of Male Asian Toads (Bufo gargarizans). Korean J. Herpetol. 2013, 5, 1–8. [Google Scholar]
- Do, M.S.; Jang, H.J.; Kim, D.I.; Koo, K.S.; Lee, S.C.; Nam, H.K. The study on habitat analysis and ecological niche of Korean Brown Frogs (Rana dybowskii, R. coreana and R. huanrensis) using the species distribution model. Korean J. Herpetol. 2018, 9, 1–11. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Environmental Variables | Contribution (%) | Mean Value of Observed Point | Habitat Range of Observed Point | Habitable Range of Modeling Result |
|---|---|---|---|---|
| Rice paddy cover (km2) | 81.9 | 0.15 ± 0.00 | 0.10–0.22 | 0.12–0.22 |
| Distance to paved road (m) | 5.1 | 89.1 ± 13.8 | 10.0–316.2 | 0–502.9 |
| NDVI (0–100) | 4.5 | 0.62 ± 0.02 | 0.31–0.73 | 0–0.78 |
| Distance to freshwater (m) | 4.1 | 115.9 ± 12.2 | 20.0–308.9 | 10.0–324.5 |
| Distance to forest (m) | 2.5 | 443.1 ± 22.7 | 123.7–710.2 | 187.9–747.3 |
| Slope (°) | 1.4 | 0.36 ± 0.03 | 0.07–0.65 | 0.07–1.19 |
| Altitude (m) | 0.5 | 3.9 ± 0.1 | 3.2–4.9 | 2.9–4.9 |
| Site | Total Area (km2) | Area of Rice Paddy Cover Over 0.12 km2 (km2) | Mean Total Area of Rice Paddy Cover (km2) | Mean Distance from Paved Road (m) |
|---|---|---|---|---|
| A | 1.05 | 0.66 (62.9%) | 0.14 ± 0.04 | 138 ± 93 |
| B | 1.05 | 0.66 (62.8%) | 0.13 ± 0.03 | 49 ± 28 |
| C | 0.67 | 0.32 (47.9%) | 0.11 ± 0.02 | 37 ± 24 |
| D | 0.78 | 0.25 (31.3%) | 0.10 ± 0.02 | 39 ± 28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, I.-K.; Park, D.; Borzée, A. Defining Conservation Requirements for the Suweon Treefrog (Dryophytes suweonensis) Using Species Distribution Models. Diversity 2021, 13, 69. https://doi.org/10.3390/d13020069
Park I-K, Park D, Borzée A. Defining Conservation Requirements for the Suweon Treefrog (Dryophytes suweonensis) Using Species Distribution Models. Diversity. 2021; 13(2):69. https://doi.org/10.3390/d13020069
Chicago/Turabian StylePark, Il-Kook, Daesik Park, and Amaël Borzée. 2021. "Defining Conservation Requirements for the Suweon Treefrog (Dryophytes suweonensis) Using Species Distribution Models" Diversity 13, no. 2: 69. https://doi.org/10.3390/d13020069
APA StylePark, I.-K., Park, D., & Borzée, A. (2021). Defining Conservation Requirements for the Suweon Treefrog (Dryophytes suweonensis) Using Species Distribution Models. Diversity, 13(2), 69. https://doi.org/10.3390/d13020069